

Carnivorous Plants from Nepenthaceae and Droseraceae as a Source of Secondary Metabolites



Abstract
1. Introduction
2. Secondary Metabolites in Nepenthaceae and Droseraceae
2.1. Phenolic Compounds
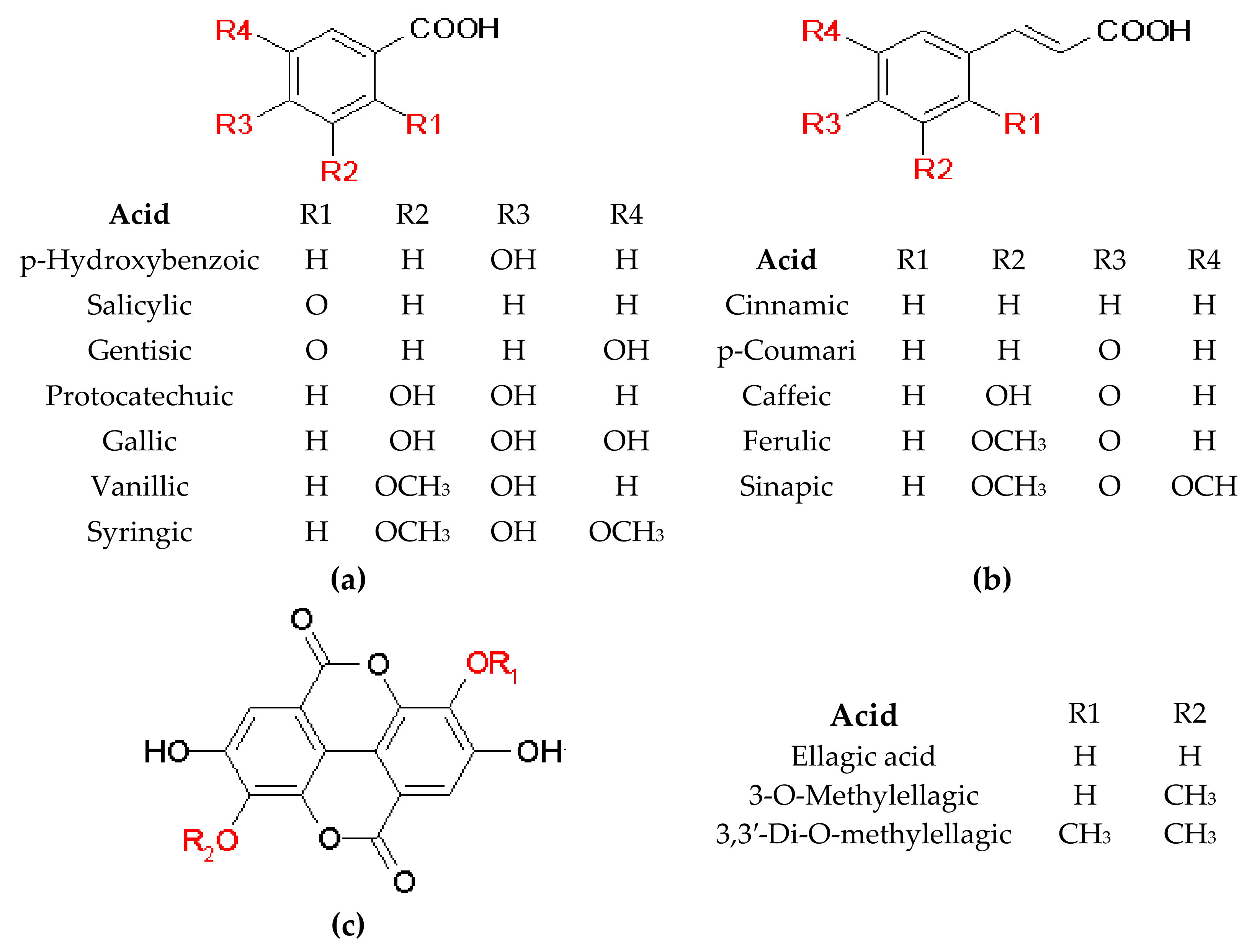
2.1.1. Phenolic Acids
General Information
Phenolic Acids and Their Derivatives in Nepenthaceae and Droseraceae
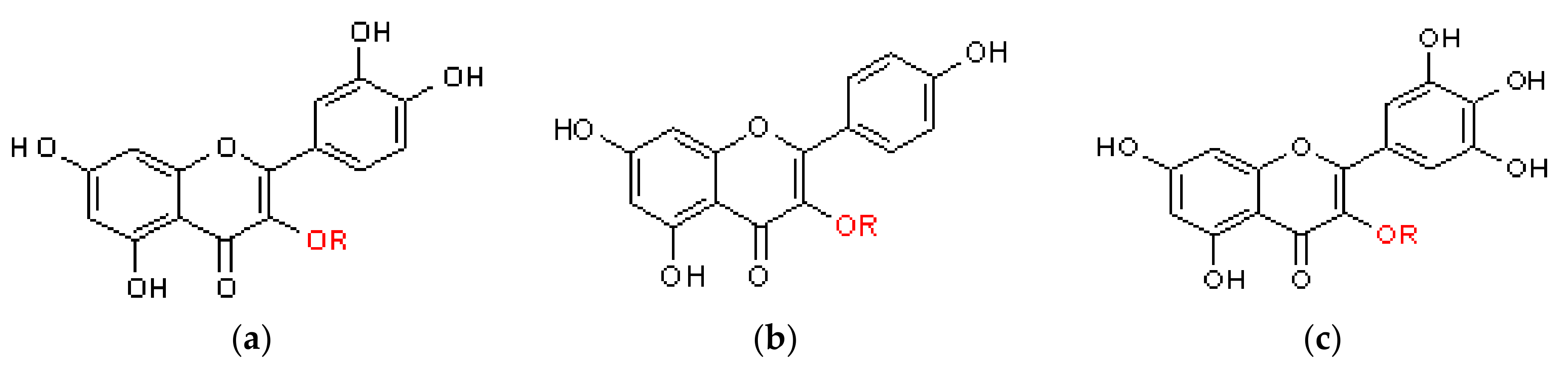
2.1.2. Flavonoids
General Information
Flavonoids in Nepenthaceae and Droseraceae
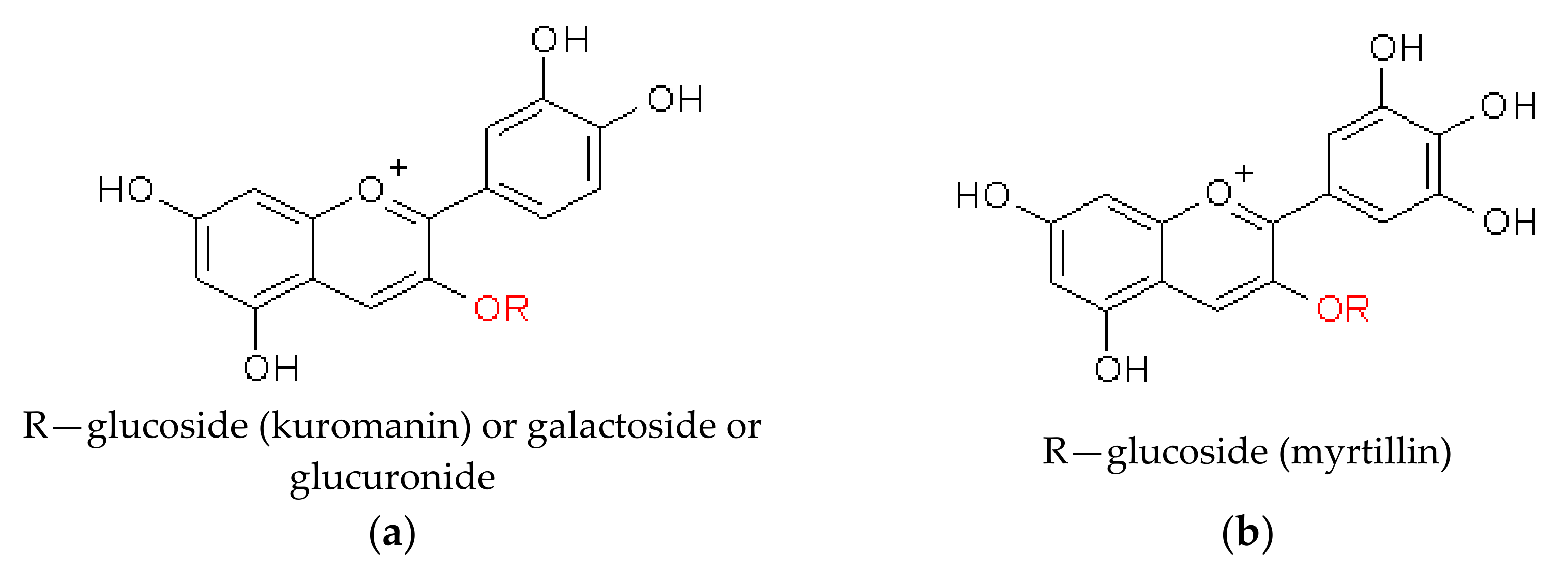
Anthocyanins
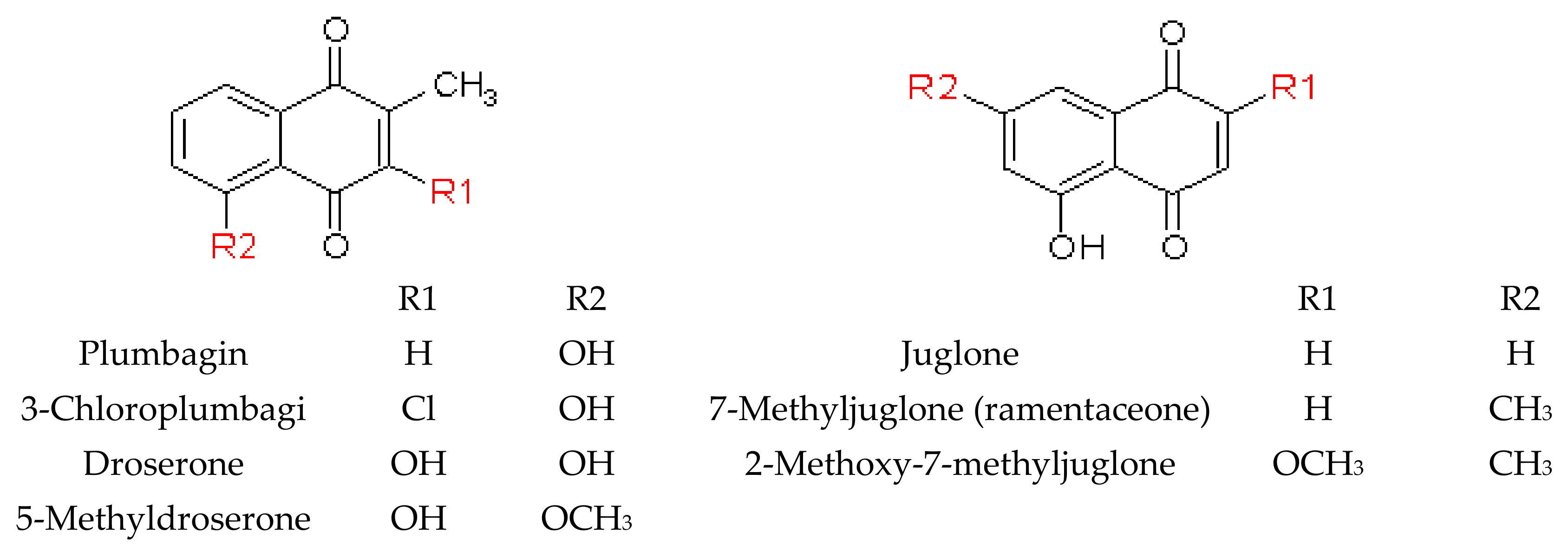
2.2. Naphthoquinones
2.2.1. General Information
2.2.2. Naphthoquinones in Nepenthaceae and Droseraceae
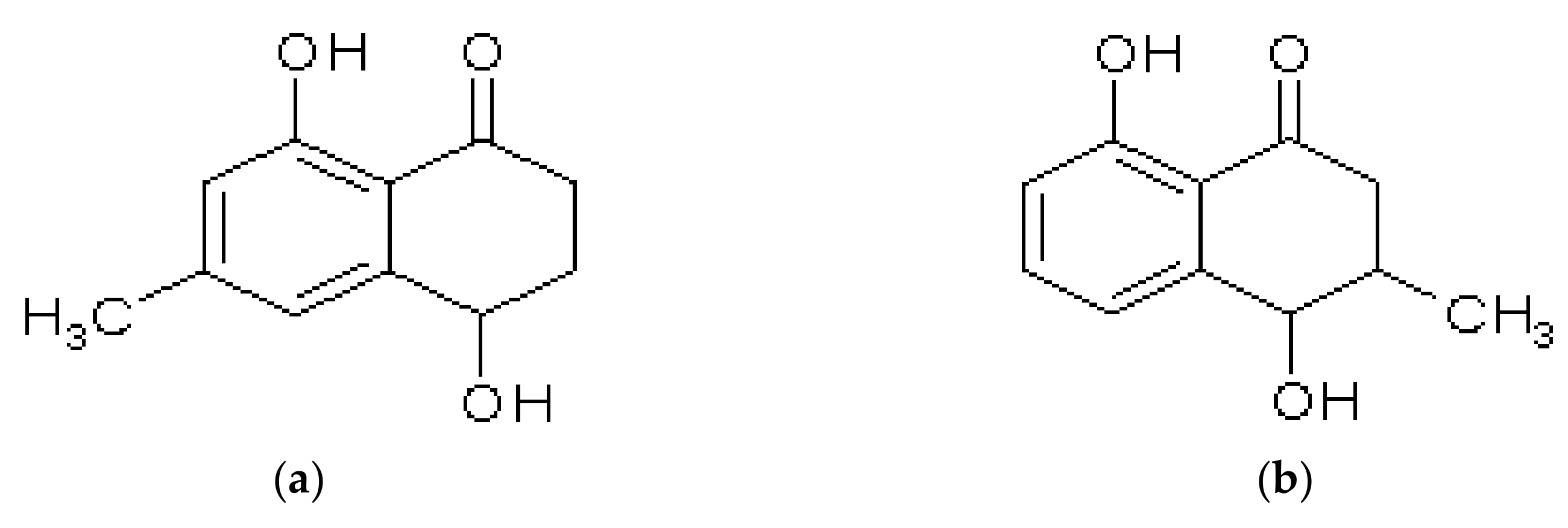
2.2.3. Acetogenic Tetralones
2.3. Other Metabolites
3. Biological Potential of Nepentheceae and Droseraceae Species
4. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Król, E.; Płachno, B.J.; Adamec, L.; Stolarz, M.; Dziubińska, H.; Trębacz, K. Quite a Few Reasons for Calling Carnivores ‘the Most Wonderful Plants in the World’. Ann. Bot. 2012, 109, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.T.; Krueger, T.A.; Gonella, P.M.; Robinson, A.S.; Fleischmann, A.S. Conservation of Carnivorous Plants in the Age of Extinction. Glob. Ecol. Conserv. 2020, 24, e01272. [Google Scholar] [CrossRef]
- Barthlott, W.; Fischer, E.; Frahm, J.-P.; Seine, R. First Experimental Evidence for Zoophagy in the Hepatic Colura. Plant Biol. 2000, 2, 93–97. [Google Scholar] [CrossRef]
- Hess, S.; Frahm, J.P.; Theisen, I. Evidence of Zoophagy in a Second Liverwort Species, Pleurozia purpurea. Bryologist 2005, 108, 212–218. [Google Scholar] [CrossRef]
- Albert, V.A.; Williams, S.E.; Chase, M.W. Carnivorous Plants: Phylogeny and Structural Evolution. Science 1992, 257, 1491–1495. [Google Scholar] [CrossRef]
- Cameron, K.M.; Wurdack, K.J.; Jobson, R.W. Molecular Evidence for the Common Origin of Snap-Traps among Carnivorous Plants. Am. J. Bot. 2002, 89, 1503–1509. [Google Scholar] [CrossRef]
- Fleischmann, A.; Schlauer, J.; Smith, S.A.; Givnish, T.J. Evolution of Carnivory in Angiosperms; Oxford University Press: Oxford, UK, 2018; Volume 1. [Google Scholar] [CrossRef]
- Silva, S.R.; Gibson, R.; Adamec, L.; Domínguez, Y.; Miranda, V.F.O. Molecular Phylogeny of Bladderworts: A Wide Approach of Utricularia (Lentibulariaceae) Species Relationships Based on Six Plastidial and Nuclear DNA Sequences. Mol. Phylogenet. Evol. 2018, 118, 244–264. [Google Scholar] [CrossRef]
- Lin, Q.; Ané, C.; Givnish, T.J.; Graham, S.W. A New Carnivorous Plant Lineage (Triantha) with a Unique Sticky-Inflorescence Trap. Proc. Natl. Acad. Sci. USA 2021, 118, e2022724118. [Google Scholar] [CrossRef]
- Ellison, A.M.; Adamec, L. Introduction: What Is a Carnivorous Plant? Oxford University Press: Oxford, UK, 2018; Volume 1. [Google Scholar] [CrossRef]
- Płachno, B.J.; Muravnik, L.E. Functional Anatomy of Carnivorous Traps; Oxford University Press: Oxford, UK, 2018; Volume 1. [Google Scholar]
- Givnish, T.J.; Sparks, K.W.; Hunter, S.J.; Pavlovič, A. Why Are Plants Carnivorous? Cost/Benefit Analysis, Whole-Plant Growth, and the Context-Specific Advantages of Botanical Carnivory; Oxford University Press: Oxford, UK, 2018; Volume 1. [Google Scholar] [CrossRef]
- Freund, M.; Graus, D.; Fleischmann, A.; Gilbert, K.J.; Lin, Q.; Renner, T.; Stigloher, C.; Albert, V.A.; Hedrich, R.; Fukushima, K. The Digestive Systems of Carnivorous Plants. Plant Physiol. 2022, 190, 44–59. [Google Scholar] [CrossRef]
- Miclea, I. Secondary Metabolites with Biomedical Applications from Plants of the Sarraceniaceae Family. Int. J. Mol. Sci. 2022, 23, 9877. [Google Scholar] [CrossRef]
- Hatcher, C.R.; Ryves, D.B.; Millett, J. The Function of Secondary Metabolites in Plant Carnivory. Ann. Bot. 2020, 125, 399–411. [Google Scholar] [CrossRef]
- Mithöfer, A. Carnivorous Pitcher Plants: Insights in an Old Topic. Phytochemistry 2011, 72, 1678–1682. [Google Scholar] [CrossRef]
- Sanusi, S.B.; Abu Bakar, M.F.; Mohamed, M.; Sabran, S.F.; Mainasara, M.M. Ethnobotanical, Phytochemical, and Pharmacological Properties of Nepenthes Species: A Review. Asian J. Pharm. Clin. Res. 2017, 10, 16–19. [Google Scholar] [CrossRef]
- Brewer, J.S.; Schlauer, J. Biogeography and Habitats of Carnivorous Plants; Oxford University Press: Oxford, UK, 2018; Volume 1. [Google Scholar]
- Adamec, L. Biological Flora of Central Europe: Aldrovanda vesiculosa L. Perspect. Plant Ecol. Evol. Syst. 2018, 35, 8–21. [Google Scholar] [CrossRef]
- Tienaho, J.; Reshamwala, D.; Karonen, M.; Silvan, N.; Korpela, L.; Marjomäki, V.; Sarjala, T. Field-Grown and In Vitro Propagated Round-Leaved Sundew (Drosera rotundifolia L.) Show Differences in Metabolic Profiles and Biological Activities. Molecules 2021, 26, 3581. [Google Scholar] [CrossRef]
- Makowski, W.; Królicka, A.; Nowicka, A.; Zwyrtková, J.; Tokarz, B.; Pecinka, A.; Banasiuk, R.; Tokarz, K.M. Transformed Tissue of Dionaea muscipula J. Ellis as a Source of Biologically Active Phenolic Compounds with Bactericidal Properties. Appl. Microbiol. Biotechnol. 2021, 105, 1215–1226. [Google Scholar] [CrossRef]
- Makowski, W.; Królicka, A.; Tokarz, B.; Miernicka, K.; Kołton, A.; Pięta, Ł.; Malek, K.; Ekiert, H.; Szopa, A.; Tokarz, K.M. Response of Physiological Parameters in Dionaea muscipula J. Ellis Teratomas Transformed with RolB Oncogene. BMC Plant Biol. 2021, 21, 564. [Google Scholar] [CrossRef]
- Dávila-Lara, A.; Rodríguez-López, C.E.; O’Connor, S.E.; Mithöfer, A. Metabolomics Analysis Reveals Tissue-Specific Metabolite Compositions in Leaf Blade and Traps of Carnivorous Nepenthes Plants. Int. J. Mol. Sci. 2020, 21, 4376. [Google Scholar] [CrossRef]
- Rosli, M.A.F.; Mediani, A.; Azizan, K.A.; Baharum, S.N.; Goh, H.-H. UPLC-TOF-MS/MS-Based Metabolomics Analysis Reveals Species-Specific Metabolite Compositions in Pitchers of Nepenthes ampullaria, Nepenthes rafflesiana, and Their Hybrid Nepenthes × hookeriana. Front. Plant Sci. 2021, 12, 655004. [Google Scholar] [CrossRef]
- Hatcher, C.R.; Sommer, U.; Heaney, L.M.; Millett, J. Metabolomic Analysis Reveals Reliance on Secondary Plant Metabolites to Facilitate Carnivory in the Cape Sundew, Drosera capensis. Ann. Bot. 2021, 128, 301–314. [Google Scholar] [CrossRef]
- Devi, S.P.; Kumaria, S.; Rao, S.R.; Tandon, P. Carnivorous Plants as a Source of Potent Bioactive Compound: Naphthoquinones. Trop. Plant Biol. 2016, 9, 267–279. [Google Scholar] [CrossRef]
- Egan, P.A.; van der Kooy, F. Phytochemistry of the Carnivorous Sundew Genus Drosera (Droseraceae)—Future Perspectives and Ethnopharmacological Relevance. Chem. Biodivers. 2013, 10, 1774–1790. [Google Scholar] [CrossRef] [PubMed]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant Polyphenols: Chemical Properties, Biological Activities, and Synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant Phenolics: Recent Advances on Their Biosynthesis, Genetics, and Ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Hano, C.; Tungmunnithum, D. Plant Polyphenols, More than Just Simple Natural Antioxidants: Oxidative Stress, Aging and Age-Related Diseases. Medicines 2020, 7, 26. [Google Scholar] [CrossRef]
- Li, Z.-H.; Wang, Q.; Ruan, X.; Pan, C.-D.; Jiang, D.-A. Phenolics and Plant Allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef]
- Kováčik, J.; Klejdus, B.; Repčáková, K. Phenolic Metabolites in Carnivorous Plants: Inter-Specific Comparison and Physiological Studies. Plant Physiol. Biochem. 2012, 52, 21–27. [Google Scholar] [CrossRef]
- Norman, E.O.; Tuohey, H.; Pizzi, D.; Saidah, M.; Bell, R.; Brkljača, R.; White, J.M.; Gasser, R.B.; Taki, A.C.; Urban, S. Phytochemical Profiling and Biological Activity of the Australian Carnivorous Plant, Drosera magna. J. Nat. Prod. 2021, 84, 964–971. [Google Scholar] [CrossRef]
- Tominaga, Y.; Sugawa, H.; Hirabayashi, K.; Ikeda, T.; Hoshi, Y.; Nagai, R. Drosera Tokaiensis Extract Containing Multiple Phenolic Compounds Inhibits the Formation of Advanced Glycation End-Products. Arch. Biochem. Biophys. 2020, 693, 108586. [Google Scholar] [CrossRef]
- Krychowiak-Maśnicka, M.; Krauze-Baranowska, M.; Godlewska, S.; Kaczyński, Z.; Bielicka-Giełdoń, A.; Grzegorczyk, N.; Narajczyk, M.; Frackowiak, J.E.; Krolicka, A. Potential of Silver Nanoparticles in Overcoming the Intrinsic Resistance of Pseudomonas aeruginosa to Secondary Metabolites from Carnivorous Plants. Int. J. Mol. Sci. 2021, 22, 4849. [Google Scholar] [CrossRef]
- Krychowiak, M.; Grinholc, M.; Banasiuk, R.; Krauze-Baranowska, M.; Głód, D.; Kawiak, A.; Królicka, A. Combination of Silver Nanoparticles and Drosera binata Extract as a Possible Alternative for Antibiotic Treatment of Burn Wound Infections Caused by Resistant Staphylococcus aureus. PLoS ONE 2014, 9, e115727. [Google Scholar] [CrossRef]
- Makowski, W.; Tokarz, K.M.; Tokarz, B.; Banasiuk, R.; Witek, K.; Królicka, A. Elicitation-Based Method for Increasing the Production of Antioxidant and Bactericidal Phenolic Compounds in Dionaea muscipula J. Ellis Tissue. Molecules 2020, 25, 1794. [Google Scholar] [CrossRef]
- Banasiuk, R.; Krychowiak, M.; Swigon, D.; Tomaszewicz, W.; Michalak, A.; Chylewska, A.; Ziabka, M.; Lapinski, M.; Koscielska, B.; Narajczyk, M.; et al. Carnivorous Plants Used for Green Synthesis of Silver Nanoparticles with Broad-Spectrum Antimicrobial Activity. Arab. J. Chem. 2020, 13, 1415–1428. [Google Scholar] [CrossRef]
- Jiang, N.; Doseff, A.; Grotewold, E. Flavones: From Biosynthesis to Health Benefits. Plants 2016, 5, 27. [Google Scholar] [CrossRef]
- Bienenfeld, W.; Katzlmeier, H. Flavonoide aus Drosera rotundifolia L. Arch. Pharm. 1966, 299, 598–602. [Google Scholar] [CrossRef]
- Schölly, T.; Kapetanidis, I. Flavonol and Naphthoquinone Glycosides of Drosera rotundifolia. Planta Med. 1989, 55, 611–612. [Google Scholar] [CrossRef]
- Iwashina, T. Flavonoid Properties of Five Families Newly Incorporated into the Order Caryophyllales (Review). Bull. Natl. Mus. Nat. Sci. Ser. 2013, 39, 25–51. [Google Scholar]
- Adam, J.H.; Omar, R.; Wilcock, C.C. Phytochemical Screening of Flavonoids in Three Hybrids of Nepenthes (Nepenthaceae) and Their Putative Parental Species from Sarawak and Sabah. J. Biol. Sci. 2002, 2, 623–625. [Google Scholar] [CrossRef][Green Version]
- Paper, D.H.; Karall, E.; Kremser, M.; Krenn, L. Comparison of the Antiinflammatory Effects of Drosera rotundifolia and Drosera madagascariensis in the HET-CAM Assay. Phytother. Res. 2005, 19, 323–326. [Google Scholar] [CrossRef]
- Fukushima, K.; Nagai, K.; Hoshi, Y.; Masumoto, S.; Mikami, I.; Takahashi, Y.; Oike, H.; Kobori, M. Drosera rotundifolia and Drosera tokaiensis Suppress the Activation of HMC-1 Human Mast Cells. J. Ethnopharmacol. 2009, 125, 90–96. [Google Scholar] [CrossRef]
- Fan, D.-H.; Wang, H.; Zhi, D.; Shen, Y.-M. CE Analysis of Endogenous Flavonoid Gallate Esters from Nepenthes gracilis (Nepenthaceae). Chromatographia 2010, 72, 1013–1016. [Google Scholar] [CrossRef]
- Marczak, Ł.; Kawiak, A.; Łojkowska, E.; Stobiecki, M. Secondary Metabolites In in Vitro Cultured Plants of the Genus Drosera. Phytochem. Anal. 2005, 16, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Ou-Yang, F.; Tsai, I.-H.; Tang, J.-Y.; Yen, C.-Y.; Cheng, Y.-B.; Farooqi, A.A.; Chen, S.-R.; Yu, S.-Y.; Kao, J.-K.; Chang, H.-W. Antiproliferation for Breast Cancer Cells by Ethyl Acetate Extract of Nepenthes thorellii x (ventricosa x maxima). Int. J. Mol. Sci. 2019, 20, 3238. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.; Ling, Y.S.; Wee, J.L.S.; Mujahid, A.; Müller, M. A Comparative UHPLC-Q/TOF–MS-Based Eco-Metabolomics Approach Reveals Temperature Adaptation of Four Nepenthes Species. Sci. Rep. 2020, 10, 21861. [Google Scholar] [CrossRef] [PubMed]
- Thanh, N.V.; Thao, N.P.; Huong, P.T.T.; Lee, S.H.; Jang, H.D.; Cuong, N.X.; Nam, N.H.; Kiem, P.V.; Kim, Y.H.; Minh, C.V. Naphthoquinone and Flavonoid Constituents from the Carnivorous Plant Nepenthes mirabilis and Their Anti-Osteoporotic and Antioxidant Activities. Phytochem. Lett. 2015, 11, 254–259. [Google Scholar] [CrossRef]
- Hake, A.; Begrow, F.; Spiegler, V.; Symma, N.; Hensel, A.; Düfer, M. Effects of Extracts and Flavonoids from Drosera rotundifolia L. on Ciliary Beat Frequency and Murine Airway Smooth Muscle. Molecules 2022, 27, 6622. [Google Scholar] [CrossRef]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A Comprehensive Review of Their Chemical Properties and Health Effects on Cardiovascular and Neurodegenerative Diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef]
- Bendokas, V.; Stanys, V.; Mažeikienė, I.; Trumbeckaite, S.; Baniene, R.; Liobikas, J. Anthocyanins: From the Field to the Antioxidants in the Body. Antioxidants 2020, 9, 819. [Google Scholar] [CrossRef]
- Timoneda, A.; Feng, T.; Sheehan, H.; Walker-Hale, N.; Pucker, B.; Lopez-Nieves, S.; Guo, R.; Brockington, S. The Evolution of Betalain Biosynthesis in Caryophyllales. New Phytol. 2019, 224, 71–85. [Google Scholar] [CrossRef]
- Foot, G.; Rice, S.P.; Millett, J. Red Trap Colour of the Carnivorous Plant Drosera rotundifolia Does Not Serve a Prey Attraction or Camouflage Function. Biol. Lett. 2014, 10, 20131024. [Google Scholar] [CrossRef]
- Dávila-Lara, A.; Reichelt, M.; Wang, D.; Vogel, H.; Mithöfer, A. Proof of Anthocyanins in the Carnivorous Plant Genus Nepenthes. FEBS Open Bio 2021, 11, 2576–2585. [Google Scholar] [CrossRef]
- Gregorio, G.J.D.; Dipalma, J.R. Anthocyanin in Dionaea muscipula Ellis (Venus Flytrap). Nature 1966, 212, 1264–1265. [Google Scholar] [CrossRef]
- Bendz, G.; Lindberg, G.; Buchardt, O.; Norin, T. Naphthoquinones and Anthocyanins from Two Drosera Species. Acta Chem. Scand. 1968, 22, 2722–2723. [Google Scholar] [CrossRef][Green Version]
- Ichiishi, S.; Nagamitsu, T.; Kondo, Y.; Iwashina, T.; Kondo, K.; Tagashira, N. Effects of Macro-Components and Sucrose in the Medium on in Vitro Red-Color Pigmentation in Dionaea muscipula Ellis and Drosera spathulata Labill. Plant Biotechnol. 1999, 16, 235–238. [Google Scholar] [CrossRef]
- Henarejos-Escudero, P.; Guadarrama-Flores, B.; García-Carmona, F.; Gandía-Herrero, F. Digestive Glands Extraction and Precise Pigment Analysis Support the Exclusion of the Carnivorous Plant Dionaea muscipula Ellis from the Caryophyllales Order. Plant Sci. 2018, 274, 342–348. [Google Scholar] [CrossRef]
- Meyer, G.W.; Bahamon Naranjo, M.A.; Widhalm, J.R. Convergent Evolution of Plant Specialized 1,4-Naphthoquinones: Metabolism, Trafficking, and Resistance to Their Allelopathic Effects. J. Exp. Bot. 2021, 72, 167–176. [Google Scholar] [CrossRef]
- Schlauer, J.; Nerz, J.; Rischer, H. Carnivorous Plant Chemistry. Acta Bot. Gall. 2005, 152, 187–195. [Google Scholar] [CrossRef]
- Eilenberg, H.; Pnini-Cohen, S.; Rahamim, Y.; Sionov, E.; Segal, E.; Carmeli, S.; Zilberstein, A. Induced Production of Antifungal Naphthoquinones in the Pitchers of the Carnivorous Plant Nepenthes khasiana. J. Exp. Bot. 2010, 61, 911–922. [Google Scholar] [CrossRef]
- Raj, G.; Kurup, R.; Hussain, A.A.; Baby, S. Distribution of Naphthoquinones, Plumbagin, Droserone, and 5-O-Methyl Droserone in Chitin-Induced and Uninduced Nepenthes khasiana: Molecular Events in Prey Capture. J. Exp. Bot. 2011, 62, 5429–5436. [Google Scholar] [CrossRef]
- Liu, W.; Lin, L.-C.; Wang, P.-J.; Chen, Y.-N.; Wang, S.-C.; Chuang, Y.-T.; Tsai, I.-H.; Yu, S.-Y.; Chang, F.-R.; Cheng, Y.-B.; et al. Nepenthes Ethyl Acetate Extract Provides Oxidative Stress-Dependent Anti-Leukemia Effects. Antioxidants 2021, 10, 1410. [Google Scholar] [CrossRef]
- Guan, H.-H.; Huang, Y.-H.; Lin, E.-S.; Chen, C.-J.; Huang, C.-Y. Plumbagin, a Natural Product with Potent Anticancer Activities, Binds to and Inhibits Dihydroorotase, a Key Enzyme in Pyrimidine Biosynthesis. Int. J. Mol. Sci. 2021, 22, 6861. [Google Scholar] [CrossRef] [PubMed]
- Rahman-Soad, A.; Dávila-Lara, A.; Paetz, C.; Mithöfer, A. Plumbagin, a Potent Naphthoquinone from Nepenthes Plants with Growth Inhibiting and Larvicidal Activities. Molecules 2021, 26, 825. [Google Scholar] [CrossRef] [PubMed]
- Dávila-Lara, A.; Rahman-Soad, A.; Reichelt, M.; Mithöfer, A. Carnivorous Nepenthes x ventrata Plants Use a Naphthoquinone as Phytoanticipin against Herbivory. PLoS ONE 2021, 16, e0258235. [Google Scholar] [CrossRef] [PubMed]
- Grevenstuk, T.; Gonçalves, S.; Domingos, T.; Quintas, C.; van der Hooft, J.J.; Vervoort, J.; Romano, A. Inhibitory Activity of Plumbagin Produced by Drosera intermedia on Food Spoilage Fungi. J. Sci. Food Agric. 2012, 92, 1638–1642. [Google Scholar] [CrossRef]
- Tian, J.; Chen, Y.; Ma, B.; He, J.; Tong, J.; Wang, Y. Drosera peltata Smith var. lunata (Buch.-Ham.) C.B. Clarke as a Feasible Source of Plumbagin: Phytochemical Analysis and Antifungal Activity Assay. World J. Microbiol. Biotechnol. 2014, 30, 737–745. [Google Scholar] [CrossRef]
- Putalun, W.; Udomsin, O.; Yusakul, G.; Juengwatanatrakul, T.; Sakamoto, S.; Tanaka, H. Enhanced Plumbagin Production from in Vitro Cultures of Drosera burmanii Using Elicitation. Biotechnol. Lett. 2010, 32, 721–724. [Google Scholar] [CrossRef]
- Adamec, L.; Gastinel, L.; Schlauer, J. Plumbagin Content in Aldrovanda vesiculosa Shoots. Carniv. Plant Newslett. 2006, 35, 52–55. [Google Scholar]
- Kawiak, A.; Królicka, A.; Łojkowska, E. In Vitro Cultures of Drosera aliciae as a Source of a Cytotoxic Naphthoquinone: Ramentaceone. Biotechnol. Lett. 2011, 33, 2309–2316. [Google Scholar] [CrossRef]
- Kawiak, A.; Zawacka-Pankau, J.; Wasilewska, A.; Stasilojc, G.; Bigda, J.; Lojkowska, E. Induction of Apoptosis in HL-60 Cells through the ROS-Mediated Mitochondrial Pathway by Ramentaceone from Drosera aliciae. J. Nat. Prod. 2012, 75, 9–14. [Google Scholar] [CrossRef]
- Aung, H.H.; Chia, L.S.; Goh, N.K.; Chia, T.F.; Ahmed, A.A.; Pare, P.W.; Mabry, T.J. Phenolic Constituents from the Leaves of the Carnivorous Plant Nepenthes gracilis. Fitoterapia 2002, 73, 445–447. [Google Scholar] [CrossRef]
- Picazo-Aragonés, J.; Terrab, A.; Balao, F. Plant Volatile Organic Compounds Evolution: Transcriptional Regulation, Epigenetics and Polyploidy. Int. J. Mol. Sci. 2020, 21, 8956. [Google Scholar] [CrossRef]
- Di Giusto, B.; Bessière, J.-M.; Guéroult, M.; Lim, L.B.L.; Marshall, D.J.; Hossaert-McKey, M.; Gaume, L. Flower-Scent Mimicry Masks a Deadly Trap in the Carnivorous Plant Nepenthes rafflesiana: Flower-Scent Mimicry in Nepenthes rafflesiana. J. Ecol. 2010, 98, 845–856. [Google Scholar] [CrossRef]
- Wells, K. Pitchers of Nepenthes rajah Collect Faecal Droppings from Both Diurnal and Nocturnal Small Mammals and Emit Fruity Odour. J. Trop. Ecol. 2011, 27, 347–353. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Scheerer, U.; Kruse, J.; Burzlaff, T.; Honsel, A.; Alfarraj, S.; Georgiev, P.; Schnitzler, J.-P.; Ghirardo, A.; Kreuzer, I.; et al. The Venus Flytrap Attracts Insects by the Release of Volatile Organic Compounds. J. Exp. Bot. 2014, 65, 755–766. [Google Scholar] [CrossRef]
- Hatano, N.; Hamada, T. Proteomic Analysis of Secreted Protein Induced by a Component of Prey in Pitcher Fluid of the Carnivorous Plant Nepenthes alata. J. Proteom. 2012, 75, 4844–4852. [Google Scholar] [CrossRef]
- Rottloff, S.; Miguel, S.; Biteau, F.; Nisse, E.; Hammann, P.; Kuhn, L.; Chicher, J.; Bazile, V.; Gaume, L.; Mignard, B.; et al. Proteome Analysis of Digestive Fluids in Nepenthes Pitchers. Ann. Bot. 2016, 117, 479–495. [Google Scholar] [CrossRef]
- Jakšová, J.; Adamec, L.; Petřík, I.; Novák, O.; Šebela, M.; Pavlovič, A. Contrasting Effect of Prey Capture on Jasmonate Accumulation in Two Genera of Aquatic Carnivorous Plants (Aldrovanda, Utricularia). Plant Physiol. Biochem. 2021, 166, 459–465. [Google Scholar] [CrossRef]
- Buch, F.; Rott, M.; Rottloff, S.; Paetz, C.; Hilke, I.; Raessler, M.; Mithöfer, A. Secreted Pitfall-Trap Fluid of Carnivorous Nepenthes Plants Is Unsuitable for Microbial Growth. Ann. Bot. 2013, 111, 375–383. [Google Scholar] [CrossRef]
- Tang, J.; Peng, S.; Cheng, Y.; Wang, C.; Farooqi, A.A.; Yu, T.; Hou, M.; Wang, S.; Yen, C.; Chan, L.; et al. Ethyl Acetate Extract of Nepenthes adrianii x clipeata Induces Antiproliferation, Apoptosis, and DNA Damage against Oral Cancer Cells through Oxidative Stress. Environ. Toxicol. 2019, 34, 891–901. [Google Scholar] [CrossRef]
- Zehl, M.; Braunberger, C.; Conrad, J.; Crnogorac, M.; Krasteva, S.; Vogler, B.; Beifuss, U.; Krenn, L. Identification and Quantification of Flavonoids and Ellagic Acid Derivatives in Therapeutically Important Drosera Species by LC–DAD, LC–NMR, NMR, and LC–MS. Anal. Bioanal. Chem. 2011, 400, 2565–2576. [Google Scholar] [CrossRef]
- Krolicka, A.; Szpitter, A.; Maciag, M.; Biskup, E.; Gilgenast, E.; Romanik, G.; Kaminski, M.; Wegrzyn, G.; Lojkowska, E. Antibacterial and Antioxidant Activity of the Secondary Metabolites from in Vitro Cultures of the Alice sundew (Drosera aliciae). Biotechnol. Appl. Biochem. 2008, 53, 175–184. [Google Scholar] [CrossRef]
- Ismail, N.A.; Kamariah, A.S.; Lim, L.B.L.; Ahmad, N. Phytochemical and Pharmacological Evaluation of Methanolic Extracts of the Leaves of Nepenthes bicalcarata Hook. F. Int. J. Pharmacogn. Phytochem. Res. 2015, 7, 1127–1138. [Google Scholar]
- Lin, E.-S.; Huang, C.-Y. Cytotoxic Activities and the Allantoinase Inhibitory Effect of the Leaf Extract of the Carnivorous Pitcher Plant Nepenthes Miranda. Plants 2022, 11, 2265. [Google Scholar] [CrossRef] [PubMed]
- Rodzali, N.N.; Mydin, M.M. Antibacterial Activity of Leaves and Pitchers Extract of Nepenthes gracilis against Bacillus subtilis and Escherichia coli. J. Fundam. Appl. Sci. 2018, 9, 81–88. [Google Scholar] [CrossRef]
- Gerschler, S.; Guenther, S.; Schulze, C. Antibiofilm Activity of Sundew Species against Multidrug-Resistant Escherichia coli Strains. Int. J. Mol. Sci. 2022, 23, 13720. [Google Scholar] [CrossRef] [PubMed]
- Kawiak, A.; Domachowska, A.; Krolicka, A.; Smolarska, M.; Lojkowska, E. 3-Chloroplumbagin Induces Cell Death in Breast Cancer Cells Through MAPK-Mediated Mcl-1 Inhibition. Front. Pharmacol. 2019, 10, 784. [Google Scholar] [CrossRef] [PubMed]
- De, U.; Son, J.Y.; Jeon, Y.; Ha, S.-Y.; Park, Y.J.; Yoon, S.; Ha, K.-T.; Choi, W.S.; Lee, B.M.; Kim, I.S.; et al. Plumbagin from a Tropical Pitcher Plant (Nepenthes alata Blanco) Induces Apoptotic Cell Death via a P53-Dependent Pathway in MCF-7 Human Breast Cancer Cells. Food Chem. Toxicol. 2019, 123, 492–500. [Google Scholar] [CrossRef]
- Gwee, P.S.; Khoo, K.S.; Ong, H.C.; Sit, N.W. Bioactivity-Guided Isolation and Structural Characterization of the Antifungal Compound, Plumbagin, from Nepenthes gracilis. Pharm. Biol. 2014, 52, 1526–1531. [Google Scholar] [CrossRef]
- Thao, N.P.; Luyen, B.T.T.; Koo, J.E.; Kim, S.; Koh, Y.S.; Thanh, N.V.; Cuong, N.X.; Kiem, P.V.; Minh, C.V.; Kim, Y.H. In Vitro Anti-Inflammatory Components Isolated from the Carnivorous Plant Nepenthes mirabilis (Lour.) Rafarin. Pharm. Biol. 2016, 54, 588–594. [Google Scholar] [CrossRef]
- Thanh, N.V.; Thao, N.P.; Dat, L.D.; Huong, P.T.T.; Lee, S.H.; Jang, H.D.; Cuong, N.X.; Nam, N.H.; Kiem, P.V.; Minh, C.V.; et al. Two New Naphthalene Glucosides and Other Bioactive Compounds from the Carnivorous Plant Nepenthes mirabilis. Arch. Pharm. Res. 2015, 38, 1774–1782. [Google Scholar] [CrossRef]
- Babula, P.; Adam, V.; Havel, L.; Kizek, R. Naftochinony a Jejich Farmakologické Vlastnosti [Naphthoquinones and Their Pharmacological Properties]. Ceska Slov. Farm. 2007, 56, 114–120. [Google Scholar]
- Liu, Y.; Cai, Y.; He, C.; Chen, M.; Li, H. Anticancer Properties and Pharmaceutical Applications of Plumbagin: A Review. Am. J. Chin. Med. 2017, 45, 423–441. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P. Arabinogalactan Proteins in the Digestive Glands of Dionaea muscipula J.Ellis Traps. Cells 2022, 11, 586. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P.; Strzemski, M.; Miranda, V.F.O. Immunocytochemical Analysis of the Wall Ingrowths in the Digestive Gland Transfer Cells in Aldrovanda vesiculosa L. (Droseraceae). Cells 2022, 11, 2218. [Google Scholar] [CrossRef]
- Rey, M.; Yang, M.; Lee, L.; Zhang, Y.; Sheff, J.G.; Sensen, C.W.; Mrazek, H.; Halada, P.; Man, P.; McCarville, J.L.; et al. Addressing Proteolytic Efficiency in Enzymatic Degradation Therapy for Celiac Disease. Sci. Rep. 2016, 6, 30980. [Google Scholar] [CrossRef]
- Mithöfer, A. A Spotlight on Prey-Induced Metabolite Dynamics in Sundew. A Commentary on: ‘Metabolomic Analysis Reveals Reliance on Secondary Plant Metabolites to Facilitate Carnivory in the Cape Sundew, Drosera capensis. Ann. Bot. 2021, 128, v–vi. [Google Scholar] [CrossRef]
- Iosip, A.-L.; Scherzer, S.; Bauer, S.; Becker, D.; Krischke, M.; Al-Rasheid, K.A.S.; Schultz, J.; Kreuzer, I.; Hedrich, R. DYSCALCULIA, a Venus Flytrap Mutant without the Ability to Count Action Potentials. Curr. Biol. 2023, 33, 589–596.e5. [Google Scholar] [CrossRef]
- Wawrosch, C. An Improved 2-Step Liquid Culture System for Efficient In Vitro Shoot Proliferation of Sundew (Drosera rotundifolia L.). Sci. Pharm. 2009, 77, 827–836. [Google Scholar] [CrossRef]
- Wawrosch, C.; Vackar, E.; Grauwald, B. Variations of Naphthoquinone Levels in Micropropaaated Drosera Species In Vitro, under Qreenhouse and Outdoor Growth Conditions. Sci. Pharm. 2005, 73, 251–262. [Google Scholar] [CrossRef][Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Investigated Species | Compound | Method | Ref. |
|---|---|---|---|
| phenols/phenolic acids and derivatives | |||
| N. anamensis (leaf, trap) | gallic: 58.9 (l), 53.7 (t) µg/g; protocatechuic: 4.48 (l), 15.1 (t) µg/g; hydroxybenzoic: 4.28 (l), 9.21 (t) µg/g; vanillic: 2.53 (l), 9.93 (f) µg/g; chlorogenic: 32.4 (l) 53.8 (f) µg/g; syringic: 10.7 (l), 14.7 (f) µg/g; caffeic: 2.6 (l), 3.57 (f) µg/g; ferulic: 1.07 (l), 7.97 (f) µg/g; sinapic: 7.09 (l) 15.7 (f) µg/g; p-coumaric 2.08 (l), 6.77 (t) µg/g; vanillin: 0.85 (l), 0.94 (f) µg/g dw | HPLC-MS | [32] |
| flavonoids: flavonols | |||
| N. minima, N. ampullaria, N. rafflesiana, N. northiana | quercetin (Q), Q 3-O-rhamnoside, Q 3-(6″-galloylglucoside), Q 3-O-(6″-n-butyl ß-D-glucuronide), rutin, miquelianin, kaempferol (K) 3-glucoside, K 3-O-beta-D-xyloside, 6,8-dihydroxy-K 3-rutinoside, afzelin, luteolin, baicalin, butin, myrciacitrin | UHPLC-Q/TOF–MS | [49] |
| N. gracilis (leaves) | quercetin esters: 3-O-ß-(3″-O-galloylxylopyranoside), 3-O-α-(3″-O-galloylrhamnopyranoside), 3-O-ß-(2″-O-galloylxylopyranoside), 3-O-α-(2″-galloylarabinofuranoside), 3-O-ß-(6″-galloylglucopyranoside) | CC/NMR, MS | [46] |
| N. thorellii × (ventricosa × maxima) (aerial parts) | quercetin 3-O-(6″-n-butyl-D-glucuronide) | HPLC-DAD, CC/NMR | [48] |
| N. mirabilis (branches and leaves) | quercitrin, quercetin (Q), Q-3-O-b-D-glucuronide, K-3-O-a-L-rhamnoside | CC/NMR, MS | [50] |
| flavonoids: anthocyanins * | |||
| N. × ventrata, N. thorelii, N. ventricosa, N. robcantleyi, N. maxima, N. fusca, N. mirabilis (pitchers: digestive zone—dz, peristome—p) | cyanidin-3-O-glucoside: ca 0.5–6 (p), up to ca 2 (dz) µM/g fw cyanidin-3-O-galactoside: ca 0.1–0.6 (p), up to ca 0.3 (dz) µM/g fw cyanidin-3-O-glucuronide: ca 0–0.1 (p, dz) µM/g fw | UHPLC/HRMS | [56] |
| other phenolic compounds | |||
| N. mirabili (branches and leaves)s | epicatechin | CC/NMR, MS | [50] |
| N. gracilis (leaves) | epicatechin 3-O-gallate | CC/NMR, MS | [46] |
| N. minima, N. ampullaria, N. rafflesiana, N. northiana (aerial parts) | syringin, catechin 5-O-gallate, coniferin, 5-galloylshikimic acid | UHPLC-Q/TOF–MS | [49] |
| naphthoquinones | |||
| N. cv. ‘Miranda’ (leaves) | plumbagin | GC-MS | [66] |
| Nepenthes × ventrata (N. alata × ventricosa) (pitcher, leaves) | plumbagin | NMR | [67] |
| Nepenthes thorellii × (ventricosa × maxima) (aerial parts) | plumbagin | HPLC-DAD, CC/NMR | [48] |
| N. × ventrata (N. alata × N. ventricosa) (leaves) | plumbagin | LC-MS/MS | [68] |
| N. khasiana (root, stem, leaves, pitchers), | plumbagin: ca 1.4% (r), 0.2% (s), 0.4% (l), 0.04% (pi) dw * droserone, 5-O-methyl droserone (pi) | GC-MS/NMR | [64] |
| N. khasiana (pitcher liquid) | 5-O-methyl droserone, droserone | HPLC/MS, UV, NMR | [63] |
| N. mirabilis (branches and leaves) | 3-methoxy-7-methyljuglone2-methoxy-7-methyljuglone, plumbagin, droserone, nepenthones C, F and G | CC/NMR, MS | [50] |
| N. alata, N. fusca, N. gracilis, N. mirabilis, N. superba, N. thorelii, N. ventricosa (pitcher fluids) | plumbagin, 7-methyl-juglone | LC/MS, NMR | [83] |
| N. adrianii × clipeata (twigs and leaves) | isoplumbagin | Isolation/NMR | [84] |
| acetogenic tetralone | |||
| N. gracilis (leaves), | Isoshinanolone, shinanolone epishinanolone | Isolation/MS, NMR | [75] |
| N. thorellii × (ventricosa × maxima) (aerial parts) | isoshinanolone | HPLC-DAD, NMR | [48] |
| N. mirabilis (branches and leaves) | cis-isoshinanolone | CC/NMR, MS | [50] |
| alkaloids | |||
| N. minima, N. ampullaria, N. rafflesiana, N. northiana (aerial parts) | trigonelline, anatoxin a, berberastine | UHPLC-Q/TOF–MS | [49] |
| Species | Compound | Method | Refs. |
|---|---|---|---|
| phenols/phenolic acids and derivatives | |||
| Dr. capensis (leaf, trap) | gallic: 167.8 (l), 195.5 (t) µg/g; protocatechuic: 1.26 (l), 12.1 (t) µg/g; chlorogenic: 28.5 (l), 29.6 (t) µg/g; ferulic: 1.15 (l), 2.21 (t) µg/g; p-coumaric: 1.52 (l), 2.06 (t) µg/g; hydroxybenzoic: 3.72 (l), 17.5 (t) µg/g; vanillic: 1.57 (l) 2.07 (t) µg/g; syringic: 7.86 (l) 10.1 (t) µg/g; caffeic: 1.91 (l) 3.01 (t) µg/g; sinapic: 0.18 (l), 0.69 (t) µg/g; vanillin: 1.93 (l), 1.62 (t) µg/g dw | HPLC-MS | [32] |
| Dr. rotundifolia, Dr. tokaiensis, Dr. spatulata | ellagic acid (EA) | HPLC | [45] |
| Dr. tokaiensis (aboveground part) | 3,3-di-O-methylellagic acid 4′-glucoside | HPLC/NMR | [34] |
| Dr. magna (bulbs and leaves) | 1-O-protochatechoyl glucoside | Isolation/NMR | [33] |
| Dr. rotundifolia field (f), in vitro propagated (p) | quinic acid (fp), monogalloyl glucose (fp), digalloyl glucose (f), coumaric acid glycoside (fp), EA (fp), EA glycoside (fp), dimethylEA glycoside ((fp), dimethylEA (fp), methylEA (f) | UPLC/DAD/MS-MS | [20] |
| Dr. anglica, Dr. intermedia, Dr. madagascariensis, Dr. rotundifolia (aerial part) | ellagic acid (0.137–1.107%), 3,3′-di-O-methylEA (0.084–0.143%) dw | LC-MS/NMR | [85] |
| Dr. binata in vitro culture | 3,3-di-O-methylellagic acid | HPLC-DAD-MS | [36] |
| flavonoids | |||
| Dr. binata | methylated myricetin (M) methylated M 3-O-glucoside, quercetin (Q), Q 3-O-glucoside; isorhamnetin (iRh) or rhamnetin (Rh); Rh/isoRh 3-O-glucoside; kaempferol (K)/fisetin (F) K/F 3-O-glucoside; K/F-3-O-rhamnosylglucoside, | HPLC-UV/MS | [47] |
| Dr. rotundifolia field (f), in vitro propagated (p) | myricetin glycoside (fp), hyperoside (fp), galloylhyperoside (fp), hydroxybenzoylhyperin (fp), dihydromyricetin (f), hexahydroxyflavonegalloyl glycoside (f), tetrahydroxyflavone (f), kaempferol-galloylglycoside (f), quercetin (Q)(f), Q-glycoside (f) Q-glycoside gallate (f), syringetin glycoside (p), spinatoside (p) | UPLC/DAD/MS | [20] |
| Dr. rotundifolia | quercetin, hyperoside and 2″-O-galloylhyperoside | UHPLC-TOF-MS | [51] |
| Dr. tokaiensis (aboveground part) | myricitrine, quercimelin | HPLC/NMR | [34] |
| Dr. magna (bulbs and leaves) | tamarixetin-3-rhamnoside, naringenin-6-C-β-D-glucopyranoside, hirsutrin, three new flavonol diglycosides, flavan-3-ol glycoside | Isolation/NMR | [33] |
| Dr. rotundifolia Dr. anglica, Dr. intermedia, Dr. madagascariensis, (aerial part) | myricetin (M) (<0.003–0.097%), M-3-O-β-galactopyranoside (0.038–0.275%), M-3-O-β-glucopyranoside, hyperoside (H) (0.048–1.530%), 2″-O-galloylH (0–2.515%), isoquercitrin (0.032–0.421%), kaempferol (K), K-3-O-β-galactopyranoside, K-3-O-(2″-O-galloyl)-β-galactopyranoside, astragalin, quercetin (0.056–0.187%) dw | LC-MS/NMR | [85] |
| naphthoquinones | |||
| Dr. binata, Dr. adelae, Dr. aliciae, Dr. capensis, Dr. cuneifolia, Dr. ramentacea | plumbagin: 0.001–0.059% fw | HPLC-UV/MS | [47] |
| Dr. binata, Dr. gigantea | plumbagin: 2.04 and 0.15 mg/g fw | HPLC-DAD-MS | [35] |
| Dr. binata | plumbagin, 3-chloroplumbagin | HPLC-DAD-MS | [36] |
| Dr. intermedia, | plumbagin | HPLC-MS/NMR | [69] |
| Dr. burmanii | plumbagin | ELISA | [71] |
| Dr. peltata Smith var. lunata | plumbagin: 11.05 mg/g dw | TLC/NMR | [70] |
| Dr. magna (bulbs and leaves) | hydroxydroserone (H), H-5-O-β-D-glucoside, droserone, plumbagin, 2,3-dihydronapthalene-1,4-diones | Isolation/NMR | [33] |
| Dr. rotundifolia field (f), in vitro propagated (p) | 7-methyljuglone, 7-methyljuglone diglycoside, 7-methyljuglone glycoside (f,p) | UPLC/DAD/MS-MS | [20] |
| Dr. aliciae in vitro culture | ramentaceone | Isolation/NMR, MS | [73,74] |
| Species | Compound | Method | Ref. |
|---|---|---|---|
| phenols/phenolic acids and derivatives | |||
| Di. muscipula (leaf, trap) | gallic: 187.8 (l), 167.4 (t) µg/g; protocatechuic: 2.74 (l), 3.21 (t) µg/g; chlorogenic: 24.5 (l), 30.8 (t) µg/g; ferulic: 0.65 (l), 0.69 (t) µg/g; p-coumaric: 0.57 (l), 0.76 (t) µg/g; salicylic: 0.28 (l), 0.19 (t) µg/g; hydroxybenzoic: 4.26 (l), 4.53 (t) µg/g; vanillic: 4.84 (l), 6.87 (t) µg/g; syringic: 8.78 (l), 12.2 (t) µg/g; caffeic: 10.8 (l), 12.0 (t) µg/g; sinapic: 17.0 (l), 42.2 (t) µg/g; vanillin: 2.45 (l), 1.97 (t) µg/g dw | HPLC-MS | [32] |
| Di. muscipula | ellagic, dimethylellagic acid isomer, 3-O-methylellagic acid, 3,3′-di-O-methylellagic acid | HPLC-DAD-MS | [35] |
| Di. muscipula | caffeic: ca 0.18 mg/g, salicylic: ca 300 mg/g, ellagic: ca 0.18 mg/g * | HPLC-DAD | [37] |
| Di. muscipula | chlorogenic:0.26 mg/g, p-coumaric: 0.04 mg/g ferulic: 0.16 mg/g, gallic: 0.31 mg/g, protocatechuic: 0.29 mg/g dw | HPLC-DAD | [22] |
| flavonoids: flavonols | |||
| Di. muscipula | myricetin: ca 20 mg/g, hyperoside: ca 0.3 mg/g, quercetin: 15 mg/g dw * | HPLC-DAD | [37] |
| Di. muscipula | kaempferol, quercetin after hydrolysis (leaf, trap) | HPLC-MS | [32] |
| Di. muscipula | kaempferol: 0.59 mg/g dw | HPLC-DAD | [22] |
| Di. muscipula | hyperoside, quercetin-3-(6″-O-galloyl)-glucoside/galactoside, kaempferol-3-(6″-O-galloyl)-glucoside | HPLC-DAD-MS | [35] |
| flavonoids: anthocyanins | |||
| Di. muscipula (digestive glands) | delphinidin-3-O-glucoside, cyanidin-3-O-glucoside, cyanidin | HPLC/MS-MS | [60] |
| naphthoquinones | |||
| Di. muscipula | plumbagin: 3.45 mg/g fw, dihydroplumbagin, 3-chloroplumbagin, 8,8′-biplumbagin | HPLC/DAD-MS | [35] |
| Di. muscipula | plumbagin: ca 50 mg/g dw * | HPLC-DAD | [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wójciak, M.; Feldo, M.; Stolarczyk, P.; Płachno, B.J. Carnivorous Plants from Nepenthaceae and Droseraceae as a Source of Secondary Metabolites. Molecules 2023, 28, 2155. https://doi.org/10.3390/molecules28052155
Wójciak M, Feldo M, Stolarczyk P, Płachno BJ. Carnivorous Plants from Nepenthaceae and Droseraceae as a Source of Secondary Metabolites. Molecules. 2023; 28(5):2155. https://doi.org/10.3390/molecules28052155
Chicago/Turabian StyleWójciak, Magdalena, Marcin Feldo, Piotr Stolarczyk, and Bartosz J. Płachno. 2023. "Carnivorous Plants from Nepenthaceae and Droseraceae as a Source of Secondary Metabolites" Molecules 28, no. 5: 2155. https://doi.org/10.3390/molecules28052155
APA StyleWójciak, M., Feldo, M., Stolarczyk, P., & Płachno, B. J. (2023). Carnivorous Plants from Nepenthaceae and Droseraceae as a Source of Secondary Metabolites. Molecules, 28(5), 2155. https://doi.org/10.3390/molecules28052155