
Renoprotective Effects of Tanshinone IIA: A Literature Review

 ,
,
Abstract
1. Introduction
1.1. Kidney Diseases
1.2. Overview of Tanshinone IIA
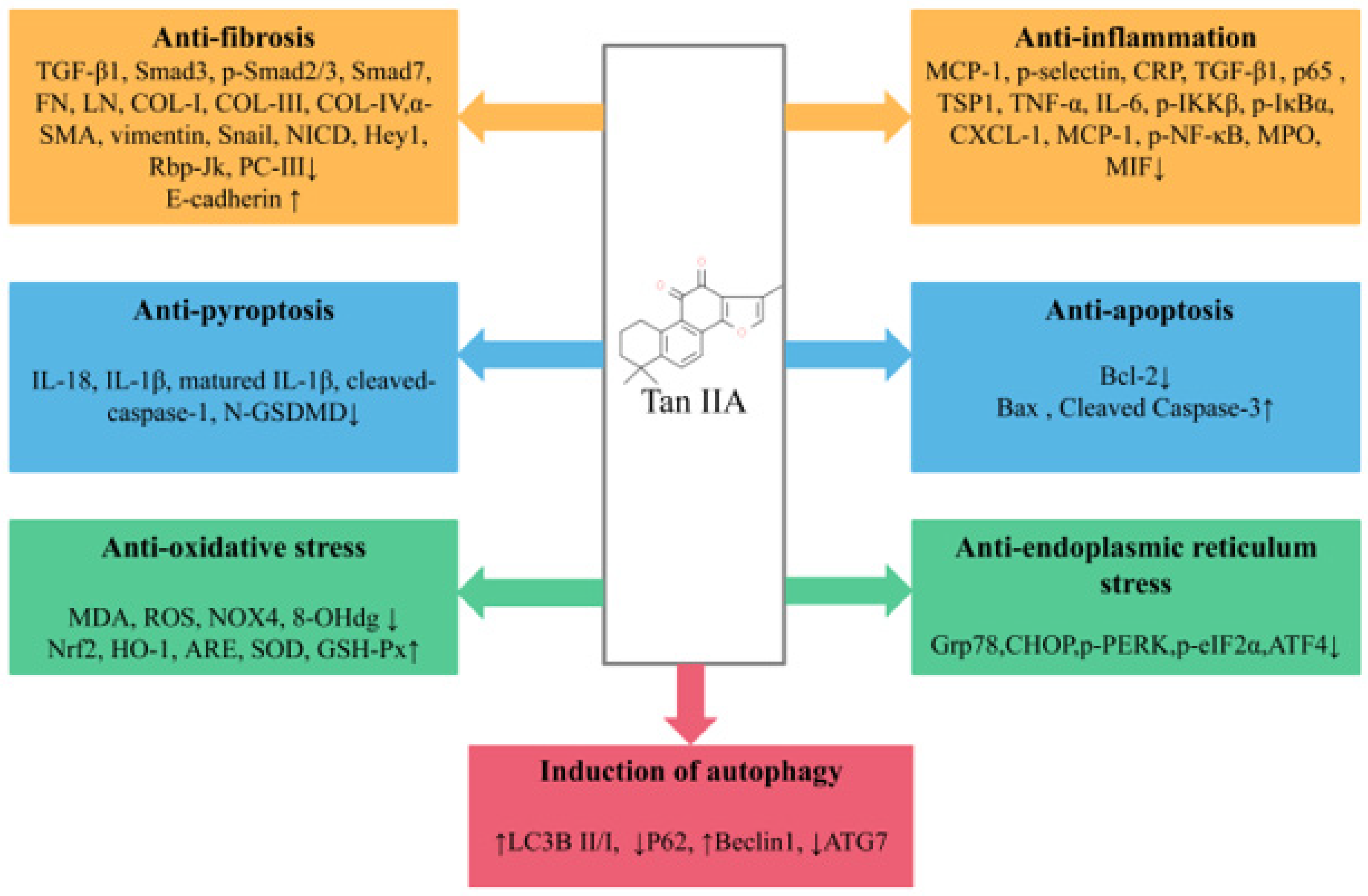
2. Renal Protective Effects of Tan IIA
2.1. Diabetic Nephropathy
2.2. Renal Fibrosis
2.3. Uric Acid Nephropathy
2.4. Renal Cell Carcinoma
2.5. Renal Ischemia and Reperfusion Injury
2.6. Drug-Induced Kidney Injury
2.6.1. Cisplatin-Induced Renal Injury
2.6.2. Vancomycin-Induced Renal Injury
2.6.3. APAP-Induced Kidney Injury
2.7. Contrast-Induced Nephropathy
2.8. Henoch–Schonlein Purpura Nephritis
2.9. Hypertensive Renal Damage
2.10. Nephrotic Syndrome
2.11. Acute Kidney Injury Induced by Sepsis

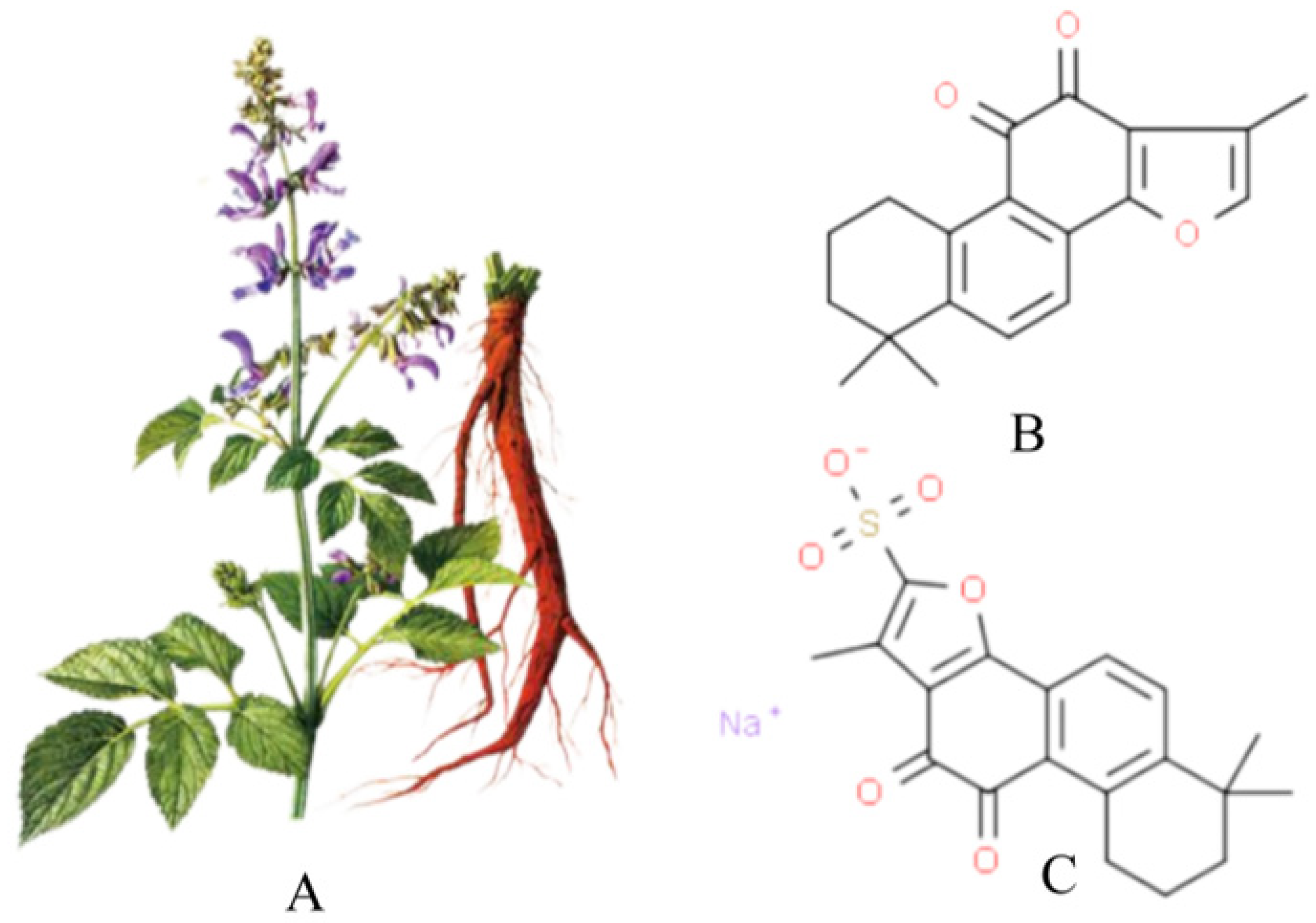{kind=link}
{kind=link}
| Disease | Models and Modeling Methods | Research Type | Tan IIA or Derivatives Dose and Treatment Schedule | Targets | References | |
|---|---|---|---|---|---|---|
| DN | SD rats; intraperitoneal injection of STZ (65 mg/kg.d) for 3 d | In vivo | Tan IIA; 10 mg/kg.d; p.o. for 12 weeks | ↓ MDA, ↑SOD, ↓ MCP-1, ↓ TGF-β1, ↓ p-selectin, ↓ CRP in renal tissue; ↓ MCP-1, ↓ TGF-β1, ↓ p-selectin, ↓ CRP in serum | [29] | |
| HBZY-1 cells; HG (30 mmol/L) for 48 h; SD rats; intraperitoneal injection of STZ (45 mg/kg.d) combined with high-fat diet for 7 weeks | In vitro and in vivo | Tan IIA; 20, 40, and 80 µM for 48 h; 8 mg/kg.d, i.m. for 3 weeks | ↓ TGFβ, ↓ p65 in cell; ↓ TGFβ, ↓ p65 in renal tissue | [30] | ||
| SD rats; intraperitoneal injection of STZ (60 mg/kg) for 2 d | In vivo | Tan IIA; 2, 4, 8 mg/kg.day, i.p. for 6 weeks | ↓ TGF-β1, ↓ TSP1, ↓ Grp78, ↓ CHOP, ↓ p-PERK, ↓ p-eIF2α, ↓ ATF4, ↑SOD in renal tissue | [31] | ||
| HK-2 cells; HG (30 mmol/L) for 48 h | In vitro | Tan IIA; 1, 5, 10 μM for 24 h | ↓ TGF-β1, ↓ TNF-α, ↓ IL-6, -caspase-3, -caspase-9, -GRP78, -CHOP, cleaved caspase-12, ↓ IL-18, ↓ IL-1β, ↓ matured IL-1β, ↓ cleaved-caspase-1, ↓ N-GSDMD, ↓ FN, ↓ COL-Ⅰ, ↓ p65, ↓ p-IKKβ, ↓ p-IκBαin cell | [32] | ||
| SD rats; STZ (65 mg/kg) was injected intraperitoneally for 72 h. | In vivo | STS; 10, 20 mg/kg.day, i.p. for 12 weeks | ↑ SOD, ↑ GSH-Px, ↓ MDA, ↓ ET, ↓ TXB2, ↓ 6-keto-PGF1α, ↓ ac FoxO1, ↑ SIRT1 in renal tissue | [33] | ||
| HMCs; AGES (1, 10, 50, 100 μg/mL) for 48 h | In vitro | STS; 0, 0.1, 1.0, 5.0, 10.0 μg/mL for 48 h | ↓ RAGE, ↑ SOD, ↑ GSH-Px, ↓ MDA in cells | [34] | ||
| RF | HK-2 cells; HG (30 mmol/L) for 24 h | In vitro | Tan IIA; 6 μg/mL for 24 h | ↓ TGF-β1, ↓ α-SMA, ↑ E-cadherin, ↓ FN, ↓ LN, ↑ HO-1 in cells | [37] | |
| HK-2 cells; HG (30 mmol/L) for 48 h | In vitro | Tan IIA; 1, 10, 50 µM for 48 h | ↑ E-cadherin, ↓ α-SMA, ↓ vimentin, ↓ FN, ↓ Snail in cells | [38] | ||
| HK-2 cells; HG (30 mmol/L) for 48 h | In vitro | Tan IIA; 5 μM, 10 μM for 48 h | ↓ α-SMA, ↑ E-cadherin, ↑ VDR, ↓ β-catenin, ↓ GSK-3β, in cells | [39] | ||
| Kunming mice; intraperitoneal injection of STZ (50mg/kg.d) three times every other day; HK-2 cell; HG (30 mmol/L) for 48 h | In vitro and In vivo | Tan IIA; 5, 10, 25 mg/kg p.o. for 30d Tan IIA; 5 mg/L, 10 mg/L, 25 mg/L for 30 min | ↑ LC 3BII/LC 3BI, ↓ P62↑ Beclin1, ↓ ATG7, ↓ Notch1, ↓ p-AKT, ↓ p-mTOR, ↑ PTEN, ↑ miR-34a-5p in renal tissue; ↑ LC 3BII/LC 3BI, ↓ P62↑ Beclin1, ↓ ATG7, ↓ Notch1, ↓ p-AKT, ↓ p-mTOR, ↑ PTEN, ↑ miR-34a-5p, ↓ Col- I, ↓ Col-III in cells | [40] | ||
| SD rats; 5/6 nephrectomy for 1 week | In vivo | Tan IIA; 10 mg/kg.d, p.o. for 16 weeks | ↓ FN, -COL-Ⅰ, ↓ COL-III, ↓ COL-IV, ↓ TGF-β1, ↓ TNF-α, ↓ CXCL-1, ↓ MCP-1, -RANTES, ↓ Smad3, ↓ p-Smad2/3, ↓ Smad7, -IKKβ, ↓ p-IKKβ, -IκBα, ↓ p-IκBα, -NF-κB, ↓ p-NF-κB in renal tissue | [41] | ||
| C57BL/6 mice; single intraperitoneal injection of folic acid (250 mg/kg) | In vivo | Tan IIA; tail vein injection, 15 mg/kg.d for 2 days | ↓ TGF-β1, ↓ MCP-1 in renal tissue | [42] | ||
| NRK/49F cells; TGFβ1 (5 ng/mL) for 0, 6, 12, 24 h | In vitro | Tan IIA; 10−6, 10−5, 10−4 mol/L for 2 h | ↓ FN, ↓ p-Smad2/3, -total Smad2, -total Smad3 in cells | [43] | ||
| SD rats; unilateral ureteral obstruction | In vivo | Tan IIA; 25 mg/kg.d, DAPT 12 mg/kg.d, Tan IIA; 25 mg/kg.d+DAPT 12mg/kg.d for 2 weeks | ↓ NICD, ↓ Hey1, ↓ Rbp-Jk, ↓ Col-Ⅰin renal tissue | [44] | ||
| SD rats; adenine solution (250 mg/kg.d) was administered by gavage for 3 weeks. | In vivo | Tan IIA; 15.0 mg/kg.d for 4 weeks | ↓ COL-Ⅳ, ↓ PC-Ⅲ, ↓ LN, ↑ FN in serum | [45] | ||
| UAN | SD rats; adenine (30 mg/kg.d) was administered by gavage for 18 d. | In vivo | Tan IIA; 3, 1.5, 0.75 g/mg/kg.d, p.o. for 23 days | ↓ MCP-1, ↓ IL-1β, ↓ Inucleus NF-κB, ↑ cytosome NF-κB, in renal tissue | [48] | |
| Kunming mice; adenine (100 mg/kg.d) and potassium oxalate (150 mg/kg.d) for 28 d HK-2 cells; stimulation with uric acid (0.2 mg/L) for 24 h | In vitro and In vivo | Tan IIA; 4, 8 or 16 mg/kg.d, p.o. for 28 d Tan IIA; 1, 5, and 10 μM for 24 h | ↓ NOX4, ↓ p22phox, ↓ iNOS, ↓ COX-2, ↓ p-p44/42, ↑ p44/42, ↑ p38, ↓ p-p38, ↓ p-JNK, ↑ JNK in renal tissue, ↓ ROS, ↓ NOX4, ↓ p22phox, ↓ p-p44/42, ↑ p44/42, ↑ p38, ↓ p-p38, ↓ p-JNK, ↑ JNK in cells | [49] | ||
| RCC | Human renal carcinoma cell line 786-O | In vitro | Tan IIA; 1, 2, 4, 8 µg/mL for 24 h | ↑ p53, ↑ p21, ↑ Bax, ↑ caspase-3 in cells | [51] | |
| Human renal carcinoma cell line 786-O and human renal carcinoma Caki-1 cell line | In vitro | Tan IIA; 1, 5, 10 or 30 mM for 24 h | -PARP, -caspase-3, -Bcl-2, -Alix, ↑ LC3Ⅱ/LC3Ⅰ, ↑ Atg7, ↑ Beclin-2, ↑ p-Beclin-2, ↓ p62, ↓ β-catenin(NE), -β-catenin(CE) in cells | [52] | ||
| RIRI | SD rats; after resection of the right kidney, the left kidney was ligated for 25 min by non-traumatic aneurysm clipping | In vivo | Tan IIA; 25 mg/kg.d, i.p. for 10 days | ↓ MPO, ↓ TNF-α, ↓ IL-6, ↓ MIF, ↓ cleaved caspase-3, ↑ Bcl-2, ↓ p-p38 MAPK in renal tissue | [54] | |
| HK-2 cells; the cells were cultured in a hypoxic incubator (94%N2, 5%CO2, 1%O2) for 24 h and then cultured in a carbon dioxide incubator for 6 h. | In vitro | Tan IIA; 5, 10, 20 μg/mL for 24 h | ↓ ROS, ↓ Bcl-2, ↑ Bax, ↑ Cleaved Caspase-3 in cells | [55] | ||
| SD rats; high-fat diet (HFD) feed for 8 weeks + the left renal artery was clamped for 30 min after right nephrectomy. | Tan IIA; 5 mg/kg.d, 10 mg/kg.d, and 20 mg/kg.d for 2 weeks | ↓ TNF-α, ↓ IL-1β, ↑ SOD, ↓ MDA, ↓ caspase-9/3, ↓ cleaved caspase-9/3, ↓ Bax, ↑ Bcl-2, ↓ PARP, ↓ Cyt-c, ↓ ROS, ↑ PI3K, ↑ Bad, ↑ Akt, ↑ p-AKT, ↑ p-Bad, ↑ PCG-1α, ↑ Nrf1, ↑ Tfam, ↓ Mfn1, ↓ Mfn2, ↑ Drp1 in renal tissue | [56] | |||
| Drug-induced kidney injury | Cisplatin-induced renal injury | C57BL/6 mice; a single dose of cisplatin (20 mg/kg) was intraperitoneally injected. | In vivo | Tan IIA; 12.5 mg/kg.d and 25 mg/kg.d, i.p. for 3 d | ↓ P105/p50, ↓ IKKβ, ↓ L-6, ↓ IL-1β, ↓ TGF-β, ↑ PXR, ↑ RXRα, ↓ pp65/p65, ↑ Cyp3a11 in renal tissue↓ TNF-α, ↓ IL-6 in serum | [57] |
| Vancomycin-induced renal injury | SD rats; 200 mg/kg vancomycin by single intraperitoneal injection | In vivo | STS; 15, 30, 60 µg/kg.d, i.p. for 10 d | ↑ SOD, ↑ GSH-Px, ↓ MDA, ↓ NO in renal tissue | [59] | |
| APAP-induced renal injury | C57BL/6J Nrf2+/− mice; 200 mg/kg APAP for 3 h HK-2 cells; APAP (1 mM) for 24, 48, 72 h | In vitro and In vivo | Tan IIA; 10 mg/kg.d and 30 mg/kg.d, p.o. for 1 week Tan IIA; 2.5, 10 μM for 24, 48, 72 h | ↑ Nrf2, ↑ MRP2, ↑ MRP4 in renal tissue ↑ Nrf2, ↑ MRP2, ↑ MRP, ↑ p-Nrf2 (in cell nucleus), ↑ p-Nrf2 (in cytoplasm), ↑ Nrf2(in cell nucleus) in cells | [61] | |
| CIN | SD rats; indomethacin (10 mg/kg) was given after tail vein injection of 50 mg/kg sodium pentobarbital, followed by tail vein injection of ioversol (3 g/kg organic iodine). HK-2 cells; the cells were incubated with 500 µmol/L H2O2 for 5 min. | In vitro and In vivo | Tan IIA; 25 mg/kg i.h. single use Tan IIA; 40 µg/mL for 1 h | ↓ MDA, ↓ 8-OHdg, ↑ Nrf2, ↑ HO-1 in renal tissue; ROS↓, ↑ Nrf2, ↑ HO-1, ↑ ARE in HK2 cells | [64] | |
| HSPN | C57BL/6 mice; BSA (4 mL/kg/every other day for 8 weeks) was administrated intragastrically, LPS (0.025%LPS0.2 mL, once at 6, 8, 10 and 12 weeks) was injected into the caudal vein, and CCL4 (0.3 mL castor oil + 0.1 mLCCL4, once a week) was injected subcutaneously. The IgA nephropathy model was established at the same time during the modeling period, and the blood fever model was combined into the anaphylactoid nephritis model. The room temperature during the modeling period was 30 °C, and 25% dry ginger water (10 mL/kg, once every other day) was given from the 9th week, and the modeling was finished at the 12th week. | In vivo | Tan IIA; 25 μmol/L 200 μL, i.p. for 4 weeks | ↑ AOPP, ↑ ERK, ↑ p-ERK, ↓ Cosmc in renal tissue | [66] | |
| Hypertensive renal damage | RRAECs; AngⅡ was treated with 5 × 10−7 mol for 24 h. | In vitro | Isoflavone 1.5 mg/L, Tan IIA 3 mg/L, isoflavone 3 mg/L+Tan IIA 3 mg/L for 24 h | ↓ ROS, ↑ eNOS in cells | [67] | |
| RRAECs; AngⅡ was treated with 5 × 10−7 mol for 24 h. | In vitro | Isoflavone 3 mg/L-Tan IIA 3 mg/L pretreated for 1 h | ↓ miRNA -200c -3p, ↑ ZEB2 in cells | [68] | ||
| NS | SD rats; single tail vein injection of doxorubicin (6.5 mg/kg) | In vivo | STS; 3.5 and 7 mg/kg.d, i.p. for 2 weeks | ↓ TGF-β1, ↓ PAI-1 in renal tissue | [71] | |
| SD rats; single tail vein injection of doxorubicin (7 mg/kg) | In vivo | Tan IIA; 0.02, 0.04 g/kg/d; p.o. for 4 weeks | ↓ TGF-β1, ↑ nephrin in renal tissue | [72] | ||
| AKI induced by sepsis | C57BL/6 mice; LPS (10 mg/kg) was intraperitoneally injected. 10 mg/kg LPS was injected and stimulated for 24 h. HK-2 cell; LPS (10 μg/mL) for 24 h | In vitro and In vivo | Tan IIA; 10 mg/kg; i.p. in advance for 15 min Tan IIA; 10 mg/L pretreated for 1 h | ↓ Cleaved-caspase3, ↓ RIP3, ↓ p18-FUNDC in renal tissue; ↓ Cleaved-caspase3, ↓ RIP3, ↓ p18-FUNDC in cells | [73] | |
3. Safety and Side Effects and Drug Interactions
4. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Inagi, R. Organelle Stress and Metabolic Derangement in Kidney Disease. Int. J. Mol. Sci. 2022, 23, 1723. [Google Scholar] [CrossRef] [PubMed]
- Capolongo, G.; Capasso, G.; Viggiano, D. A Shared Nephroprotective Mechanism for Renin-Angiotensin-System Inhibitors, Sodium-Glucose Co-Transporter 2 Inhibitors, and Vasopressin Receptor Antagonists: Immunology Meets Hemodynamics. Int. J. Mol. Sci. 2022, 23, 3915. [Google Scholar] [CrossRef] [PubMed]
- Perkovic, V.; Jardine, M.J.; Neal, B.; Bompoint, S.; Heerspink, H.; Charytan, D.M.; Edwards, R.; Agarwal, R.; Bakris, G.; Bull, S.; et al. Mahaffey. Canagliflozin and Renal Outcomes in Type 2 Diabetes and Nephropathy. N. Engl. J. Med. 2019, 380, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.; Tarnow, L.; Rossing, P.; Hansen, B.V.; Parving, H.H. Renoprotective effects of angiotensin II receptor blockade in type 1 diabetic patients with diabetic nephropathy. Kidney Int. 2000, 57, 601–606. [Google Scholar] [CrossRef]
- Abecassis, M.; Bartlett, S.T.; Collins, A.J.; Davis, C.L.; Delmonico, F.L.; Friedewald, J.J.; Hays, R.; Howard, A.; Jones, E.; Leichtman, A.B.; et al. Kidney transplantation as primary therapy for end-stage renal disease: A National Kidney Foundation/Kidney Disease Outcomes Quality Initiative (NKF/KDOQITM) conference. Clin. J. Am. Soc. Nephrol. 2008, 3, 471–480. [Google Scholar] [CrossRef]
- Wolfe, R.A.; Ashby, V.B.; Milford, E.L.; Ojo, A.O.; Ettenger, R.E.; Agodoa, L.Y.; Held, P.J.; Port, F.K. Comparison of mortality in all patients on dialysis, patients on dialysis awaiting transplantation, and recipients of a first cadaveric transplant. N. Engl. J. Med. 1999, 341, 1725–1730. [Google Scholar] [CrossRef]
- Chen, D.Q.; Hu, H.H.; Wang, Y.N.; Feng, Y.L.; Cao, G.; Zhao, Y.Y. Natural products for the prevention and treatment of kidney disease. Phytomedicine 2018, 50, 50–60. [Google Scholar] [CrossRef]
- Jiang, Z.; Gao, W.; Huang, L. Tanshinones, Critical Pharmacological Components in Salvia miltiorrhiza. Front. Pharmacol. 2019, 10, 202. [Google Scholar] [CrossRef]
- Feng, J.; Li, S.; Chen, H. Tanshinone IIA ameliorates apoptosis of cardiomyocytes induced by endoplasmic reticulum stress. Exp. Biol. Med. 2016, 241, 2042–2048. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, P.; Ye, M.; Kim, S.H.; Jiang, C.; Lü, J. Tanshinones: Sources, pharmacokinetics and anti-cancer activities. Int. J. Mol. Sci. 2012, 13, 13621–13666. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, H. Anti-Inflammatory and Immunomodulatory Mechanism of Tanshinone IIA for Atherosclerosis. Evid. Based Complement Alternat. Med 2014, 2014, 267976. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Liu, Z.; Li, H.; Little, P.J.; Liu, P.; Xu, S. Cardiovascular actions and therapeutic potential of tanshinone IIA. Atherosclerosis 2012, 220, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Xu, H.; Huang, L. Tanshinone IIA: A Promising Natural Cardioprotective Agent. Evid. Based Complement Alternat. Med. 2012, 2012, 716459. [Google Scholar] [CrossRef]
- Wang, P.; Zhou, S.; Xu, L.; Lu, Y.; Yuan, X.; Zhang, H.; Li, R.; Fang, J.; Liu, P. Hydrogen peroxide-mediated oxidative stress and collagen synthesis in cardiac fibroblasts: Blockade by tanshinone IIA. J. Ethnopharmacol. 2013, 145, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, Y.; Xing, C.; Zhang, M. Tanshinone IIA represses inflammatory response and reduces radiculopathic pain by inhibiting IRAK-1 and NF-kappaB/p38/JNK signaling. Int. Immunopharmacol. 2015, 28, 382–389. [Google Scholar] [CrossRef]
- Bi, Z.; Wang, Y.; Zhang, W. A comprehensive review of tanshinone IIA and its derivatives in fibrosis treatment. Biomed. Pharmacother. 2021, 137, 111404. [Google Scholar] [CrossRef]
- Zhang, H.S.; Chen, X.; Wu, T.; Zhang, F. Tanshinone II A inhibits tat-induced HIV-1 transactivation through redox-regulated AMPK/Nampt pathway. J. Cell. Physiol. 2014, 229, 1193–1201. [Google Scholar] [CrossRef]
- Subedi, L.; Gaire, B. Tanshinone IIA: A phytochemical as a promising drug candidate for neurodegenerative diseases. Pharmacol. Res. 2021, 169, 105661. [Google Scholar] [CrossRef]
- Maione, F.; De Feo, V.; Caiazzo, E.; De Martino, L.; Cicala, C.; Mascolo, N. Tanshinone IIA, a major component of Salvia milthorriza Bunge, inhibits platelet activation via Erk-2 signaling pathway. J. Ethnopharmacol. 2014, 155, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.J.; Dong, B.; Yang, W.N.; Su, S.B.; Zhang, H. Preventive and therapeutic role of Tanshinone ⅡA in hepatology. Biomed. Pharmacother. 2019, 112, 108676. [Google Scholar] [CrossRef]
- Zhang, W.; He, H.; Liu, J.; Wang, J.; Zhang, S.; Zhang, S.; Wu, Z. Pharmacokinetics and atherosclerotic lesions targeting effects of tanshinone IIA discoidal and spherical biomimetic high density lipoproteins. Biomaterials 2013, 34, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.Y.; Zhao, W.R.; Zhang, J.; Chen, X.L.; Tang, J.Y. Tang. Sodium tanshinone IIA sulfonate: A review of pharmacological activity and pharmacokinetics. Biomed. Pharmacother. 2019, 118, 109362. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhou, L. The Signaling of Cellular Senescence in Diabetic Nephropathy. Oxid. Med. Cell. Longev. 2019, 2019, 7495629. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.C.; Wu, H.; Kirita, Y.; Uchimura, K.; Ledru, N.; Rennke, H.G.; Welling, P.A.; Waikar, S.S.; Humphreys, B.D. The single-cell transcriptomic landscape of early human diabetic nephropathy. Proc. Natl. Acad. Sci. USA 2019, 116, 19619–19625. [Google Scholar] [CrossRef]
- Tesch, G.H. Diabetic nephropathy-is this an immune disorder? Clin. Sci. 2017, 131, 2183–2199. [Google Scholar] [CrossRef] [PubMed]
- Flyvbjerg, A. The role of the complement system in diabetic nephropathy. Nat. Rev. Nephrol. 2017, 13, 311–318. [Google Scholar] [CrossRef]
- Rossing, P.; Persson, F.; Frimodt-Moller, M. Prognosis and treatment of diabetic nephropathy: Recent advances and perspectives. Nephrol. Ther. 2018, 14 (Suppl. 1), S31–S37. [Google Scholar] [CrossRef]
- Ruiz-Ortega, M.; Rodrigues-Diez, R.R.; Lavoz, C.; Rayego-Mateos, S. Special Issue “Diabetic Nephropathy: Diagnosis, Prevention and Treatment”. J. Clin. Med. 2020, 9, 813. [Google Scholar] [CrossRef]
- Chen, X.; Wu, R.; Kong, Y.; Yang, Y.; Gao, Y.; Sun, D.; Liu, Q.; Dai, D.; Lu, Z.; Wang, N.; et al. Tanshinone IIA attenuates renal damage in STZ-induced diabetic rats via inhibiting oxidative stress and inflammation. Oncotarget 2017, 8, 31915–31922. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, X.; Li, C.; Lin, Y.; Meng, Y.; Tang, S. Role of the TGFbeta/p65 pathway in tanshinone ⅡA-treated HBZY-1 cells. Mol. Med. Rep. 2014, 10, 2471–2476. [Google Scholar] [CrossRef]
- Xu, S.; He, L.; Ding, K.; Zhang, L.; Xu, X.; Wang, S.; Qian, X. Tanshinone IIA Ameliorates Streptozotocin-Induced Diabetic Nephropathy, Partly by Attenuating PERK Pathway-Induced Fibrosis. Drug Des. Devel. Ther. 2020, 14, 5773–5782. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Deng, X.; Zhuang, W.; Li, Y.; Xue, H.; Lv, X.; Zhu, S. Tanshinone IIA down-regulates -transforming growth factor beta 1 to relieve renal tubular epithelial cell inflammation and pyroptosis caused by high glucose. Bioengineered 2022, 13, 12224–12236. [Google Scholar] [CrossRef] [PubMed]
- Yong, C.; Cairong, L.; Fei, C. Effect of sodium tanshinone IIA sulfonic acid on SIRT1 and Fox01 in kidney of diabetic rats. Pharmacol. Clin. Chin. Mater. Med. 2015, 31, 47–50. (In Chinese) [Google Scholar]
- Wei, C.; Ji-Xiong, X.; Ling-Yan, Z.; Jun-Ren, X.; Gang, L. Effect of Tanshinone ⅡA on RAGE Expression and Oxidative Stress Status in Human Mesangial Cells Induced by AGE. Chin. Gen. Pract. 2014, 17, 3585–3589. (In Chinese) [Google Scholar]
- Kalantar-Zadeh, K.; Jafar, T.H.; Nitsch, D.; Neuen, B.L.; Perkovic, V. Chronic kidney disease. Lancet 2021, 398, 786–802. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, A.; Pires, M.; Oliveira, P. Pathophysiological Mechanisms of Renal Fibrosis: A Review of Animal Models and Therapeutic Strategies. In Vivo 2017, 31, 1–22. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.G.; Yang, X.K.; Li, M.; Liu, S.J.; Zhang, T.Y.; Wang, Y.Q. Tanshinone IIA suppresses fibrosis induced by high glucose conditions in HK-2 cells via inhibition of extracellular matrix deposition, reduction of oxidative stress, and inhibition of epithelial to mesenchymal transition. Trop. J. Pharm. Res. 2020, 19, 739–744. [Google Scholar] [CrossRef]
- Cao, L.; Huang, B.; Fu, X.; Yang, J.; Lin, Y.; Lin, F. Effects of tanshinone IIA on the regulation of renal proximal tubular fibrosis. Mol. Med. Rep. 2017, 15, 4247–4252. [Google Scholar] [CrossRef]
- Zeng, J.; Bao, X. Tanshinone IIA attenuates high glucose-induced epithelial-to-mesenchymal transition in HK-2 cells through VDR/Wnt/beta-catenin signaling pathway. Folia Histochem. Cytobiol. 2021, 59, 259–270. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, F. Tanshinone IIA improves diabetes-induced renal fibrosis by regulating the miR-34-5p/Notch1 axis. Food Sci. Nutr. 2022, 10, 4019–4040. [Google Scholar] [CrossRef]
- Wang, D.T.; Huang, R.H.; Cheng, X.; Zhang, Z.H.; Yang, Y.J.; Lin, X. Tanshinone IIA attenuates renal fibrosis and inflammation via altering expression of TGF-beta/Smad and NF-kappaB signaling pathway in 5/6 nephrectomized rats. Int. Immunopharmacol. 2015, 26, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Shao, Q.; Jin, B.; Gong, R.; Zhang, M.; Xu, B. Tanshinone IIA Attenuates Renal Fibrosis after Acute Kidney Injury in a Mouse Model through Inhibition of Fibrocytes Recruitment. Biomed. Res. Int. 2015, 2015, 867140. [Google Scholar] [CrossRef]
- Tang, J.; Zhan, C.; Zhou, J. Effects of tanshinone IIA on transforming growth factor beta1-Smads signal pathway in renal interstitial fibroblasts of rats. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2008, 28, 539–542. (In Chinese) [Google Scholar] [CrossRef] [PubMed]
- Ding-Hao, F.; Jun, L.; Jing-Ya, X.; Li-Zhi, Z. Effect of Tanshinone ⅡA Combined with DAPT on Notch/Jagged Pathway Proteins in Renal Tissue of UUO Rat Models. Chin. J. Integr. Tradit. West. Med. 2019, 39, 826–831. [Google Scholar]
- Quan, W.; Yan-Min, Z.; Wei-Dong, C.; Tao-Tao, H.; Huan-Lan, W.; Hong, L. Effect of tanshinone ⅡA on the renal pathological damage and renal interstitial fibrosis of chronic renal failure rats. Chin. J. Clin. Pharmacol. 2019, 35, 2609–2612. (In Chinese) [Google Scholar]
- Pan, J.; Shi, M.; Li, L.; Liu, J.; Guo, F.; Feng, Y.; Ma, L.; Fu, P. Pterostilbene, a bioactive component of blueberries, alleviates renal fibrosis in a severe mouse model of hyperuricemic nephropathy. Biomed. Pharmacother. 2019, 109, 1802–1808. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Fang, Y.; Yu, X.; Guo, L.; Zhang, X.; Xia, D. The flavonoid-rich fraction from rhizomes of Smilax glabra Roxb. ameliorates renal oxidative stress and inflammation in uric acid nephropathy rats through promoting uric acid excretion. Biomed. Pharmacother. 2019, 111, 162–168. [Google Scholar] [CrossRef]
- Wu, X.; Liu, L.; Xie, H.; Liao, J.; Zhou, X.; Wan, J.; Yu, K.; Li, J.; Zhang, Y. Tanshinone IIA prevents uric acid nephropathy in rats through NF-kappaB inhibition. Planta Med. 2012, 78, 866–873. [Google Scholar]
- Zhang, X.W.; Zhou, M.; An, L.; Zhang, P.; Li, P.; Chen, J. Lipophilic Extract and Tanshinone IIA Derived from Salvia miltiorrhiza Attenuate Uric Acid Nephropathy through Suppressing Oxidative Stress-Activated MAPK Pathways. Am. J. Chin. Med. 2020, 48, 1455–1473. [Google Scholar] [CrossRef]
- Capitanio, U.; Bensalah, K.; Bex, A.; Boorjian, S.A.; Bray, F.; Coleman, J.; Gore, J.L.; Sun, M.; Wood, C.; Russo, P. Epidemiology of Renal Cell Carcinoma. Eur. Urol. 2019, 75, 74–84. [Google Scholar] [CrossRef]
- Wei, X.; Zhou, L.; Hu, L.; Huang, Y. Tanshinone IIA arrests cell cycle and induces apoptosis in 786-O human renal cell carcinoma cells. Oncol. Lett. 2012, 3, 1144–1148. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.Y.; Jung, Y.Y.; Yang, M.H.; Chinnathambi, A.; Govindasamy, C.; Narula, A.S.; Namjoshi, O.A.; Blough, B.E.; Ahn, K.S. Tanshinone IIA exerts autophagic cell death through down-regulation of beta-catenin in renal cell carcinoma cells. Biochimie 2022, 200, 119–130. [Google Scholar] [CrossRef]
- Salvadori, M.; Rosso, G.; Bertoni, E. Update on ischemia-reperfusion injury in kidney transplantation: Pathogenesis and treatment. World J. Transplant 2015, 5, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.M.; Ding, G.H.; Huang, J.; Xiong, Y. Tanshinone IIA pretreatment attenuates ischemia/reperfusion-induced renal injury. Exp. Ther. Med. 2016, 12, 2741–2746. [Google Scholar] [CrossRef] [PubMed]
- Ziyi, Y.; Gang, C.; Yunxiao, Z.; Chaoyu, X.; Han, C.; Shuang, W.; Yi, L. Effects of Tanshinone ⅡA on hypoxia/reperfusion human kidney-2 cells and their mechanism. J. Chongqing Med. Univ. 2018, 43, 1427–1432. (In Chinese) [Google Scholar]
- Tai, H.; Cui, X.Z.; He, J.; Lan, Z.M.; Li, S.M.; Li, L.B.; Yao, S.C.; Jiang, X.L.; Meng, X.S.; Kuang, J.S. Renoprotective effect of Tanshinone IIA against kidney injury induced by ischemia-reperfusion in obese rats. Aging 2022, 14, 8302–8320. [Google Scholar] [CrossRef]
- Dou, J.Y.; Zhang, M.; Cen, H.; Chen, Y.Q.; Wu, Y.F.; Lu, F.; Zhou, J.; Liu, X.S.; Gu, Y.Y. Salvia miltiorrhiza Bunge (Danshen) and Bioactive Compound Tanshinone IIA Alleviates Cisplatin-Induced Acute Kidney Injury Through Regulating PXR/NF-kappaB Signaling. Front. Pharmacol. 2022, 13, 860383. [Google Scholar]
- Iwamoto, T.; Kagawa, Y.; Kojima, M. Clinical efficacy of therapeutic drug monitoring in patients receiving vancomycin. Biol. Pharm. Bull. 2003, 26, 876–879. [Google Scholar] [CrossRef]
- Jiaxi, X.; Huajun, Z.; Xiaoyu, C. Study on the Protective Effect of Tanshinone ⅡA Sulfonate Injection on Vancomycin-induced Renal Injury Model Rats and Its Mechanism. China Pharm. 2016, 27, 3081–3084. (In Chinese) [Google Scholar]
- Chen, Y.G.; Lin, C.L.; Dai, M.S.; Chang, P.Y.; Chen, J.H.; Huang, T.C.; Wu, Y.Y.; Kao, C.H. Risk of Acute Kidney Injury and Long-Term Outcome in Patients with Acetaminophen Intoxication: A Nationwide Population-Based Retrospective Cohort Study. Medicine 2015, 94, e2040. [Google Scholar] [CrossRef]
- Zhang, X.; Long, F.; Li, R.; Yang, Y.; Wang, T.; He, Q.; Xu, M.; Wang, L.; Jiang, X. Tanshinone IIA prevents acetaminophen-induced nephrotoxicity through the activation of the Nrf2-Mrp2/4 pathway in mice. Environ. Toxicol. 2022, 37, 1618–1628. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Cheng, D.; Yin, J.; Wu, R.; Zhang, G.; Zhao, Q.; Wang, N.; Wang, F.; Liang, M. Antithrombin III Protects Against Contrast-Induced Nephropathy. EBioMedicine 2017, 17, 101–107. [Google Scholar] [CrossRef]
- Tan, X.; Zheng, X.; Huang, Z.; Lin, J.; Xie, C.; Lin, Y. Involvement of S100A8/A9-TLR4-NLRP3 Inflammasome Pathway in Contrast-Induced Acute Kidney Injury. Cell Physiol. Biochem. 2017, 43, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Zhao, Q.; Jian, G.; Cheng, D.; Wang, N.; Zhang, G.; Wang, F. Tanshinone IIA Attenuates Contrast-Induced Nephropathy via Nrf2 Activation in Rats. Cell Physiol. Biochem. 2018, 46, 2616–2623. [Google Scholar] [CrossRef]
- Chen, J.Y.; Mao, J. Henoch-Schonlein purpura nephritis in children: Incidence, pathogenesis and management. World J. Pediatr. 2015, 11, 29–34. [Google Scholar] [CrossRef]
- Yun, Y.; Xixing, M.; Dahu, W.; Huanhuan, Z.; Yaohui, M.; Cuimin, R.; Qiang, L. Influence of tanshinone ⅡA on expressins of Cosmc and AOPP in kidney tissue of allergic purpura nephritis mice and its mechanism. J. Jilin Univ. 2019, 45, 867–871+987. (In Chinese) [Google Scholar]
- Yao, L.; Yue-Hua, J.; Wei, L. Mechanism study of Calycosin-Tanshinone ⅡA on regulating endothelial cell function involved in hypertensive renal damage. China J. Tradit. Chin. Med. Pharm. 2020, 35, 4598–4601. (In Chinese) [Google Scholar]
- Yao, L.; Cong, H.; Wei, L. Effect of active ingredients of Astragalus-Danshen pair on proliferation and migration of rat renal artery endothelial cells through regulation of miRNA-200c -3p /ZEB2. Lishizhen Med. Mater. Med. Res. 2022, 33, 516–520. (In Chinese) [Google Scholar]
- Wang, Y.; Wang, Y.; Tay, Y.; Harris, D. Progressive adriamycin nephropathy in mice: Sequence of histologic and immunohistochemical events. Kidney Int. 2000, 58, 1797–1804. [Google Scholar] [CrossRef]
- Bertani, T.; Poggi, A.; Pozzoni, R.; Delaini, F.; Sacchi, G.; Thoua, Y.; Mecca, G.; Remuzzi, G.; Donati, M.B. Donati. Adriamycin-induced nephrotic syndrome in rats: Sequence of pathologic events. Lab. Investig. 1982, 46, 16–23. [Google Scholar]
- Bo, L.; Maobo, Y.; Hongbin, X. Research on Intervention Mechanism of Tanshinone Type IIA Sulfonate of Adriamycin Nephrosis Rats Kidney Injury. Chin. Arch. Tradit. Chin. Med. 2015, 33, 2470–2472+26. (In Chinese) [Google Scholar]
- Jian, W.; Rui, H. Effects of Tanshinone ⅡA on Renal Nephrin and Transforming Growth Factor-β1 of Adriamycin Nephritic Rats. Chin. J. Exp. Tradit. Med. Formulae 2011, 17, 245–251. (In Chinese) [Google Scholar]
- Shu, Z.; Baolin, S.; Liangliang, W.; Shuifu, T.; Gangyi, C. Tanshinone ⅡA alleviates lipopolysaccharide-induced renal tubular epithelial cell apoptosis by inhibiting RIP3/FUNDC1 signaling pathway. J. South. Med. Univ. 2022, 42, 1852–1857. (In Chinese) [Google Scholar]
- Gu, Y.; Liang, Z.; Wang, H.; Jin, J.; Zhang, S.; Xue, S.; Chen, J.; He, H.; Duan, K.; Wang, J.; et al. Tanshinone IIA protects H9c2 cells from oxidative stress-induced cell death via microRNA-133 upregulation and Akt activation. Exp. Ther. Med. 2016, 12, 1147–1152. [Google Scholar] [CrossRef]
- Yang, L.J.; Jeng, C.J.; Kung, H.N.; Chang, C.C.; Wang, A.G.; Chau, G.Y.; Don, M.J.; Chau, Y.P. Tanshinone IIA isolated from Salvia miltiorrhiza elicits the cell death of human endothelial cells. J. Biomed. Sci. 2005, 12, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, C.; Wu, Q.; Zheng, K.; Chen, J.; Lan, Y.; Qin, Y.; Mei, W.; Wang, B. Evaluation of Tanshinone IIA Developmental Toxicity in Zebrafish Embryos. Molecules 2017, 22, 660. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Wang, L.; Zhao, X.; Guo, L.; Lin, Q.; Wang, X.; Dai, X.; Shang, H.; Zhang, M.; Hinek, A. Efficacy of Sodium Tanshinone IIA Sulfonate in Patients with Non-ST Elevation Acute Coronary Syndrome Undergoing Percutaneous Coronary Intervention: Results from a Multicentre, Controlled, Randomized Trial. Cardiovasc. Drugs Ther. 2021, 35, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.-H. Anaphylactic shock induced by sodium Tanshinone IIA sulfonate injection. Nei Mong. J. Tradit. Chin. Med. 2008, 3, 28. (In Chinese) [Google Scholar]
- Jiang, S.-X.; Zheng, L.-Y. Anaphylactic shock induced by sodium Tanshinone IIA sulfonate injection. Advers. Drug React. J. 2007, 1, 61. (In Chinese) [Google Scholar]
- Shi, B.; Li, Q.; Feng, Y.; Dai, X.; Zhao, R.; Zhao, Y.; Jia, P.; Wang, S.; Yu, J.; Liao, S.; et al. Pharmacokinetics of 13 active components in a rat model of middle cerebral artery occlusion after intravenous injection of Radix Salviae miltiorrhizae-Lignum dalbergiae odoriferae prescription. J. Sep. Sci. 2020, 43, 531–546. [Google Scholar] [CrossRef]
- Li, K.; Liu, W.; Zhao, Q.; Wu, C.; Fan, C.; Lai, H.; Li, S. Combination of tanshinone IIA and doxorubicin possesses synergism and attenuation effects on doxorubicin in the treatment of breast cancer. Phytother. Res. 2019, 33, 1658–1669. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, T.; Yang, Y.; Li, R.; Chen, Y.; Li, R.; Jiang, X.; Wang, L. Tanshinone IIA attenuates acetaminophen-induced hepatotoxicity through HOTAIR-Nrf2-MRP2/4 signaling pathway. Biomed. Pharmacother. 2020, 130, 110547. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, X.; Cai, Z.; Lee, F.S. Effect of tanshinone IIA on the noncovalent interaction between warfarin and human serum albumin studied by electrospray ionization mass spectrometry. J. Am. Soc. Mass Spectrom. 2008, 19, 1568–1575. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jia, Q.; Zhou, Y.; Jiang, X.; Song, L.; Wu, Y.; Wang, A.; Chen, W.; Wang, S.; Lu, Y. Tanshinone IIA attenuates the stemness of breast cancer cells via targeting the miR-125b/STARD13 axis. Exp. Hematol. Oncol. 2022, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shu, H.; Su, W.; Li, B.; Zhang, H.; Li, L.; Lin, C.; Yi, W.; Zhan, X.Y.; Chen, C.; et al. Tanshinone IIA Has a Potential Therapeutic Effect on Kawasaki Disease and Suppresses Megakaryocytes in Rabbits with Immune Vasculitis. Front. Cardiovasc. Med. 2022, 9, 873851. [Google Scholar] [CrossRef]
- Gong, Y.; Liu, Y.C.; Ding, X.L.; Fu, Y.; Cui, L.J.; Yan, Y.P. Tanshinone IIA Ameliorates CNS Autoimmunity by Promoting the Differentiation of Regulatory T Cells. Neurotherapeutics 2020, 17, 690–703. [Google Scholar] [CrossRef]
- Ni, H.; Ruan, G.; Sun, C.; Yang, X.; Miao, Z.; Li, J.; Chen, Y.; Qin, H.; Liu, Y.; Zheng, L.; et al. Tanshinone IIA inhibits gastric cancer cell stemness through inducing ferroptosis. Environ. Toxicol. 2022, 37, 192–200. [Google Scholar] [CrossRef]
- Hua, G.; Yang, G. Clinical observation of sodium tanshinone ⅡA sulfonate injection in the treatment of chronic renal failure. Shandong Med. J. 2011, 51, 80–81. (In Chinese) [Google Scholar]
- Zhi, W.; Chun-Hua, L.; Dong-Mei, L. Efficacy of irbesartan tablets combined with sodium tanshinone ⅡA sulfonate in the treatment of early diabetic nephropathy. Tianjin Med. J. 2010, 38, 901–902. (In Chinese) [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Feng, H.; Peng, C.; Zhang, Z.; Yuan, Q.; Gao, H.; Tang, S.; Xie, C. Renoprotective Effects of Tanshinone IIA: A Literature Review. Molecules 2023, 28, 1990. https://doi.org/10.3390/molecules28041990
Chen Z, Feng H, Peng C, Zhang Z, Yuan Q, Gao H, Tang S, Xie C. Renoprotective Effects of Tanshinone IIA: A Literature Review. Molecules. 2023; 28(4):1990. https://doi.org/10.3390/molecules28041990
Chicago/Turabian StyleChen, Zhengtao, Haoyue Feng, Chuan Peng, Zehua Zhang, Qianghua Yuan, Hong Gao, Shiyun Tang, and Chunguang Xie. 2023. "Renoprotective Effects of Tanshinone IIA: A Literature Review" Molecules 28, no. 4: 1990. https://doi.org/10.3390/molecules28041990
APA StyleChen, Z., Feng, H., Peng, C., Zhang, Z., Yuan, Q., Gao, H., Tang, S., & Xie, C. (2023). Renoprotective Effects of Tanshinone IIA: A Literature Review. Molecules, 28(4), 1990. https://doi.org/10.3390/molecules28041990