
Characterization of the Glucan-Branching Enzyme GlgB Gene from Swine Intestinal Bacteria



Abstract
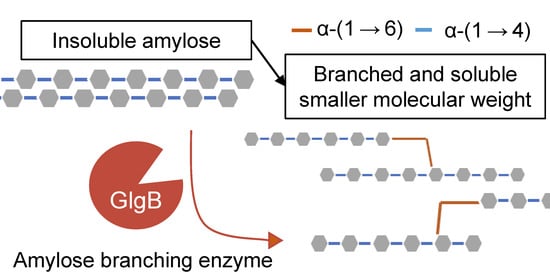
1. Introduction
2. Results
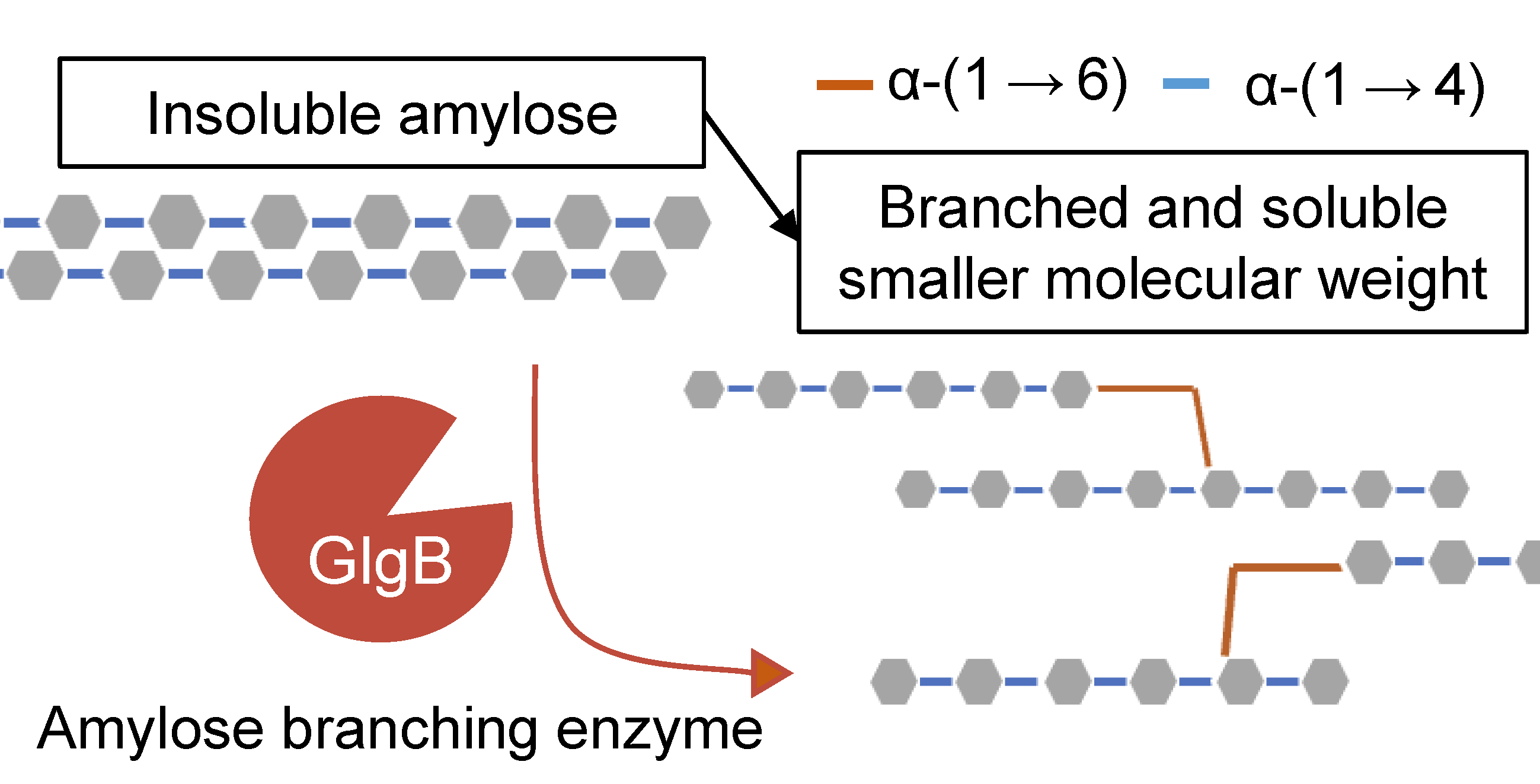
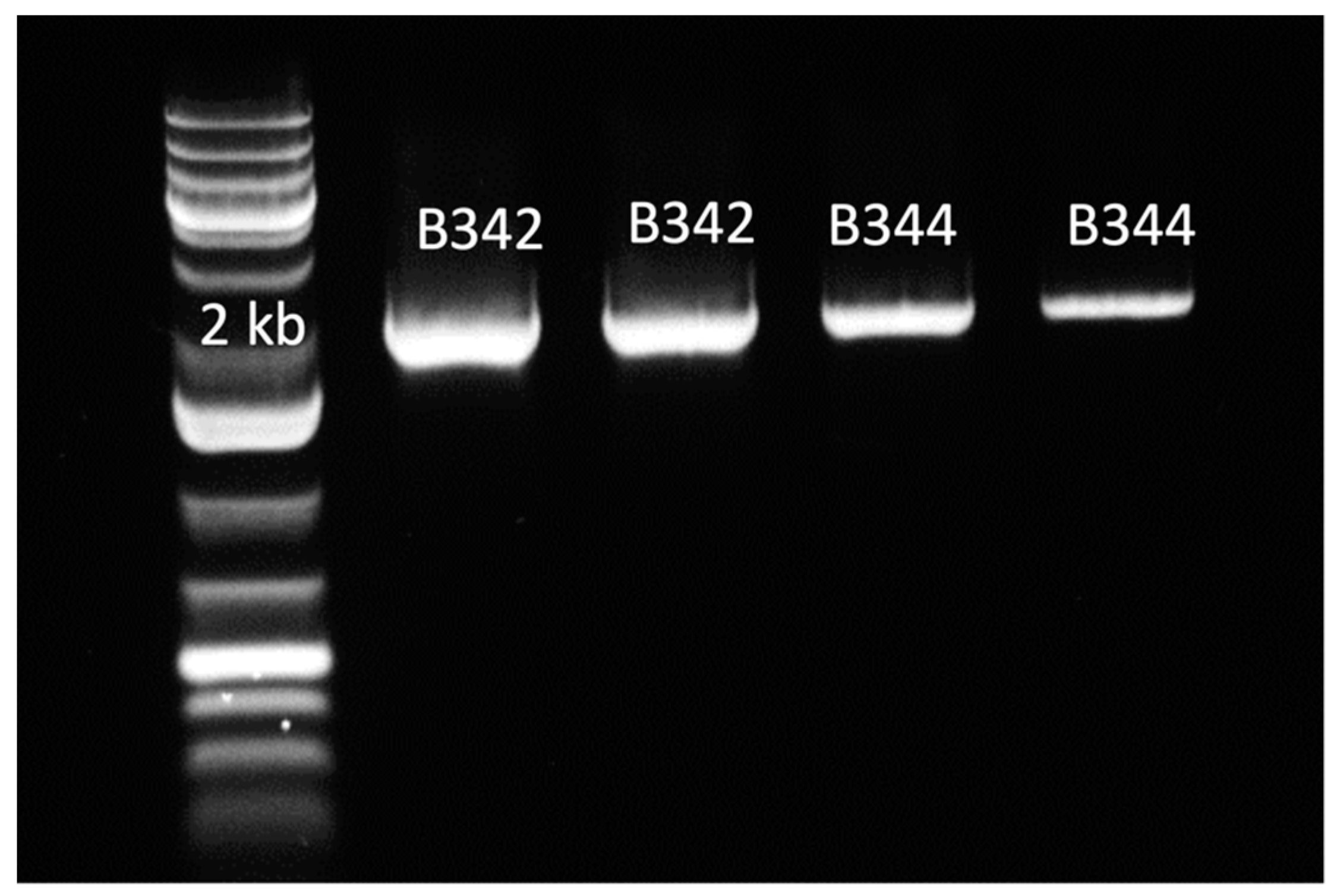
2.1. Cloning of Glgb from Swine Intestinal Bacteria in E. coli and Protein Purification
2.2. Quantitative Determination of Iodine-Binding Amylose
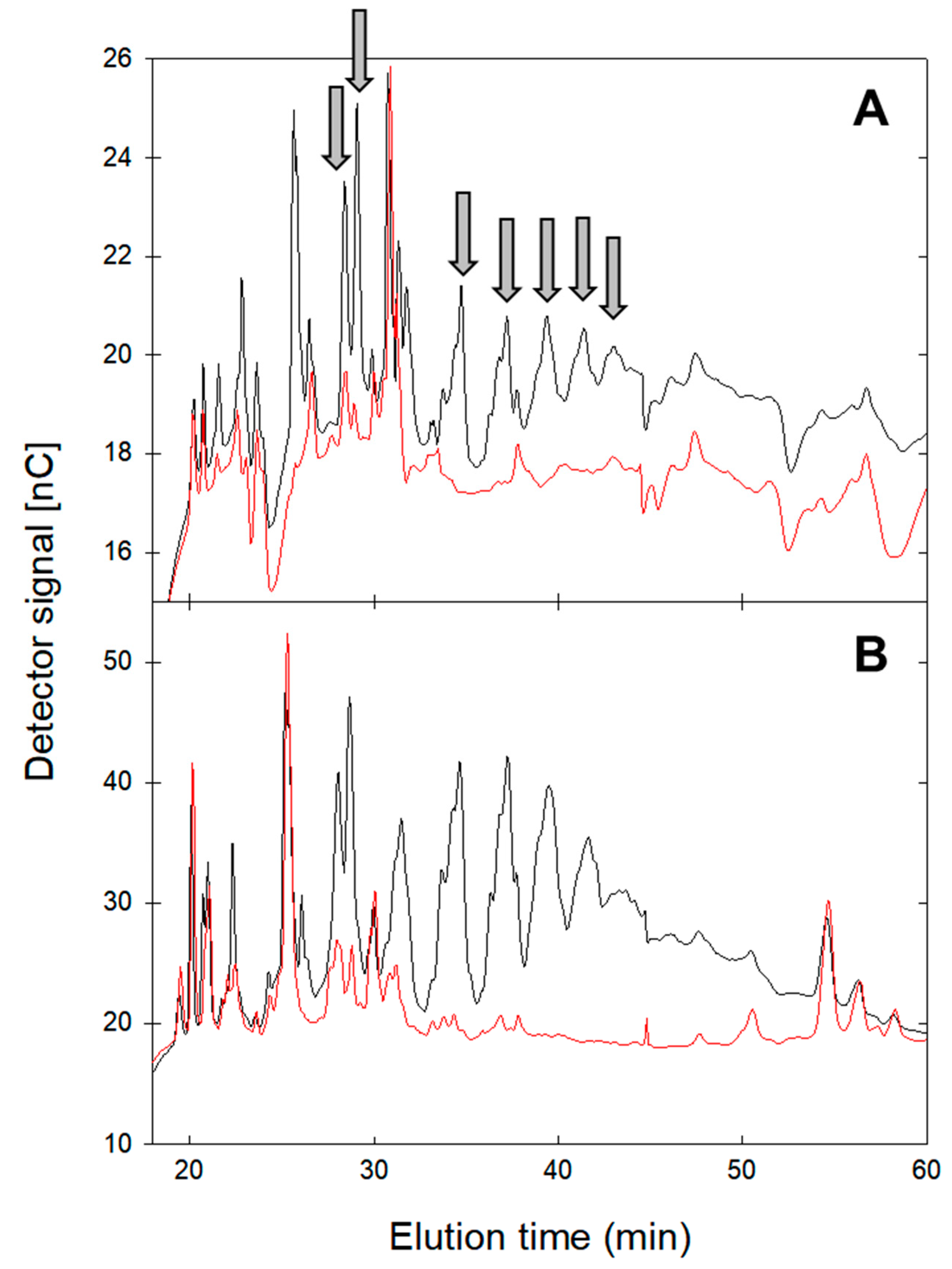
2.3. Oligosaccharide Profile after Amylase Hydrolysis
2.4. Molecular Size Distribution of GlgB-Treated Starch
2.5. Quantitative Determination of Reducing Ends after Debranching
2.6. In Vitro Digestion of GlgB-Treated Starch Products
3. Discussion
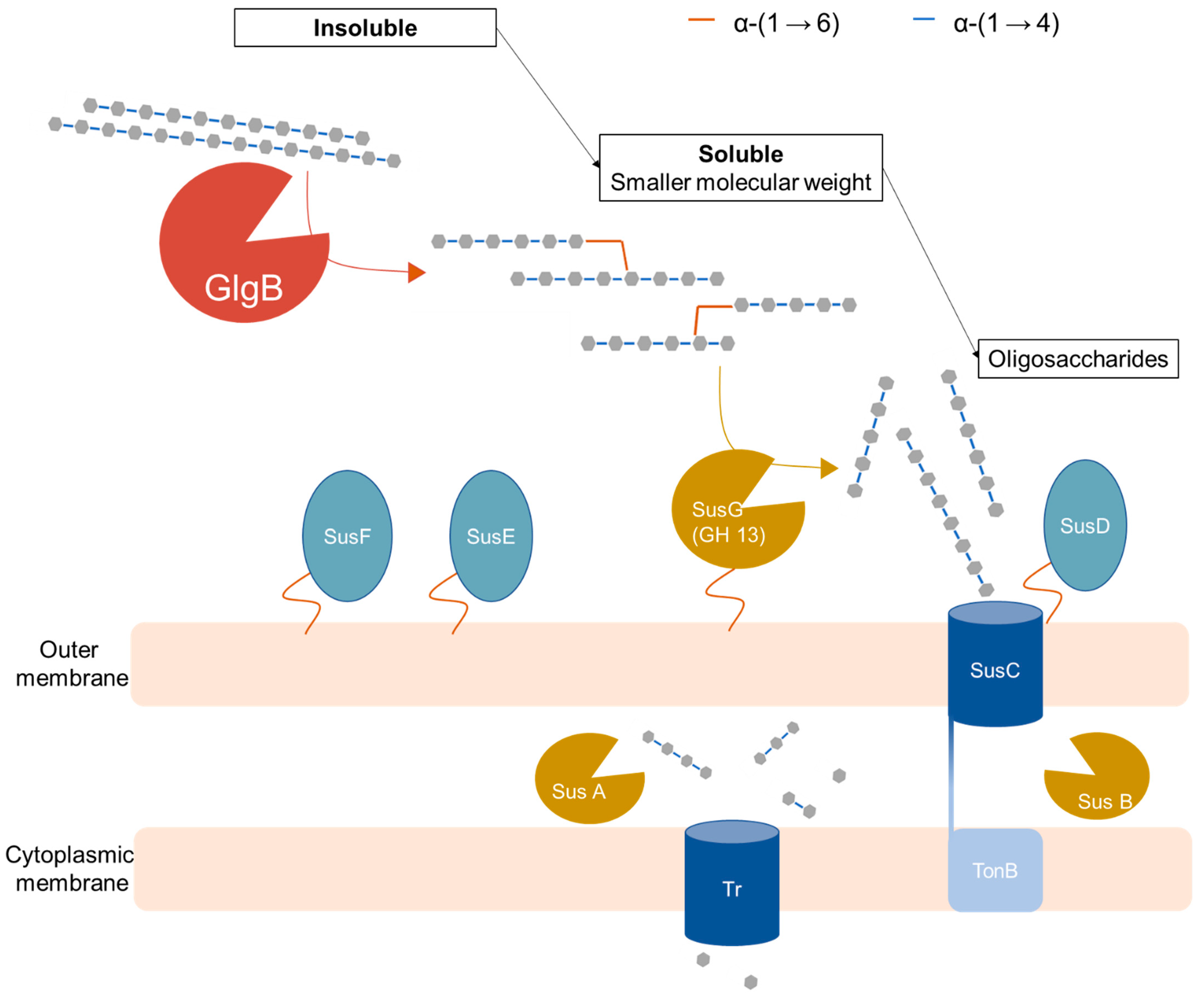
3.1. Biochemical Characteristics of Glucan-Branching Enzyme GlgB
3.2. Physiological Function of Glucan-Branching Enzyme GlgB in Colonic Starch Digestion
3.3. The Use of Glucan-Branching Enzyme GlgB for Starch Conversion in Food Applications
4. Materials and Methods
4.1. Materials
4.2. Bacteria, Plasmid and Growth Condition
4.3. Cloning of Glgb in E. coli
4.4. Overexpression of GlgB and Protein Purification
4.5. Determination of GlgB Activity
4.6. Analysis of Oligosaccharides Profiles by HPAEC-PAD
4.7. Analysis of Molecular Size Distribution by HPSEC-RI
4.8. Determination of GlgB Branching Activity
4.9. In Vitro Digestibility Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Flint, H.J.; Bayer, E.A.; Rincon, M.T.; Lamed, R.; White, B.A. Polysaccharide utilization by gut bacteria: Potential for new insights from genomic analysis. Nat. Rev. Microbiol. 2008, 6, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.; Fouhse, J.M.; Tiwari, U.P.; Li, L.; Willing, B.P. Dietary fiber and intestinal health of monogastric animals. Front. Vet. Sci. 2019, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.L.; Hu, Y.; Gänzle, M.G. Prebiotics, FODMAPs and dietary fiber—Conflicting concepts in development of functional food products? Curr. Opin. Food Sci. 2018, 20, 30–37. [Google Scholar] [CrossRef]
- Nugent, A.P. Health properties of resistant starch. Nutr. Bull. 2005, 30, 27–54. [Google Scholar] [CrossRef]
- Leszczyñski, W. Resistant starch–classification, structure, production. Pol. J. Food Nutr. Sci. 2004, 13, 37–50. [Google Scholar]
- Fouhse, J.M.; Gänzle, M.G.; Regmi, P.R.; van Kempen, T.A.; Zijlstra, R.T. High amylose starch with low in vitro digestibility stimulates hindgut fermentation and has a bifidogenic effect in weaned pigs. J. Nutr. 2015, 145, 2464–2470. [Google Scholar] [CrossRef]
- Wang, W.; Hu, H.; Zijlstra, R.T.; Zheng, J.; Gänzle, M.G. Metagenomic reconstructions of gut microbial metabolism in weanling pigs. Microbiome 2019, 7, 48. [Google Scholar] [CrossRef]
- Deehan, E.C.; Duar, R.M.; Armet, A.M.; Perez-Muñoz, M.E.; Jin, M.; Walter, J. Modulation of the gastrointestinal microbiome with nondigestible fermentable carbohydrates to improve human health. Microbiol. Spectr. 2017, 5, BAD-0019-2017. [Google Scholar] [CrossRef]
- Anderson, K.L.; Salyers, A.A. Biochemical evidence that starch breakdown by Bacteroides thetaiotaomicron involves outer membrane starch-binding sites and periplasmic starch-degrading enzymes. J. Bacteriol. 1989, 171, 3192–3198. [Google Scholar] [CrossRef]
- Reeves, A.R.; Wang, G.R.; Salyers, A.A. Characterization of four outer membrane proteins that play a role in utilization of starch by Bacteroides thetaiotaomicron. J. Bacteriol. 1997, 179, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Shipman, J.A.; Cho, K.H.; Siegel, H.A.; Salyers, A.A. Physiological characterization of SusG, an outer membrane protein essential for starch utilization by Bacteroides thetaiotaomicron. J. Bacteriol. 1999, 181, 7206–7211. [Google Scholar] [CrossRef] [PubMed]
- Ze, X.; Duncan, S.H.; Louis, P.; Flint, H.J. Ruminococcus bromii is a keystone species for the degradation of resistant starch in the human colon. ISME J. 2012, 6, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Cantarel, B.; Henrissat, B.; Gevers, D.; Birren, B.W.; Huttenhower, C.; Ko, G. Gene-targeted metagenomic analysis of glucan-branching enzyme gene profiles among human and animal fecal microbiota. ISME J. 2014, 8, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.G.; Morell, M.K. From bacterial glycogen to starch: Understanding the biogenesis of the plant starch granule. Annu. Rev. Plant Biol. 2003, 54, 207–233. [Google Scholar] [CrossRef]
- Ao, Z.; Simsek, S.; Zhang, G.; Venkatachalam, M.; Reuhs, B.L.; Hamaker, B.R. Starch with a slow digestion property produced by altering its chain length, branch density, and crystalline structure. J. Agric. Food Chem. 2007, 55, 4540–4547. [Google Scholar] [CrossRef]
- Hu, Y.; Heyer, C.M.E.; Wang, W.; Zijlstra, R.T.; Gänzle, M.G. Digestibility of branched and linear α-gluco-oligosaccharides in vitro and in ileal-cannulated pigs. Food Res. Int. 2020, 127, 108726. [Google Scholar] [CrossRef]
- Rumbak, E.; Rawlings, D.E.; Lindsey, G.G.; Woods, D.R. Characterization of the Butyrivibrio fibrisolvens glgB gene, which encodes a glycogen-branching enzyme with starch-clearing activity. J. Bacteriol. 1991, 173, 6732–6741. [Google Scholar] [CrossRef]
- Li, D.; Fei, T.; Wang, Y.; Zhao, Y.; Dai, L.; Fu, X.; Li, X. A cold-active 1,4-α-glucan branching enzyme from Bifidobacterium longum reduces the retrogradation and enhances the slow digestibility of wheat starch. Food Chemistry 2020, 324, 126855. [Google Scholar] [CrossRef]
- Hu, Y.; Winter, V.; Gänzle, M. In vitro digestibility of commercial and experimental isomalto-oligosaccharides. Food Res. Int. 2020, 134, 109250. [Google Scholar] [CrossRef]
- Ban, X.; Li, C.; Gu, Z.; Bao, C.; Qiu, Y.; Hong, Y.; Cheng, L.; Li, Z. Expression and biochemical characterization of a thermostable branching enzyme from Geobacillus thermoglucosidans. Microb. Physiol. 2016, 26, 303–311. [Google Scholar] [CrossRef]
- Ban, X.; Dhoble, A.S.; Li, C.; Gu, Z.; Hong, Y.; Cheng, L.; Holler, T.P.; Kaustubh, B.; Li, Z. Bacterial 1,4-α-glucan branching enzymes: Characteristics, preparation and commercial applications. Crit. Rev. Biotechnol. 2020, 40, 380–396. [Google Scholar] [CrossRef]
- Beck, T.C.; Shinohara, M.L.; Ihara, M.; Abo, M.; Hashida, M.; Takagi, S. A novel thermostable branching enzyme from an extremely thermophilic bacterial species, Rhodothermus obamensis. Appl. Microbiol. Biotechnol. 2001, 57, 653–659. [Google Scholar] [CrossRef]
- Kim, E.-J.; Ryu, S.-I.; Bae, H.-A.; Huong, N.T.; Lee, S.-B. Biochemical characterisation of a glycogen branching enzyme from Streptococcus mutans: Enzymatic modification of starch. Food Chem. 2008, 110, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Kittisuban, P.; Lee, B.-H.; Suphantharika, M.; Hamaker, B.R. Slow glucose release property of enzyme-synthesized highly branched maltodextrins differs among starch sources. Carbohydr. Polym. 2014, 107, 182–191. [Google Scholar] [CrossRef] [PubMed]
- van der Maarel, M.J.E.C.; Vos, A.; Sanders, P.; Dijkhuizen, L. Properties of the glucan branching enzyme of the hyperthermophilic bacterium Aquifex aeolicus. Biocatal. Biotransformation 2003, 21, 199–207. [Google Scholar] [CrossRef]
- Ye, X.; Liu, W.; Ma, S.; Chen, X.; Qiao, Y.; Zhao, Y.; Fan, Q.; Li, X.; Dong, C.; Fang, X.; et al. Expression and characterization of 1,4-α-glucan branching enzyme from Microvirga sp. MC18 and its application in the preparation of slowly digestible starch. Protein Expr. Purif. 2021, 185, 105898. [Google Scholar] [CrossRef]
- Zhang, X.; Leemhuis, H.; van der Maarel, M.J.E.C. Characterization of the GH13 and GH57 glycogen branching enzymes from Petrotoga mobilis SJ95 and potential role in glycogen biosynthesis. PLoS ONE 2019, 14, e0219844. [Google Scholar] [CrossRef]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch—Composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Roussel, X.; Lancelon-Pin, C.; Viksø-Nielsen, A.; Rolland-Sabaté, A.; Grimaud, F.; Potocki-Véronèse, G.; Buléon, A.; Putaux, J.-L.; D’Hulst, C. Characterization of substrate and product specificity of the purified recombinant glycogen branching enzyme of Rhodothermus obamensis. Biochim. Biophys. Acta BBA Gen. Subj. 2013, 1830, 2167–2177. [Google Scholar] [CrossRef]
- Cerqueira, F.M.; Photenhauer, A.L.; Pollet, R.M.; Brown, H.A.; Koropatkin, N.M. Starch digestion by gut bacteria: Crowdsourcing for carbs. Trends Microbiol. 2020, 28, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Koropatkin, N.M.; Smith, T.J. SusG: A unique cell-membrane-associated α-amylase from a prominent human gut symbiont targets complex starch molecules. Structure 2010, 18, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-H.; Yan, L.; Phillips, R.J.; Reuhs, B.L.; Jones, K.; Rose, D.R.; Nichols, B.L.; Quezada-Calvillo, R.; Yoo, S.-H.; Hamaker, B.R. Enzyme-synthesized highly branched maltodextrins have slow glucose generation at the mucosal α-glucosidase level and are slowly digestible in vivo. PLoS ONE 2013, 8, e59745. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; Jiang, B.; Jin, Z.; BeMiller, J.N. Microbial starch-converting enzymes: Recent insights and perspectives. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1238–1260. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, C.; Gu, Z.; Qiu, Y.; Cheng, L.; Hong, Y.; Li, Z. Relationship between structure and retrogradation properties of corn starch treated with 1,4-α-glucan branching enzyme. Food Hydrocoll. 2016, 52, 868–875. [Google Scholar] [CrossRef]
- Wu, S.; Liu, Y.; Yan, Q.; Jiang, Z. Gene cloning, functional expression and characterisation of a novel glycogen branching enzyme from Rhizomucor miehei and its application in wheat breadmaking. Food Chem. 2014, 159, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Na, Y.; Kim, J.; Kang, S.D.; Park, K.-H. Properties and applications of starch modifying enzymes for use in the baking industry. Food Sci. Biotechnol. 2017, 27, 299–312. [Google Scholar] [CrossRef]
- Li, H.; Gidley, M.J.; Dhital, S. High-amylose starches to bridge the “Fiber Gap”: Development, structure, and nutritional functionality. Compr. Rev. Food Sci. Food Saf. 2019, 18, 362–379. [Google Scholar] [CrossRef]
- Li, C.; Dhital, S.; Gidley, M.J. High-amylose wheat bread with reduced in vitro digestion rate and enhanced resistant starch content. Food Hydrocoll. 2022, 123, 107181. [Google Scholar] [CrossRef]
- van Hung, P.; Yamamori, M.; Morita, N. Formation of enzyme-resistant starch in bread as affected by high-amylose wheat flour substitutions. Cereal Chem. J. 2005, 82, 690–694. [Google Scholar] [CrossRef]
- Waffenschmidt, S.; Jaenicke, L. Assay of reducing sugars in the nanomole range with 2, 2′-bicinchoninate. Anal. Biochem. 1987, 165, 337–340. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amylose Content (g/L) | ||
|---|---|---|
| Control | GlgB Treatment | |
| Amylose | 7.65 ± 0.60 | 0.23 ± 0.05 |
| Pea | 4.53 ± 0.31 | 0.14 ± 0.03 |
| Fava bean | 4.37 ± 0.14 | 0.14 ± 0.03 |
| Potato | 3.91 ± 0.16 | 0.12 ± 0.02 |
| Corn | 3.56 ± 0.28 | 0.44 ± 0.07 |
| Wheat | 3.21 ± 0.31 | 0.31 ± 0.04 |
| Barley | 2.98 ± 0.25 | 0.46 ± 0.02 |
| Reducing Ends μM/g Starch | ||
|---|---|---|
| Control | GlgB Treatment | |
| Amylose | 71.4 ± 3.6 | 239 ± 11 * |
| Pea starch | 223 ± 5.7 | 250 ± 15 |
| Fava bean starch | 215 ± 2.1 | 248 ± 10 * |
| Potato starch | 239 ± 10 | 252 ± 15 |
| Corn starch | 236 ± 8.2 | 258 ± 11 * |
| Wheat starch | 229 ± 1.5 | 249 ± 18 |
| Barley starch | 234 ± 0.1 | 251 ± 14 |
| Glucose Release (g/L) | ||
|---|---|---|
| Control | GlgB Treatment | |
| Pea starch | 5.06 ± 0.31 | 4.44 ± 0.23 * |
| Fava bean starch | 5.10 ± 0.44 | 4.25 ± 0.47 * |
| Potato starch | 5.35 ± 0.33 | 4.87 ± 0.24 |
| Corn starch | 4.96 ± 0.78 | 4.48 ± 0.57 |
| Wheat starch | 5.05 ± 0.42 | 4.40 ± 0.45 * |
| Barley starch | 5.35 ± 0.55 | 4.69 ± 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, Y.; Wang, W.; Hu, Y.; Gänzle, M.G. Characterization of the Glucan-Branching Enzyme GlgB Gene from Swine Intestinal Bacteria. Molecules 2023, 28, 1881. https://doi.org/10.3390/molecules28041881
Shao Y, Wang W, Hu Y, Gänzle MG. Characterization of the Glucan-Branching Enzyme GlgB Gene from Swine Intestinal Bacteria. Molecules. 2023; 28(4):1881. https://doi.org/10.3390/molecules28041881
Chicago/Turabian StyleShao, Yuqi, Weilan Wang, Ying Hu, and Michael G. Gänzle. 2023. "Characterization of the Glucan-Branching Enzyme GlgB Gene from Swine Intestinal Bacteria" Molecules 28, no. 4: 1881. https://doi.org/10.3390/molecules28041881
APA StyleShao, Y., Wang, W., Hu, Y., & Gänzle, M. G. (2023). Characterization of the Glucan-Branching Enzyme GlgB Gene from Swine Intestinal Bacteria. Molecules, 28(4), 1881. https://doi.org/10.3390/molecules28041881