
Effects of Melaleuca alternifolia Chell (Tea Tree) and Eucalyptus globulus Labill. Essential Oils on Antibiotic-Resistant Bacterial Biofilms

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. Qualitative and Semi-Quantitative Analysis
2.2. Determination of the Fractional Inhibitory (FIC) Index
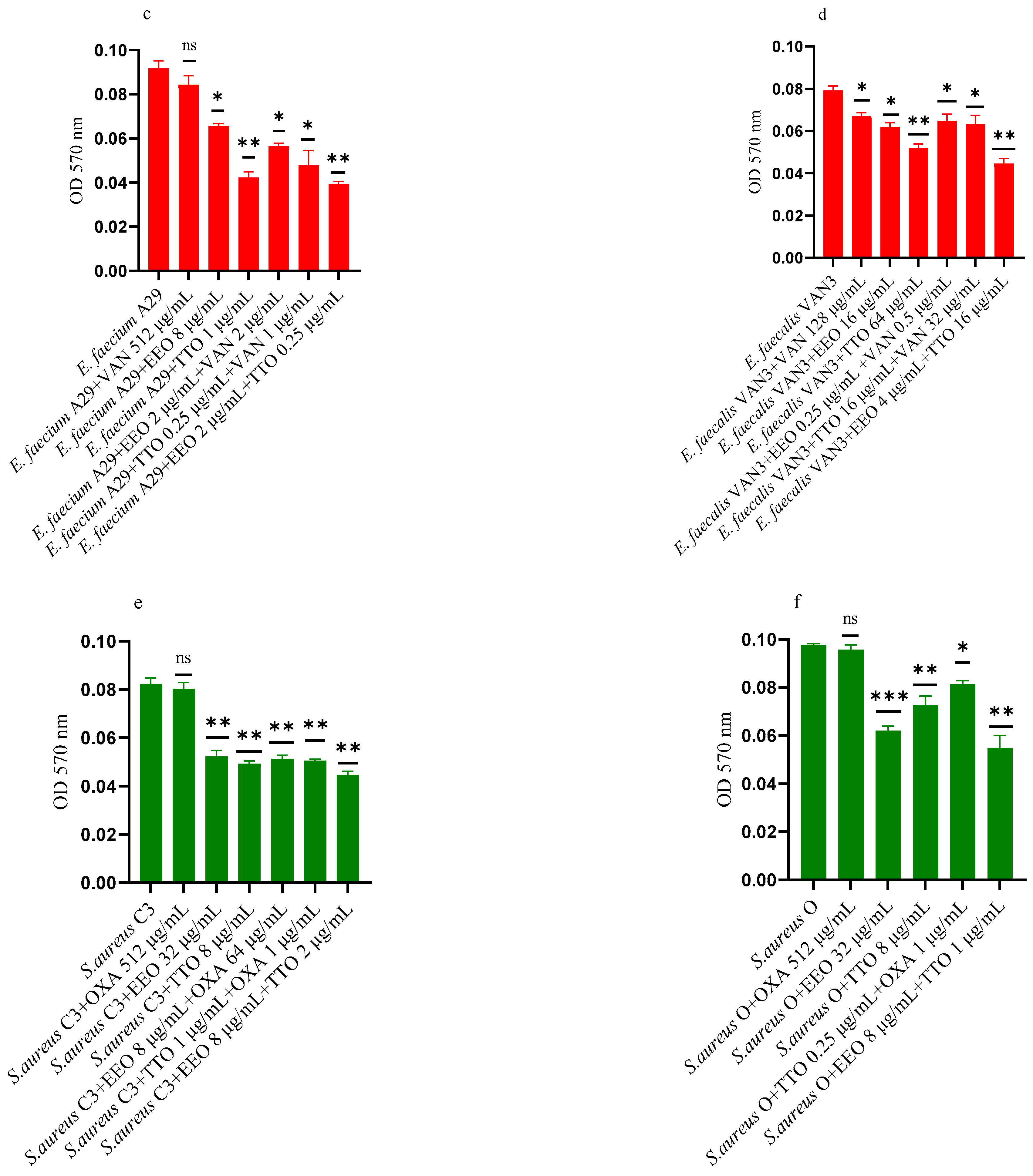
2.3. Effect of EOs on Mono-Species Biofilm Formation
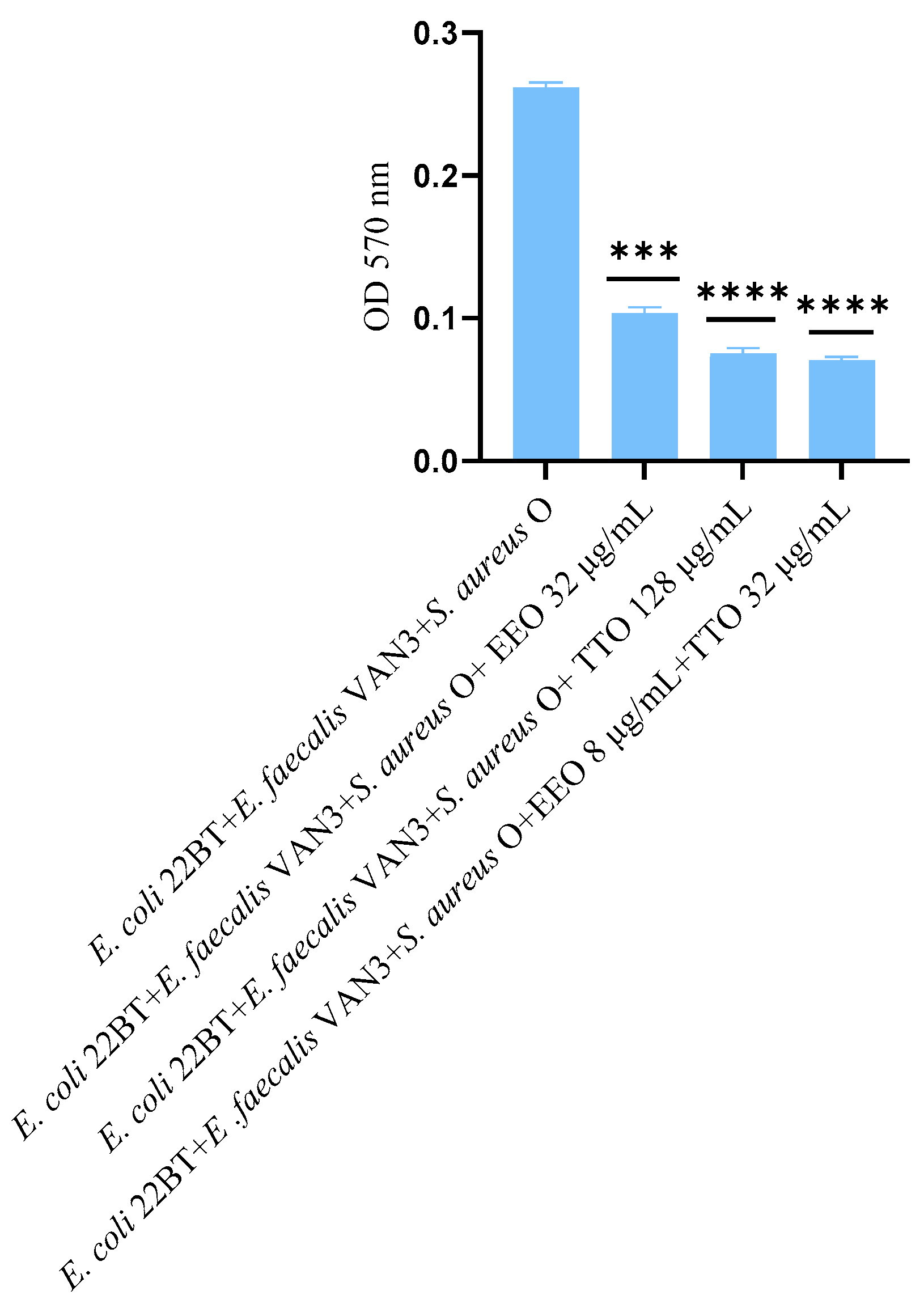
2.4. Effect of EOs on Multi-Species Biofilm Formation
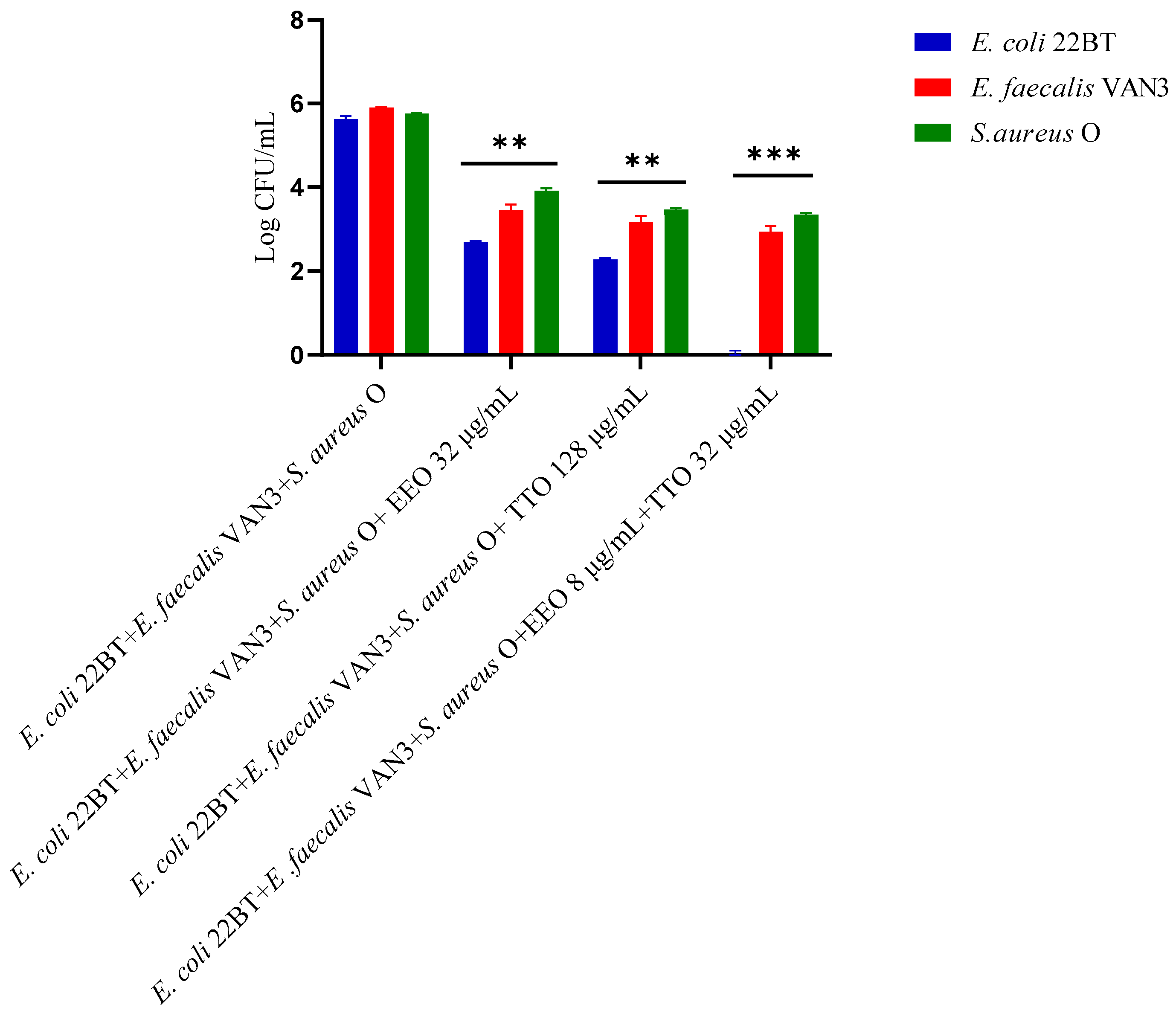
2.5. Quantification of Viable Cells in Multi-Species Biofilm Formation
2.6. Effect of EOs on Mono-Species Mature Biofilm
2.7. Effect of EOs on Multi-Species Mature Biofilm
2.8. Quantification of Viable Cells in Multi-Species Mature Biofilm
2.9. Epifluorescence Microscopy Observation of Mature Biofilm
3. Discussion
4. Strength and Limits of Research
5. Materials and Methods
5.1. Microbial Strains and Essential Oils
5.2. GC-MS Analysis
5.3. GC-FID Analysis
5.4. Qualitative and Semi-Quantitative Analysis
5.5. The Minimum Inhibitory Concentration (MIC) of EOs and Antibiotics
5.6. Determination of the Fractional Inhibitory (FIC) Index
5.7. EOs Activity on Biofilm Produced by Single and Mixed Strains
5.8. Quantification of Viable Cells in Multi-Species Biofilm
5.9. Epifluorescence Microscopy Observation of Mature Biofilm
5.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 625–655. [Google Scholar] [CrossRef]
- World Health Organization. Ten Threats to Global Health in 2019. 2019. Available online: https://www.who.int/emergencies/ten-threats-to-global-health-in-2019 (accessed on 17 January 2019).
- Bereket, W.; Hemalatha, K.; Getenet, B.; Wondwossen, T.; Solomon, A.; Zeynudin, A.; Kannan, S. Update on bacterial nosocomial infections. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 1039–1044. [Google Scholar]
- Davey, M.E.; O’Toole, G.A. Microbial Biofilms: From Ecology to Molecular Genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef]
- Evena, C.; Marlièreb, C.; Ghigoc, J.M.; Allaind, J.M.; Marcellanf, A.; Raspauda, E. Recent advances in studying single bacteria and biofilm mechanics. Adv. Colloid Interface Sci. 2017, 247, 573–588. [Google Scholar] [CrossRef]
- Messi, P. Biofilm formation, development and relevance. In Biofilm in Bioengineering; Nova Science Publishers, Inc.: New York, NY, USA, 2013; Volume 268, pp. 1–26. [Google Scholar]
- Rasmussen, T.B.; Givskov, M. Quorum-sensing inhibitors as anti-pathogenic drugs. Int. J. Med. Microbiol. 2006, 296, 149–161. [Google Scholar] [CrossRef]
- Evans, D.J.; Brown, M.R.W.; Allison, D.G.; Gilbert, P. Susceptibility of bacterial biofilms to tobramycin: Role of specific growth rate and phase in the division cycle. J. Antimicrob. Chemother. 1990, 25, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Batoni, G.; Maisetta, G.; Brancatisano, F.L.; Esin, S.; Campa, M. Use of antimicrobial peptides against microbial biofilms: Advantages and limits. Curr. Med. Chem. 2011, 18, 256–279. [Google Scholar] [CrossRef]
- Newell, D.G.; Fearnley, C. Sources of Campylobacter colonization in broiler chickens. Appl. Environ. Microbiol. 2003, 69, 4343–4351. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef]
- Singhai, M.; Malik, A.; Shahid, M.; Malik, M.A.; Goyal, R. A study on device-related infections with special reference to biofilm production and antibiotic resistance. J. Glob. Infect. Dis. 2012, 4, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Murugaiyan, J.; Kumar, P.A.; Rao, G.S.; Iskandar, K.; Hawser, S.; Hays, J.P.; Mohsen, Y.; Adukkadukkam, S.; Awuah, W.A.; Jose, R.A.M.; et al. Progress in Alternative Strategies to Combat Antimicrobial Resistance: Focus on Antibiotics. Antibiotics 2022, 11, 200. [Google Scholar] [CrossRef] [PubMed]
- Deans, S.G.; Ritchie, G. Antibacterial properties of plant essential oils. Int. J. Food Microbiol. 1987, 5, 165–180. [Google Scholar] [CrossRef]
- Oussalah, M.; Caillet, S.; Lacroix, M. Mechanism of action of Spanish oregano, Chinese cinnamon, and savory essential oils against cell membranes and walls of Escherichia coli O157: H7 and Listeria monocytogenes. J. Food Prot. 2006, 69, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.K.; Malik, A. Liquid and vapour-phase antifungal activities of selected essential oils against Candida albicans: Microscopic observations and chemical characterization of Cymbopogon citratus. BMC Complement. Altern. Med. 2010, 10, 65. [Google Scholar] [CrossRef]
- Kim, S.I.; Roh, J.Y.; Kim, D.H.; Lee, H.S.; Ahn, Y.J. Insecticidal activities of aromatic plant extracts and essential oils against Sitophilus oryzae and Callosobruchus chinensis. J. Stored. Prod. Res. 2003, 39, 293–303. [Google Scholar] [CrossRef]
- Garozzo, A.; Timpanaro, R.; Stivala, A.; Bisignano, G.; Castro, A. Activity of Melaleuca alternifolia (tea tree) oil on influenza virus A/PR/8: Study on the mechanism of action. Antivir. Res. 2011, 89, 83–88. [Google Scholar] [CrossRef] [PubMed]
- George, D.R.; Smith, T.J.; Shiel, R.S.; Sparagano, O.A.E.; Guy, J.H. Mode of action and variability in efficacy of plant essential oils showing toxicity against the poultry red mite, Dermanyssus gallinae. Vet. Parasitol. 2009, 161, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.H.; Mohamed, M.S.M.; Khalil, M.S.; Azmy, M.; Mabrouk, M.I. Combination of essential oil and ciprofloxacin to inhibit/eradicate biofilms in multidrug-resistant Klebsiella pneumoniae. J. Appl. Microbiol. 2018, 125, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Sagar, P.K.; Singh, R. Anti-quorum sensing activity of clove (Syzygium aromaticum) bud extract and its combined efficacy with ceftazidime against multidrug-resistant Pseudomonas aeruginosa. Res. J. Biotechnol. 2021, 16, 75–82. [Google Scholar]
- Brożyna, M.; Paleczny, J.; Kozłowska, W.; Chodaczek, G.; Dudek-Wicher, R.; Felińczak, A.; Gołębiewska, J.; Górniak, A.; Junka, A. The antimicrobial and antibiofilm in vitro activity of liquid and vapour phases of selected essential oils against Staphylococcus aureus. Pathogens 2021, 10, 1207. [Google Scholar] [CrossRef]
- Liu, T.; Wang, J.; Gong, X.; Wu, X.; Liu, L.; Chi, F. Rosemary and Tea Tree essential oils exert antibiofilm activities in vitro against Staphylococcus aureus and Escherichia coli. J. Food Prot. 2020, 83, 1261–1267. [Google Scholar] [CrossRef]
- Francisconi, R.S.; Huacho, P.M.M.; Tonon, C.C.; Bordini, E.A.F.; Correia, M.F.; Sardi, J.C.O.; Spolidorio, D.M.P. Antibiofilm efficacy of tea tree oil and of its main component terpinen-4-ol against Candida albicans. Braz. Oral Res. 2020, 34, e050. [Google Scholar] [CrossRef]
- Luís, Â.; Duarte, A.; Gominho, J.; Domingues, F.; Duarte, A.P. Chemical composition, antioxidant, antibacterial and anti-quorum sensing activities of Eucalyptus globulus and Eucalyptus radiata essential oils. Ind. Crops Prod. 2016, 79, 274–282. [Google Scholar]
- Merghni, A.; Noumi, E.; Hadded, O.; Dridi, N.; Panwar, H.; Ceylan, O.; Mastouri, M.; Snoussi, M. Assessment of the antibiofilm and antiquorum sensing activities of Eucalyptus globulus essential oil and its main component 1,8-cineole against methicillin-resistant Staphylococcus aureus strains. Microb. Pathog. 2018, 118, 74–80. [Google Scholar] [CrossRef]
- National Institute of Standards and Technology Library Database. Available online: https://webbook.nist.gov/ (accessed on 15 December 2022).
- Babushok, V.I.; Zenkevich, I.G. Retention indices for most frequently reported essential oil compounds in GC. Chromatographia 2009, 69, 257–269. [Google Scholar] [CrossRef]
- Dong, G.; Bai, X.; Aimila, A.; Aisa, H.A.; Maiwulanjiang, M. Study on Lavender essential oil chemical compositions by GC-MS and improved pGC. Molecules 2020, 25, 3166. [Google Scholar] [CrossRef]
- Carson, C.F.; Hammer, K.A.; Riley, T.V. Melaleuca alternifolia (Tea Tree) oil: A review of antimicrobial and other medicinal properties. Clin. Microbiol. Rev. 2006, 19, 50–62. [Google Scholar] [CrossRef]
- Noumi, E.; Snoussi, M.; Hajlaoui, H.; Trabelsi, N.; Ksouri, R.; Valentin, E.; Bakhrouf, A. Chemical composition, antioxidant and antifungal potential of Melaleuca alternifolia (Tea Tree) and Eucalyptus globulus essential oils against oral Candida species. J. Med. Plants Res. 2011, 5, 4147–4156. [Google Scholar]
- Brun, P.; Bernabè, G.; Filippini, R.; Piovan, A. In vitro antimicrobial activities of commercially available Tea Tree (Melaleuca alternifolia) essential oils. Curr. Microbiol. 2019, 76, 108–116. [Google Scholar] [CrossRef]
- Kim, H.J.; Chen, F.; Wu, C.; Wang, X.; Chung, H.Y.; Jin, Z. Evaluation of antioxidant activity of australian Tea Tree (Melaleuca alternifolia) oil and its components. J. Agric. Food Chem. 2004, 52, 2849–2854. [Google Scholar] [CrossRef]
- Elmi, A.; Ventrella, D.; Barone, F.; Carnevali, G.; Filippini, G.; Pisi, A.; Benvenuti, S.; Scozzoli, M.; Bacci, M.L. In vitro effects of Tea Tree oil (Melaleuca alternifolia essential oil) and its principal component terpinen-4-ol on swine spermatozoa. Molecules 2019, 24, 1071. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential oils as antimicrobial agents-myth or real alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef]
- Ghaffar, A.; Yameen, M.; Kiran, S.; Kamal, S.; Jalal, F.; Munir, B.; Saleem, S.; Rafiq, N.; Ahmad, A.; Saba, I.; et al. Chemical composition and in-vitro evaluation of the antimicrobial and antioxidant activities of essential oils extracted from seven Eucalyptus species. Molecules 2015, 20, 20487–20498. [Google Scholar] [CrossRef]
- Alipanah, H.; Abdollahi, A.; Firooziyan, S.; Zarenezhad, E.; Jafari, M.; Osanloo, M. Nanoemulsion and nanogel containing Eucalyptus globulus essential oil; larvicidal activity and antibacterial properties. Interdiscip. Perspect. Infect. Dis. 2022, 2022, 1616149. [Google Scholar] [CrossRef]
- Quatrin, P.M.; Verdi, C.M.; de Souza, M.E.; de Godoi, S.N.; Klein, B.; Gunde, A.; Wagner, R.; de Almeida Vaucher, R.; Ourique, A.F.; Santos, R.C.V. Antimicrobial and antibiofilm activities of nanoemulsions containing Eucalyptus globulus oil against Pseudomonas aeruginosa and Candida spp. Microb. Pathog. 2017, 112, 230–242. [Google Scholar] [CrossRef]
- Iseppi, R.; Mariani, M.; Condò, C.; Sabia, C.; Messi, P. Essential oils: A natural weapon against antibiotic-resistant bacteria responsible for nosocomial infections. Antibiotics 2021, 10, 417. [Google Scholar] [CrossRef]
- Yap, P.S.; Lim, S.H.; Hu, C.P.; Yiap, B.C. Combination of essential oils and antibiotics reduce antibiotic resistance in plasmid conferred multidrug resistant bacteria. Phytomedicine 2013, 20, 710–713. [Google Scholar] [CrossRef]
- Cordeiro, L.; Figueired, P.; Souza, H.; Sousa, A.; Andrade-Júnior, F.; Medeiros, D.; Nóbrega, J.; Silva, D.; Martins, E.; Barbosa-Filho, J.; et al. Terpinen-4-ol as an antibacterial and antibiofilm agent against Staphylococcus Aureus. Int. J. Mol. Sci. 2020, 21, 4531. [Google Scholar] [CrossRef]
- Borotová, P.; Galovičová, L.; Vukovic, N.L.; Vukic, M.; Tvrdá, E.; Kačániová, M. Chemical and biological characterization of Melaleuca alternifolia essential oil. Plants 2022, 11, 558. [Google Scholar] [CrossRef]
- Hendry, E.R.; Worthington, T.; Conway, B.R.; Lambert, P.A. Antimicrobial efficacy of Eucalyptus oil and 1,8-cineole alone and in combination with chlorhexidine digluconate against microorganisms grown in planktonic and biofilm cultures. J. Antimicrob. Chemother. 2009, 64, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, P.; Pruss, A.; Grygorcewicz, B.; Wojciuk, B.; Dołęgowska, B.; Giedrys-Kalemba, S.; Kochan, E.; Sienkiewicz, M. Preliminary study on the antibacterial activity of essential oils alone and in combination with Gentamicin against extended-spectrum-β-lactamase-producing and New Delhi metallo-β-Lactamase-1-Producing Klebsiella pneumoniae isolates. Microb. Drug Resist. 2018, 24, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Owen, L.; Webb, J.P.; Green, J.; Smith, L.J.; Laird, K. From formulation to in vivo model: A comprehensive study of a synergistic relationship between vancomycin, carvacrol, and cuminaldehyde against Enterococcus faecium. Phytother. Res. 2020, 34, 1638–1649. [Google Scholar] [CrossRef] [PubMed]
- Mulyaningsih, S.; Sporer, F.; Zimmermann, S.; Reichling, J.; Wink, M. Synergistic properties of the terpenoids aromadendrene and 1,8-cineole from the essential oil of Eucalyptus globulus against antibiotic-susceptible and antibiotic-resistant pathogens. Phytomedicine 2010, 17, 1061–1066. [Google Scholar] [CrossRef]
- Alexopoulos, A.; Kimbaris, A.C.; Plessas, S.; Mantzouran, I.; Voidarou, C.; Pagonopoulou, O.; Tsigalou, C.; Fournomiti, M.; Bontsidis, C.; Stavropoulou, E.; et al. Combined action of Piperitenone Epoxide and antibiotics against clinical isolates of Staphylococcus aureus and Escherichia coli. Front. Microbiol. 2019, 10, 2607. [Google Scholar] [CrossRef]
- Kon, K.V.; Rai, M.K. Plant essential oils and their constituents in coping with multidrug-resistant bacteria. Expert Rev. Anti-Infect. Ther. 2012, 10, 775–790. [Google Scholar] [CrossRef]
- Scandorieiro, S.; Rodrigues, B.C.D.; Nishio, E.K.; Panagio, L.A.; de Oliveira, A.G.; Durán, N.; Nakazato, G.; Kobayashi, R.K.T. Biogenic silver nanoparticles strategically combined with Origanum vulgare derivatives: Antibacterial mechanism of action and effect on multidrug-resistant strains. Front. Microbiol. 2022, 13, 842600. [Google Scholar] [CrossRef]
- Kwieciński, J.; Eick, S.; Wójcik, K. Effects of tea tree (Melaleuca alternifolia) oil on Staphylococcus aureus in biofilms and stationary growth phase. Int. J. Antimicrob. Agents 2009, 33, 343–347. [Google Scholar] [CrossRef]
- Tranchida, P.Q.; Shellie, R.A.; Purcaro, G.; Conte, L.S.; Dugo, P.; Dugo, G.; Mondello, L. Analysis of fresh and aged tea tree essential oils by using GCxGC-qMS. J. Chromatogr Sci. 2010, 48, 262–266. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing: Chicago, IL, USA, 2007. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019; Volume 39. [Google Scholar]
- Hemaiswaryaa, S.; Kruthiventib, A.K.; Doblea, M. Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 2008, 15, 639–652. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| a Lit. RI | b Exp. RI | Melaleuca alternifolia | Eucalyptus globulus | |
|---|---|---|---|---|
| α-thujene | 928 | 926 | 0.88 c | - |
| α-pinene | 936 | 932 | 2.56 ± 0.1 | 2.33 ± 0.1 |
| camphene | 950 | 947 | - | 0.51 c |
| sabinene | 973 | 972 | 0.07 c | - |
| β-pinene | 978 | 975 | 0.73 c | 0.99 c |
| β-myrcene | 989 | 991 | 0.80 c | 0.83 c |
| α-phellandrene | 1004 | 1004 | 0.45 c | 0.23 c |
| α-terpinene | 1017 | 1017 | 8.89 ± 0.2 | 0.09 c |
| p-cymene | 1024 | 1024 | 2.84 ± 0.1 | - |
| limonene | 1029 | 1028 | 1.88 ± 0.1 | 0.93 c |
| 1,8-cineole | 1032 | 1030 | 2.35 ± 0.1 | 58.07 ± 0.2 |
| cis-β-ocimene | 1038 | 1039 | - | 0.21 c |
| γ-terpinene | 1060 | 1059 | 20.16 ± 0.2 | 1.11 c |
| linalool | 1099 | 1100 | 0.49 c | 12.05 ± 0.2 |
| camphor | 1143 | 1146 | - | 4.39 ± 0.1 |
| borneol | 1166 | 1167 | - | 1.00 c |
| terpinen-4-ol | 1177 | 1179 | 43.29 ± 0.2 | 0.66 c |
| α-terpineol | 1190 | 1192 | 2.99 ± 0.1 | 0.71 c |
| linalyl acetate | 1263 | 1264 | - | 10.95 ± 0.2 |
| α-cubebene | 1351 | 1353 | 0.06 c | - |
| α-copaene | 1376 | 1379 | 0.10 c | - |
| α-gurjunene | 1409 | 1414 | 0.32 c | - |
| β-caryophyllene | 1420 | 1426 | 0.32 c | 0.98 c |
| aromadendrene | 1440 | 1445 | 1.07 c | - |
| α-humulene | 1453 | 1460 | 0.09 c | 0.22 c |
| allo-aromadendrene | 1460 | 1467 | 0.47 c | - |
| germacrene D | 1481 | 1488 | - | 0.11 c |
| α-selinene | 1493 | 1496 | 0.14 c | - |
| ledene | 1495 | 1501 | 1.31 c | - |
| δ-cadinene | 1523 | 1530 | 1.03 c | - |
| globulol | 1582 | 1586 | 0.06 c | - |
| caryophyllene oxide | 1589 | 1593 | 0.19 c | 0.06 c |
| viridiflorol | 1591 | 1601 | 0.17 c | - |
| Total identified | 97.07 | 96.55 |
| Strains | Drug/EO | MIC Alone (μg/mL) | MIC EO/EO and EO/Drug (μg/mL) | DCL | Strains | Drug/EO | MIC Alone (μg/mL) | MIC EO/EO and EO/Drug (μg/mL) | DCL |
|---|---|---|---|---|---|---|---|---|---|
| E. coli 22BT | E. coli 45DT | ||||||||
| CTX EEO | 64 32 | 16 8 | 4-fold 4-fold | CTX EEO | 32 128 | 8 32 | 4-fold 4-fold | ||
| CTX TTO | 64 128 | 16 32 | 4-fold 4-fold | CTX TTO | n.s | n.s | n.s | ||
| EEO TTO | n.s | n.s | n.s | EEO TTO | 128 1 | 32 0.25 | 4-fold 4-fold | ||
| E. faecium A29 | E. faecalis VAN3 | ||||||||
| VAN EEO | 512 8 | 2 2 | 256-fold 4-fold | VAN EEO | 128 16 | 0.5 0.25 | 256-fold 64-fold | ||
| VAN TTO | 512 1 | 1 0.25 | 512-fold 4-fold | VAN TTO | 128 64 | 32 16 | 4-fold 4-fold | ||
| EEO TTO | 8 1 | 2 0.25 | 4-fold 4-fold | EEO TTO | 16 64 | 4 16 | 4-fold 4-fold | ||
| S. aureus C3 | S. aureus O | ||||||||
| OXA EEO | 512 32 | 64 8 | 8-fold 4-fold | OXA EEO | n.s | n.s | n.s | ||
| OXA TTO | 512 8 | 1 1 | 512-fold 8-fold | OXA TTO | 512 8 | 1 0.25 | 512-fold 32-fold | ||
| EEO TTO | 32 8 | 8 2 | 4-fold 4-fold | EEO TTO | 32 8 | 8 1 | 4-fold 8-fold | ||
| Strains | EEO (µg/mL) | TTO (µg/mL) | CTX (µg/mL) | VAN (µg/mL) | OXA (µg/mL) |
|---|---|---|---|---|---|
| Escherichia coli 22BT | 32 | 128 | 64 | n.a | n.a |
| Escherichia coli 45DT | 128 | 1 | 32 | n.a | n.a |
| Enterococcus faecium A29 | 8 | 1 | n.a | 512 | n.a |
| Enterococcus faecalis VAN 3 | 16 | 64 | n.a | 128 | n.a |
| Staphylococcus aureus C3 | 32 | 8 | n.a | n.a | 512 |
| Staphylococcus aureus O | 32 | 8 | n.a | n.a | 512 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iseppi, R.; Mariani, M.; Benvenuti, S.; Truzzi, E.; Messi, P. Effects of Melaleuca alternifolia Chell (Tea Tree) and Eucalyptus globulus Labill. Essential Oils on Antibiotic-Resistant Bacterial Biofilms. Molecules 2023, 28, 1671. https://doi.org/10.3390/molecules28041671
Iseppi R, Mariani M, Benvenuti S, Truzzi E, Messi P. Effects of Melaleuca alternifolia Chell (Tea Tree) and Eucalyptus globulus Labill. Essential Oils on Antibiotic-Resistant Bacterial Biofilms. Molecules. 2023; 28(4):1671. https://doi.org/10.3390/molecules28041671
Chicago/Turabian StyleIseppi, Ramona, Martina Mariani, Stefania Benvenuti, Eleonora Truzzi, and Patrizia Messi. 2023. "Effects of Melaleuca alternifolia Chell (Tea Tree) and Eucalyptus globulus Labill. Essential Oils on Antibiotic-Resistant Bacterial Biofilms" Molecules 28, no. 4: 1671. https://doi.org/10.3390/molecules28041671
APA StyleIseppi, R., Mariani, M., Benvenuti, S., Truzzi, E., & Messi, P. (2023). Effects of Melaleuca alternifolia Chell (Tea Tree) and Eucalyptus globulus Labill. Essential Oils on Antibiotic-Resistant Bacterial Biofilms. Molecules, 28(4), 1671. https://doi.org/10.3390/molecules28041671