
Postbiotics Derived from L. paracasei ET-22 Inhibit the Formation of S. mutans Biofilms and Bioactive Substances: An Analysis

 , and
, and
Abstract
1. Introduction
2. Results
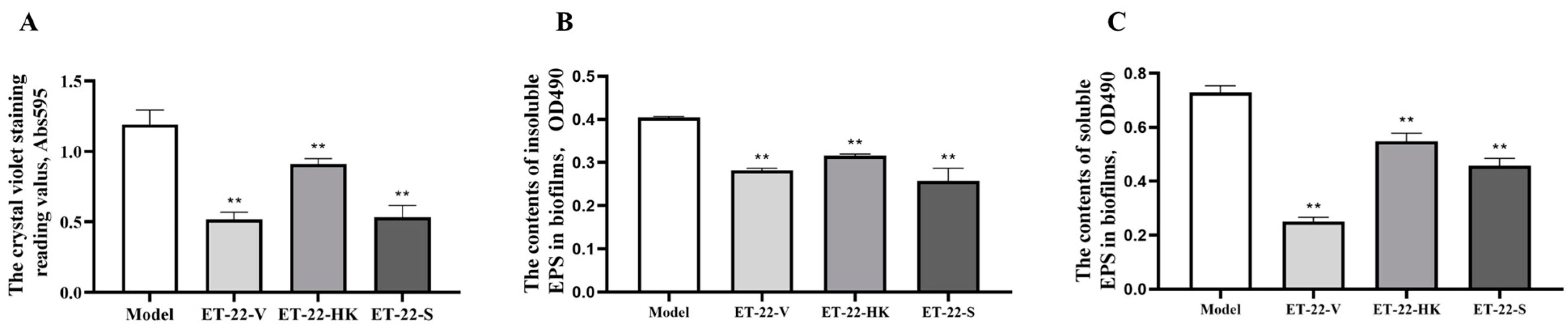
2.1. Effects of L. paracasei ET-22 on the Biofilm Formation and EPS Production of S. mutans
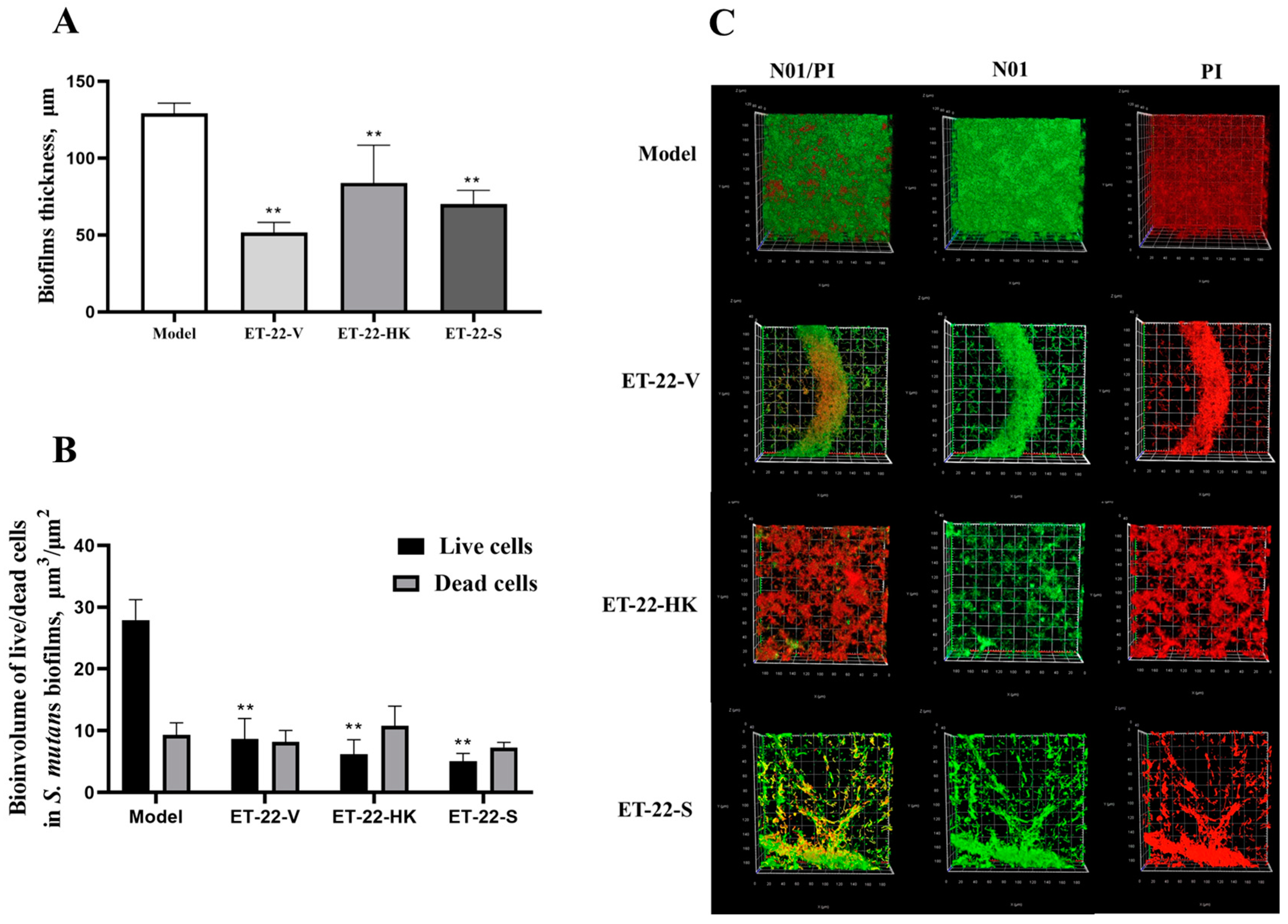
2.2. Biofilm Microstructure Is Changed by L. paracasei ET-22
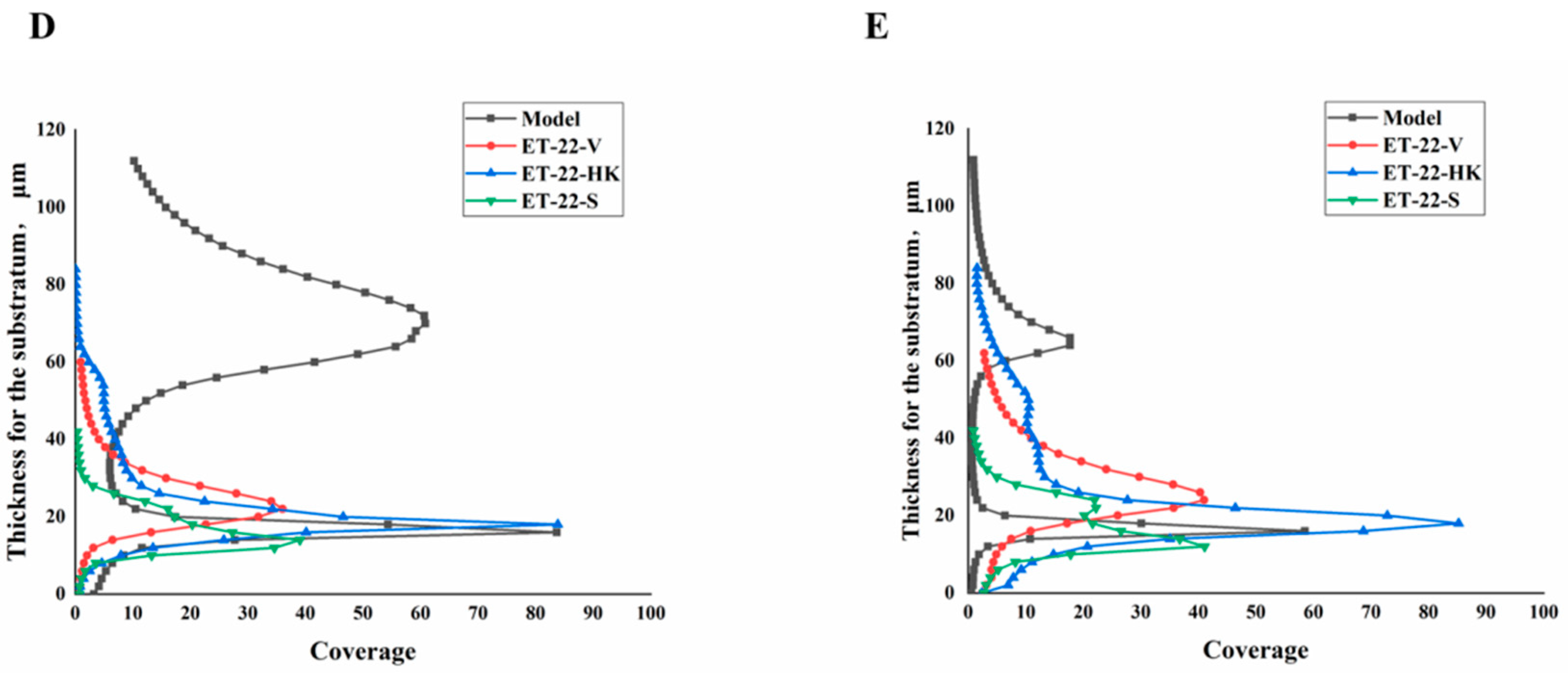
2.3. Biofilm Thickness Is Changed by L. paracasei ET-22
2.4. The Expression of Virulence Genes and the Quorum-Sensing System Are Regulated by L. paracasei ET-22
2.5. The Bioactive Substances for Resisting Formations of S. mutans Biofilms in Heat-Killed Bacteria and Secretions of L. paracasei ET-22
3. Discussion
3.1. Live L. paracasei ET-22 and Its Postbiotics Inhibit the Formation of S. mutans Biofilms
3.2. Live L. paracasei ET-22 and Its Postbiotics Inhibit the Formation of S. mutans Biofilms by Blocking the Initial Adhesion
3.3. Live L. paracasei ET-22 and Its Postbiotics Inhibit S. mutans Biofilms by Interfering with the QS System and the Expression of Virulence Factors
3.4. The Inhibitory Effect of Live L. paracasei ET-22 and Its Postbiotics on S. mutans Biofilms Is Mediated by Multiple Components
4. Materials and Methods
4.1. Bacterial Strains and Culture Medium
4.2. Preparation of Live Bacteria, Heat-Killed Bacteria, and Secretions of L. paracasei ET-22
4.3. Treatment for the Formation of S. mutans Biofilms
4.4. Extracellular-Polysaccharide Production in Biofilms
4.5. Biofilm Microstructure Observed Using Scanning Electron Microscopy
4.6. Detection of Biofilm Thickness with Confocal Laser Scanning Microscopy
4.7. RT-qPCR for Gene-Expression Levels
4.8. Active-Substances Determinations in Heat-Killed L. paracasei ET-22 by Non-Targeted Metabonomics
4.9. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- GBD 2017 Disease and Injury Incidence and Prevalence Collaborators. Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the global burden of disease study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef]
- Lin, Y.; Zhou, X.; Li, Y. Strategies for Streptococcus mutans biofilm dispersal through extracellular polymeric substances disruption. Mol. Oral Microbiol. 2022, 37, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, T.; Peng, W.; Zhu, Y. Effects of resveratrol on cariogenic virulence properties of Streptococcus mutans. BMC Microbiol. 2020, 20, 99. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Bidossi, A.; De Grandi, R.; Toscano, M.; Bottagisio, M.; De Vecchi, E.; Gelardi, M.; Drago, L. Probiotics Streptococcus salivarius 24SMB and Streptococcus oralis 89a interfere with biofilm formation of pathogens of the upper respiratory tract. BMC Infect. Dis. 2018, 18, 653. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Hwang, G.; Liu, Y.; Gao, L.; Kilpatrick-Liverman, L.; Santarpia, P.; Zhou, X.; Koo, H. L-arginine modifies the exopolysaccharide matrix and thwarts Streptococcus mutans outgrowth within mixed-species oral biofilms. J. Bacteriol. 2016, 198, 2651–2661. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.N.; Choi, H.M.; Jeon, J.G. Relationship between sucrose concentration and bacteria proportion in a multispecies biofilm. J. Oral Microbiol. 2021, 13, 1910443. [Google Scholar] [CrossRef]
- Alshahrani, A.M.; Gregory, R.L. In vitro Cariostatic effects of cinnamon water extract on nicotine-induced Streptococcus mutans biofilm. BMC Complement. Med. Ther. 2020, 20, 45. [Google Scholar] [CrossRef]
- Sun, Y.; Jiang, W.; Zhang, M.; Zhang, L.; Shen, Y.; Huang, S.; Li, M.; Qiu, W.; Pan, Y.; Zhou, L.; et al. The inhibitory effects of ficin on Streptococcus mutans biofilm formation. BioMed Res. Int. 2021, 2021, 6692328. [Google Scholar] [CrossRef]
- Bowen, W.H.; Koo, H. Biology of Streptococcus mutans-derived glucosyltransferases: Role in extracellular matrix formation of cariogenic biofilms. Caries Res. 2011, 45, 69–86. [Google Scholar] [CrossRef]
- Simón-Soro, A.; Mira, A. Solving the etiology of dental caries. Trends Microbiol. 2015, 23, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Söderling, E.M.; Marttinen, A.M.; Haukioja, A.L. Probiotic lactobacilli interfere with Streptococcus mutans biofilm formation in vitro. Curr. Microbiol. 2011, 62, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Falsetta, M.L.; Klein, M.I.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.J.; Bowen, W.H.; et al. Symbiotic relationship between Streptococcus mutans and Candida albicans synergizes virulence of plaque biofilms in vivo. Infect. Immun. 2014, 82, 1968–1981. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, C.; Chen, J.; Zhou, S.; Zhao, Y.; Xu, M.; Xu, H. Dual mode of anti-biofilm action of G3 against Streptococcus mutans. ACS Appl. Mater. Interfaces 2020, 12, 27866–27875. [Google Scholar] [CrossRef]
- Pedret, A.; Valls, R.M.; Calderón-Pérez, L.; Llauradó, E.; Companys, J.; Pla-Pagà, L.; Moragas, A.; Martín-Luján, F.; Ortega, Y.; Giralt, M.; et al. Effects of daily consumption of the probiotic Bifidobacterium animalis subsp. lactis CECT 8145 on anthropometric adiposity biomarkers in abdominally obese subjects: A randomized controlled trial. Int. J. Obes. 2019, 43, 1863–1868. [Google Scholar] [CrossRef] [PubMed]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Hannig, C.; Hannig, M. The oral cavity—A key system to understand substratum-dependent bioadhesion on solid surfaces in man. Clin. Oral Investig. 2009, 13, 123–139. [Google Scholar] [CrossRef]
- Maehata, H.; Arai, S.; Iwabuchi, N.; Abe, F. Immuno-modulation by heat-killed Lacticaseibacillus paracasei MCC1849 and its application to food products. Int. J. Immunopathol. Pharmacol. 2021, 35. [Google Scholar] [CrossRef]
- Gulube, Z.; Patel, M. Effect of Punica granatum on the virulence factors of cariogenic bacteria Streptococcus mutans. Microb. Pathog. 2016, 98, 45–49. [Google Scholar] [CrossRef]
- Torino, M.I.; Taranto, M.P.; Sesma, F.; De Valdez, G.F. Heterofermentative pattern and exopolysaccharide production by Lactobacillus helveticus ATCC 15807 in response to environmental pH. J. Appl. Microbiol. 2001, 91, 846–852. [Google Scholar] [CrossRef]
- Deng, Y.; Yang, Y.; Zhang, B.; Chen, H.; Lu, Y.; Ren, S.; Lei, L.; Hu, T. The vicK gene of Streptococcus mutans mediates its cariogenicity via exopolysaccharides metabolism. Int. J. Oral Sci. 2021, 13, 45. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.T.; Yates, D.; Ahn, S.J.; Burne, R.A. Biofilm formation and virulence expression by Streptococcus mutans are altered when grown in dual-species model. BMC Microbiol. 2010, 10, 111. [Google Scholar] [CrossRef] [PubMed]
- De Alvarenga, J.A.; De Barros, P.P.; De Camargo Ribeiro, F.; Rossoni, R.D.; Garcia, M.T.; Dos Santos Velloso, M.; Shukla, S.; Fuchs, B.B.; Shukla, A.; Mylonakis, E.; et al. Probiotic effects of Lactobacillus paracasei 28.4 to inhibit Streptococcus mutans in a gellan-based formulation. Probiotics Antimicrob. Proteins 2021, 13, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; He, X.; Chen, J.; Tang, B.; Zheng, T.; Jing, M.; Lin, Y.; Pan, Y.; Ma, Q.; Li, Y.; et al. Transcriptional profiling reveals the importance of RcrR in the regulation of multiple sugar transportation and biofilm formation in Streptococcus mutans. mSystems 2021, 6, e0078821. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, Q. Influence of surface energy of modified surfaces on bacterial adhesion. Biophys. Chem. 2005, 117, 39–45. [Google Scholar] [CrossRef]
- Teughels, W.; Van Assche, N.; Sliepen, I.; Quirynen, M. Effect of material characteristics and/or surface topography on biofilm development. Clin. Oral Implants Res. 2006, 17, 68–81. [Google Scholar] [CrossRef]
- Yang, J.; Deng, D.; Brandt, B.W.; Nazmi, K.; Wu, Y.; Crielaard, W.; Ligtenberg, A. Diversity of SpaP in genetic and salivary agglutinin mediated adherence among Streptococcus mutans strains. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Katsumata, T.; Nguyen-Tra Le, M.; Kawada-Matsuo, M.; Taniguchi, Y.; Ouhara, K.; Oogai, Y.; Nakata, M.; Mizuno, N.; Nishitani, Y.; Komatsuzawa, H. Comprehensive characterization of sortase A-dependent surface proteins in Streptococcus mutans. Microbiol. Immunol. 2022, 66, 145–156. [Google Scholar] [CrossRef]
- Ciandrini, E.; Campana, R.; Baffone, W. Live and heat-killed Lactobacillus spp. interfere with Streptococcus mutans and Streptococcus oralis during biofilm development on titanium surface. Arch. Oral Biol. 2017, 78, 48–57. [Google Scholar] [CrossRef]
- Kuhnert, W.L.; Quivey, R.G., Jr. Genetic and biochemical characterization of the F-ATPase operon from Streptococcus sanguis 10904. J. Bacteriol. 2003, 185, 1525–1533. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, B.; Weir, M.D.; Homayounfar, N.; Fay, G.G.; Martinho, F.; Lei, L.; Bai, Y.; Hu, T.; Xu, H. S. mutans gene-modification and antibacterial resin composite as dual strategy to suppress biofilm acid production and inhibit caries. J. Dent. 2020, 93, 103278. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; McLean, J.S.; Lux, R.; He, X.; Shi, W. The well-coordinated linkage between acidogenicity and aciduricity via insoluble glucans on the surface of Streptococcus mutans. Sci. Rep. 2015, 5, 18015. [Google Scholar] [CrossRef]
- Cao, L.; Zhang, Z.Z.; Xu, S.B.; Ma, M.; Wei, X. Farnesol inhibits development of caries by augmenting oxygen sensitivity and suppressing virulence-associated gene expression in Streptococcus mutans. J. Biomed. Res. 2017, 31, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Bitoun, J.P.; Liao, S.; Yao, X.; Ahn, S.J.; Isoda, R.; Nguyen, A.H.; Wen, Z.T. BrpA is involved in regulation of cell envelope stress responses in Streptococcus mutans. Appl. Environ. Microbiol. 2012, 78, 2914–2922. [Google Scholar] [CrossRef] [PubMed]
- Al-Ansari, M.M.; Al-Dahmash, N.D.; Ranjitsingh, A. Synthesis of silver nanoparticles using gum arabic: Evaluation of its inhibitory action on Streptococcus mutans causing dental caries and endocarditis. J. Infect. Public Health 2021, 14, 324–330. [Google Scholar] [CrossRef]
- Lu, J.; Cheng, L.; Huang, Y.; Jiang, Y.; Chu, C.H.; Peng, X.; Li, M.; Xu, H.; Zhou, X.; Ren, B. Resumptive Streptococcus mutans persisters induced from dimethylaminododecyl methacrylate elevated the cariogenic virulence by up-regulating the Quorum-Sensing and VicRK pathway genes. Front. Microbiol. 2020, 10, 3102. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Jiang, W.; Wang, K.; Luo, J.; Li, W.; Zhou, X.; Zhang, L. Antimicrobial peptide GH12 suppresses cariogenic virulence factors of Streptococcus mutans. J. Oral Microbiol. 2018, 10, 1442089. [Google Scholar] [CrossRef]
- Wu, H.; Guang, C.; Zhang, W.; Mu, W. Recent development of phenyllactic acid: Physicochemical properties, biotechnological production strategies and applications. Crit. Rev. Biotechnol. 2021, 1–16. [Google Scholar] [CrossRef]
- Liu, F.; Sun, Z.; Wang, F.; Liu, Y.; Zhu, Y.; Du, L.; Wang, D.; Xu, W. Inhibition of biofilm formation and exopolysaccharide synthesis of Enterococcus faecalis by phenyllactic acid. Food Microbiol. 2020, 86, 103344. [Google Scholar] [CrossRef]
- Shakya, S.; Danshiitsoodol, N.; Noda, M.; Inoue, Y.; Sugiyama, M. 3-Phenyllactic acid generated in medicinal plant extracts fermented with plant-derived lactic acid bacteria inhibits the biofilm synthesis of Aggregatibacter actinomycetemcomitans. Front. Microbiol. 2022, 13, 991144. [Google Scholar] [CrossRef]
- Jiang, X.; Jiang, C.; Yu, T.; Jiang, X.; Kang, R.; Ren, S.; Chen, H.; Zhang, Y.; Li, Y.; Meng, H.; et al. Phenyllactic acid application to control Listeria monocytogenes biofilms and its growth in milk and spiced beef. Int. J. Food Microbiol. 2022, 381, 109910. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, D.; Sun, J.; Sun, Z.; Liu, F.; Du, L.; Wang, D. Synergistic antibiofilm effects of ultrasound and phenyllactic acid against Staphylococcus aureus and Salmonella enteritidis. Foods 2021, 10, 2171. [Google Scholar] [CrossRef] [PubMed]
- Shariff, M.; Chatterjee, M.; Morris, S.D.; Paul, V.; Vasudevan, A.K.; Mohan, C.G.; Paul-Prasanth, B.; Biswas, R. Enhanced inhibition of Pseudomonas aeruginosa virulence factor production and biofilm development by sublethal concentrations of eugenol and phenyllactic acid. Lett. Appl. Microbiol. 2022, 75, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Pertusati, F.; Pileggi, E.; Richards, J.; Wootton, M.; Van Leemputte, T.; Persoons, L.; De Coster, D.; Villanueva, X.; Daelemans, D.; Steenackers, H.; et al. Drug repurposing: Phosphate prodrugs of anticancer and antiviral FDA-approved nucleosides as novel antimicrobials. J. Antimicrob. Chemother. 2020, 75, 2864–2878. [Google Scholar] [CrossRef] [PubMed]
- Vermilyea, D.M.; Ottenberg, G.K.; Davey, M.E. Citrullination mediated by PPAD constrains biofilm formation in P. gingivalis strain 381. npj Biofilms Microbiomes 2019, 5, 7. [Google Scholar] [CrossRef]
- Xiong, Z.; Zhu, X.; Geng, J.; Xu, Y.; Wu, R.; Li, C.; Fan, D.; Qin, X.; Du, Y.; Tian, Y.; et al. Intestinal Tuft-2 cells exert antimicrobial immunity via sensing bacterial metabolite N-undecanoylglycine. Immunity 2022, 55, 686–700. [Google Scholar] [CrossRef]
- Wasfi, R.; El-Rahman, O.A.A.; Zafer, M.M.; Ashour, H.M. Probiotic Lactobacillus sp. inhibit growth, biofilm formation and gene expression of caries-inducing Streptococcus mutans. J. Cell Mol. Med. 2018, 22, 1972–1983. [Google Scholar] [CrossRef]
- Choi, J.H.; Moon, C.M.; Shin, T.S.; Kim, E.K.; McDowell, A.; Jo, M.K.; Joo, Y.H.; Kim, S.E.; Jung, H.K.; Shim, K.N.; et al. Lactobacillus paracasei-derived extracellular vesicles attenuate the intestinal inflammatory response by augmenting the endoplasmic reticulum stress pathway. Exp. Mol. Med. 2020, 52, 423–437. [Google Scholar] [CrossRef]
- Ayebo, A.D.; Angelo, I.A.; Shahani, K.M. Effect of ingesting Lactobacillus acidophilus milk upon fecal flora and enzyme activity in humans. Milchwissenschaft 1980, 35, 730–733. [Google Scholar] [CrossRef]
- Jiang, Q.; Stamatova, I.; Kainulainen, V.; Korpela, R.; Meurman, J.H. Interactions between Lactobacillus rhamnosus GG and oral micro-organisms in an in vitro biofilm model. BMC Microbiol. 2016, 16, 149. [Google Scholar] [CrossRef]
- Bijle, M.N.; Neelakantan, P.; Ekambaram, M.; Lo, E.; Yiu, C. Effect of a novel synbiotic on Streptococcus mutans. Sci. Rep. 2020, 10, 7951. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yin, L.; Ramage, G.; Li, B.; Tao, Y.; Zhi, Q.; Lin, H.; Zhou, Y. Assessing the impact of curcumin on dual-species biofilms formed by Streptococcus mutans and Candida albicans. Microbiologyopen 2019, 8, e937. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Huang, Y.; Chen, M.; Hu, C.; Chen, Y. Functional synergy of antimicrobial peptides and chlorhexidine acetate against gram-negative/gram-positive bacteria and a fungus in vitro and in vivo. Infect. Drug Resist. 2019, 12, 3227–3239. [Google Scholar] [CrossRef] [PubMed]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersbøll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D. Real-time quantitative PCR. Methods 2001, 4, 383–385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, Y.; Zuo, Y.; Tang, C.; Zhou, F.; Cui, X.; Wang, L. Effects of Rhein-8-O-β-D-glucopyranoside on the biofilm formation of Streptococcus mutans. Curr. Microbiol. 2021, 78, 323–328. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Metabolite | Formula | Peak Area (%) | No. | Metabolite | Formula | Peak Area (%) |
|---|---|---|---|---|---|---|---|
| Organic acids | 7 | Arginylproline | C11H21N5O3 | 3.24 | |||
| 1 | 9,10-Epoxy-18-hydroxy-octadecanoic acid | C18H34O4 | 14.55 | 8 | 1-(D-3-Mercapto-2-methyl -1-oxopropyl)-L-proline | C9H15NO3S | 3.01 |
| 2 | 9-Hydroxylinoleic acid | C18H32O3 | 13.12 | 9 | L-beta-aspartyl-L-leucine | C10H18N2O5 | 2.93 |
| 3 | 6-Hydroxyhexanoic Acid | C6H12O3 | 12.23 | 10 | Valylserine | C8H16N2O4 | 2.43 |
| 4 | 3-Aminopentanedioic Acid | C5H9NO4 | 9.48 | 11 | Others | - | 8.58 |
| 5 | D-2-Hydroxyglutaric acid | C5H8O5 | 7.44 | Amino acid | |||
| 6 | 2-Hydroxyadipic acid | C6H10O5 | 7.36 | 1 | L-Leucine | C6H13NO2 | 32.23 |
| 7 | DL-2-hydroxy stearic acid | C18H36O3 | 5.96 | 2 | L-Lysine | C6H14N2O2 | 10.83 |
| 8 | Behenic acid | C22H44O2 | 5.74 | 3 | Citrulline | C6H13N3O3 | 9.73 |
| 9 | Phytanic acid | C20H40O2 | 4.26 | 4 | L-Carnitine | C7H15NO3 | 9.40 |
| 10 | Phenyllactic acid | C9H10O3 | 3.06 | 5 | L-Tyrosine | C9H11NO3 | 8.14 |
| 11 | Others | - | 16.80 | 6 | L-prolinamide | C5H10N2O | 4.41 |
| Nucleotides | 7 | L-4-Hydroxyglutamate semialdehyde | C5H9NO4 | 3.92 | |||
| 1 | Adenosine 3′-monophosphate | C10H14N5O7P | 28.17 | 8 | N-Undecanoylglycine | C13H25NO3 | 3.62 |
| 2 | Hypoxanthine | C5H4N4O | 15.92 | 9 | Cholylmethionine | C29H49NO6S | 3.23 |
| 3 | 5′-Methylthioadenosine | C11H15N5O3S | 11.31 | 10 | L-Glutamic Acid | C5H9NO4 | 1.65 |
| 4 | Zidovudine monophosphate | C10H14N5O7P | 3.83 | 11 | Others | - | 12.84 |
| 5 | ADP-ribose | C15H23N5O14P2 | 3.56 | Lipid | |||
| 6 | Adenosine monophosphate | C10H14N5O7P | 3.29 | 1 | PA(PGE1/20:1(11Z)) | C43H77O11P | 9.35 |
| 7 | Pseudouridine 5′-phosphate | C9H13N2O9P | 3.18 | 2 | SM(d16:2(4E,8Z)/20:5(6E, 8Z,11Z,14Z,17Z)-OH(5)) | C41H71N2O7P | 8.59 |
| 8 | Guanidylic acid (guanosine monophosphate) | C10H14N5O8P | 3.16 | 3 | PA(TXB2/22:1(13Z)) | C45H81O12P | 8.03 |
| 9 | Xanthine | C5H4N4O2 | 3.15 | 4 | PI(18:0/18:2(9Z,12Z)) | C45H83O13P | 6.88 |
| 10 | 3′,5′-Cyclic AMP | C10H12N5O6P | 3.13 | 5 | PA(TXB2/20:0) | C43H79O12P | 6.61 |
| 11 | Others | - | 21.30 | 6 | PG(a-21:0/20:4(7E,9E, 11Z,13E)3OH(5S,6R,15S)) | C47H85O13P | 6.52 |
| Terpene | 7 | PE(LTE4/20:1(11Z)) | C48H85N2O11PS | 6.33 | |||
| 1 | Glycinoeclepin B | C31H42O9 | 16.02 | 8 | PA(20:4(8Z,11Z,14Z,17Z)/PGF1alpha) | C43H73O11P | 4.98 |
| 2 | Soyasapogenol F | C30H50O3 | 14.99 | 9 | PG(i-12:0/i-19:0) | C37H73O10P | 3.36 |
| 3 | Alpha-Campholonic acid | C10H16O3 | 10.71 | 10 | PI(PGF1alpha/22:2(13Z,16Z)) | C51H91O16P | 2.55 |
| 4 | Zedoarol | C15H18O3 | 9.55 | 11 | Others | - | 36.80 |
| 5 | Geranylcitronellol | C20H36O | 7.68 | Amines | |||
| 6 | Arctiopicrin | C19H26O6 | 5.88 | 1 | N1,N12-Diacetylspermine | C14H30N4O2 | 54.59 |
| 7 | Canavalioside | C26H42O12 | 5.28 | 2 | N1,N8-Diacetylspermidine | C11H23N3O2 | 21.17 |
| 8 | Manoalide | C25H36O5 | 5.21 | 3 | Oleamide | C18H35NO | 11.07 |
| 9 | Cinncassiol C | C20H28O7 | 4.91 | 4 | N-Palmitoyl Cysteine | C14H22O4 | 2.38 |
| 10 | 3-O-cis-Coumaroylmaslinic acid | C39H54O6 | 2.88 | 5 | N1-Acetylspermine | C12H28N4O | 1.55 |
| 11 | Others | - | 16.89 | 6 | 9-Octadecenamide | C18H35NO | 1.27 |
| Peptides | 7 | N-[4-[Acetyl(3-aminopropyl)amino]butyl]-N-(3-aminopropyl)acetamide | C14H30N4O2 | 1.21 | |||
| 1 | Permetin A | C54H92N12O12 | 52.51 | 8 | 2-Phenylacetamide | C8H9NO | 0.92 |
| 2 | Prolyl-Asparagine | C9H15N3O4 | 5.82 | 9 | N-Acetylcadaverine | C7H16N2O | 0.64 |
| 3 | Sarcodon scabrosus Depsipeptide | C23H39N3O8 | 6.44 | 10 | Manumycin A | C31H38N2O7 | 0.60 |
| 4 | Cyclo(his-pro) | C11H14N4O2 | 6.01 | 11 | Others | - | 4.60 |
| 5 | Etelcalcetide | C38H73N21O10S2 | 4.91 | ||||
| 6 | Asp-Phe | C13H16N2O5 | 4.12 | ||||
| No. | Metabolite | Formula | Peak Area (%) | No. | Metabolite | Formula | Peak Area (%) |
|---|---|---|---|---|---|---|---|
| Organic acids | 10 | Gamma-Glutamylmethionine | C10H18N2O5S | 2.47 | |||
| 1 | 6-Hydroxyhexanoic Acid | C6H12O3 | 58.01 | 11 | Others | - | 20.30 |
| 2 | Phenyllactic acid | C9H10O3 | 14.12 | Amino acid | |||
| 3 | 3-Aminopentanedioic Acid | C5H9NO4 | 11.02 | 1 | N-Acetyl-DL-Leucine | C8H15NO3 | 20.23 |
| 4 | 3-(4-Hydroxyphenyl)lactate | C9H10O4 | 7.03 | 2 | N-Acetyl-DL-Phenylalanine | C11H13NO3 | 14.96 |
| 5 | 9,10-Epoxy-18-hydroxy-octadecanoic acid | C18H34O4 | 2.51 | 3 | L-Leucine | C6H13NO2 | 10.88 |
| 6 | D-2-Hydroxyglutaric acid | C5H8O5 | 2.00 | 4 | Gamma-Glutamylvaline | C10H18N2O5 | 10.31 |
| 7 | Citramalic Acid | C5H8O5 | 1.70 | 5 | Phenyl-Alanine | C9H11NO2 | 9.47 |
| 8 | acexamic acid | C8H15NO3 | 1.13 | 6 | Homocysteine | C4H9NO2S | 7.09 |
| 9 | 12-hydroxyheptadecanoic acid | C17H34O3 | 0.37 | 7 | Tyrosine lactate | C12H15NO5 | 6.87 |
| 10 | Azelaic Acid | C9H16O4 | 0.31 | 8 | L-Phenylalanine | C9H11NO2 | 5.20 |
| 11 | Others | - | 1.80 | 9 | L-Tyrosine | C9H11NO3 | 2.80 |
| Nucleotides | 10 | L-Lysine | C6H14N2O2 | 2.49 | |||
| 1 | Hypoxanthine | C5H4N4O | 58.42 | 11 | Others | - | 9.70 |
| 2 | FAPy-adenine | C5H7N5O | 15.41 | Lipid | |||
| 3 | Xanthine | C5H4N4O2 | 9.06 | 1 | PA(TXB2/22:1(13Z)) | C45H81O12P | 11.21 |
| 4 | 7-Methylguanine | C6H7N5O | 6.08 | 2 | PA(PGE1/20:1(11Z)) | C43H77O11P | 10.69 |
| 5 | Adenine | C5H5N5 | 4.01 | 3 | PG(a-21:0/20:4(7E,9E,11Z,13E)-3OH(5S,6R,15S)) | C47H85O13P | 10.22 |
| 6 | Oxypurinol | C5H4N4O2 | 1.72 | 4 | PE(LTE4/20:1(11Z)) | C48H85N2O11PS | 9.05 |
| 7 | 1-(2-Hydroxyethyloxymethyl)-6-phenyl thiothymine | C14H16N2O4S | 1.05 | 5 | PI(18:0/18:2(9Z,12Z)) | C45H83O13P | 8.81 |
| 8 | Morph | C10H16N2O4 | 0.86 | 6 | PA(TXB2/20:0) | C43H79O12P | 6.45 |
| 9 | Deoxyribose | C5H10O4 | 0.30 | 7 | SM(d16:2(4E,8Z)/20:5 (6E,8Z,11Z,14Z,17Z)-OH(5)) | C41H71N2O7P | 6.42 |
| 10 | 5-Methyldeoxycytidine | C10H15N3O4 | 0.20 | 8 | PA(20:4(8Z,11Z,14Z,17Z)/PGF1alpha) | C43H73O11P | 6.08 |
| 11 | Others | - | 2.90 | 9 | PG(i-12:0/i-19:0) | C37H73O10P | 4.04 |
| Terpene | 10 | PE(20:5(5Z,8Z,11Z,14Z,17Z)/18:2(9Z,12Z)) | C43H72NO8P | 3.63 | |||
| 1 | Fusarin C | C23H29NO7 | 57.37 | 11 | Others | - | 23.40 |
| 2 | Ineketone | C20H30O3 | 14.98 | Amines | |||
| 3 | 3-trans-p-Coumaroylrotundic acid | C39H54O7 | 13.58 | 1 | Lauryldiethanolamine | C16H35NO2 | 30.15 |
| 4 | 8-Isobutanoylneosolaniol | C23H32O9 | 7.65 | 2 | N1,N8-Diacetylspermidine | C11H23N3O2 | 28.41 |
| 5 | 4a-Methylzymosterol-4-carboxylic acid | C29H46O3 | 1.61 | 3 | N1,N12-Diacetylspermine | C14H30N4O2 | 9.58 |
| 6 | Polyporenic acid C | C31H46O4 | 0.97 | 4 | N-Acetylcadaverine | C7H16N2O | 5.22 |
| 7 | Methyl lucidenate F | C28H38O6 | 0.80 | 5 | 2-Phenylacetamide | C8H9NO | 4.16 |
| 8 | Auraptene | C19H22O3 | 0.61 | 6 | N-Cyclohexylformamide | C7H13NO | 2.47 |
| 9 | Eremanthin | C15H18O2 | 0.52 | 7 | Repaglinide aromatic amine | C22H28N2O4 | 2.15 |
| 10 | 8-Butanoylneosolaniol | C23H32O9 | 0.41 | 8 | 2-Naphthylamine | C10H9N | 2.06 |
| 11 | Others | - | 1.50 | 9 | N-[4-[Acetyl(3-aminopropyl) amino]butyl]-N-(3-aminopropyl) acetamide | C14H30N4O2 | 1.68 |
| Peptides | 10 | Fructosamine | C6H13NO5 | 1.32 | |||
| 1 | Pyro-L-glutaminyl-L-glutamine | C10H15N3O5 | 13.17 | 11 | Others | - | 12.80 |
| 2 | L-beta-aspartyl-L-leucine | C10H18N2O5 | 11.56 | ||||
| 3 | H-Gly-Arg-Gly-Asp-D-Ser-Pro-OH | C22H37N9O10 | 10.17 | ||||
| 4 | Gamma-Glutamylglutamic acid | C10H16N2O7 | 9.32 | ||||
| 5 | Mauritine A | C32H41N5O5 | 9.03 | ||||
| 6 | Isoleucyl-Valine | C11H22N2O3 | 8.44 | ||||
| 7 | Histidyltyrosine | C15H18N4O4 | 7.17 | ||||
| 8 | Glutamylisoleucine | C11H20N2O5 | 4.87 | ||||
| 9 | Bursopoietin | C14H25N7O3 | 3.50 | ||||
| Groups | Strains | Cultural Conditions |
|---|---|---|
| Model | S. mutans, PBS | Anaerobic culture at 37 °C for 24 h |
| ET-22-L | S. mutans, ET-22-L | Anaerobic culture at 37 °C for 24 h |
| ET-22-HK | S. mutans, ET-22-HK | Anaerobic culture at 37 °C for 24 h |
| ET-22-S | S. mutans, ET-22-S | Anaerobic culture at 37 °C for 24 h |
| Primers | Forward Sequences (5′–3′) | Reverse Sequences (5′–3′) | Tm (°C) | References |
|---|---|---|---|---|
| SpaP | TGATGTTGCTTCTTCTATGGAG | CAGGTTAGTGTATGTAAGCTGT | 53.95 | [24] |
| gtfB | CGAACAGCTTCTAATGGTGAAAAGCTT | TTGGCTGCATTGCTATCATCA | 55.88 | [9] |
| RecA | GCCTATGCTGCTGCTCTTG | TCACCAATATCTCCGTCAATCTC | 56.66 | [52] |
| gbpB | AGGGCAATGTACTTGGGGTG | TTTGGCCACCTTGAACACCT | 56.43 | [9] |
| gbpC | TCTGGTTTTTCTGGCGGTGT | GTCAATGCTGATGGAACGCC | 56.43 | [9] |
| gbpD | TTGACTCAGCAGCCTTTCGT | CTTCTGGTTGATAGGCGGCA | 56.43 | [9] |
| Ffh | TGGGAATGGGAGACTTGCTTA | GCTCGGAGTTAGGAGGTCAG | 57.56 | [52] |
| relA | ACAAAAAGGGTATCGTCCGTACAT | AATCACGCTTGGTATTGCTAATTG | 55.30 | [3] |
| comDE | ACAATTCCTTGAGTTCCATCCAAG | TGGTCTGCTGCCTGTTGC | 56.67 | [3] |
| ldh | CTTGATACTGCTCGTTTCCGTC | GAGTCACCATGTTCACCCAT | 56.54 | [3] |
| brpA | GGAGCAGGCCCTCGTTTATT | ATGACCTCACGTTGACGCTT | 56.43 | [9] |
| 16S RNA | CCTACGGGAGGCAGCAGTAG | CAACAGAGCTTTACGATCCGAAA | 58.77 | [9] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Wu, J.; Sun, Z.; Fan, J.; Liu, F.; Zhao, W.; Liu, W.-H.; Zhang, M.; Hung, W.-L. Postbiotics Derived from L. paracasei ET-22 Inhibit the Formation of S. mutans Biofilms and Bioactive Substances: An Analysis. Molecules 2023, 28, 1236. https://doi.org/10.3390/molecules28031236
Zhao Z, Wu J, Sun Z, Fan J, Liu F, Zhao W, Liu W-H, Zhang M, Hung W-L. Postbiotics Derived from L. paracasei ET-22 Inhibit the Formation of S. mutans Biofilms and Bioactive Substances: An Analysis. Molecules. 2023; 28(3):1236. https://doi.org/10.3390/molecules28031236
Chicago/Turabian StyleZhao, Zhi, Jianmin Wu, Zhe Sun, Jinbo Fan, Fudong Liu, Wen Zhao, Wei-Hsien Liu, Ming Zhang, and Wei-Lian Hung. 2023. "Postbiotics Derived from L. paracasei ET-22 Inhibit the Formation of S. mutans Biofilms and Bioactive Substances: An Analysis" Molecules 28, no. 3: 1236. https://doi.org/10.3390/molecules28031236
APA StyleZhao, Z., Wu, J., Sun, Z., Fan, J., Liu, F., Zhao, W., Liu, W.-H., Zhang, M., & Hung, W.-L. (2023). Postbiotics Derived from L. paracasei ET-22 Inhibit the Formation of S. mutans Biofilms and Bioactive Substances: An Analysis. Molecules, 28(3), 1236. https://doi.org/10.3390/molecules28031236