Abstract
The aim of the present study was the phytochemical analysis of the aerial parts of Heliotropium procumbens Mill., a herb from Boraginaceae plant family not previously studied. The methanol (ME) and aqueous extracts (WE) of the aerial parts were assayed for their total phenolic and flavonoid content and antioxidant properties, using free radical scavenging (DPPH, ABTS), reducing power (FRAP, CUPRAC), phosphomolybdenum and metal chelating assays. The extracts displayed considerable free radical scavenging activity against DPPH and ABTS radicals, with potential values of 46.88 and 68.31 mg TE/g extract for ME, and 93.43 and 131.48 mg TE/g extract for WE, respectively. Key clinical enzymes involved in neurodegenerative diseases AChE and BChE, diabetes (α-amylase and α-glucosidase) and skin whitening (tyrosinase) were also assayed. The phytochemical profile of the studied species was determined through UHPLC–HRMS, whereby 26 secondary metabolites were identified, three of which (luteolin-7-glucoside, lithospermic and rosmarinic acids) were isolated and structurally determined by NMR spectral means. H. procubens was found to harbor bioactive metabolites and could, hence, serve as a source of biological activities which could be further explored and exploited for potential applications.
1. Introduction
The Boraginaceae plant family comprises 156 genera and 2650 species of herbs, shrubs, and trees, with a worldwide distribution, occurring mainly in Europe, Asia and North America [1]. Heliotropium, Arnebia, Martensia, Cordia, and Trichedesma are the main genera of this family, with therapeutic effects mostly based on the presence of phenolic metabolites [2,3,4]. Moreover, almost all genera of the family Boraginaceae produce and serve as a natural source of pyrrolizidine alkaloids, which are related with serious health problems [4,5].
Heliotropium (Boraginaceae) is a genus of herbs, distributed in tropical and temperate regions of the world comprising approximately 300 species commonly known as heliotropes. The name “heliotrope” derives from the fact that these plants turn their leaves to the direction of the sun and, as a result, the name includes the word “helios”, a Greek word for “sun”, and “tropein” which means “to turn”. Heliotropes [5] also exhibit several therapeutic properties, such as antimicrobial, anti-inflammatory, analgesic, healing, etc., and they have been used in folk medicine against rheumatism, menstrual dysfunction, billiary disorders, and noxious bites [6,7,8]. The potential active secondary metabolites from the Heliotropium species comprise phenolic compounds (phenolic acids, flavonoids, quinones: alkannins–shikonins), terpenoids and pyrrolizidine alkaloids (PAs).
This study has been focused on the species Heliotropium procumbens Mill., a herb with white flowers, native to America, as well as East to West Indies, is used by locals in animal feed, as it is considered to exhibit a high nutritional value with a notably high protein content [9]. H. procumbens occurs in the USA Gulf Coastal Region (Texas, Mississippi, and Louisiana), and it has been questioned whether it is a native or adventive species in the area. After comparing the time and location of the various Texan specimens assembled over the past 150 years, it has been concluded that H. procumbens is native to the area under consideration [10].
In the framework of our research on Boraginaceae plants [3,4,11,12,13], we herein present the phytochemical study on the aerial parts of H. procumbens from Panama in order to evaluate its chemical composition, the total phenolic and flavonoid content, antioxidant activity, inhibitory potentials against key clinical enzymes involved in neurodegenerative diseases (cholinesterases AChE, BChE), in skin whitening (tyrosinase), and in diabetes (α-amylase, α-glucosidase).
2. Results
2.1. Phytochemical Analysis
Through our chemical analysis of secondary metabolites, characteristics of the Heliotropium genus, as well as of the whole plant family of Boraginaceae, have been identified and further examined. Several among them contributed to the exerted biological properties, such as the caffeic derivatives, rosmarinic acid (RA), and lithospermic acid together with the flavone luteolin-7-O-glucoside [14]. All three of them have been isolated and structurally determined through NMR.
Furthermore, through UHPLC–HRMS, 26 secondary metabolites have been identified (Figure 1, Table 1, Supplementary Materials).
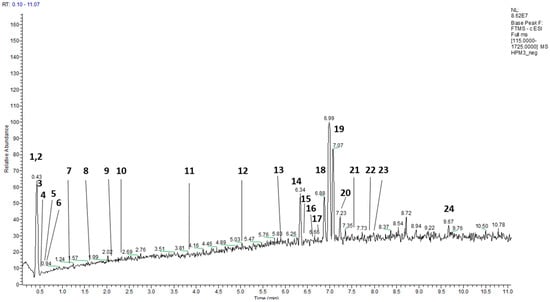
Figure 1.
Characterization of the phenolic constituents of Heliotropium procubens extract by UHPLC–MS.

Table 1.
Secondary metabolites identified from methanolic extract and fractions through UHPLC–HRMS.
2.2. Total Phenolic (TPC)/Flavonoid (TFC) Content and Antioxidant Activity Results
The antioxidative capacity of the aerial parts from H. procumbens extracts was determined using different assays (DPPH•, ABTS•+, CUPRAC, FRAP), phosphomolybdenum and ferrous ion chelating test, as well as the TPC and TFC which are correlated with the antioxidant activity. The results (Table 2) show a high TPC (32.20 mg GAE/g for ME and 53.47 mg GAE/g for WE) and a lower TFC (0.70 mg REs/g extract for ME and 12.39 mg REs/g extract for WE), assuming that this plant is a rich source of phenolics, which is in accordance with the presented phytochemical profile.
Table 2.
Antioxidant properties of H. procumbens.
The DPPH and ABTS assays on the H. procumbens methanolic extract showed good inhibition of the free radicals (46.88 ± 0.64 inhibition for the DPPH assay and 68.31 ± 0.69 for the ABTS assay) and, therefore, a significant antioxidative profile, while the water extract showed a higher scavenging capacity (93.43 ± 0.11 for the DPPH assay and 131.48 ± 6.66 for the ABTS assay).
2.3. Enzyme Inhibitory Activity
The results obtained from enzyme inhibitory assays from H. procumbens extracts are summarized in Table 3. Regarding the acetylocholinesterase inhibition, the methanolic and aqueous extracts did not exhibit any activity (0.48 ± 0.04 mg GALAEs/g extract for WE), while no inhibition was detected in the case of the butyrylcholinesterases assay (1.17 ± 0.25 mg GALAEs/g extract for ME).
Table 3.
Enzyme inhibitory effects of H. procumbens.
Regarding the inhibitors, which control glucose level in blood, the assayed extracts exhibited higher α-glucosidase inhibitory activity (1.97 ± 0.14 mmol ACAE/g extract ME and 2.08 ± 0.01 mmol ACAE/g extract WE) compared to α-amylase inhibitory activity (0.22 ± 0.01 mmol ACAE/g extract ME and 0.07 ± 0.01 mmol ACAE/g extract WE) but both are not valuable.
In addition, ME showed higher inhibitory capacity against tyrosinase, an enzyme involved in skin whitening, (25.05 ± 0.17 mg KAE/g extract) in comparison with the WE (8.82 ± 0.80 mg KAE/g extract).
3. Discussion
From the studied H. procumbens, 26 secondary metabolites have been presented so far. Several coumaric and caffeic acid derivatives have been revealed, previously identified in other Heliotropium species as H. lasiocarpium, H. suaveolens [17] and H. strigosum [18], and they have been strongly related with the plant’s antioxidant properties. Syringic acid has been also previously identified in H. curassavicum [19] and H. strigosum [18]. Isobergapten, among coumarins, has been identified previously in several heliotropes, among which are H. lasiocarpium, H. suaveolens and H. crispum [17]. Methyl-catechin has never been identified in the genus, while catechin has been found in H. strigosum [27], H. crispum [28] and H. curassavicum [19]. Dihydroxy methoxy benzoic acid (pyruvic acid) has been previously identified in H. crispum through UHPLC-Q-TOF-MS together with other benzoic acids’ derivatives and/or substituted hydroxy-methoxy benzoic acids [17]. Vanillic acid has been previously found in H. thermophilum [29] and H. strigosum [18]. Ferulic acid is a coumaric derivative previously reported as either iso, cis or trans-ferulic acid in H. crispum [17] and H. strigosum [18], not easily distinguished due to common fragmentation between the isomers. Among the most abundant metabolites in the studied heliotrope species were the caffeic acid derivatives, RA and lithospermic acid, together with the flavone luteolin-7-glucoside which were identified but also isolated and structurally determined.
The identified by UHPLC–HRMS caffeic derivatives in this study were caffeic acid itself, caffeoyl hexoside, RA and its methyl ester, lithospermic acid and salvianolic acid B.
RA, a polyphenolic compound, is structurally an ester of caffeic acid and is derived from hydroxycinnamic acid, widespread and well-known for its multitude of bioactivities, such as antioxidant, antiviral, antimicrobial and anti-inflammatory. It possesses mainly neuroprotective, anti-acetyl cholinesterase and hepatoprotective properties, and is rapidly eliminated from human and rat blood circulation after per os administration, metabolized predominantly to caffeic, coumaric and ferulic acids. According to the results of previous phytochemical studies, RA is considered as a chemotaxonomic marker among Boraginaceae taxa [11,12,13,14], while it has been also isolated from H. foertherianum [30] and H. angiospermum [31], together with ethyl lithosprmate in the latter.
Moreover, caffeic acid has very recently been identified in H. ramosissimum [5], while the taxonomic significance of neither RA nor caffeic acid in heliotropes and the Boraginacae plant family has been referred to and/or explained in the existing overview for heliotropes [5].
Within heliotropes, flavonoids are a common class of secondary metabolites, among which several express strong bioactivities. In the existing literature, several research surveys have characterized and reported various flavonoids, such as 5,4′-dihydroxy-7-methoxyflavanone, 4′-acetyl-5-hydroxy-7-methoxyflavanone, methoxy-3-[7′-methyl-3′-hydroxymethyl-2′,6′-octadienyl]phenol (in H. glutinosum), 5,3′-dihydroxy-7,4′-dimethoxyflavanone, 7-O-methyleriodictyol, 3-O-methylgalangin, filifolinol, filifolinyl senecionate (in H. filifolium), naringenin, filifolinoic acid, filifolinone, 3-oxo-2-arylbenzofuran (in H. taltalense, H. sclerocarpum), dihydroquercetine, quercetin (H. strigosum) [5].
In the present study, in H. procubens, the flavones luteolin-7-O-glucoside and trihydroxy-flavone hexoside, the flavanone naringenin-7-O-glucoside, as well as the flavonols kaempferol and quercetine-7-O-rutinoside (rutin) have been identified. Among them, luteolin-7-glucoside is a well-known metabolite isolated before from H. tenellum [32], but it is unfortunately not included in the phytochemical overview of Heliotropium genus [5]. Furthermore, luteolin-8-C-glucoside (orientin) has also been also reported for H. ramosissimum [16].
Additionally, naringenin-7-O-glucoside, a trivial flavonoid, has been previously identified in many heliotropes such as H. taltalense Phil., H. sclerocarpum Phil. [8], H. sinuatum [6,33], H. glutinosum Phil [7], H. filifolium and H. huascoense [5], together with the flavanone chrysin and sakuranetin in H ellipticum [34,35]
Among flavonols, kaempferol and quercetin (the aglycon of rutin) and myricetin have been reported in H. crispum [36] so far.
In H. crispum, among other long chain carbon fatty acids, 5,8,12-trihydroxy-9-octadecenoic acid has been identified [17]. This chemical structure is commonly found in several plants of the Boraginaceae family as the basic molecule of octadecanoic acid (stearic acid), and different substitutions with hydroxyl groups have been determined in several plant species of the family, such as Anchusa strigosa, Phyllocara aucheri, Cynoglottis barrelieri [13].
There is a large demand from the market for natural extracts with antioxidant properties, especially from a plant origin, as many side effects caused by synthetic compounds have been reported to affect human health [37]. The high inhibition shown by the DPPH and ABTS assays is most likely related to the high phenolic content, as the identified phenolic compounds are extremely polar, containing multiple hydroxyl groups known for their antioxidative capacity. The mean values of the antioxidant potential of the extract in the ABTS assay were higher (68.31 ± 0.69 and 131.48 ± 6.66) compared to the DPPH assay (46.88 ± 0.64 and 93.43 ± 0.11). These results are explained by the difference in the mechanism of the radical interaction of these two assays, due to the fact that ABTS acts as both a hydrophobic and hydrophilic system, while DPPH acts only as a hydrophobic system. A connection between antioxidant activity and phenolic content has already been established for plants of the Boraginaceae family [12]. It is linked that much of the plant family’s biological effects (mostly antioxidant and anti-inflammatory) are relative to the concentration of phenolic compounds in such plants.
Regarding the enzyme inhibition, it is noteworthy that the methanolic extract of the plant showed moderate inhibitory capacity against tyrosinase, an enzyme involved in skin whitening.
4. Materials and Methods
4.1. Plant Material
The plant material was collected by Prof. M. Gupta and identified by M. Correa, Headmistress at the Herbarium of Panama University. The characteristics of the plant are plant number: 6476; family: Boraginaceae; name: Heliotropium procumbens Mill.; part of the plant: aerial part; collection habitat: Rio Grande; area: Cerca del Puente; amount: 1041 g.
4.2. Extraction
The air-dried aerial parts of H. procumbens Mill. (900 g) were successively extracted in methanol (550 g) for 24 h at room temperature (3 × 0.3 L) and the rest (450 g) were extracted with water. The extracts were evaporated under reduced pressure to dryness, to afford 14.8 g of methanolic extract and 12.7 g of aqueous extract.
4.3. Isolation of Phenolic Compounds
For the isolation of secondary metabolites, methanol extract (5 g) was fractioned on microcrystalline cellulose (20–160 μm, Merck) column chromatography with solvents (HPLC grade) of increasing polarity (saturated in H2O), gradient of cyclohexane(c-hex)/ethyl acetate (EtOAc) from 100:0 to 0:100, followed by EtOAc/MeOH (99:1 to 80:20) to yield 46 fractions (C1–C46).
Fraction C8 (267 mg) was further subjected to column chromatography with Sephadex LH-20 (25–100 μm, Pharmacia) as the stationary phase and MeOH (HPLC grade) 100% as the mobile phase to isolate, and purify rosmarinic acid (21 mg), which was further identified through NMR (chloroform-d) and compared with bibliographic data [38]. The fractions C12 (48 mg) and C26 (100 mg) were lithospermic acid A and luteolin-7-O-β-D-glucoside, respectively. Both the compounds were structurally elucidated by NMR (chloroform-d) and a comparison of their spectroscopic data was made with those reported in the literature [39,40].
In order to evaluate the phenolic compounds through UHPLC–HRMS analysis, 1.9 g of the methanolic extract was subjected to chromatographic separation via open column chromatography with Sephadex LH-20 and MeOH as the mobile phase to give 55 fractions (HC.S1-HC.S55). The fractions HC.S8, HC.S10, HC.S17 and HC.S20, as well as the crude methanolic extract, were analyzed through UHPLC–HRMS analysis.
4.4. UHPLC–HRMS Analysis of Phenolic Compounds
Characterization of the phytochemical profile of methanolic extract was performed using ultra-high-performance liquid chromatography–mass spectrometry (UHPLC–HRMS).
The UHPLC was performed employing a Vanquish UHPLC system (Thermo Scientific, Bremen, Germany) equipped with a binary pump, an autosampler, an online vacuum degasser, and a temperature-controlled column compartment. LC-MS grade methanol (MeOH) and formic acid (FA) were purchased from Fisher Scientific (Fisher Optima, UK) and LC-MS water was produced from a Barnstead MicroPure Water Purification System (Thermo Scientific, Germany). An Accucore Vanquish UPLC C18 (2.1 × 50 mm, 1.5 μm) reversed-phased column (Thermo Scientific, Germany) was used for the analysis. High-resolution mass spectrometry was performed on a Orbitrap Exactive Plus mass spectrometer (Thermo Scientific, Germany). Samples were injected at a concentration of 100 ppm diluted in MeOH/H2O 50:50. The mobile phase consisted of solvents A—aqueous 0.1% (v/v) FA, and B—acetonitrile. Different gradient elutions were performed for positive and negative ion mode detection and after optimization of the chromatography, the gradient applied was t = 0 min, 5% B; t = 3 min, 5% B; t = 21 min, 95% B; t = 26 min, 95% B. The flow rate was 0.3 mL/min and the injection volume was 5 μL. The column temperature was kept at 40 °C while the sample tray temperature was set at 10 °C. The ionization was performed at HESI, both in positive and negative mode. The conditions for HRMS for both negative and positive ionization modes were set as follows: capillary temperature, 320 °C; spray voltage, 2.7 kV; S-lense Rf level, 50 V; sheath gas flow, 40 arb. units; aux gas flow, 8 arb. units; aux. gas heater temperature, 50 °C. Analysis was performed using the Fourier transform mass spectrometry mode (FTMS) in the full scan ion mode, applying a resolution of 70,000, while the acquisition of mass spectra was performed in every case using the centroid mode. The data dependent acquisition capability was also used at 35,000 resolution, allowing for MS/MS fragmentation of the three most intense ions of every peak exceeding the predefined threshold, applying a 10 s dynamic exclusion. Normalized collision energy was set at 35. Data acquisition and analysis were undertaken by employing Xcalibur 2.1.
4.5. Total Phenolic Content (TPC)
The TPC of the extracts was calculated using the Folin–Ciocalteu method. Quantification was achieved by using a gallic acid reference curve. All measurements were conducted three times. Results were expressed as mg of gallic acid equivalents per gr of extract. UV/Vis absorption values at 765 nm were obtained using an Infinite M200 PRO TECAN reader (Tecan Group, Männedorf, Switzerland) [12].
4.6. Total Flavonoid Content (TFC)
The TFC of the extracts was calculated by means of an AlCl3 colorimetric assay. The results were obtained from a reference calibration curve of quercetin and expressed as mg of quercetin equivalents per g of extract. UV/Vis absorption values at 415 nm were obtained using an Infinite M200 PRO TECAN reader (Tecan Group, Männedorf, Switzerland) [12].
4.7. Radical Scavenging Activity
4.7.1. 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) Diammonium Salt (ABTS) Assay
The free radical-scavenging activity of the extracts was determined by a ABTS radical cation (ABTS+) decolorization assay [12]. The sample absorbance was read at 734 nm. Trolox was used as a positive control and the radical scavenging activity of the tested extracts was expressed as mg of Trolox equivalents (mg TE/g extract).
4.7.2. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Assay
For the DPPH• assay, discoloration of the extract solution was measured to determine the antioxidant activity thereof [12]. The sample absorbance was read at 517 nm. Trolox was used as a positive control and the radical scavenging activity of the tested extracts was expressed as mg of Trolox equivalents (mg TE/g extract).
4.8. Reducing Power FRAP and CUPRAC Assays
FRAP (ferric reducing antioxidant power) and CUPRAC (cupric ion reducing antioxidant capacity) assays were used to determine the reductive activity of the extracts. Experimental procedures were undertaken as described previously [12] and reductive capability was expressed as mg of Trolox equivalents (mg TE/g extract) for both assays.
4.9. Total Antioxidant Capacity and Metal Chelating Activity
The total antioxidant activity of both extracts was evaluated by the phosphomolybdenum method [12]. Trolox was used as a positive control and the results were expressed as millimoles of Trolox equivalents (mmol TE/g extract). Metal chelating activity was determined, EDTA was used as a positive control and the results were expressed as milligrams of EDTA equivalents (mg EDTAE/g extract).
4.10. Enzyme Inhibitory Activities
The ability of the methanol and aqueous extracts to inhibit the activity of selective enzymes was measured [12]. Cholinesterases (acetylcholinesterase (AChE) and butyrylcholinesterase (BChE)) inhibitory activity was expressed as mg of galanthamine equivalents (mg GALAE/g extract), tyrosinase inhibition as mg of kojic acid equivalents (mg KAE/g extract), while for α-amylase and α-glucosidase the results were expressed as millimoles of acarbose equivalents (mmol ACAE/g extract).
4.11. Expression of Results
All analyses were performed in triplicate. The results were expressed as mean values and standard deviation (SD). The differences in the extracts were investigated by using the Student t-test (p < 0.05) and this test was performed in Xlstat 2018.
5. Conclusions
The presented study is the first record on Heliotropium procumbens and it is focused on the determination of the phenolic profile of the plant as well as the evaluation of in vitro antioxidative, neuroprotective and antidiabetic effects. It is noteworthy that the taxonomic significance of the reported phenolic secondary metabolites and especially the caffeic derivatives (mostly caffeic acid and RA) has been proved, as well as the most common flavonoids, and their potential contribution to the expressed bioactivities. The importance of exploiting natural antioxidants of herbal origin has increased significantly at present due to the fact that synthetic antioxidants have been suspected to express adverse reactions to human health [14]. According to the results, Heliotropium procumbens showed a rich phenolic profile, which is related to its strong antioxidant activity as determined by TPC, DPPH and ABTS assays, as well as moderate tyrosinase inhibition, which is also of high interest towards required systematic procedures necessary to rich extracts in various active secondary metabolites for further safe exploitation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/molecules28031008/s1, Photo 1: Heliotropium procubens Mill.; Figure S1: Characterization of the phenolic constituents of Heliotropium procubens extract by UHPLC-MS; Figure S2: UHPLC-MS chromatogram of Heliotropium procubens extract (HPM3, top chromatogram) and fractions (HP8, HP10, HP17, HP20); Figure S3: Spectra of 3: Left: Mass spectrum of 3 (m/z = 215), Right: MS2 spectrum of 3 (at m/z = 215); Figure S4: Spectra of 4: Left: Mass spectrum of 4 (m/z = 197), Right: MS2 spectrum of 4 (at m/z = 197); Figure S5: Spectra of 5: Left: Mass spectrum of 5 (m/z = 153), Right: MS2 spectrum of 5 (at m/z = 153); Figure S6: Spectra of 6: Left: Mass spectrum of 6 (m/z = 321), Right: MS2 spectrum of 6 (at m/z = 321); Figure S7: Spectra of 7: Left: Mass spectrum of 7 (m/z = 137), Right: MS2 spectrum of 7 (at m/z = 137); Figure S8: Spectra of 8: Left: Mass spectrum of 8 (m/z = 183), Right: MS2 spectrum of 8 (at m/z = 183); Figure S9: Spectra of 9: Left: Mass spectrum of 9 (m/z = 179), Right: MS2 spectrum of 9 (at m/z = 179); Figure S10: Spectra of 10: Left: Mass spectrum of 10 (m/z = 329), Right: MS2 spectrum of 10 (at m/z = 329); Figure S11: Spectra of 11: Left: Mass spectrum of 11 (m/z = 305), Right: MS2 spectrum of 11 (at m/z = 305); Figure S12: Spectra of 12: Left: Mass spectrum of 12 (m/z = 341), Right: MS2 spectrum of 12 (at m/z = 341); Figure S13: Spectra of 13: Left: Mass spectrum of 13 (m/z = 609), Right: MS2 spectrum of 13 (at m/z = 609); Figure S14: Spectra of 14: Left: Mass spectrum of 14 (m/z = 447), Right: MS2 spectrum of 14 (at m/z = 447); Figure S15: Spectra of 15: Left: Mass spectrum of 15 (m/z = 479), Right: MS2 spectrum of 15 (at m/z = 479); Figure S16: Spectra of 16: Left: Mass spectrum of 16 (m/z = 303), Right: MS2 spectrum of 16 (at m/z = 303); Figure S17: Spectra of 17: Left: Mass spectrum of 17 (m/z = 193), Right: MS2 spectrum of 17 (at m/z = 193); Figure S18: Spectra of 18: Left: Mass spectrum of 18 (m/z = 431), Right: MS2 spectrum of 18 (at m/z = 431); Figure S19: Spectra of 19: Left: Mass spectrum of 19 (m/z = 359), Right: MS2 spectrum of 19 (at m/z = 359); Figure S20: Spectra of 20: Left: Mass spectrum of 20 (m/z = 537), Right: MS2 spectrum of 20 (at m/z = 537); Figure S21: Spectra of 21: Left: Mass spectrum of 21 (m/z = 717), Right: MS2 spectrum of 21 (at m/z = 717); Figure S22: Spectra of 22: Left: Mass spectrum of 22 (m/z = 373), Right: MS2 spectrum of 22 (at m/z = 373); Figure S23: Spectra of 23: Left: Mass spectrum of 23 (m/z = 285), Right: MS2 spectrum of 23 (at m/z = 285); Figure S24: Spectra of 24: Left: Mass spectrum of 24 (m/z = 329), Right: MS2 spectrum of 24 (at m/z = 329); Figure S25: The structure and the main fragments of the isolated compounds.
Author Contributions
Conceptualization, I.C.; methodology, A.C., G.Z. and K.G.; formal analysis, K.-M.O.-P., A.C., G.Z., K.G., C.G. and I.C.; investigation, K.-M.O.-P., G.Z. and G.-A.K.; resources, K.-M.O.-P., A.C., G.Z. and G.-A.K.; data curation, K.-M.O.-P., A.C., G.Z., K.G., C.G. and I.C.; writing—original draft preparation, K.-M.O.-P. and I.C.; writing—review and editing, A.C., K.G. and I.C.; visualization, I.C.; supervision, I.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank the late deceased Emeritus Researcher Mahabir Gupta for the plant collection and the botanist Mireya Correa, Headmistress of the Herbarium of the University of Panama, for the species identification.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Da Silva, S.A.S.; Agra, M.d.F.; Tavares, J.F.; da-Cunha, E.V.L.; Barbosa-Filho, J.M.; Silva, M.S.d. Flavanones from aerial parts of Cordia globosa (Jacq.) Kunth, Boraginaceae. Rev. Bras. Farmacogn. 2010, 20, 682–685. [Google Scholar] [CrossRef]
- Dresler, S.; Szymczak, G.; Wójcik, M. Comparison of some secondary metabolite content in the seventeen species of the Boraginaceae family. Pharm. Biol. 2017, 55, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Orfanou, I.M.; Damianakos, H.; Bazos, I.; Graikou, K.; Chinou, I. Pyrrolizidine Alkaloids from Onosma kaheirei Teppner (Boraginaceae). Rec. Nat. Prod. 2016, 10, 221. [Google Scholar]
- Damianakos, H.; Jeziorek, M.; Sykłowska-Baranek, K.; Buchwald, W.; Pietrosiuk, A.; Chinou, I. Pyrrolizidine alkaloids from Cynoglossum columnae Ten. (Boraginaceae). Phytochem. Lett. 2016, 15, 234–237. [Google Scholar] [CrossRef]
- Fayed, M.A.A. Heliotropium; a genus rich in pyrrolizidine alkaloids: A systematic review following its phytochemistry and pharmacology. Phytomedicine Plus 2021, 1, 100036. [Google Scholar] [CrossRef]
- Goyal, N.; Sharma, K.S. Bioactive phytoconstituents and plant extracts from genus Heliotropium. Int. J. Green Pharm. 2014, 8, 217. [Google Scholar] [CrossRef]
- Modak, B.; Rojas, M.; Torres, R.; Rodilla, J.; Luebert, F. Antioxidant Activity Of A New Aromatic Geranyl Derivative of The Resinous Exudates From Heliotropium glutinosum Phil. Molecules 2007, 12, 1057–1063. [Google Scholar] [CrossRef]
- Ghori, M.K.; Ghaffari, M.A.; Nawaz, S.; Manzoor, M.; Aziz, M.; Sarwer, W. Ethnopharmacological, Phytochemical and Pharmacognostic Potential of Genus Heliotropium L. Turk. J. Pharm. Sci. 2016, 13, 143–168. [Google Scholar] [CrossRef]
- Nunes, A.T.; Cabral, D.L.V.; Amorim, E.L.C.; Santos, M.V.E.; dos Albuquerque, U.P. Plants used to feed ruminants in semi-arid Brazil: A study of nutritional composition guided by local ecological knowledge. J. Arid. Environ. 2016, 135, 96–103. [Google Scholar] [CrossRef]
- Turner, B.L.; Williams, J. Heliotropium procumbens (Boraginaceae): Native or Adventive in the USA. Phytologia 2010, 92, 2. [Google Scholar]
- Ganos, C.; Aligiannis, N.; Chinou, I.; Naziris, N.; Chountoulesi, M.; Mroczek, T.; Graikou, K. Rindera graeca (Boraginaceae) phytochemical profile and biological activities. Molecules 2020, 25, 3625. [Google Scholar] [CrossRef] [PubMed]
- Varvouni, E.-F.; Zengin, G.; Graikou, K.; Ganos, C.; Mroczek, T.; Chinou, I. Phytochemical analysis and biological evaluation of the aerial parts from Symphytum anatolicum Boiss. and Cynoglottis Barrelieri (All.) Vural & Kit Tan (Boraginaceae). Biochem. Syst. Ecol. 2020, 92, 104128. [Google Scholar] [CrossRef]
- Varvouni, E.-F.; Zengin, G.; Graikou, K.; Ganos, C.; Mroczek, T.; Chinou, I. Chemical profile and biological properties of the endemic Turkish species Phyllocara aucheri. S. Afr. J. Bot. 2021, 137, 340–344. [Google Scholar] [CrossRef]
- Graikou, K.; Damianakos, H.; Ganos, C.; Sykłowska-Baranek, K.; Jeziorek, M.; Pietrosiuk, A.; Roussakis, C.; Chinou, I. Chemical Profile and Screening of Bioactive Metabolites of Rindera graeca (A. DC.) Bois. & Heldr. (Boraginaceae) In Vitro Cultures. Plants 2021, 10, 834. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.J.; Serró, R.F. Isolation and identification of O-α-d-galactopyranosyl-myo-inositol and of myo-inositol from juice of the sugar beet (Beta vulgaris). J. Am. Chem. Soc. 1953, 75, 1040–1042. [Google Scholar] [CrossRef]
- Fayed, M.A.A.; Abouelela, M.E.; Refaey, M.S. Heliotropium ramosissimum metabolic profiling, in silico and in vitro evaluation with potent selective cytotoxicity against colorectal carcinoma. Sci. Rep. 2022, 12, 12539. [Google Scholar] [CrossRef] [PubMed]
- Arshad, A.; Ahemad, S.; Saleem, H.; Saleem, M.; Zengin, G.; Abdallah, H.H.; Tousif, M.I.; Ahemad, N.; Fawzi Mahomoodally, M. RP-UHPLC-MS Chemical Profiling, Biological and in Silico Docking studies to Unravel the therapeutic potential of Heliotropium crispum Desf. As A Novel Source of Neuroprotective Bioactive Compounds. Biomolecules 2021, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, A.; Sarfraz, R.A.; Ashraf, A.; Adil, S. Phenolic composition and biological (anti diabetic and antioxidant) activities of different solvent extracts of an endemic plant (Heliotropium strigosum). J. Chil. Chem. Soc. 2016, 61, 2903–2906. [Google Scholar] [CrossRef]
- Abd-Elgawad, A.M.; Elshamy, A.I.; Al-Rowaily, S.L.; El-Amier, Y.A. Habitat affects the chemical profile, allelopathy, and antioxidant properties of essential oils and phenolic enriched extracts of the invasive plant Heliotropium curassavicum. Plants 2019, 8, 482. [Google Scholar] [CrossRef]
- Gören, A.C.; Tümen, G.; Çelik, A.; Çıkrıkçı, S. Fatty acid composition of Heliotropium species (Boraginaceae): A first chemical report on the new species H. thermophilum. Nat. Prod. Commun. 2008, 3, 1934578X0800301030. [Google Scholar] [CrossRef]
- FoodB. Available online: www.foodb.com (accessed on 10 October 2022).
- He, J.; Feng, Y.; Ouyang, H.; Yu, B.; Chang, Y.; Pan, G.; Dong, G.; Wang, T.; Gao, X. A Sensitive LC–MS/MS method for simultaneous determination of six flavonoids in rat plasma: Application to a pharmacokinetic study of total flavonoids from Mulberry leaves. J. Pharm. Biomed. Anal. 2013, 84, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; He, W.; Xiang, C.; Han, J.; Wu, L.; Guo, D.; Ye, M. Qualitative and quantitative analyses of flavonoids in spirodela polyrrhiza by high-performance liquid chromatography coupled with Mass Spectrometry: Chemical Analysis for Spirodela polyrrhiza by LC/MS. Phytochem. Anal. 2011, 22, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Liu, H.; Lin, L. Simultaneous determination of vitexin and isovitexin in rat plasma after oral administration of Santalum album L. leaves extract by liquid chromatography tandem mass spectrometry: Simultaneous determination of vitexin and isovitexin. Biomed. Chromatogr. 2013, 27, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Metlin. Available online: www.metlin.com (accessed on 10 October 2022).
- Wang, X.; Qian, Y.; Li, X.; Jia, X.; Yan, Z.; Han, M.; Qiao, M.; Ma, X.; Chu, Y.; Zhou, S.; et al. Rapid determination of rosmarinic acid and its two bioactive metabolites in the plasma of rats by LC–MS/MS and application to a pharmacokinetics study. Biomed. Chromatogr. 2021, 35, e4984. [Google Scholar] [CrossRef]
- Ghori, K.M.; Chaudhry, A.B.; Uzair, M.; Janbaz, K.; Sarwar, W.; Manzoor, M.; Hussain, S.; Qaisar, M.N.; Ghori, U.M. Antiglycation and Insecticidal Potential of Heliotropium strigosum Willd. J. Nat. Prod. Plant Res. 2016, 6, 1–7. [Google Scholar]
- Shahbaz, A.; Abbasi, B.A.; Iqbal, J.; Fatima, I.; Anber Zahra, S.; Kanwal, S.; Devkota, H.P.; Capasso, R.; Ahmad, A.; Mahmood, T. Chemical Composition of Gastrocotyle Hispida (Forssk.) Bunge and Heliotropium crispum Desf. and evaluation of their multiple in vitro biological potentials. Saudi J. Biol. Sci. 2021, 28, 6086–6096. [Google Scholar] [CrossRef] [PubMed]
- Yıldırım, S.; Kadıoğlu, A.; Sağlam, A.; Yaşar, A. Determination of phenolic acids and rutin in Heliotropium thermophilum by High-Performance Liquid Chromatography with Photodiode Array Detection. Instrum. Sci. Technol. 2017, 45, 35–48. [Google Scholar] [CrossRef]
- Rossi, F.; Jullian, V.; Pawlowiez, R.; Kumar-Roiné, S.; Haddad, M.; Darius, H.T. Protective effect of Heliotropium foertherianum (Boraginaceae) folk remedy and its active compound, rosmarinic acid, against a Pacific ciguatoxin. J. Ethnopharmacol. 2012, 143, 33–40. [Google Scholar] [CrossRef]
- dos Santos, G.L.D.; de Araújo, D.I.A.F.; Raimundo e Silva, J.P.; do Nascimento, Y.M.; de Souza, T.A.; Opretzka, L.C.F.; Villarreal, C.F.; Abreu, L.S.; dos Santos Junior, F.M.; de Melo, J.I.M.; et al. Sucrose Diester of Aryldihydronaphthalene-Type Lignan with Anti-inflammatory Activity from Heliotropium angiospermum. Rev. Brasil Farmacognosia 2022, 32, 734–740. [Google Scholar] [CrossRef]
- Kagan, J. Luteolin 7-glucoside, the flavonoid pigment of Heliotropium tenellum. Phytochemistry 1968, 7, 505–506. [Google Scholar] [CrossRef]
- Modak, B.; Torres, R.; Lissi, E.; Monache, F.D. Antioxidant capacity of flavonoids and a new aryl-phenol of the resinous exudate from Heliotropium sinuatum. Nat. Prod. Res. 2003, 17, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Sahu, P.M.; Sharma, R.A. In vitro and in vivo Evaluation of Flavonoids from Heliotropium ellipticum Exudate for Antioxidant, Antineoplastic and Cytotoxic Activities II Indian. J. Pharm. Sci. 2017, 79, 939–947. [Google Scholar]
- Singh, B.; Sharma, R.A. Anti-Inflammatory and Antimicrobial Effects of Flavonoids from Heliotropium ellipticum Exudate. Curr. Bioact. Compnds 2016, 12, 123–131. [Google Scholar] [CrossRef]
- Fatima, S.F.; Ishtiaq, S.; Lashkar, M.O.; Ashour, M.L.; Elhady, S.S. Metabolic Profiling of Heliotropium crispum Aerial Parts Using HPLC and FTIR and In Vivo Evaluation of Its Anti-Ulcer Activity Using an Ethanol Induced Acute Gastric Ulcer Model. Metabolites 2022, 12, 750. [Google Scholar] [CrossRef] [PubMed]
- Sarikurkcu, C.; Zengin, G.; Aktumsek, A.; Ceylan, O.; Uysal, S. Screening of Possible In Vitro Neuroprotective, Skin Care, Antihyperglycemic, and Antioxidative Effects of Anchusa undulata L. subsp. hybrida (Ten.) Coutinho from Turkey and Its Fatty Acid Profile. Int. J. Food Prop. 2015, 18, 1491–1504. [Google Scholar] [CrossRef]
- Lu, Y.; Foo, L.Y. Rosmarinic acid derivatives from Salvia officinalis. Phytochemistry 1999, 51, 91–94. [Google Scholar] [CrossRef]
- Murata, T.; Oyama, K.; Fujiyama, M.; Oobayashi, B.; Umehara, K.; Miyase, T.; Yoshizaki, F. Diastereomers of lithospermic acid and lithospermic acid B from Monarda fistulosa and Lithospermum erythrorhizon. Fitoterapia 2013, 91, 51–59. [Google Scholar] [CrossRef]
- Lin, L.-C.; Pai, Y.-F.; Tsai, T.-H. Isolation of Luteolin and Luteolin-7-O-glucoside from Dendranthema morifolium Ramat Tzvel and Their Pharmacokinetics in Rats. J. Agric. Food Chem. 2015, 63, 7700–7706. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).