
Combined Transcriptomics and Metabolomics Analysis Reveals the Effect of Selenium Fertilization on Lycium barbarum Fruit

Abstract
:1. Introduction
2. Results
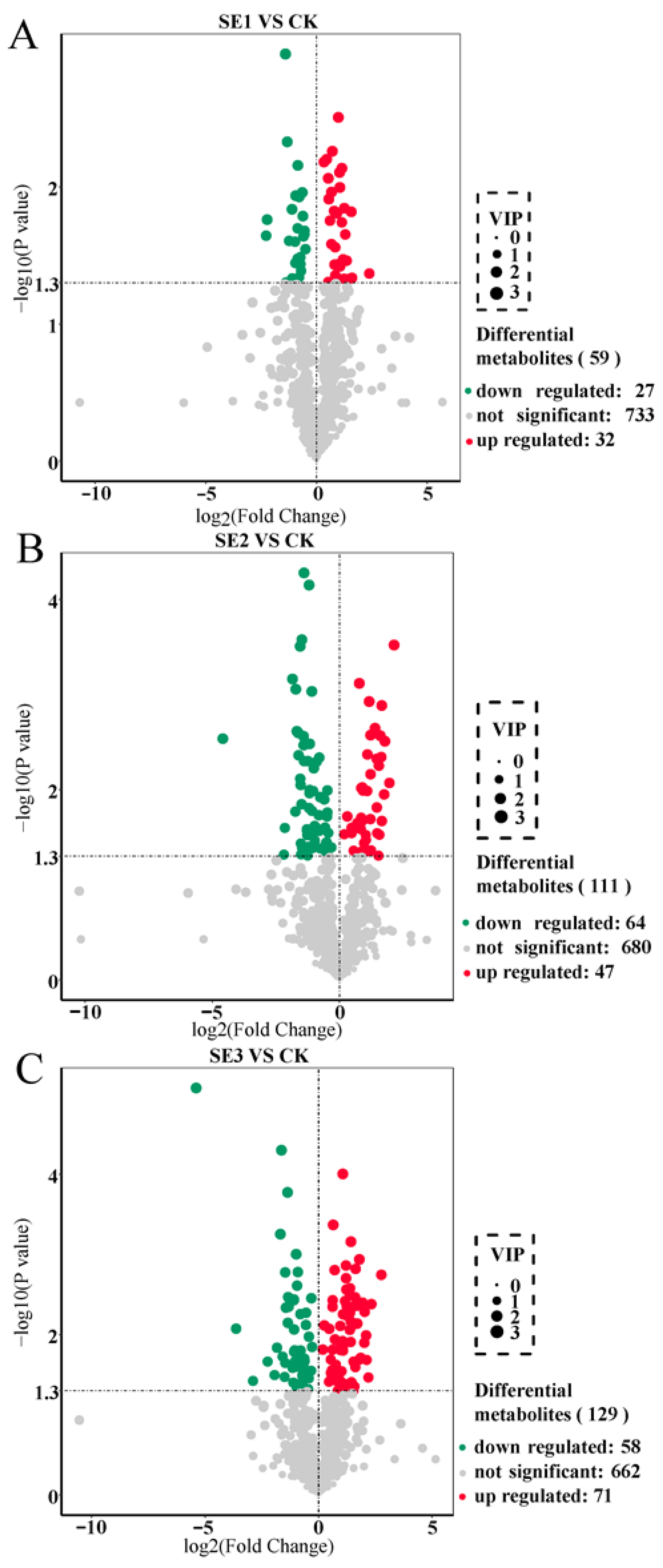
2.1. Screening for Significantly Different Metabolites in Lycium barbarum
2.2. Effect of Exogenous Selenium on Substances Related to Amino Acid Metabolism in Lycium barbarum
2.3. Effect of Exogenous Selenium on Flavonoid-Metabolism-Related Substances in Lycium barbarum
2.4. Effect of Exogenous Selenium on Substances Related to the Alkaloid Metabolism
2.5. Correlation Analysis of Fruit Metabolites of Lycium barbarum
2.6. Analysis of the Transcriptomes of Differentially Expressed Genes in Lycium barbarum
2.6.1. RNA-Seq Data Quality Assessment
2.6.2. Differentially Expressed Genes
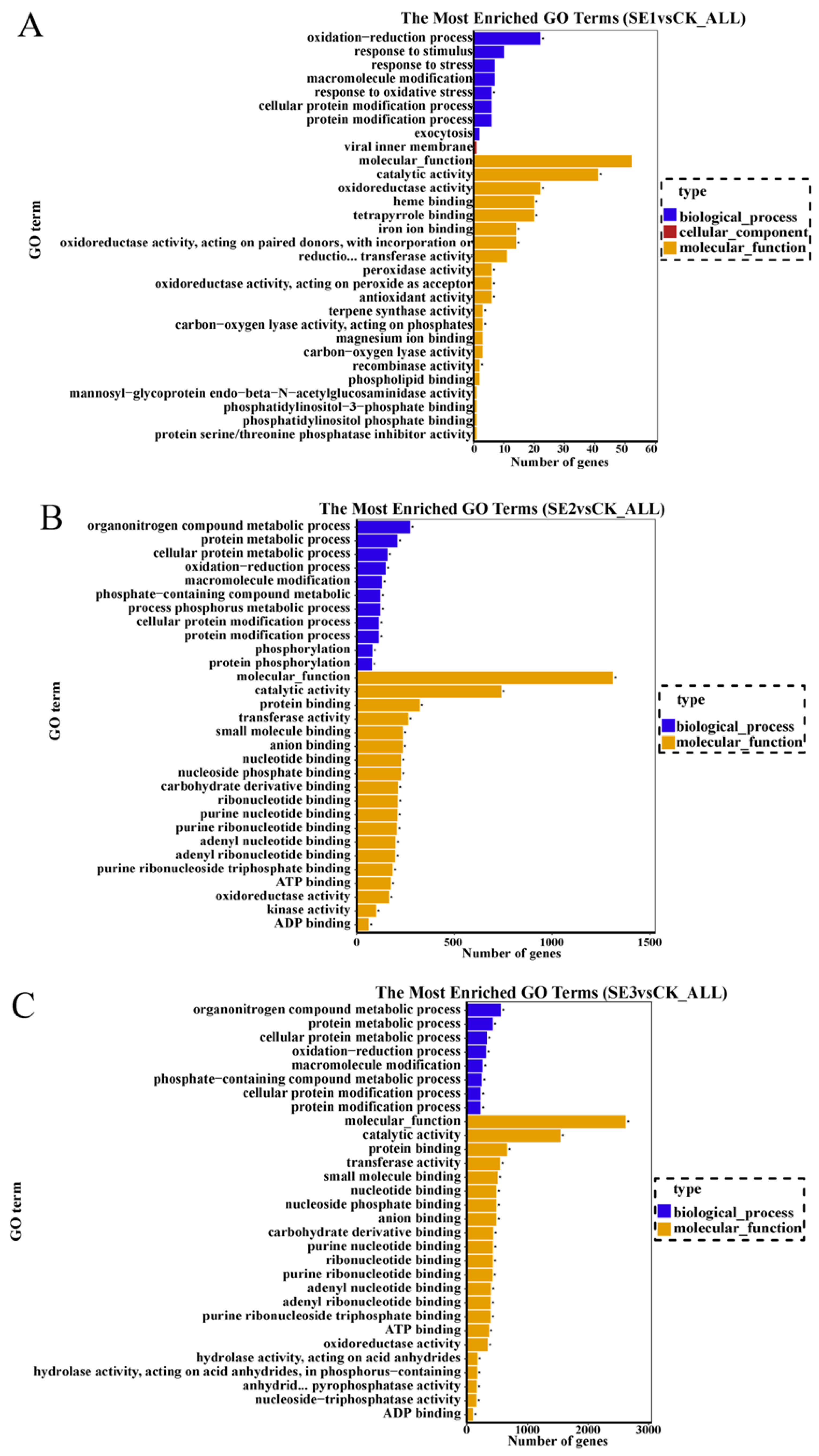
2.6.3. GO Enrichment Analysis of Differentially Expressed Genes
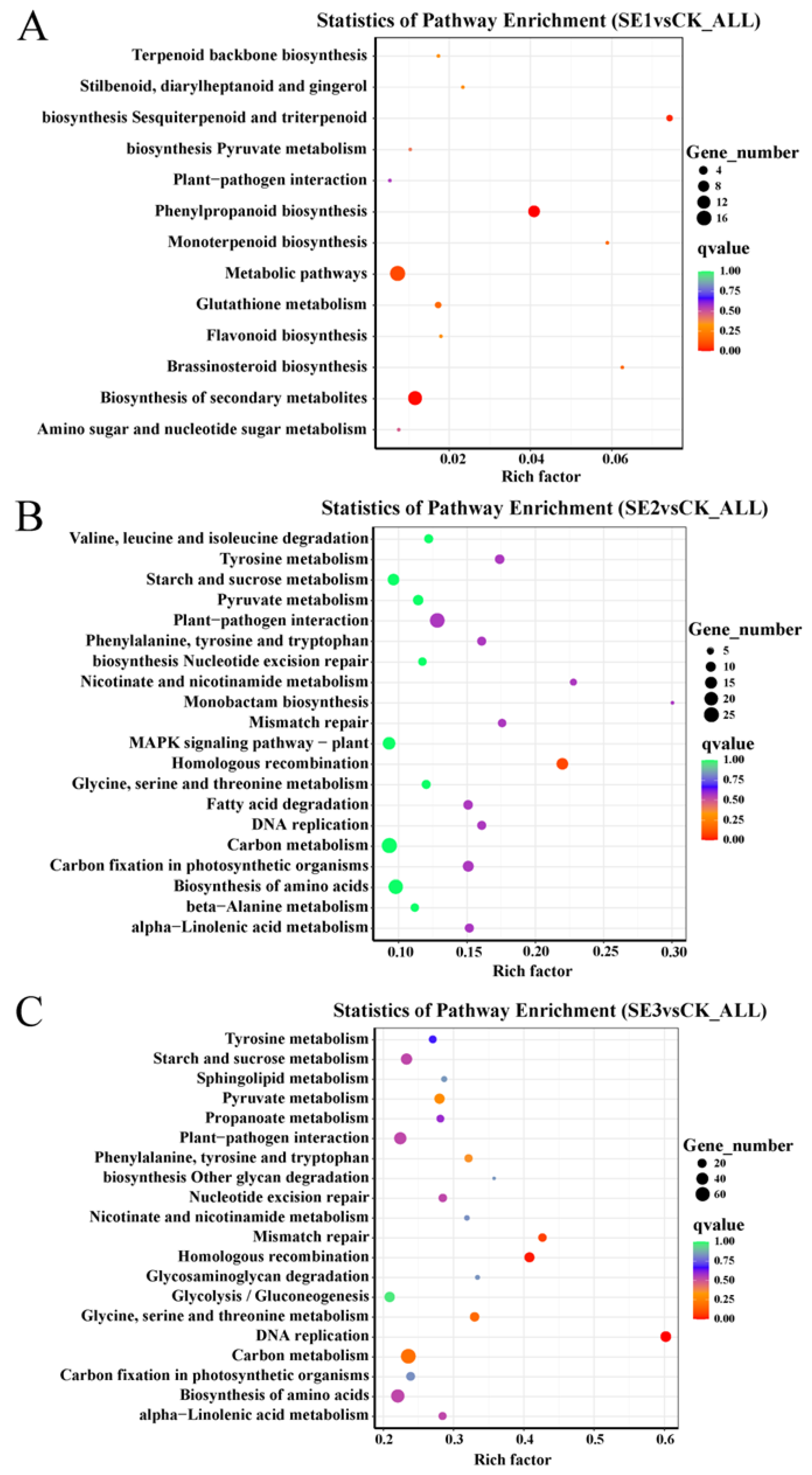
2.6.4. Results of KEGG Enrichment Analysis of Differentially Expressed Genes
2.6.5. Metabolomics and Transcriptomics Association Analysis
3. Discussion
4. Materials and Methods
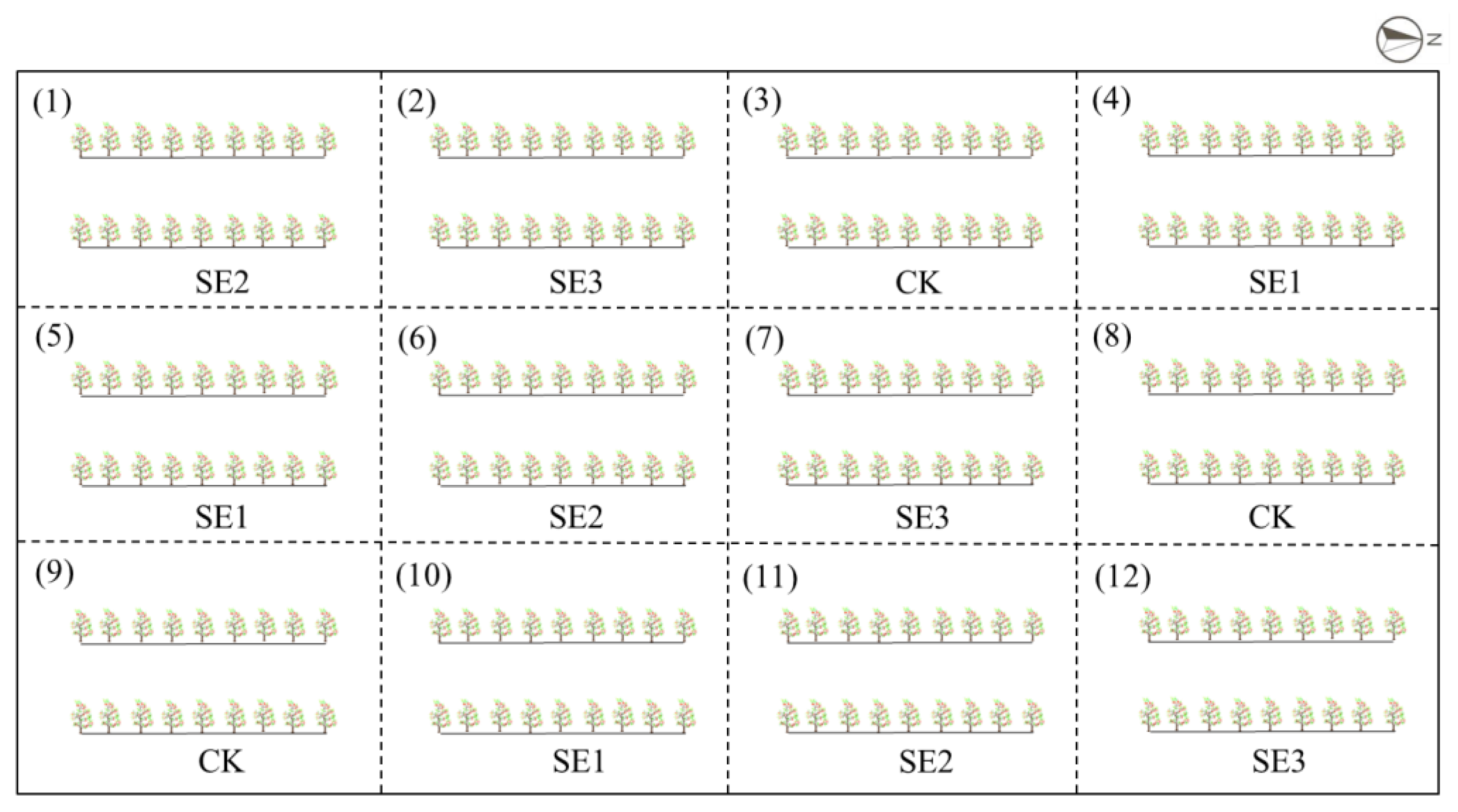
4.1. Study Region
4.2. Study Methods
4.3. Sample Collection
4.4. Experimental Methods
4.4.1. Metabolome Analyses
4.4.2. Transcriptomics Analysis
4.5. Data Processing
4.5.1. Metabolomic Data Analysis
4.5.2. Transcriptome Data Analysis
4.5.3. Differential Gene GO Enrichment and KEGG Pathway Analysis
4.5.4. Combined Analysis of Metabolomic Data and Transcriptomic Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, Y.J.; Lai, Y.K.; Chen, Z.S.Z.; Qu, H.X.; Ma, S.M.; Wang, Y.; Jiang, Y.M. Evolution of physiological characteristics and nutritional quality in fresh goji berry (Lycium barbarum) stored under different temperatures. J. Food Process. Preserv. 2020, 44, e14835. [Google Scholar] [CrossRef]
- Gao, Y.J.; Wei, Y.F.; Wang, Y.Q.; Gao, Y.; Chen, Z.G. Lycium barbarum: A Traditional Chinese Herb and a Promising Anti-Aging Agent. Aging Dis. 2017, 8, 778–791. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.F.; Hao, Z.; Li, Y.H.; Li, H.R.; Wang, L.; Zang, Z.F.; Liao, X.Y.; Zhang, R. Distribution of selenium and zinc in soil-crop system and their relationship with environmental factors. Chemosphere 2020, 242, 125289. [Google Scholar] [CrossRef] [PubMed]
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: It’s role in regulation of inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Mahmud, J.A.; Nahar, K.; Fujita, M. Selenium in plants: Boon or bane? Environ. Exp. Bot. 2020, 178, 104170. [Google Scholar] [CrossRef]
- Shinde, H.; Dudhate, A.; Sathe, A.; Paserkar, N.; Wagh, S.G.; Kadam, U.S. Gene coexpression analysis identifies genes associated with chlorophyll content and relative water content in pearl millet. Plants 2023, 12, 1412. [Google Scholar] [CrossRef]
- Reich, H.J.; Hondal, R.J. Why Nature Chose Selenium. ACS Chem. Biol. 2016, 11, 821–841. [Google Scholar] [CrossRef] [PubMed]
- Fernández, V.; Gil-Pelegrín, E.; Eichert, T. Foliar water and solute absorption: An update. Plant J. 2021, 105, 870–883. [Google Scholar] [CrossRef]
- Trippe, R.C., III; Pilon-Smits, E.A.H. Selenium transport and metabolism in plants: Phytoremediation and biofortification implications. J. Hazard. Mater. 2021, 404, 124178. [Google Scholar] [CrossRef]
- Schiavon, M.; Lima, L.W.; Jiang, Y.; Hawkesford, M.J. Effects of selenium on plant metabolism and implications for crops and consumers. Selenium Plants 2017, 11, 257–275. [Google Scholar]
- Gupta, M.; Shikha, G. An overview of selenium uptake, metabolism, and toxicity in plants. Front. Plant Sci. 2017, 7, 2074. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Zhao, L.; Li, C.R.; Zhao, G.H.; Hu, X.S. Purification and characterization of a novel fungi Se-containing protein from Se-enriched Ganoderma Lucidum mushroom and its Se-dependent radical scavenging activity. Eur. Food Res. Technol. 2007, 224, 659–665. [Google Scholar] [CrossRef]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Wang, C.Y.; Li, C.H.; Wang, X.J. Effect of Selenium Applicationon Selenium Content and Qualities of Lycium barbarum. North. Hortic. 2016, 159–162. (In Chinese) [Google Scholar]
- Niu, Y.; Wang, C.Y.; Li, C.H.; Wu, Y.; Wang, X.J. Effects of Selenium Application on Accumulation of Trace Elements of Lycium barbarum. J. Anhui Agric. Sci. 2016, 44, 114–115+119. (In Chinese) [Google Scholar]
- Ma, Y.P.; Wang, Z.J.; Li, Y.M.; Feng, X.R.; Song, L.H.; Gao, H.D.; Gao, B. Fruit morphological and nutritional quality features of goji berry (Lycium barbarum L.) during fruit development. Sci. Hortic. 2023, 308, 111555. [Google Scholar] [CrossRef]
- Shi, Z.G.; Wei, F.; Wan, R.; Li, Y.X.; Wang, Y.J.; An, W.; Qin, K.; Dai, G.L.; Cao, Y.L.; Feng, J.Y. Impact of nitrogen fertilizer levels on metabolite profiling of the Lycium barbarum L. fruit. Molecules 2019, 24, 3879. [Google Scholar] [CrossRef]
- Chen, C.L.; Xu, M.L.; Wang, C.P.; Qiao, G.X.; Wang, W.W.; Tan, Z.Y.; Wu, T.T.; Zhang, Z.S. Characterization of the Lycium barbarum fruit transcriptome and development of EST-SSR markers. PLoS ONE 2017, 12, e0187738. [Google Scholar] [CrossRef]
- Wei, F.; Wan, R.; Shi, Z.G.; Ma, W.L.; Wang, H.; Chen, Y.J.; Bo, J.H.; Li, Y.X.; An, W.; Qin, K.; et al. Transcriptomics and Metabolomics Reveal the Critical Genes of Carotenoid Biosynthesis and Color Formation of Goji (Lycium barbarum L.) Fruit Ripening. Plants 2023, 12, 2791. [Google Scholar] [CrossRef]
- Golovinskaia, O.; Wang, C.K. Review of functional and pharmacological activities of berries. Molecules 2021, 26, 3904. [Google Scholar] [CrossRef]
- Mocan, A.; Moldovan, C.; Zengin, G.; Bender, O.; Locatelli, M.; Simirgiotis, M.; Atalay, A.; Vodnar, D.C.; Rohn, S.; Crișan, G. UHPLC-QTOF-MS analysis of bioactive constituents from two Romanian Goji (Lycium barbarum L.) berries cultivars and their antioxidant, enzyme inhibitory, and real-time cytotoxicological evaluation. Food Chem. Toxicol. 2018, 115, 414–424. [Google Scholar] [CrossRef]
- Ruffo, M.; Parisi, O.I.; Amone, F.; Malivindi, R.; Gorgoglione, D.; Biasio, F.D.; Scrivano, L.; Pezzi, V.; Puoci, F. Calabrian goji vs. Chinese goji: A comparative study on biological properties. Foods 2017, 6, 30. [Google Scholar] [CrossRef]
- Jiang, Y.Q.; Fang, Z.X.; Leonard, W.; Zhang, P.Z. Phenolic compounds in Lycium berry: Composition, health benefits and industrial applications. J. Funct. Foods 2021, 77, 104340. [Google Scholar] [CrossRef]
- Huang, X.X.; Tang, Q.; Chen, C.J.; Li, Q.; Lin, H.Y.; Bai, S.L.; Zhao, J.; Li, J.; Wang, K.B.; Zhu, M.Z. Combined analysis of transcriptome and metabolome provides insights into nano-selenium foliar applications to improve summer tea quality (Camellia sinensis). LWT 2023, 175, 114496. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, Y.R.; Hao, J.H.; Fan, S.X.; Dong, R.F.; Zeng, H.H.; Liu, C.J.; Han, Y.Y. Effects of selenate and selenite on selenium accumulation and speciation in lettuce. Plant Physiol. Biochem. 2022, 192, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Wang, C.; Chen, F.; Yue, L.; Cao, X.S.; Liu, X.F.; Yao, Y.S.; Wang, Z.Y.; Xing, B.S. Multiomics understanding of improved quality in cherry radish (Raphanus sativus L. var. radculus pers) after foliar application of selenium nanomaterials. Sci. Total Environ. 2022, 824, 153712. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Jia, Z.S.; Tang, M.C.; Wu, J.M. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar]
- Chen, J.W.; Zhu, Z.Q.; Hu, T.X.; Zhu, D.Y. Structure-activity relationship of natural flavonoids in hydroxyl radical-scavenging effects. Acta Pharmacol. Sin. 2002, 23, 667–672. [Google Scholar] [PubMed]
- Deng, K.; Li, L.; Li, L.L.; Xu, F.; Yuan, H.H.; Zha, S.x.; Xiao, X.; Yu, J.; Cheng, S.Y.; Cheng, H. Molecular mechanism of selenium affecting the synthesis of flavonoids in G. biloba leaves. Plant Mol. Biol. Report. 2022, 40, 232–246. [Google Scholar] [CrossRef]
- Ahmad, P.; Abd Allah, E.F.; Hashem, A.; Sarwat, M.; Gucel, S. Exogenous application of selenium mitigates cadmium toxicity in Brassica juncea L. (Czern & Cross) by up-regulating antioxidative system and secondary metabolites. J. Plant Growth Regul. 2016, 35, 936–950. [Google Scholar]
- Li, D.; Zhou, C.; Zou, N.; Wu, Y.L.; Zhang, J.B.; An, Q.s.; Li, J.Q.; Pan, C.P. Nanoselenium foliar application enhances biosynthesis of tea leaves in metabolic cycles and associated responsive pathways. Environ. Pollut. 2021, 273, 116503. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.R.; Yue, C.; Zhou, C.; Huang, Y.T.; Cao, H.L.; Hao, X.Y.; Wang, X.C.; Zeng, J.M. Cloning and Expression Analysis of Sulfate Transporter CsSUL3.5 Gene Tea Plant. Acta Hortic. Sin. 2015, 42, 2306–2314. (In Chinese) [Google Scholar]
- Roy, A. A review on the alkaloids an important therapeutic compound from plants. IJPB 2017, 3, 1–9. [Google Scholar]
- Pilon-Smits, E.A.H.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Kalaei, M.H.R.; Abdossi, V.; Danaee, E. Evaluation of foliar application of selenium and flowering stages on selected properties of Iranian Borage as a medicinal plant. Sci. Rep. 2022, 12, 12568. [Google Scholar] [CrossRef]
- González-Lemus, U.; Medina-Pérez, G.; Espino-García, J.J.; Fernández-Luqueño, F.; Campos-Montiel, R.; Almaraz-Buendía, I.; Reyes-Munguía, A.; Urrutia-Hernández, T. Nutritional Parameters, Biomass Production, and Antioxidant Activity of Festuca arundinacea Schreb. Conditioned with Selenium Nanoparticles. Plants 2022, 11, 2326. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, K.; Pajoro, A.; Angenent, G.C. Regulation of transcription in plants: Mechanisms controlling developmental switches. Nat. Rev. Genet. 2010, 11, 830–842. [Google Scholar] [CrossRef]
- Hameed, A.; Sharma, I.; Kumar, A.; Azooz, M.M.; Lone, H.A.; Ahmad, P. Glutathione metabolism in plants under environmental stress. In Oxidative Damage to Plants; Academic Press: Cambridge, MA, USA, 2014; pp. 183–200. [Google Scholar]
- Noctor, G.; Queval, G.; Mhamdi, A.; Chaouch, S.; Foyer, C.H. Glutathione. Arab. Book 2011, 9, e0142. [Google Scholar] [CrossRef]
- May, M.J.; Vernoux, T.; Leaver, C.; Montagu, M.V.; Inzé, D. Glutathione homeostasis in plants: Implications for environmental sensing and plant development. J. Exp. Bot. 1998, 49, 649–667. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-garcia, B.; Queval, G.; Foyer, C. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef]
- Patra, B.; Schluttenhofer, C.; Wu, Y.M.; Pattanaik, S.; Yuan, L. Transcriptional regulation of secondary metabolite biosynthesis in plants. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2013, 1829, 1236–1247. [Google Scholar] [CrossRef]
- De Luca, V.; St Pierre, B. The cell and developmental biology of alkaloid biosynthesis. Trends Plant Sci. 2000, 5, 168–173. [Google Scholar] [CrossRef]
- Winkel, B.S.J. Metabolic channeling in plants. Annu. Rev. Plant Biol. 2004, 55, 85–107. [Google Scholar] [CrossRef] [PubMed]
- Butelli, E.; Titta, L.; Giorgio, M.; Mock, H.P.; Matros, A.; Peterek, S.; Schijlen, E.G.W.M.; Hall, R.D.; Bovy, A.G.; Luo, J.; et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 2008, 26, 1301–1308. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Barrieu, F.; Bogs, J.; Kappel, C.; Delrot, S.; Lauvergeat, V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J. Exp. Bot. 2011, 62, 2465–2483. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.F.; Lu, C.H.; Huang, L.Q.; Zhang, H.Q.; Zhao, Z.Q.; Liu, X.W. Effects of spraying selenium in different forms and at different stages on selenium absorption and accumulation and main quality indexes of ‘Jintao’ kiwifruit. J. Fruit Sci. 2018, 35, 1385–1392. (In Chinese) [Google Scholar]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw Reads | Raw Bases | Clean Reads | Clean Bases | Error Rate | Q20 | Q30 | GC Content |
|---|---|---|---|---|---|---|---|---|
| CK-1 | 45,330,910 | 6.79 G | 44,340,314 | 6.65 G | 0.03% | 97.82% | 93.72% | 42.53% |
| CK-2 | 48,021,620 | 7.2 G | 47,150,434 | 7.07 G | 0.03% | 97.82% | 93.71% | 42.44% |
| CK-3 | 41,223,694 | 6.18 G | 40,393,914 | 6.06 G | 0.03% | 97.94% | 93.99% | 42.51% |
| SE1-1 | 46,940,776 | 7.04 G | 46,199,336 | 6.93 G | 0.03% | 97.91% | 93.89% | 42.56% |
| SE1-2 | 46,966,566 | 7.04 G | 46,364,144 | 6.95 G | 0.03% | 97.91% | 93.85% | 42.40% |
| SE1-3 | 40,378,554 | 6.05 G | 39,796,962 | 5.97 G | 0.03% | 97.87% | 93.82% | 42.45% |
| SE2-1 | 44,866,144 | 6.72 G | 44,070,522 | 6.61 G | 0.03% | 97.62% | 93.28% | 42.20% |
| SE2-2 | 53,107,194 | 7.96 G | 52,151,418 | 7.82 G | 0.03% | 97.82% | 93.74% | 42.28% |
| SE2-3 | 49,901,306 | 7.48 G | 49,416,786 | 7.41 G | 0.03% | 97.83% | 93.72% | 42.60% |
| SE3-1 | 41,495,698 | 6.22 G | 40,914,436 | 6.14 G | 0.03% | 97.90% | 93.87% | 42.48% |
| SE3-2 | 45,808,966 | 6.87 G | 44,980,762 | 6.75 G | 0.03% | 97.86% | 93.81% | 42.44% |
| SE3-3 | 47,927,700 | 7.18 G | 47,208,160 | 7.08 G | 0.03% | 97.96% | 94.02% | 42.39% |
| Fertilizer Types | Selenium Content | Fertilization Concentration | Solubility | Water Solubility |
|---|---|---|---|---|
| Sodium selenite, SE1 | 44.70% | 100mg/L | 95.5% | Soluble in water |
| Nano selenium, SE2 | 4.00% | 100mg/L | 99.0% | Soluble in water |
| Organic selenium, SE3 | 0.60% | 100mg/L | 99.0% | Extremely soluble in water |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.; Jiang, T.; Wang, Y.; Wang, X.; Wang, R. Combined Transcriptomics and Metabolomics Analysis Reveals the Effect of Selenium Fertilization on Lycium barbarum Fruit. Molecules 2023, 28, 8088. https://doi.org/10.3390/molecules28248088
Yang W, Jiang T, Wang Y, Wang X, Wang R. Combined Transcriptomics and Metabolomics Analysis Reveals the Effect of Selenium Fertilization on Lycium barbarum Fruit. Molecules. 2023; 28(24):8088. https://doi.org/10.3390/molecules28248088
Chicago/Turabian StyleYang, Wenqin, Tingting Jiang, Yaqi Wang, Xiaojing Wang, and Rui Wang. 2023. "Combined Transcriptomics and Metabolomics Analysis Reveals the Effect of Selenium Fertilization on Lycium barbarum Fruit" Molecules 28, no. 24: 8088. https://doi.org/10.3390/molecules28248088
APA StyleYang, W., Jiang, T., Wang, Y., Wang, X., & Wang, R. (2023). Combined Transcriptomics and Metabolomics Analysis Reveals the Effect of Selenium Fertilization on Lycium barbarum Fruit. Molecules, 28(24), 8088. https://doi.org/10.3390/molecules28248088