
Influence of Amino Acid Substitutions in ApoMb on Different Stages of Unfolding of Amyloids

 , , , ,
, , , ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
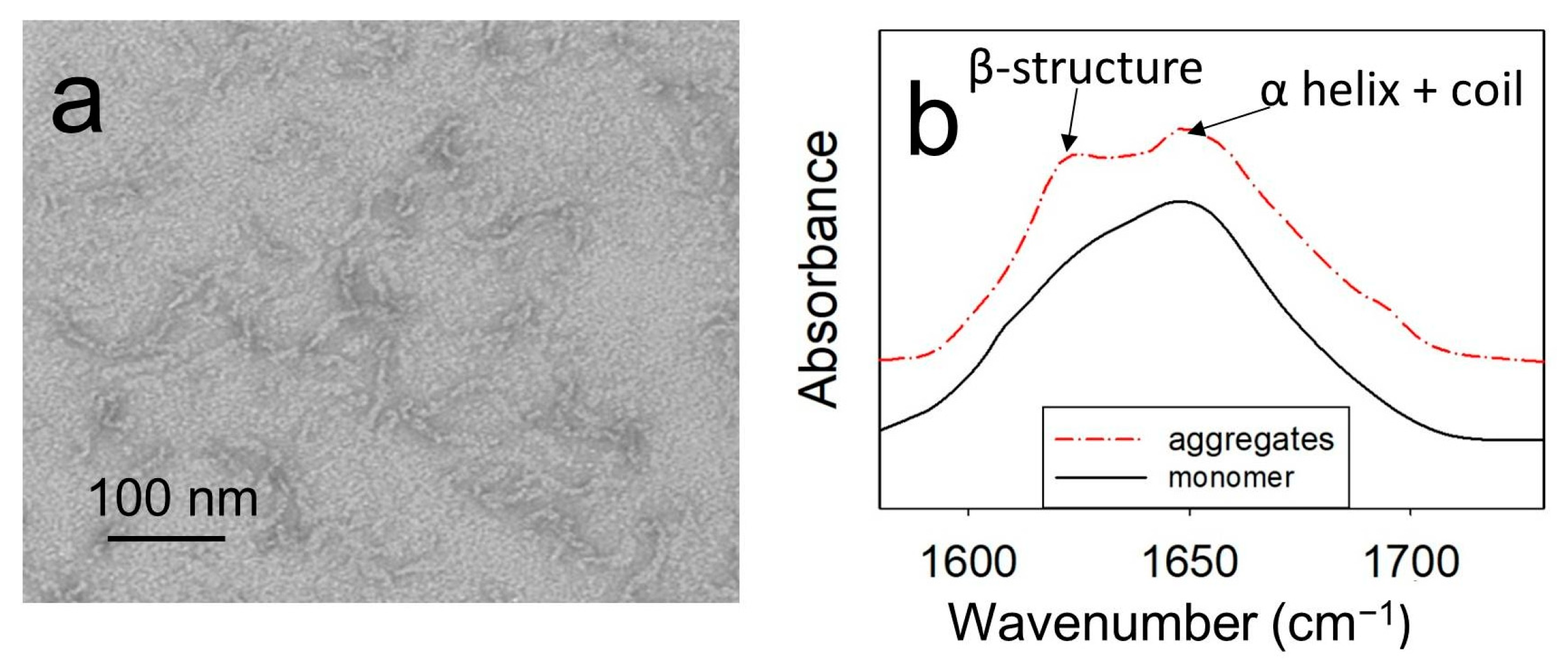
2.1. At a Temperature of 40 °C and pH 5.5, ApoMb Forms Amyloids
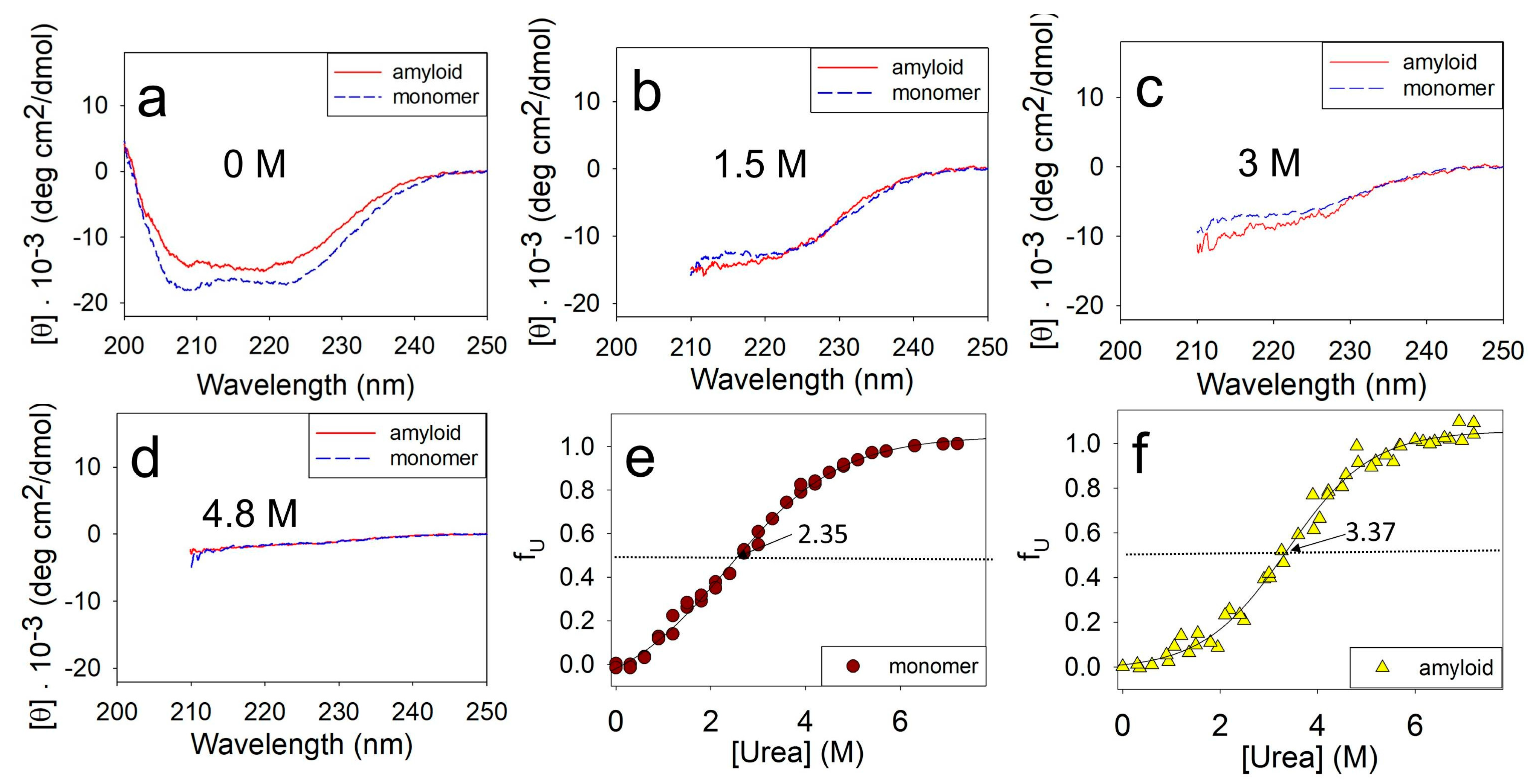
2.2. Changes in the Secondary Structure during Amyloids’ Unfolding
2.3. Influence of Amino Acid Substitutions on the Stability of Amyloids’ Secondary Structure
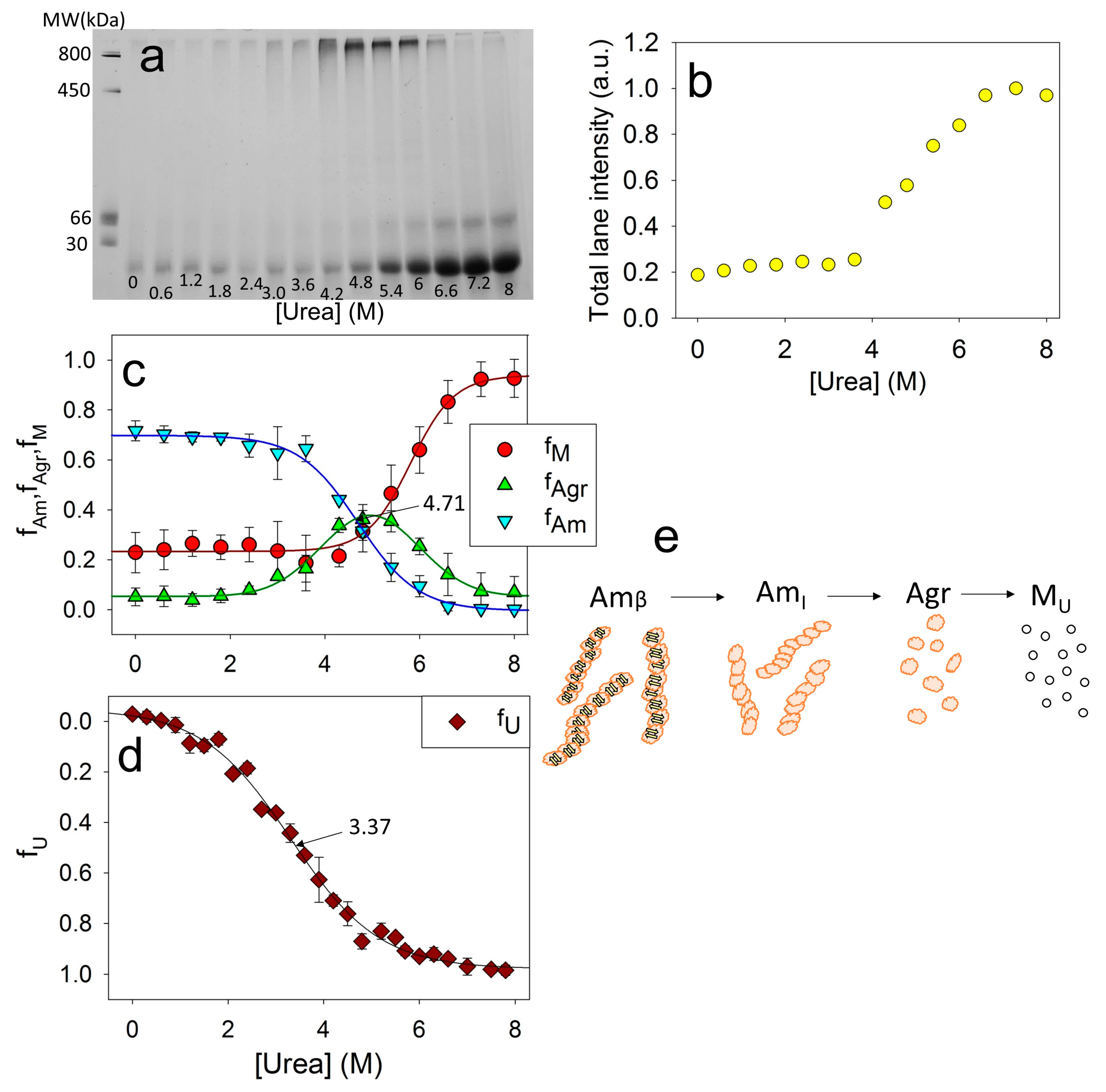
2.4. Changes in the Molecular Weight of the Aggregates during Amyloids’ Unfolding
2.5. Influence of Amino Acid Substitutions on the Stability of Amyloids Relative to the Intermediate Aggerates and the Stability of Intermediate Aggregates Relative to the Unfolded Monomer
3. Discussion
4. Materials and Methods
4.1. Protein Expression and Purification
4.2. Amyloids’ Formation
4.3. Electron Microscopy
4.4. Fourier Transforms Infrared Spectroscopy (FTIR)
4.5. Amyloid Unfolding by Urea
4.6. Circular Dichroism (CD)
4.7. Blue Native-PAGE (BN-PAGE) Electrophoresis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dill, K.A.; MacCallum, J.L. The protein-folding problem, 50 years on. Science 2012, 338, 1042–1046. [Google Scholar] [CrossRef]
- Dobson, C.M. Protein folding and misfolding. Nature 2003, 426, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Jahn, T.R.; Radford, S.E. Folding versus aggregation: Polypeptide conformations on competing pathways. Arch. Biochem. Biophys. 2008, 469, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Iadanza, M.G.; Jackson, M.P.; Hewitt, E.W.; Ranson, N.A.; Radford, S.E. A new era for understanding amyloid structures and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 755–773. [Google Scholar] [CrossRef] [PubMed]
- Chiti, F.; Dobson, C.M. Protein misfolding, functional amyloid, and human disease. Annu. Rev. Biochem. 2006, 75, 333–366. [Google Scholar] [CrossRef]
- Knowles, T.P.; Vendruscolo, M.; Dobson, C.M. The amyloid state and its association with protein misfolding diseases. Nat. Rev. Mol. Cell Biol. 2014, 15, 384–396. [Google Scholar] [CrossRef]
- Otzen, D.; Riek, R. Functional amyloids. Cold Spring Harb. Perspect. Biol. 2019, 11, a0338608. [Google Scholar] [CrossRef]
- Maji, S.K.; Perrin, M.H.; Sawaya, M.R.; Jessberger, S.; Vadodaria, K.; Rissman, R.A.; Singru, P.S.; Nilsson, K.P.R.; Simon, R.; Schubert, D.; et al. Functional amyloids as natural storage of peptide hormones in pituitary secretory granules. Science 2009, 325, 328–332. [Google Scholar] [CrossRef]
- Knowles, T.P.J.; Mezzenga, R. Amyloid fibrils as building blocks for natural and artificial functional materials. Adv. Mater. 2016, 28, 6546–6561. [Google Scholar] [CrossRef]
- MacPhee, C.E.; Woolfson, D.N. Engineered and designed peptide-based fibrous biomaterials. Curr. Opin. Solid State Mater. Sci. 2004, 8, 141–149. [Google Scholar] [CrossRef]
- Eisenberg, D.; Jucker, M. The amyloid state of proteins in human diseases. Cell 2012, 148, 1188–1203. [Google Scholar] [CrossRef] [PubMed]
- Makin, O.S.; Atkins, E.; Sikorski, P.; Johansson, J.; Serpell, L.C. Molecular basis for amyloid fibril formation and stability. Proc. Natl. Acad. Sci. USA 2005, 102, 315–320. [Google Scholar] [CrossRef]
- Tycko, R. Solid-state NMR studies of amyloid fibrils structure. Annu. Rev. Phys. Chem. 2011, 62, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Lomakin, A.; Chung, D.S.; Benedek, G.B.; Kirschner, D.A.; Teplow, D.B. On the nucleation and growth of amyloid beta-protein fibrils: Detection of nuclei and quantification of rate constants. Proc. Natl. Acad. Sci. USA 1996, 93, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Chiti, F.; Stefani, M.; Taddei, N.; Ramponi, G.; Dobson, C.M. Rationalization of the effects of mutations on peptide and protein aggregation rates. Nature 2003, 424, 805–808. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, A.W.; Saibil, H.R. Cryo-EM of amyloid fibrils and cellular aggregates. Curr. Opin. Struct. Biol. 2019, 58, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Michaels, T.C.T.; Saric, A.; Meisl, G.; Heller, G.T.; Curk, S.; Arosio, P.; Linse, S.; Dobson, C.M.; Vendruscolo, M.; Knowles, T.P.J. Thermodynamic and kinetic design principles for amyloid-aggregation inhibitors. Proc. Natl. Acad. Sci. USA 2020, 117, 24251–24257. [Google Scholar] [CrossRef]
- Narimoto, T.; Sakurai, K.; Okamoto, A.; Chatani, E.; Hoshito, M.; Hasegawa, K.; Naiki, H.; Goto, Y. Conformational stability of amyloid fibrils of beta2-microglobulin probed by guanidine-hydrochloride-induced unfolding. FEBS Lett. 2004, 576, 313–319. [Google Scholar] [CrossRef]
- Gazit, E. The “Correctly Folded” state of proteins: Is it a metastable state? Angew. Chem. Int. Ed. Engl. 2002, 41, 257–259. [Google Scholar] [CrossRef]
- Noda, S.; So, M.; Adachi, M.; Kardos, J.; Akazawa-Ogawa, Y.; Hagihara, Y.; Goto, Y. Thioflavin T-silent denaturation intermediates support the main-chain-dominated architecture of amyloid fibrils. Biochemistry 2016, 55, 3937–3948. [Google Scholar] [CrossRef]
- Sulatsky, M.I.; Sulatskaya, A.I.; Stepanenko, O.V.; Povarova, O.I.; Kuznetsova, I.M.; Turoverov, K.K. Denaturant effect on amyloid fibrils: Declasterization, depolymerization, denaturation and reassembly. Int. J. Biol. Macromol. 2020, 150, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Breydo, L.; Makarava, N.; Yang, Q.; Bocharova, O.V.; Baskakov, I.V. Site-specific conformational studies of prion protein (PrP) amyloid fibrils revealed two cooperative folding domains within amyloid structure. J. Biol. Chem. 2007, 282, 9090–9097. [Google Scholar] [CrossRef] [PubMed]
- Samatova, E.N.; Katina, N.S.; Balobanov, V.A.; Melnik, B.S.; Dolgikh, D.A.; Bychkova, V.E.; Finkelstein, A.V. How strong are side chain interactions in the folding intermediate? Protein Sci. 2009, 18, 2152–2159. [Google Scholar] [CrossRef] [PubMed]
- Stagg, L.; Samiotakis, A.; Homouz, D.; Cheung, M.S.; Wittung-Stafshede, P. Residue-specific analysis of frustration in the folding landscape of repeat beta/alpha protein apoflavodoxin. J. Mol. Biol. 2010, 396, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Eliezer, D.; Yao, J.; Dyson, H.J.; Wright, P.E. Structural and dynamic characterization of partially folded states of apomyoglobin and implications for protein folding. Nat. Struct. Biol. 1998, 5, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, J.T.; Kao, Y.H.; Cocco, M.J. The native state of apomyoglobin described by proton NMR spectroscopy: The A-B-G-H interface of wild-type sperm whale apomyoglobin. Proteins 1996, 25, 267–285. [Google Scholar] [CrossRef]
- Hughson, F.M.; Wright, P.E.; Baldwin, R.L. Structural characterization of a partly folded apomyoglobin intermediate. Science 1990, 249, 1544–1548. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, C.; Dyson, H.J.; Wright, P.E. Identification of native and non-native structure in kinetic folding intermediates of apomyoglobin. J. Mol. Biol. 2006, 355, 139–156. [Google Scholar] [CrossRef]
- Fandrich, M.; Forge, V.; Buder, K.; Kittler, M.; Dobson, C.M.; Diekmann, S. Myoglobin forms amyloid fibrils by association of unfolded polypeptide segments. Proc. Natl. Acad. Sci. USA 2003, 100, 15463–15468. [Google Scholar] [CrossRef]
- Sirangelo, I.; Malmo, C.; Casillo, M.; Mezzogiorno, A.; Papa, M.; Irace, G. Tryptophanyl substitutions in apomyoglobin determine protein aggregation and amyloid-like fibril formation at physiological pH. J. Biol. Chem. 2002, 277, 45887–45891. [Google Scholar] [CrossRef]
- Katina, N.S.; Balobanov, V.A.; Ilyina, N.B.; Vasiliev, V.D.; Marchenkov, V.V.; Glukhov, A.S.; Nikulin, A.D.; Bychkova, V.E. sw ApoMb amyloid aggregation under nondenaturing conditions: The role of native structure stability. Biophys. J. 2017, 113, 991–1001. [Google Scholar] [CrossRef][Green Version]
- Picotti, P.; Franceschi, G.D.; Frare, E.; Spolaore, B.; Zambonin, M.; Chiti, F.; Polverino de Laureto, P.; Fontana, A. Amyloid fibril formation and disaggregation of fragment 1-29 of apomyoglobin: Insights into the effect of pH on protein fibrillogenesis. J. Mol. Biol. 2007, 367, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Zandomeneghi, G.; Krebs, M.R.H.; McCammon, M.G.; Fandrich, M. FTIR reveals structural differences between native beta-sheet proteins and amyloid fibrils. Protein Sci. 2004, 13, 3314–3321. [Google Scholar] [CrossRef] [PubMed]
- Fauchere, J.L.; Pliska, V. Hydrophobic parameters-p of amino acid side-chains from the partitioning of N-acetyl aminoacid amide. Eur. J. Med. Chem. 1983, 18, 369–375. [Google Scholar]
- Schuck, P.; Perugini, M.A.; Gonzales, N.R.; Howlett, G.J.; Schubert, D. Size-distribution analysis of proteins by analytical ultracentrifugation: Strategies and application to model system. Biophys. J. 2002, 82, 1096–1111. [Google Scholar] [CrossRef] [PubMed]
- Adamcik, J.; Mezzenga, R. Amyloid polymorphism in the protein folding and aggregation energy landscape. Angew. Chem. Int. Ed. Engl. 2018, 57, 8370–8382. [Google Scholar] [CrossRef]
- Baryshnikova, E.N.; Melnik, B.S.; Finkelstein, A.V.; Semisotnov, G.V.; Bychkova, V.E. Three-state protein folding: Experimental determination of free-energy profile. Protein Sci. 2005, 14, 2658–2667. [Google Scholar] [CrossRef]
- Gill, S.C.; von Hippel, P.H. Calculation of protein extinction coefficients from amino acid sequence data. Anal. Biochem. 1989, 182, 319–326. [Google Scholar] [CrossRef]
- Marchenkov, V.; Ivashina, T.; Marchenko, N.; Ryabova, N.; Selivanova, O.; Timchenko, A.; Kihara, H.; Ksenzenko, V.; Semisotnov, G. In vivo incorporation of photoproteins into GroEL chaperonin retaining structural and functional properties. Molecules 2023, 28, 1901. [Google Scholar] [CrossRef]
- Schagger, H.; Jagow, G. Blue native electrophoresis for isolation of membrane protein complexes in enzymatically active form. Anal. Biochem. 1991, 199, 223–231. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ApoMb Variant | [urea]1/2β-U (M) | fN (%) Data from [31] | ∆hydrophobicity (kcal/mol) Octanol/Water Fraction, Data from [34] | [urea]1/2Am-Agr (M) | [urea]1/2Agr-M (M) |
|---|---|---|---|---|---|
| WT | 3.9 ± 0.06 | 83.4 ± 0.6 | 0 | 5.2 ± 0.3 | 6.0 ± 0.4 |
| V10A | 3.0 ± 0.09 | 81.8 ± 0.9 | −1.2 | 4.2 ± 0.1 | 4.8 ± 0.2 |
| V10F | 3.4 ± 0.06 | 74.7 ± 0.6 | 0.8 | 4.7 ± 0.2 | 5.8 ± 0.1 |
| L115A | 2.9 ± 0.07 | 63.8 ± 0.8 | −1.9 | 4.4 ± 0.1 | 5.6 ± 0.2 |
| L115F | 3.4 ± 0.08 | 74.9 ± 2.1 | 0.1 | 5.5 ± 0.2 | 5.7 ± 0.2 |
| M131A | 2.7± 0.05 | 56.5 ± 2.4 | −1.2 | 4.1 ± 0.2 | 4.8 ± 0.1 |
| M131W | 3.0 ± 0.04 | 87.9 ± 1.7 | 1.4 | 4.6 ± 0.2 | 5.1 ± 0.1 |
| Position of Substitution | ∆[urea]1/2β-U (M) | ∆[urea]1/2Am-Agr (M) | ∆[urea]1/2Agr-M (M) |
|---|---|---|---|
| V10 | 0.4 ± 0.2 | 0.5 ± 0.3 | 0.9 ± 0.3 |
| L115 | 0.5 ± 0.2 | 1.0 ± 0.3 | 0.2 ± 0.2 |
| M131 | 0.3 ± 0.2 | 0.5 ± 0.4 | 0.3 ± 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katina, N.; Marchenkov, V.; Ryabova, N.; Ilyina, N.; Marchenko, N.; Balobanov, V.; Finkelstein, A. Influence of Amino Acid Substitutions in ApoMb on Different Stages of Unfolding of Amyloids. Molecules 2023, 28, 7736. https://doi.org/10.3390/molecules28237736
Katina N, Marchenkov V, Ryabova N, Ilyina N, Marchenko N, Balobanov V, Finkelstein A. Influence of Amino Acid Substitutions in ApoMb on Different Stages of Unfolding of Amyloids. Molecules. 2023; 28(23):7736. https://doi.org/10.3390/molecules28237736
Chicago/Turabian StyleKatina, Natalya, Victor Marchenkov, Natalya Ryabova, Nelly Ilyina, Natalia Marchenko, Vitalii Balobanov, and Alexey Finkelstein. 2023. "Influence of Amino Acid Substitutions in ApoMb on Different Stages of Unfolding of Amyloids" Molecules 28, no. 23: 7736. https://doi.org/10.3390/molecules28237736
APA StyleKatina, N., Marchenkov, V., Ryabova, N., Ilyina, N., Marchenko, N., Balobanov, V., & Finkelstein, A. (2023). Influence of Amino Acid Substitutions in ApoMb on Different Stages of Unfolding of Amyloids. Molecules, 28(23), 7736. https://doi.org/10.3390/molecules28237736