
Effect of Chestnut (Castanea Mollissima Blume) Bur Polyphenol Extract on Shigella dysenteriae: Antibacterial Activity and the Mechanism

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
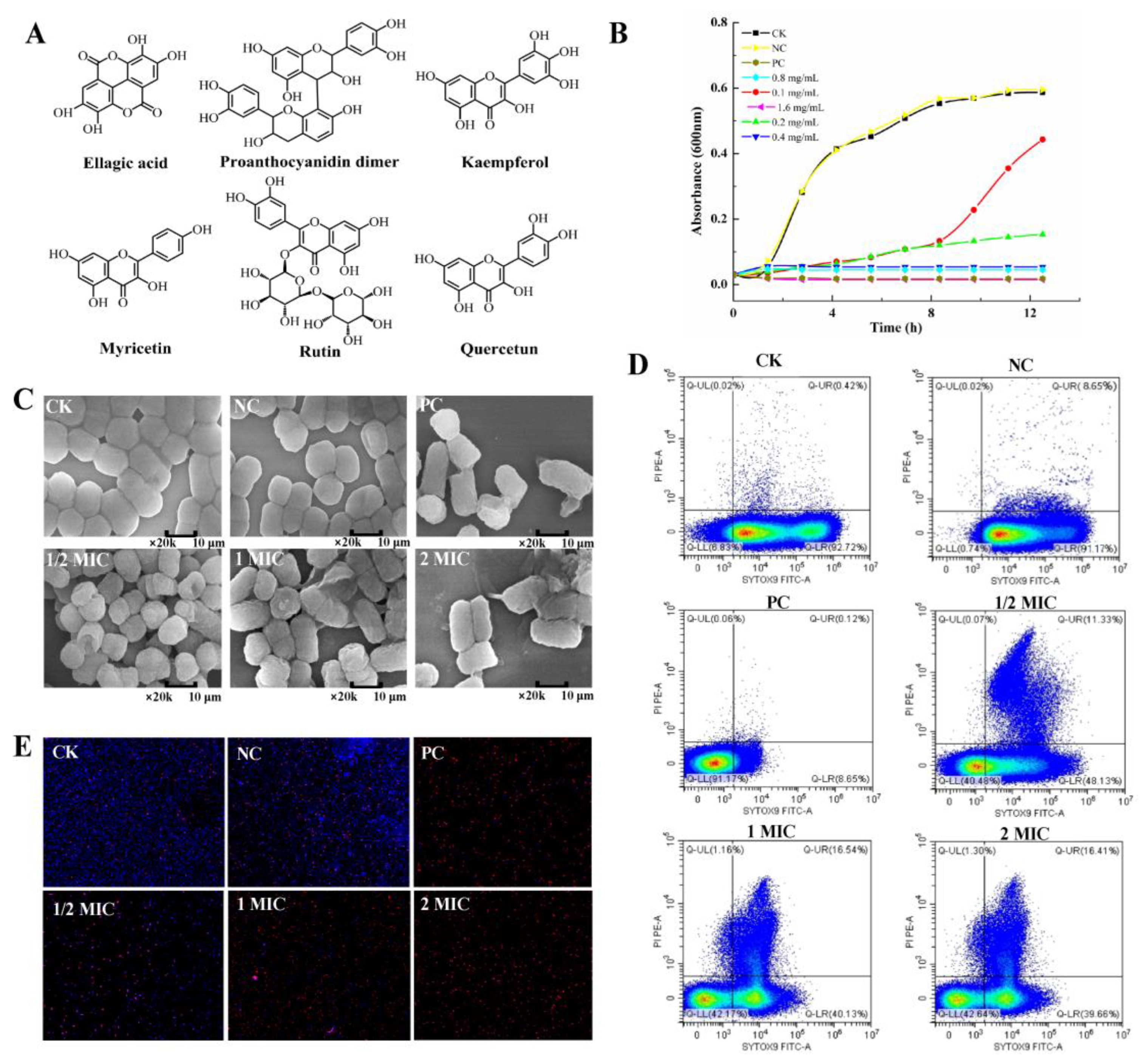
2.1. CBPE Components
2.2. Antibacterial Activity of CBPE against S. dysenteriae
2.3. Effect of CBPE on the Cell Membrane and Cell Wall of S. dysenteriae
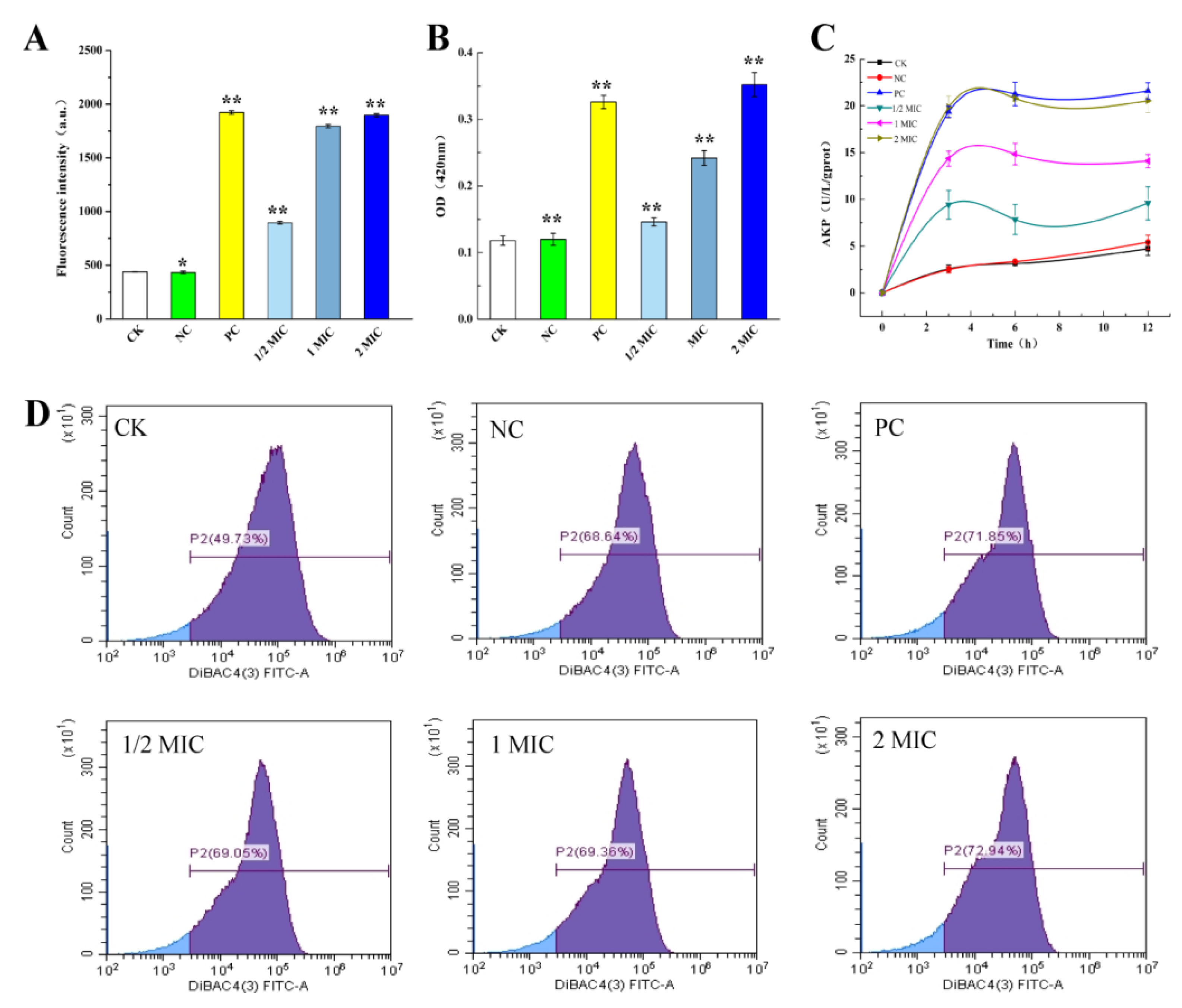
2.3.1. Effect of CBPE on the Cell Membrane Permeability of S. dysenteriae
2.3.2. Effect of CBPE on the Cell Wall Integrity of S. dysenteriae
2.3.3. Effect of CBPE on the Cell Membrane Potential of S. dysenteriae
2.4. Effects of CBPE on Cell Contents
2.4.1. Effect of CBPE on the Cell Membrane Potential of S. dysenteriae
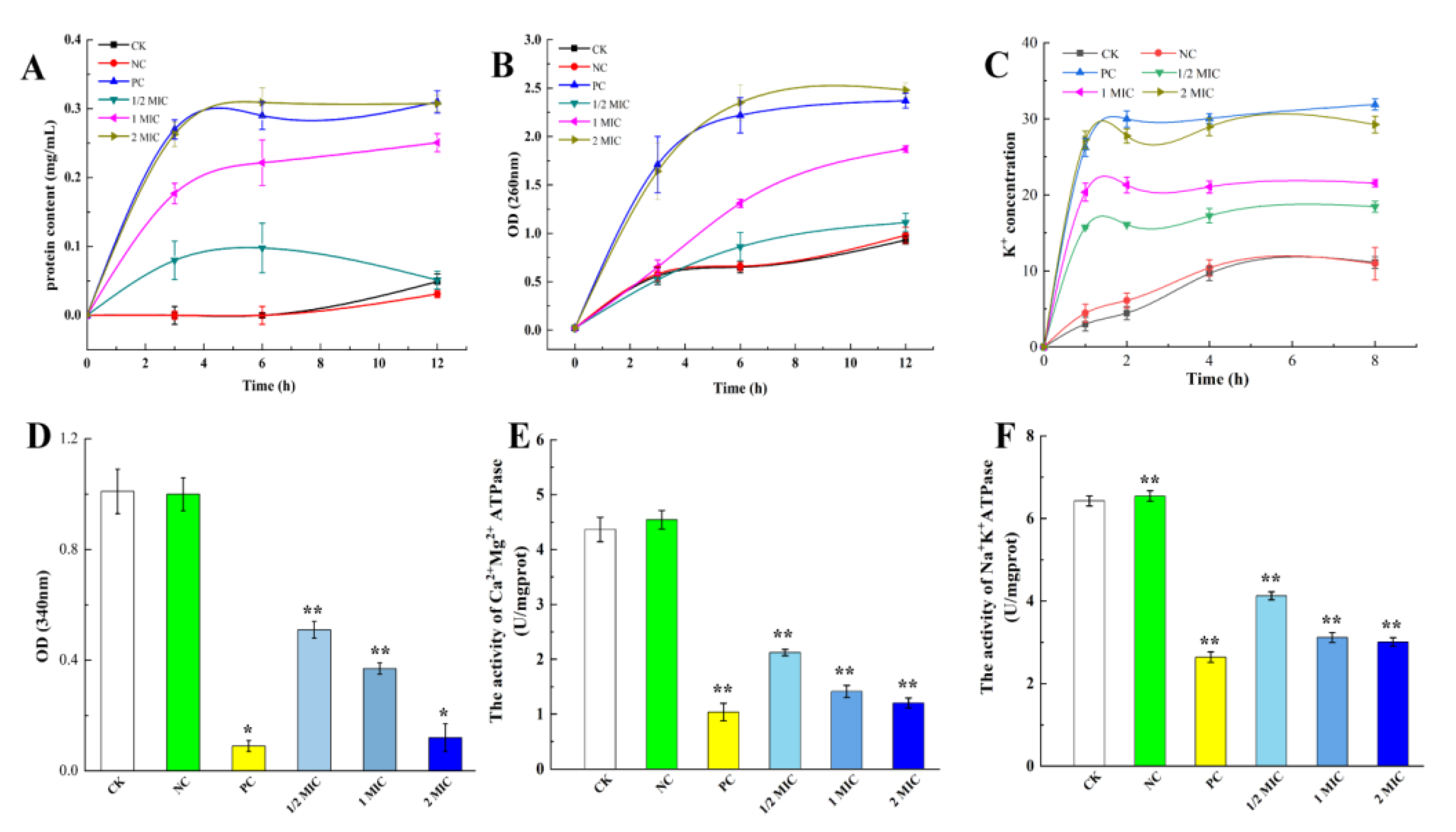
2.4.2. Effect of CBPE on the Macromolecular Leakage of S. dysenteriae
2.4.3. Effect of CBPE on the Extracellular K+ Content of S. dysenteriae
2.4.4. Effects of CBPE on the ATP Content and ATPase Activity in S. dysenteriae Cells
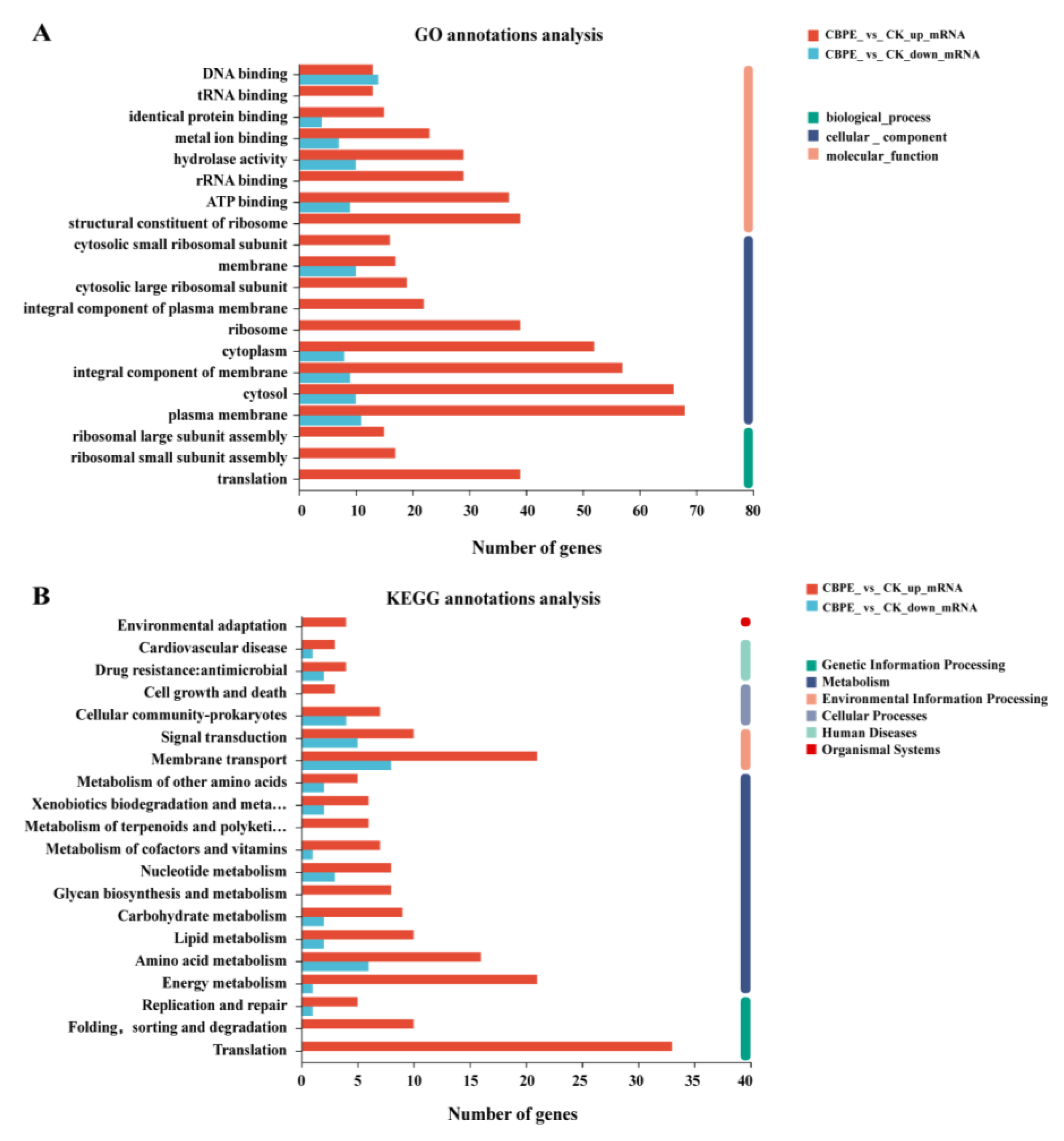
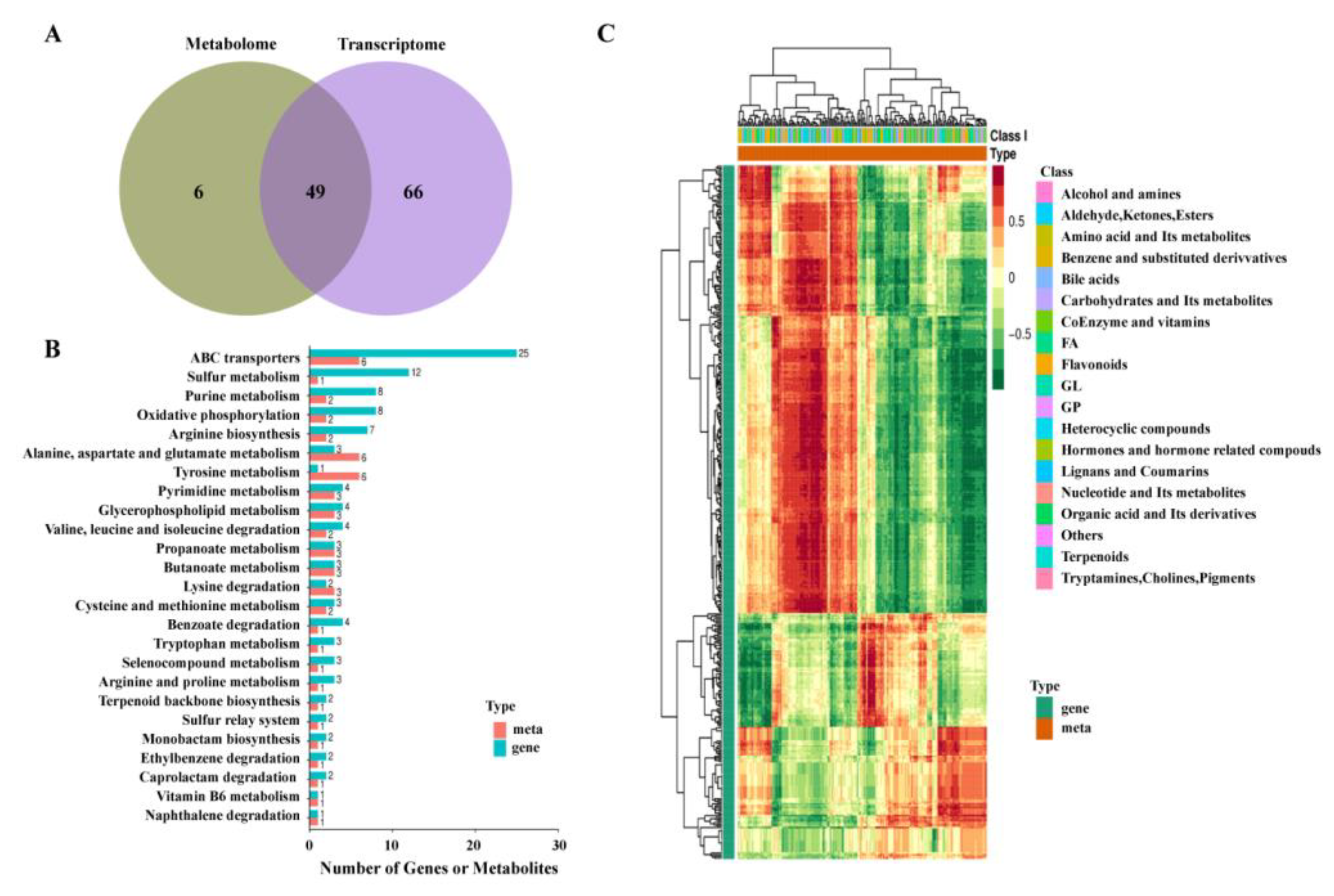
2.5. Transcriptomics Analysis
2.6. Metabolomics Analysis
2.7. Correlation Analysis of Metabolism and Transcription
3. Discussion
4. Materials and Methods
4.1. Reagents and Strains
4.2. Plant Materials and Extraction
4.3. Analysis of CBPE
4.4. Determination of CBPE Antibacterial Activity
4.4.1. Determination of the Minimum Inhibitory Concentration (MIC) and Maximum Bactericidal Concentration (MBC)
4.4.2. Growth Curve Assay
4.5. Inhibition Mechanism of CBPE
4.5.1. Observation of S. dysenteriae Morphology by Scanning Electron Microscope (SEM)
4.5.2. Observation of Bacterial Activity using Confocal Laser Scanning Microscope (CLSM)
4.5.3. Cell Viability Assay
4.5.4. Analysis of Outer/Inner Membrane Permeability
4.5.5. Detection of Cell Membrane Potential
4.5.6. Cell Wall Integrity Assay
4.5.7. Assay of Macromolecular Substances and Protein Leakage
4.5.8. Determination of K+ Content
4.5.9. Determination of ATP Content and ATPase Activity
4.6. Transcriptomic Analysis
4.7. Non-Targeted Metabolomics
4.7.1. Metabolome Sample Processing
4.7.2. UPLC-MS Analysis
4.7.3. Data Processing
4.7.4. Bioinformatics Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ranganathan, S.; Doucet, M.L.; Grassel, C.; Delaine-Elias, B.C.; Zachos, N.; Barry, E.M. Evaluating Shigella flexneri pathogenesis in the human enteroid model. Infect. Immun. 2019, 87, e00740-18. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, E.; Shirazi, F.; Khamesipour, F. Isolation and study of the antibiotic resistance properties of Shigella species in meat and meat products. J. Food Process. Preserv. 2017, 41, e12947. [Google Scholar] [CrossRef]
- Najmi, A.; Javed, S.A.; Al Bratty, M.; Alhazmi, H.A. Modern approaches in the discovery and development of plant-based natural products and their analogues as potential therapeutic agents. Molecules 2022, 27, 349. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Huang, X.; Zhang, C.; Zhang, M.; Huang, C.; Yang, H. Amino acid composition and nutritional value evaluation of Chinese chestnut (Castanea mollissima Blume) and its protein subunit. RSC Adv. 2018, 8, 2653–2659. [Google Scholar] [CrossRef] [PubMed]
- Squillaci, G.; Apone, F.; Sena, L.M.; Carola, A.; Tito, A.; Bimonte, M.; De Lucia, A.; Colucci, G.; La Cara, F.; Morana, A. Chestnut (Castanea sativa Mill.) industrial wastes as a valued bioresource for the production of active ingredients. Process Biochem. 2018, 64, 228–236. [Google Scholar] [CrossRef]
- Sorice, A.; Siano, F.; Capone, F.; Guerriero, E.; Picariello, G.; Budillon, A.; Ciliberto, G.; Paolucci, M.; Costantini, S.; Volpe, M.G. Potential anticancer effects of polyphenols from chestnut shell extracts: Modulation of cell growth, and cytokinomic and metabolomic profiles. Molecules 2016, 21, 1411. [Google Scholar] [CrossRef]
- Braga, N.; Rodrigues, F.; Oliveira, M.B.P.P. Castanea sativa by-products: A review on added value and sustainable application. Nat. Prod. Res. 2015, 29, 1–18. [Google Scholar] [CrossRef]
- Vella, F.M.; Laratta, B.; La Cara, F.; Alessandra, M. Recovery of bioactive molecules from chestnut (Castanea sativa Mill) by-products through extraction by different solvents. Nat. Prod. Res. 2018, 32, 1022–1032. [Google Scholar] [CrossRef]
- Fernández-Agulló, A.; Freire, M.S.; Antorrena, G.; Pereira, J.; González-Álvarez, J. Effect of the extraction technique and operational conditions on the recovery of bioactive compounds from chestnut (Castanea sativa) bur and shell. Sep. Sci. Technol. 2014, 49, 267–277. [Google Scholar] [CrossRef]
- Vázquez, G.; González-Álvarez, J.; Freire, M.S.; Fernández-Agulló, A.; Santos, J.; Antorrena, G. Chestnut burs as a source of natural antioxidants. Chem. Eng. Trans. 2009, 17, 855–860. [Google Scholar] [CrossRef]
- Zhao, Y.N.; Li, H.Z.; Zhang, Z.J.; Ren, Z.Q.; Yang, F.H. Extraction, preparative monomer separation and antibacterial activity of total polyphenols from Perilla frutescens. Food Funct. 2022, 13, 880–890. [Google Scholar] [CrossRef]
- Latos-Brozio, M.; Masek, A. Structure-activity relationships analysis of monomeric and polymeric polyphenols (quercetin, rutin and catechin) obtained by various polymerization methods. Chem. Biodivers. 2019, 16, e1900426. [Google Scholar] [CrossRef] [PubMed]
- Mattio, L.M.; Dallavalle, S.; Musso, L.; Filardi, R.; Franzetti, L.; Pellegrino, L.; Incecco, P.; Mora, D.; Pinto, A.; Arioli, S. Antimicrobial activity of resveratrol-derived monomers and dimers against foodborne pathogens. Sci. Rep. 2019, 9, 19525. [Google Scholar] [CrossRef] [PubMed]
- Esposito, T.; Silva, N.; Almeida, A.; Silvestre, A.; Piccinelli, A.; Aquino, R.; Sansone, F.; Mencherini, T.; Vilela, C.; Freire, C. Valorisation of chestnut spiny burs and roasted hazelnut skins extracts as bioactive additives for packaging films. Ind. Crop. Prod. 2020, 151, 112491. [Google Scholar] [CrossRef]
- Genovese, C.; Addamo, A.; Malfa, G.A.; Acquaviva, R.; Giacomo, C.D.; Tomasello, B.; Mantia, A.L.; Ragusa, M.; Toscano, M.A.; Lupo, G.; et al. Antioxidant, antimicrobial and anticancer activities of Castanea sativa (Fagaceae) extract: New therapeutic perspectives. Plant Biosyst. 2021, 155, 1032–1040. [Google Scholar] [CrossRef]
- Silva, V.; Falco, V.; Dias, M.I.; Barros, L.; Silva, A.; Capita, R.; Alonso-Calleja, C.S.; Amaral, J.; Igrejas, G.C.F.R.; Ferreira, I.; et al. Evaluation of the phenolic profile of Castanea sativa Mill. by-products and their antioxidant and antimicrobial activity against multiresistant bacteria. Antioxidants 2020, 9, 87. [Google Scholar] [CrossRef]
- Živković, J.; Zeković, Z.; Mujić, I.; Vidović, S.; Cvetković, D.; Lepojević, Z.; Nikolić, G.; Trutić, N. Scavenging capacity of superoxide radical and screening of antimicrobial activity of Castanea sativa Mill. extracts. Czech J. Food Sci. 2010, 28, 61–68. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Wu, X.; Sun, Y.; Liu, Z. Effect of thyme essential oil against Bacillus cereus planktonic growth and biofilm formation. Appl. Microb. Cell Physiol. 2018, 102, 10209–10218. [Google Scholar] [CrossRef]
- Briaud, P.; Camus, L.; Bastien, S.; Anne, D.; Vandenesch, F.; Moreau, K. Coexistence with Pseudomonas aeruginosa alters Staphylococcus aureus transcriptome, antibiotic resistance and internalization into epithelial cells. Sci. Rep. 2019, 9, 16564–16579. [Google Scholar] [CrossRef]
- Puzari, M.; Sharma, M.; Chetia, P. Emergence of antibiotic resistant Shigella species: A matter of concern. J. Infect. Public Health 2018, 11, 451–454. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Shimamoto, T. Molecular characterization of multidrug-resistant Shigella spp. of food origin. Int. J. Food Microbiol. 2015, 194, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Liu, L.; Liu, Y.; Wang, X. Ferulic acid inactivates Shigella flexneri through cell membrane destruction, biofilm retardation, and altered gene expression. J. Agric. Food Chem. 2020, 68, 7121–7131. [Google Scholar] [CrossRef] [PubMed]
- Rismondo, J.; Percy, M.G.; Gründling, A. Discovery of genes required for lipoteichoic acid glycosylation predicts two distinct mechanism for wall teichoic acid glycosylation. J. Biol. Chem. 2018, 293, 3293–3306. [Google Scholar] [CrossRef] [PubMed]
- Diao, M.M.; Qi, D.P.; Xu, M.M.; Lu, Z.X. Antibacterial activity and mechanism of monolauroyl-galactosylglycerol against Bacillus cereus. Food Control. 2018, 85, 339–344. [Google Scholar] [CrossRef]
- Huang, J.; Yang, L.; Zou, Y.; Luo, S.; Wang, X.; Liang, Y.; Du, Y.; Feng, R.; Wei, Q. Antibacterial activity and mechanism of three isomeric terpineols of Cinnamomum longepaniculatum leaf oil. Folia Microbiol. 2021, 66, 59–67. [Google Scholar] [CrossRef]
- Liu, F.; Wang, F.; Du, L.; Zhao, T.; Doyle, M.; Wang, D.; Zhang, X.; Sun, Z.; Xu, W. Antibacterial and antibiofilm activity of phenyllactic acid against Enterobacter cloacae. Food Control. 2018, 84, 442–448. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. R. 2010, 12, 9–16. [Google Scholar] [CrossRef]
- Bonora, M.; Patergnani, S.; Rimessi, A.; Marchi, E.; Suski, J.; Bononi, A.; Giorgi, C.; Marchi, S.; Missiroli, S.; Poletti, F.; et al. ATP synthesis and storage. Purinerg. Signal. 2012, 8, 343–357. [Google Scholar] [CrossRef]
- Moussatova, A.; Kandt, C.; O’Mara, M.L.; Peter, T.D. ATP-binding cassette transporters in Escherichia coli. BBA-Biochim Biophys Acta 2008, 1778, 1757–1771. [Google Scholar] [CrossRef]
- Fernández, L.; Hancock, R. Adaptive and mutational resistance: Role of porins and efflux pumps in drug resistance. Clin. Microbiol. Rev. 2012, 25, 661–681. [Google Scholar] [CrossRef]
- Klein, J. Membrane breakdown in acute and chronic neurodegeneration: Focus on choline-containing phospholipids. J. Neural Transm. 2000, 107, 1027–1063. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.X.; Yuan, Y.H.; Jia, H.; Zeng, X.J.; Wang, Z.L.; Hu, Z.Q.; Gao, Z.P.; Yue, T.L. Antimicrobial and anti-biofilm activity of thymoquinone against Shigella flexneri. Appl. Microbiol. Biotechnol. 2021, 105, 4709–4718. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.J.; Jia, J.J.; Zhang, B.K.; Mi, S.J.; Zhang, L.; Xie, X.M.; Guo, H.C.; Shi, J.S.; Tu, C.C. Serum metabolomic profiling of piglets infected with virulent classical swine fever virus. Front. Microbiol. 2017, 8, 731. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Xu, F.; Zhang, X.; Shao, X.F.; Wei, Y.Y.; Wang, H.F. Transcriptomic analysis reveals antibacterial mechanism of flavonoids from Sedum aizoon L. against Pseudomonas fragi. Food Control 2022, 134, 108755. [Google Scholar] [CrossRef]
- Liu, M.M.; Feng, M.X.; Yang, K.; Cao, Y.F.; Zhang, J.; Xu, J.N.; Hernández, S.H.; Wei, X.Y.; Fan, M.T. Transcriptomic and metabolomic analyses reveal antibacterial mechanism of astringent persimmon tannin against Methicillin-resistant Staphylococcus aureus isolated from pork. Food Chem. 2020, 309, 125692. [Google Scholar] [CrossRef]
- Johnson, L.; Mulcahy, H.; Kanevets, U.; Shi, Y.; Lewenza, S. Surface-localized spermidine protects the Pseudomonas aeruginosa outer membrane from antibiotic treatment and oxidative stress. J. Bacteriol. 2012, 194, 813–826. [Google Scholar] [CrossRef]
- Huang, Z.; Maness, P.C.; Blake, D.M.; Wolfrum, E.J.; Smolinski, S.L.; Jacoby, W.A. Bactericidal mode of titanium dioxide photocatalysis. J. Photochem. Photobiol. A 2000, 130, 163–170. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, F.; Chen, L.; Wang, X.; Yu, Z.; Cheng, C.; Yang, Y. Effect of Chestnut (Castanea Mollissima Blume) Bur Polyphenol Extract on Shigella dysenteriae: Antibacterial Activity and the Mechanism. Molecules 2023, 28, 6990. https://doi.org/10.3390/molecules28196990
Peng F, Chen L, Wang X, Yu Z, Cheng C, Yang Y. Effect of Chestnut (Castanea Mollissima Blume) Bur Polyphenol Extract on Shigella dysenteriae: Antibacterial Activity and the Mechanism. Molecules. 2023; 28(19):6990. https://doi.org/10.3390/molecules28196990
Chicago/Turabian StylePeng, Fei, Linan Chen, Xiuping Wang, Zuoqing Yu, Caihong Cheng, and Yuedong Yang. 2023. "Effect of Chestnut (Castanea Mollissima Blume) Bur Polyphenol Extract on Shigella dysenteriae: Antibacterial Activity and the Mechanism" Molecules 28, no. 19: 6990. https://doi.org/10.3390/molecules28196990
APA StylePeng, F., Chen, L., Wang, X., Yu, Z., Cheng, C., & Yang, Y. (2023). Effect of Chestnut (Castanea Mollissima Blume) Bur Polyphenol Extract on Shigella dysenteriae: Antibacterial Activity and the Mechanism. Molecules, 28(19), 6990. https://doi.org/10.3390/molecules28196990