1. Introduction
Sarcoptic mange, also known as scabies in human patients, is an extremely contagious skin condition caused by
Sarcoptes scabiei, an astigmatic mite that burrows into the epidermis, actively penetrating the stratum corneum [
1]. The adult mites mate, and the females lay eggs in the skin. The hatched larvae create small burrows, referred to as molting pouches, where they molt into nymphs and eventually mature into adults. These parasites have a worldwide distribution, affecting over 150 host species, and they demonstrate remarkable epidemiological adaptability, enabling transmission among diverse hosts [
2]. The disease can also manifest as a mild infection in animals, presenting symptoms such as itching papules, erythema, scales, and alopecia. In chronic cases, hyperkeratosis and/or exudative crust formation may be observed [
3,
4].
Sarcoptic mange is an exceptionally contagious skin disease that spreads through direct skin-to-skin contact, fomites or contact with contaminated environments frequented by heavily affected hosts [
5,
6]. Nymphs and females have remarkable survival capabilities off the host, lasting up to 21 days, and exhibit higher resistance compared to larvae and males [
7,
8]. Thus, the implementation of biocides and repellents in the environment becomes paramount. Wildlife, unfortunately, is particularly susceptible to sarcoptic mange, and its outbreaks can lead to heightened morbidity and even fatal consequences [
9], especially when naive populations are affected [
10]. This disease is rapidly emerging [
11] and has been implicated in the decline of wildlife populations [
12,
13], resulting in reduced reproduction rates and triggering mass mortality events [
14,
15].
The
Rhipicephalus microplus tick (Acari:
Ixodidae) is a profoundly impactful tick species with devastating effects on bovine well-being and productivity. Its widespread presence in various tropical and subtropical regions worldwide, particularly in Pakistan and India [
16], poses a severe threat to cattle populations. Infestations by these ticks lead to blood loss, resulting in anemia and substantial economic losses due to reduced growth and production. Moreover, these ticks serve as vectors for various diseases, including
Babesia bigemina and
Anaplasma marginale, which cause economically significant and very deadly illnesses, particularly in high-yielding crossbred and exotic cattle herds [
17]. Ticks and tick-borne illnesses generate an estimated USD 13.9 to 18.7 billion loss in cattle alone, with an annual deficit of nearly 3 billion pieces of hide and skin [
18]. Therefore, it is of the highest significance to create highly efficient ways for managing cattle ticks and guaranteeing optimum health and productivity in dairy animals.
Monotheca buxifolia, a member of the
Sapotaceae family, is a very important indigenous evergreen tree or thorny shrub known for its tasty fruits that grows in a variety of habitats including northwest Pakistan, Afghanistan, and Iran. This excellent plant performs a variety of essential functions, including feed, fuel, tiny wood, and fences. Its importance extends to traditional medicine, where the fruit has been valued for its digestive, hematinic, vermicide, and laxative characteristics, and the aerial portions are used to treat rheumatism and urinary tract infections [
19]. The plant extracts have been scientifically proven to have a wide range of beneficial properties, including “hepatoprotective, urease inhibitory, reno-protective, antidepressant, antiproliferative, antibacterial, antioxidant, anxiolytic, antimalarial, analgesic, antipyretic, and anti-inflammatory activities” [
19,
20,
21,
22,
23,
24].
Aspartic protease is an enzymatic protein responsible for catalyzing the hydrolysis of peptide bonds with the aid of a trapped water molecule [
25]. These proteases contain a pair of aspartic acid residues at their active site, facilitating various crucial physiological processes in parasitic organisms, including “tissue invasion, migration, digestion, molting, and reproduction”. For ticks, hemoglobin serves as a primary nutrient that is crucial for their development and reproduction. Consequently, the digestion of hemoglobin is a vital metabolic pathway occurring within digestive vesicles absorbed into gut epithelial cells through the action of aspartic proteases [
26]. In a
S. scabiei cDNA library, researchers identified a single aspartic protease, and functional assays revealed its ability to digest human “hemoglobin, serum albumin, fibronectin, and fibrinogen” while not affecting collagen III or laminin. The presence of this aspartic protease (SsAP) opens up possibilities for interfering with its function, which may have implications for mite survival [
27].
In this context, the present study aimed to assess the potential of M. buxifolia plant extract to inhibit ticks and mites in vitro, as well as to investigate the inhibitory interactions between the plant’s phytochemical and the aspartic protease proteins of ticks and mites using in silico methods. It is worth noting that, prior to our research, there has been limited exploration of M. buxifolia plant extract for its acaricidal potential against these ticks and mites. Furthermore, the specific phytochemicals derived from this plant have not been previously utilized in docking experiments aimed at understanding their interactions with the target proteins in ticks and mites. This novelty adds significance to our study, as it contributes to the emerging knowledge about the potential use of M. buxifolia in pest control and sheds light on the underlying molecular mechanisms.
3. Discussion
The necessity for tick management puts the dairy industry’s viability at risk in locations where ticks thrive and proliferate [
34]. The same is the case for mite infestations. Chemical acaricides are often employed for this purpose, although [
35] revealed that the development of acaricide resistance in tick species is a serious problem. Furthermore, the use of chemical acaricides may pollute the environment and contaminate cattle meat and milk, as well as promote tick and mite resistance [
36]. In response to these issues, there is increased interest in the use of natural plant-based tick-management solutions. Many studies have looked at the efficiency of plant extracts and phytochemicals as acaricides, and the findings have been encouraging [
25].
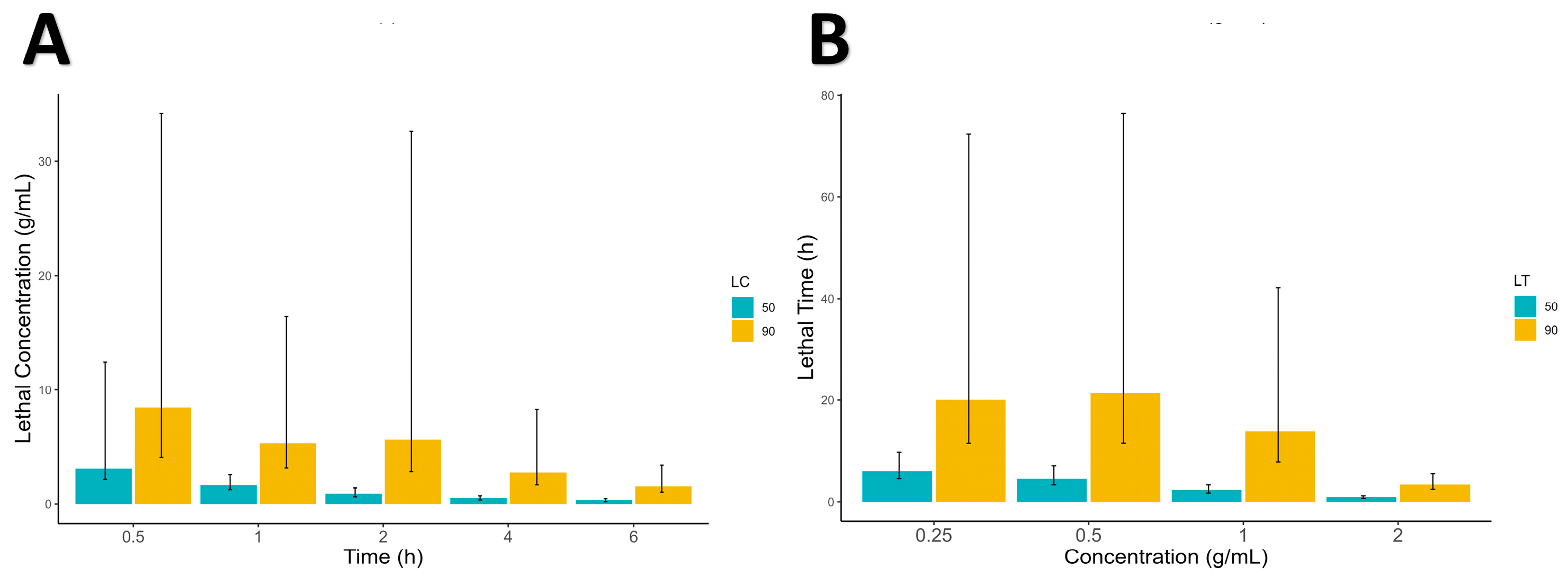
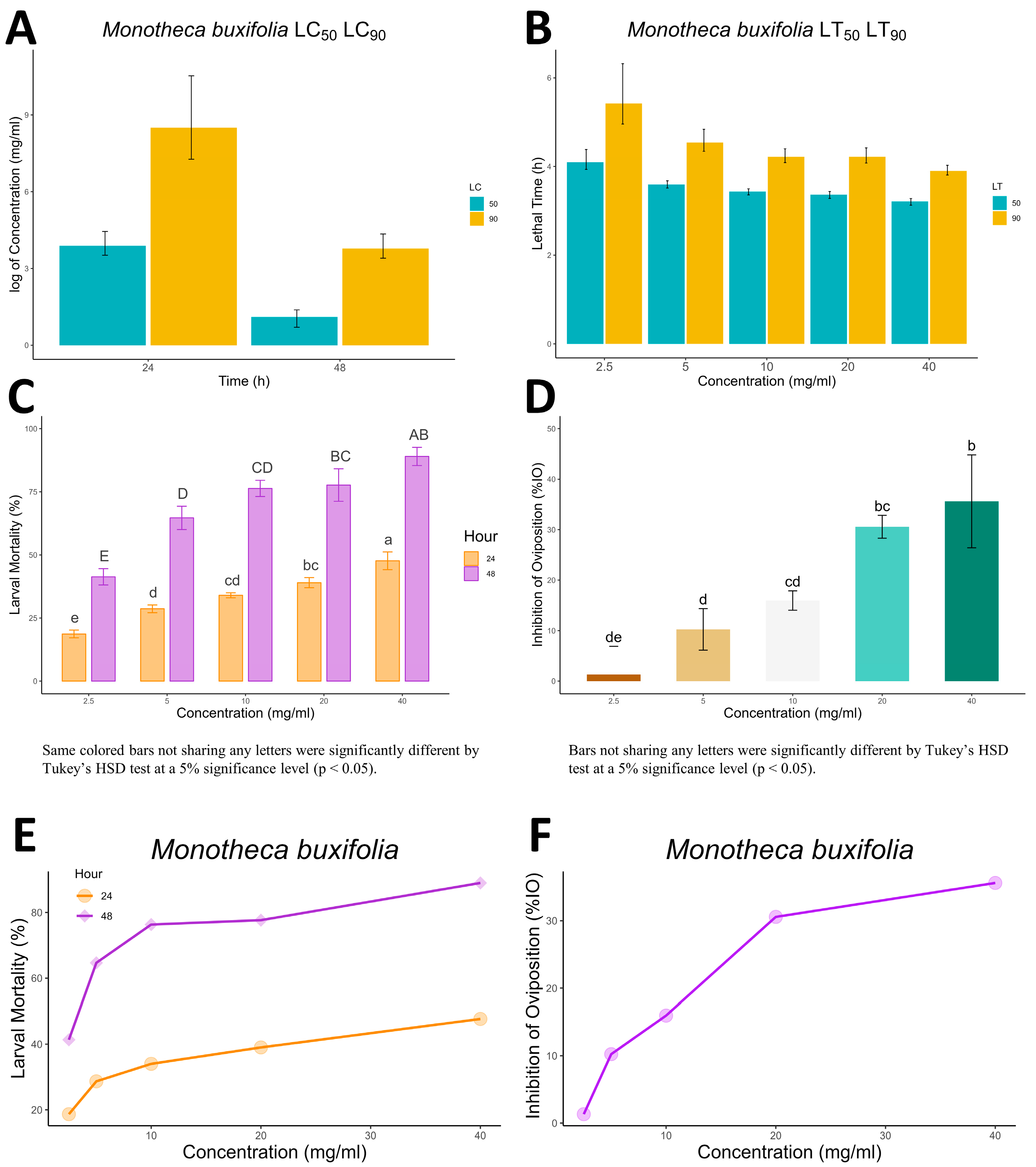
The current study revealed that the mean mortality of adult ticks was increased significantly with increased dosage (concentration) and exposure time after in vitro treatment for the tested botanicals. The same was the case for
S. scabiei mites, whose mortality was also time- and dose-dependent. These results are in line with the findings of [
37,
38], in which the mortality effect of botanicals was indicated to be dose-, concentration- and exposure-time-dependent. Our study also revealed that all methanolic extracts of the tested botanical leaves at the tested concentrations induced a significant acaricidal effect against
S. scabiei mites and
R. microplus compared with the negative control.
Lead and target identification, followed by lead optimization, was a lengthy and costly process in the traditional drug development cycle. In the modern age, computational biology has created a low-cost, rapid method for finding novel drugs [
39]. Potential therapeutic candidates for use against our target proteins are identified using molecular docking, an analysis of the likely binding affinities of two structures. The aspartic protease enzymes of
S. scabiei mites and
R. microplus were found to play an essential role in the survival and sustenance of the parasites by facilitating the breakdown of host hemoglobin during the blood-feeding process [
27,
40]. Therefore, preventing ticks and mites from producing aspartic protease may disrupt their blood-feeding process and perhaps restrict their survival and reproductive success by preventing them from digesting host hemoglobin. This method has the potential to be investigated as a means of decreasing the harm ticks and mites do to their hosts. Butelin, Lupenol, and Buterol are examples of phytochemicals. Alpha-Indole-3-deoxyestradiol, 13-cis-epoxydocosanoic acid, 14-epoxydocosanoic acid, tocopherol, 5HTP, Ascorbic Acid, and both 6-stearate and 3,4,4-trimethyl-5-pyrazolone isolated from
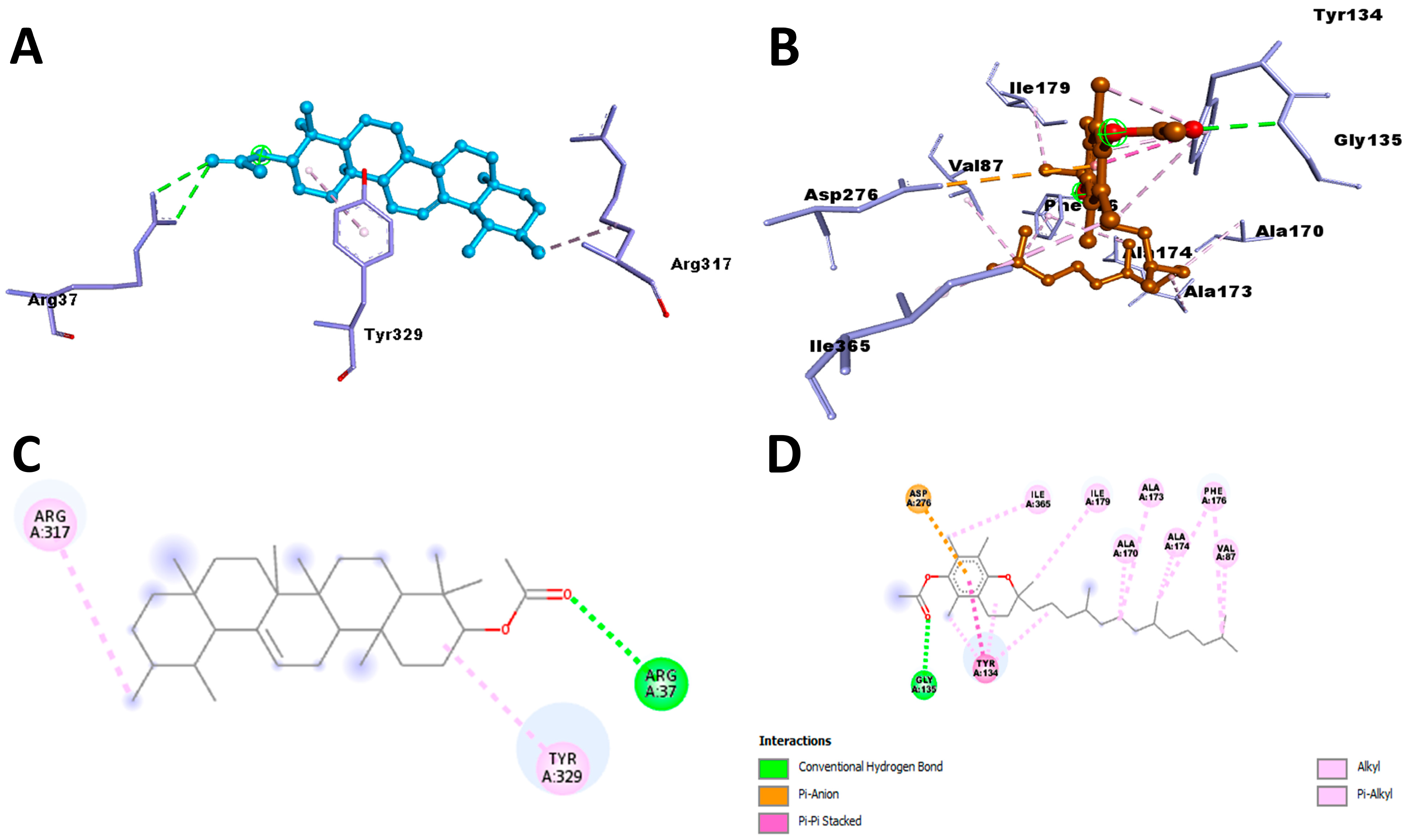
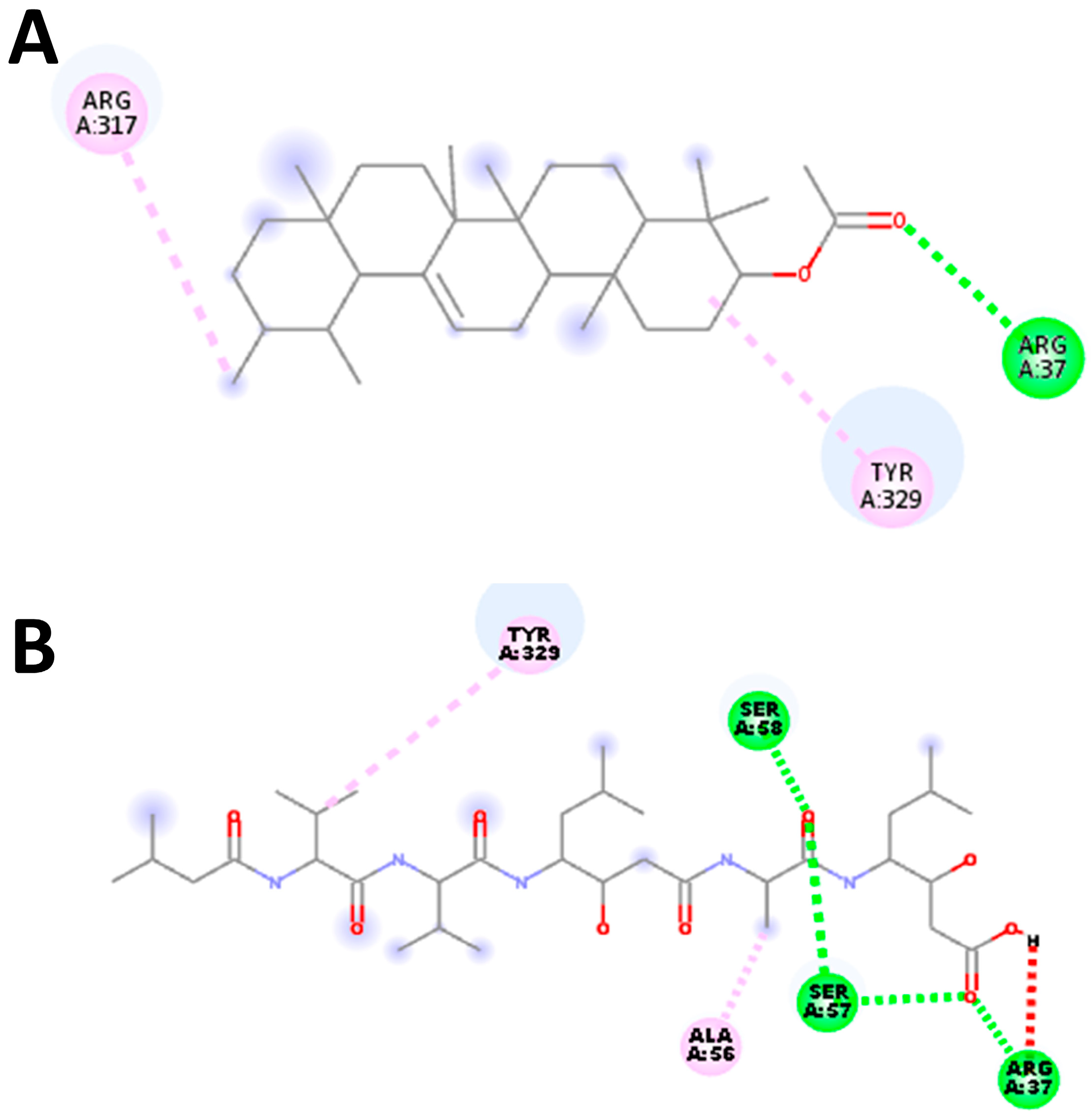
M. buxifolia have been shown to meet all the requirements for classification as pharmaceuticals. Thus, the aforementioned phytochemicals were used in docking experiments with the aspartic protease parasite enzymes. Since we wanted to stop these specific enzymes from working, we bound inhibitors to their active sites. All of the phytochemicals were shown to have stronger affinities to the active pockets and lower binding energies compared to the synthetic drug. Furthermore, the existence of typical hydrogen bonds in the protein–ligand complex’s 2D bond contacts demonstrated robust binding. Aspartic protease inhibitory action was shown by plant-derived compounds with significant affinities, such as Alpha-Amyrenyl acetate against
R. microplus and alpha-Tocopherol against
S. scabiei.
4. Materials and Methods
4.1. Plant Collection, Identification and Extract Preparation
Aerial parts from
M. buxifolia were collected from Abdul Wali Khan University Mardan’s (AWKUM) botanical garden in Khyber Pakhtunkhwa (coordinates: 34.1917° N, 72.0347° E). The plant materials were inspected for damage and rinsed. The leaves were then taken to the herbarium of the Department of Botany, AWKUM, and assigned the accession number Awkum.Bot. They were then air-dried for 2 weeks. After 15 days, the dried leaves were ground into powder, and stock solutions were prepared as described by [
34]. The resulting solution was concentrated, yielding a stock extract for further dilutions and tests. For the mite bioassay, the stock solution was diluted to 0.25, 0.5, 1, and 2 g/mL, whereas for the adult immersion test (AIT) and the larval packet test (LPT), the concentrations were diluted to 2.5, 5, 10, 20, and 40 mg/mL concentrations.
4.2. Collection and Identification of Mites
Mites were collected from rabbits at AWKUM’s rabbit farms, where hay served as bedding and was changed daily. Upon detecting signs of mange, skin scraps were taken from the rabbits using the method described by [
23] and immediate treatment was administered. The skin scraps were then examined under a compound microscope to identify
S. scabiei var cuniculi.
4.3. Collection and Identification of Ticks
In strict adherence to the guidelines outlined by the World Association for the Advancement of Veterinary Parasitology [
41] a careful manual collection of fully engorged
R. microplus ticks was conducted from cattle and buffaloes at various farms located in Mardan, Khyber Pakhtunkhwa, with precise coordinates provided as 34.1986° N, 72.0404° E. Subsequently, the ticks underwent a comprehensive cleaning process and were morphologically identified as
R. microplus through the utilization of standard tick identification keys, employing a microscope [
42]. A total of 300 adult engorged female ticks were carefully selected for inclusion in the study. Some of these ticks were employed to obtain larvae for the larval packet test, while the remaining ticks were carefully divided into separate groups to conduct the adult immersion test, with a focus on evaluating the acaricidal effects of a fungal extract.
4.4. Contact Bioassay
The study was first approved by the ethical committee of AWKUM. Skin scraps were taken from the infested rabbits raised at the rabbit form adhering to the Guide for the Care and Use of Laboratory Animals (8th edition) [
43]. The infested skin was first briefly cleaned and then scraped into a micro Petri plate using sterile surgical blade until the skin appeared red. The Petri plate containing skin scraps was incubated at 37 °C for the mites to emerge out of the skin scraps. The experimental setup involved the inoculation of ten mites into individual Petri plates using a fine needle. Following that, 0.5 mL samples of the plant extract were introduced directly onto the mites within the Petri plates. This procedure was carried out independently for each concentration of the extract, and each concentration was replicated three times.
4.5. Adult Immersion Test and Larval Packet Test
The extracts efficacy against
R. microplus ticks was determined using the adult immersion test (AIT) and larval packet test (LPT) according to the protocol outlined by Ayub et al, [
37] and Matos et al. [
44]. The tick’s larval mortality was recorded at 24 and 48 h for different concentrations. Similarly, female ticks and their laid eggs were weighed in AIT. The effectiveness of the crude extracts in AIT was evaluated by calculating the percent inhibition of oviposition (%
IO) using the following formula [
45]:
where egg-laying index = mean weight of eggs laid ÷ mean weight of engorged females.
4.6. Sequence Retrieval
The amino acid sequences of aspartic protease protein from S. scabiei (accession numbers: V5NEJ5) and R. microplus aspartic proteinase protein with accession number C3UTE0 were obtained from the UniProtKB database. Both the protein sequences were retrieved in FASTA format and used for further analysis.
4.7. Characterization of the Physicochemical Properties
The protein’s physicochemical characterization required the calculation of many critical characteristics using well-established methodologies. Expasy’s ProtParam server [
46] was used to calculate the theoretical isoelectric point (pI), molecular weight, total number of positive and negative residues, extinction coefficient [
47], instability index [
48], aliphatic index [
49], and grand average hydropathy (GRAVY) [
50]. The corresponding results can be found in the provided table.
4.8. Functional Characterization
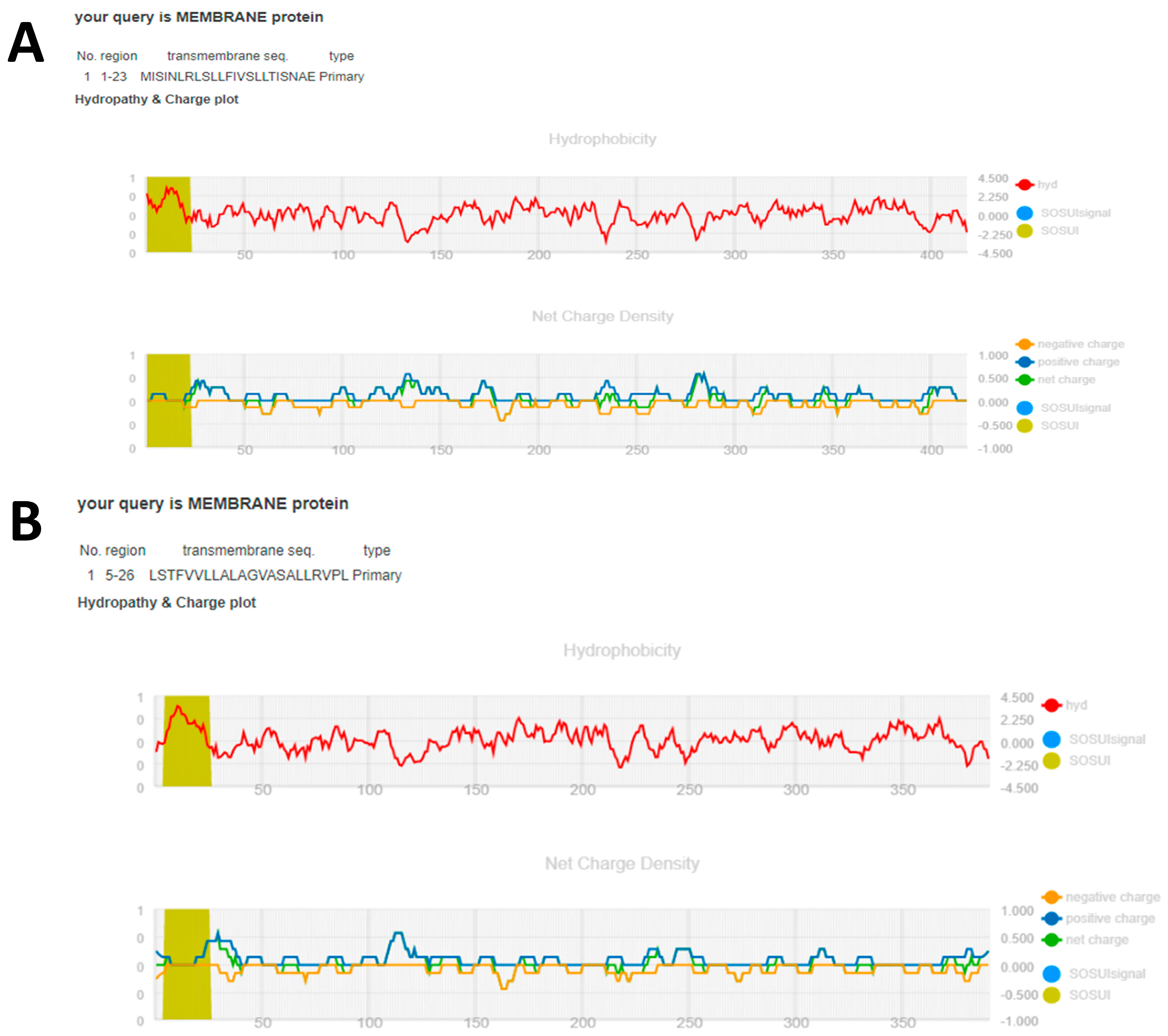
In this study, the SOSUI server (
https://harrier.nagahama-i-bio.ac.jp/sosui/, accessed on 22 May 2023 [
51]) was employed to assess the amphiphilicity index and hydropathy index of the aspartic protease protein found in
R. microplus and
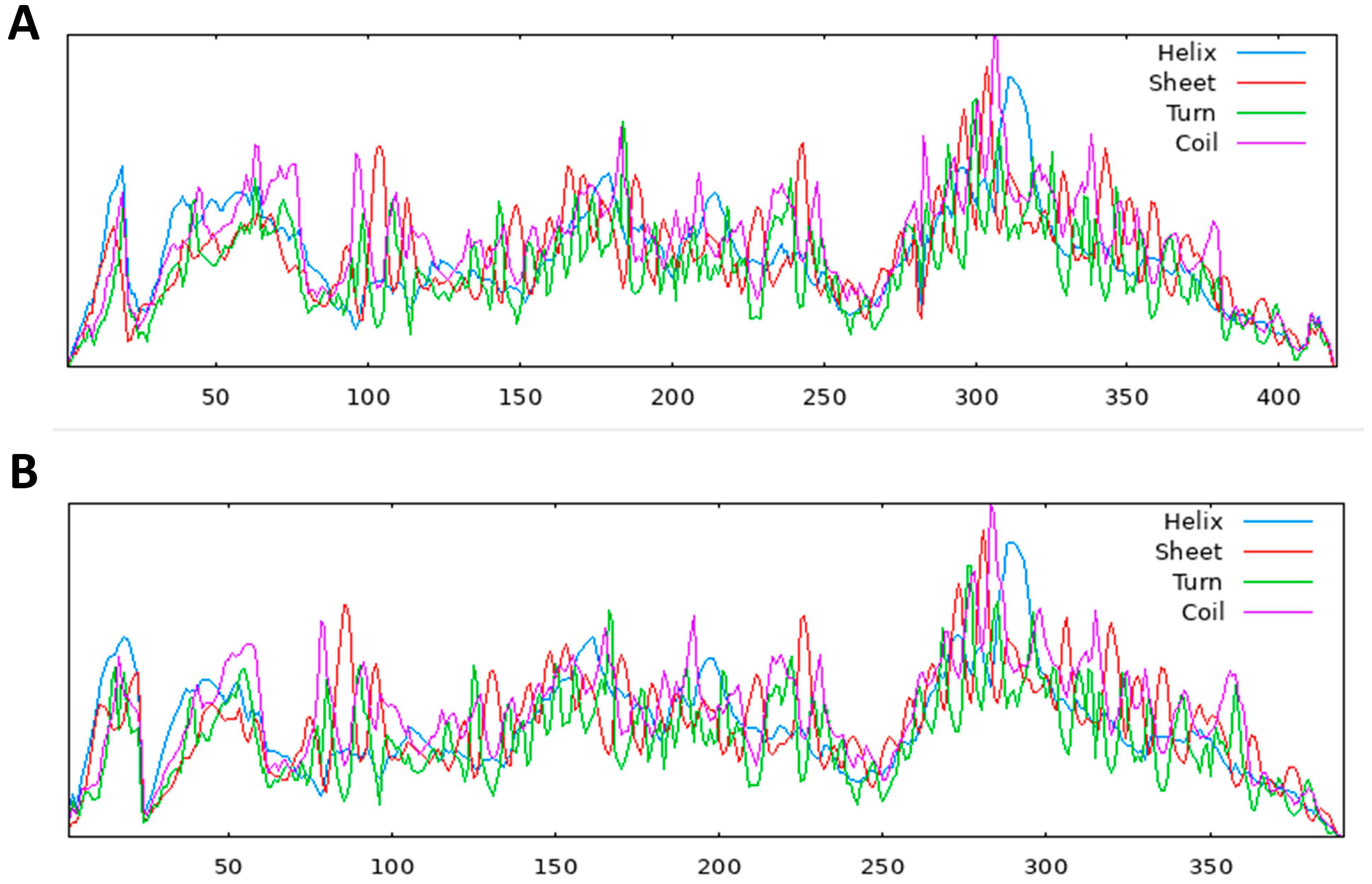
S. scabiei mite. These indices were used to estimate whether the protein was in the cytoplasm or bridged the transmembrane. Furthermore, the SOSUI server’s numerous tools, such as SOSUI (Batch), SOSUIsignal, SOSUIgramN, and SOSUImp1, were used to predict parts of the secondary structure of the aspartic protease protein from its supplied amino acid sequence [
52]. This prediction helps in locating the hypothetical protein inside a cell.
4.9. Secondary Structure Prediction
4.10. Building and Evaluation of the 3D Structure Model
The FASTA sequences of the target proteins were submitted to the SWISS-MODEL server (
https://swissmodel.expasy.org/, accessed on 25 May 2023) for prediction of the 3D structure model using automated comparative modeling as described by [
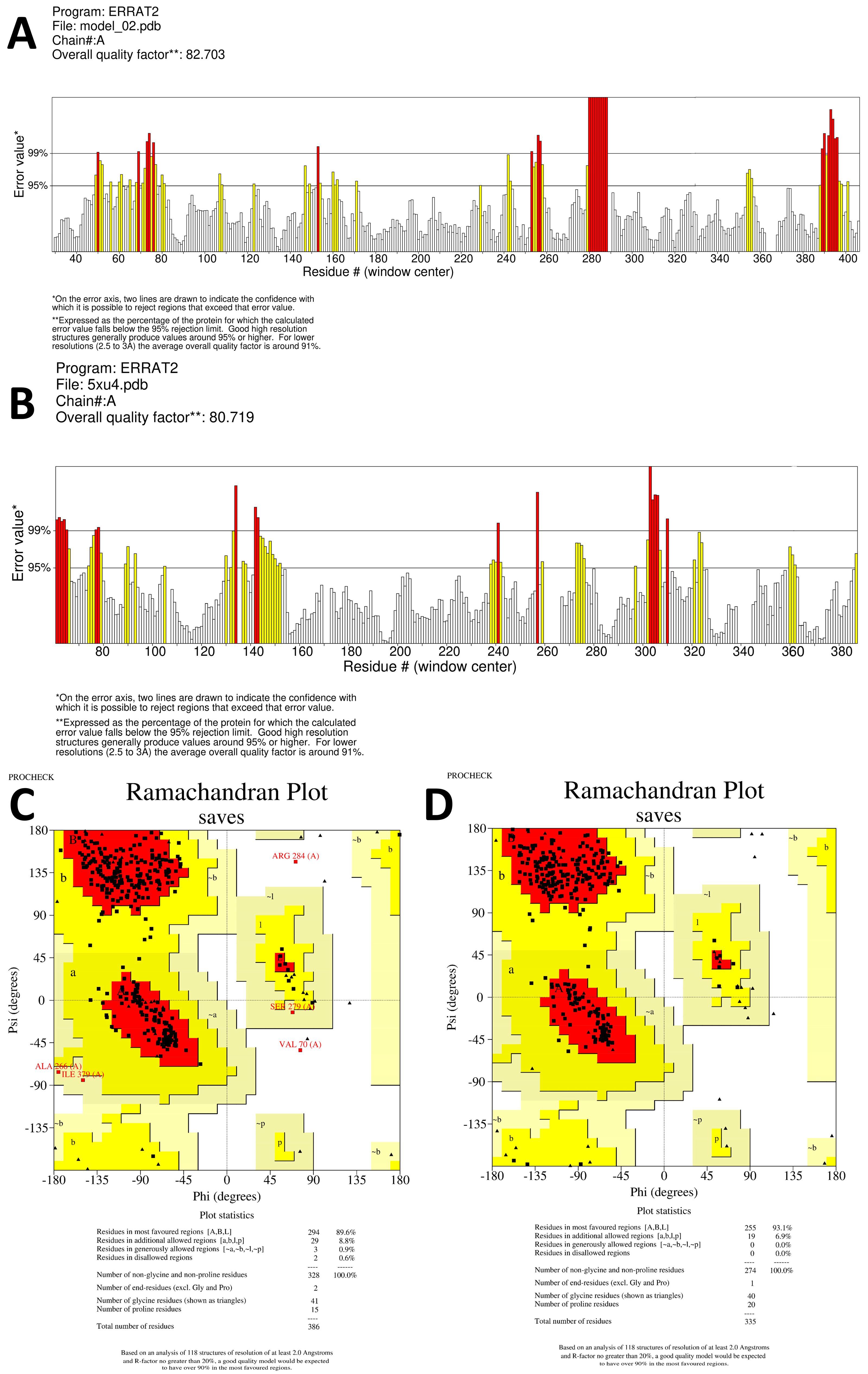
53]. To ensure the reliability of the predicted 3D structure model for the hypothetical protein, multiple quality assessment tools were employed. Firstly, the 3D model quality was assessed using SAVES (
https://saves.mbi.ucla.edu/, accessed on 26 May 2023, which is a comprehensive platform for protein structure validation. The Ramachandran plot was constructed using the PROCHECK server [
54] to visualize the distribution of backbone dihedral angles (ψ against φ) for all amino acid residues in the protein structure. The analysis of this plot helps to evaluate the stereochemical quality and identify any potential deviations from ideal conformations in the model. Furthermore, the protein structure was cross-validated using the ERRAT server (
https://services.mbi.ucla.edu/ERRAT, accessed on 1 June 2023). This tool assesses the statistics of nonbonded interactions between different atom types within a 9-residue sliding window [
55]. The plot of the error function versus the window position provides valuable insights into the accuracy and consistency of the protein crystallographic structure. By integrating the results from these various assessment methods, the reliability and accuracy of the predicted 3D structure model for the hypothetical protein were thoroughly verified.
4.11. Active Site Determination
The use of the Computed Atlas of Surface Topography of Proteins (CASTp) server (
http://sts.bioe.uic.edu/castp/, accessed on 2 June 2023) has proven to be an immensely valuable and indispensable tool for determining the active sites of proteins. Through CASTp, a highly detailed, comprehensive, and quantitative analysis of the topographical features of proteins is achieved. This analysis enables the precise localization and measurement of active pockets on both the protein’s surface and its interior 3D structure. As a result, CASTp facilitates the accurate prediction of critical regions and key residues within the protein that play a significant role in interacting with ligands [
56]. To enhance the understanding and interpretation of the CASTp results, the visualization of the outcomes has been effectively accomplished using the PyMOL (version 2.5.4) software.
4.12. Ligand Preparation
The thirteen structures of chemical constituents of
M. buxifolia, Alpha-Amyrenyl acetate, Lupenol, Butelin, 3-Deoxyestradiol, cis-13,14-Epoxydocosanoic acid, Indole, Alpha-Tocopherol, 5-Hydroxytryptamine, Ascorbyl 6-stearate and 3,4,4-Trimethyl-5-pyrazolone, were collected from published literatures [
21,
57]. The ligands’ 2D chemical structures were drawn using ChemDraw Ultra 2008, and their energy minimization was performed using Chem3D Ultra. The resulting ligand structures were saved in .pdb format.
4.13. Molecular Docking Analysis
The binding mode and interaction of target proteins with individual chemical constituents of M. buxifolia were thoroughly investigated using AutoDock Vina software (version 1.1.2). The docking process aimed to explore a range of possible conformations and orientations for the ligands at the binding sites. Prior to docking, the protein structures were prepared in PyRx (version 0.8) software, generating PDBQT files with added hydrogen atoms to all polar residues. The ligands’ bonds were set to be rotatable to enhance flexibility during docking. To carry out the protein-fixed ligand-flexible docking, the Lamarckian Genetic Algorithm (LGA) method was employed for all calculations. The binding site on the target proteins, S. scabiei aspartic proteinase protein and R. microplus aspartic proteinase protein, was defined by establishing a grid box with a grid spacing of 0.375 Å. For the S. scabiei aspartic proteinase protein, the grid box was centered on coordinates X: 21.8344, Y: −5.6644 and Z: 19.7712 Å, while for the R. microplus aspartic proteinase protein, it was centered on coordinates X: 21.6564, Y: −10.6987 and Z: 22.5169 Å. To ensure the reliability of the results, ten runs with AutoDock Vina were performed for each ligand structure in all cases, saving the best pose from each run. The final affinity value was determined by averaging the affinity of the best poses obtained in the ten runs. The analysis of protein–ligand conformations, including interactions such as hydrogen bonds and bond lengths, was conducted using PyMol (version 2.5.4) and Discovery Studio Visualizer (version 4.5), allowing for a comprehensive examination of the complex structures and their interactions.
4.14. In Silico Docking Protocol Validation
The validation of our docking protocol aimed to establish its precision and reliability in replicating the binding model and molecular interactions observed in experimentally modeled protein structures during our current in silico studies. To validate the procedure, we utilized an aspartic protease inhibitor as a test case. The validation process involved manually removing modeled proteins and redocking the inhibitor into the active site using AutoDock Vina (version 1.1.2) software. This entailed extracting inhibitor heteroatoms from the protein complex, saving them as a separate inhibitor in PDB format, while keeping the grid parameters consistent. This rigorous validation ensured the accurate binding of the inhibitor to the active site cleft, with minimal deviation compared to the actual co-crystallized complex. Subsequently, we superimposed the redocked complex onto the reference co-crystallized complex using PyMOL 2.3, calculated the root mean square deviation (RMSD), and generated a 2-dimensional image highlighting the relevant amino acid residues with Discovery Studio (version 4.5) software. This process ensured the reliability of our docking methodology and its fidelity in reproducing binding characteristics observed in experimental crystallographic data.
4.15. Statistical Analysis
All statistical approaches were made in R and RStudio. The data were first arranged in Microsoft Excel (v 2302) and then imported into the R working environment for further statistical analysis. Descriptive statistics of the data were calculated and presented in a table with mean ± standard deviation. The significant difference between the different concentrations were calculated using one-way ANOVA (analysis of variance) followed by the Tukey honesty significance difference (HSD) test. Furthermore, 50% and 90% lethal concentration and lethal time (LC50, LC90 and LT50, LT90) were calculated in R using the “ecotox” package, and all the data were graphically presented using “ggplot2 and ggpubr” R package.



 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}