
Antioxidant and Anti-Aging Phytoconstituents from Faucaria tuberculosa: In Vitro and In Silico Studies


 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Evaluation of TPC and TFC of Ethanolic Extract
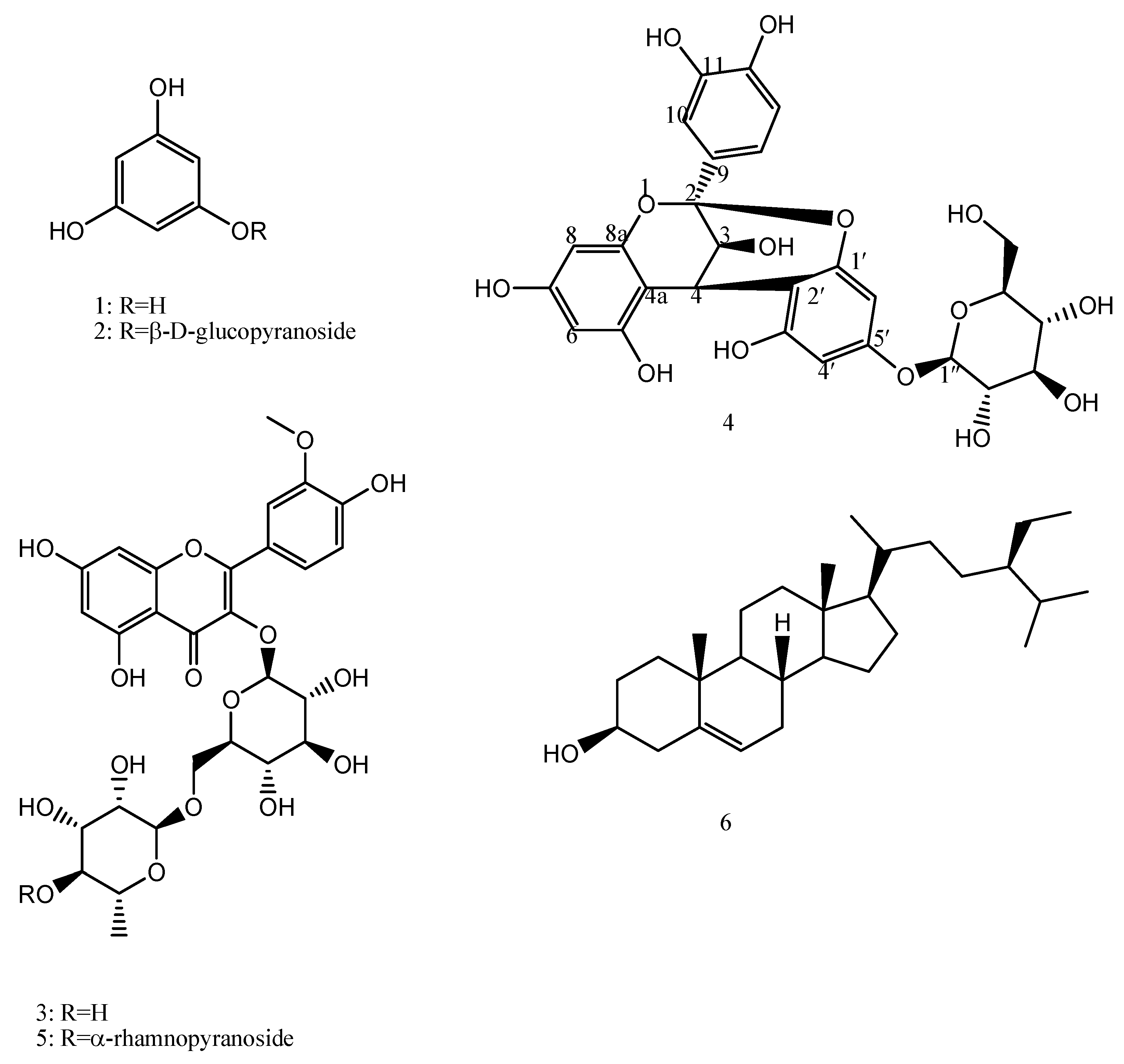
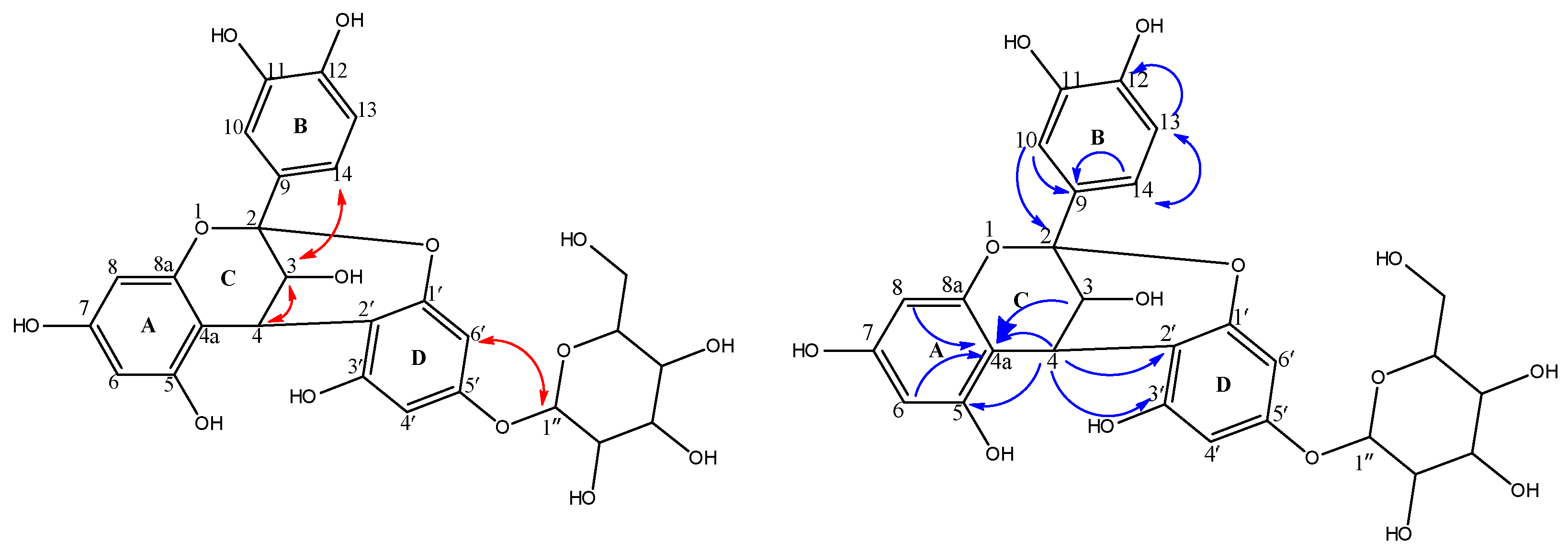
2.2. Spectroscopic Data of Isolated Compounds
2.3. Antioxidant Activities of Isolated Compounds
2.4. Anti-Aging Activities of Isolated Compounds
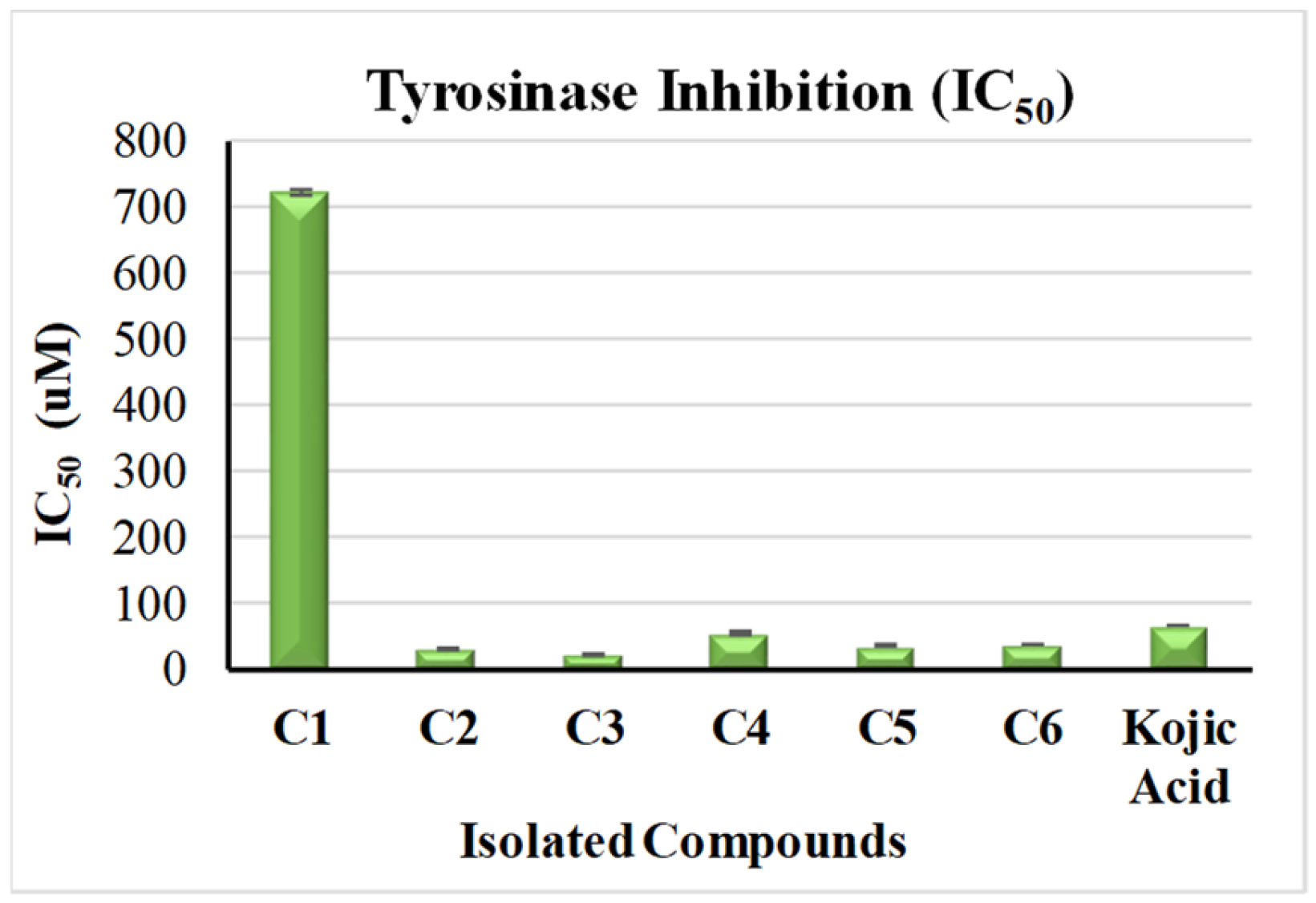
2.4.1. Tyrosinase Inhibitory Assay
2.4.2. Hyaluronidase Inhibitory Assay
2.5. In Silico Molecular Docking
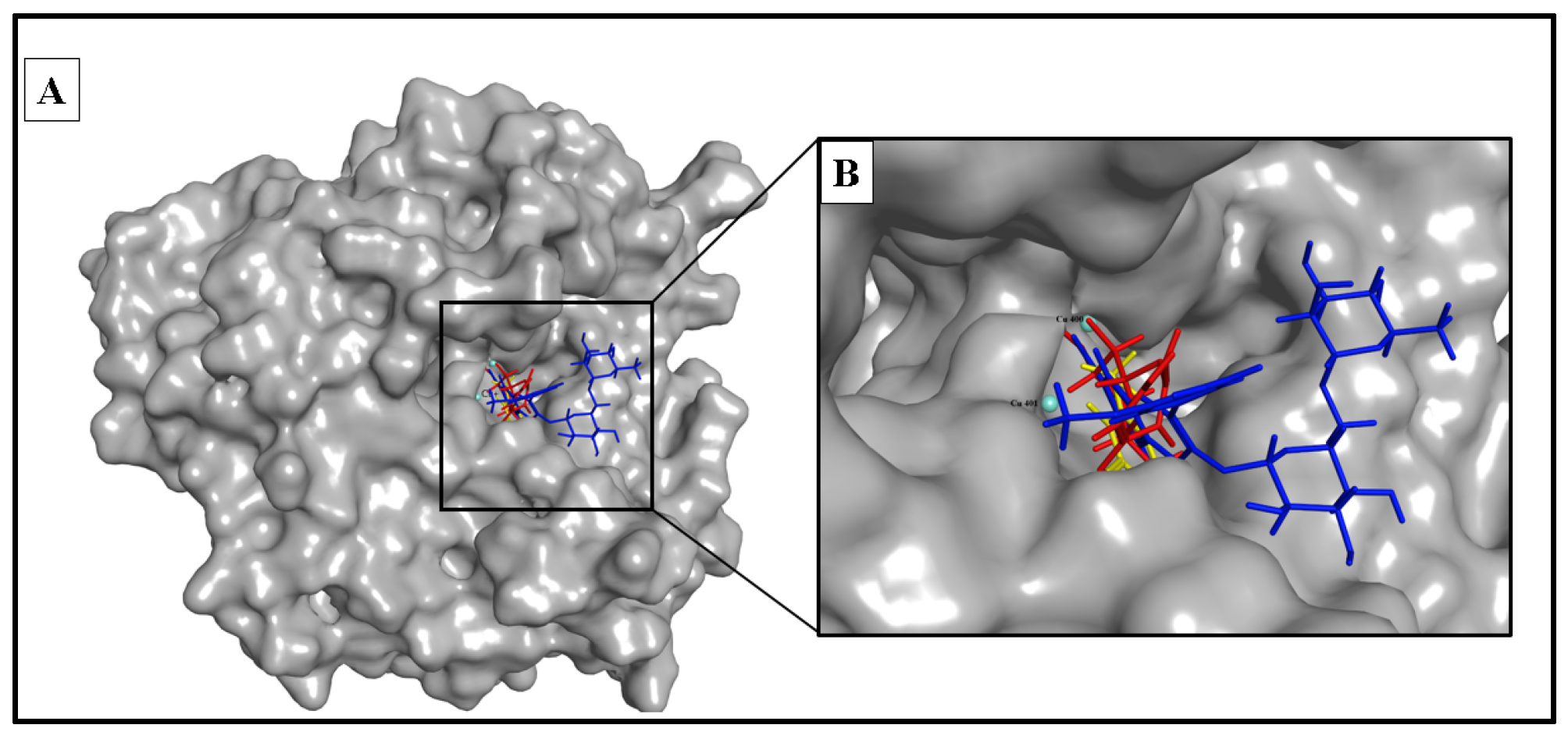
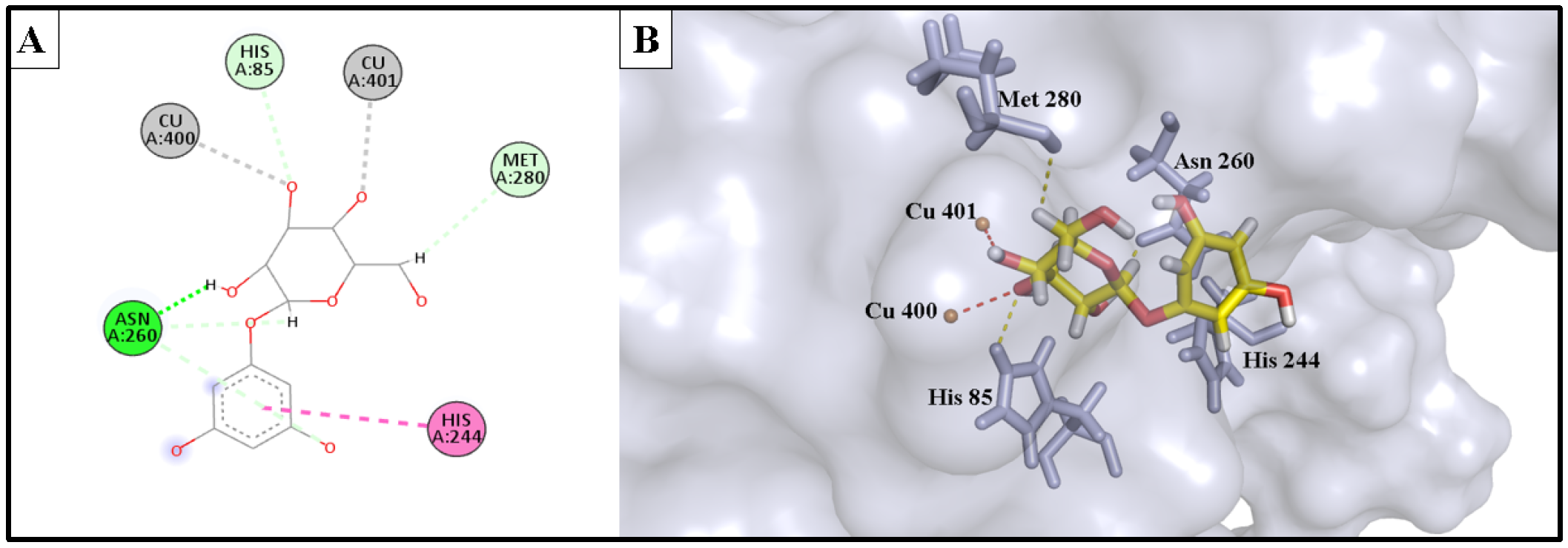
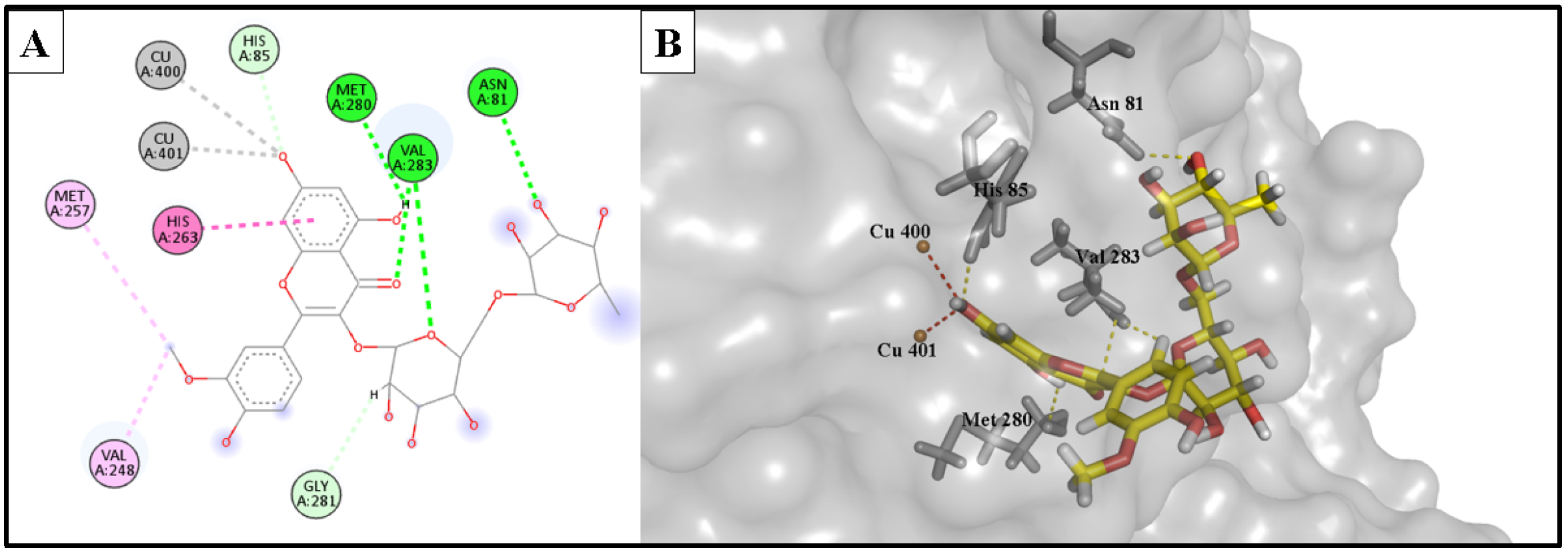
2.5.1. Anti-Tyrosinase Activity
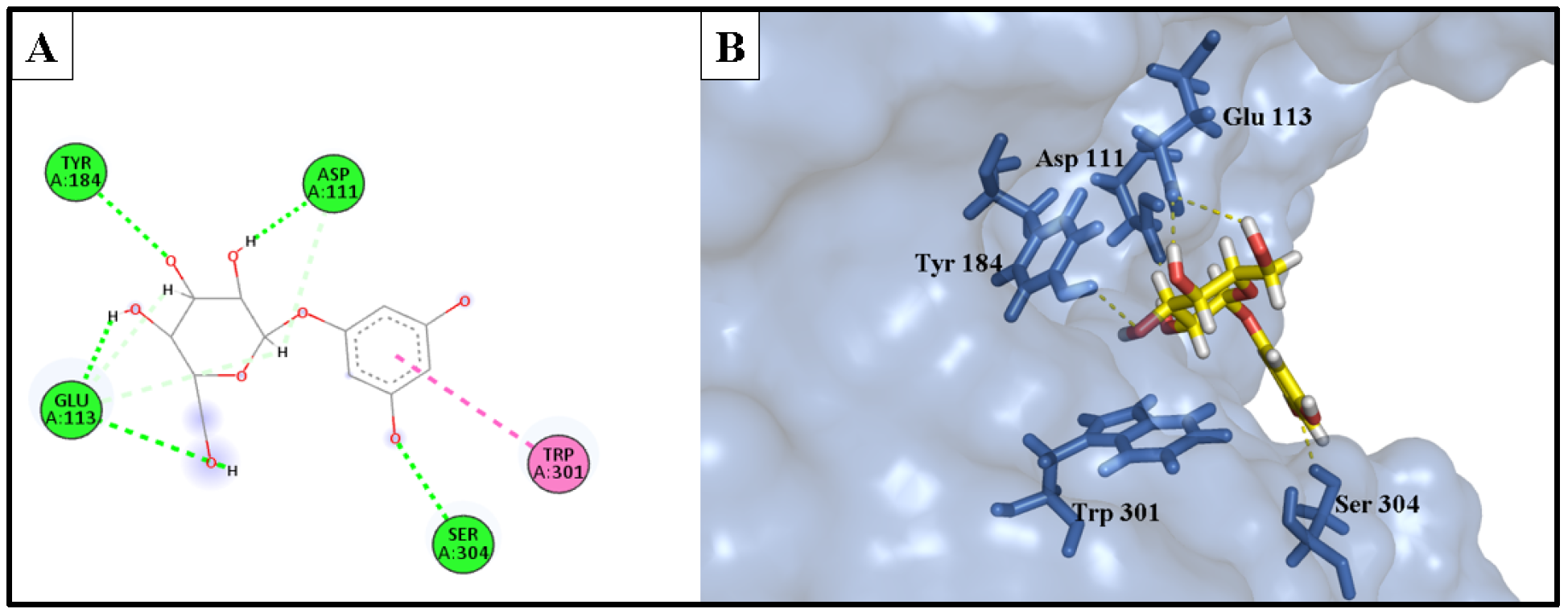
2.5.2. Anti-Hyaluronidase Activity
3. Materials and Methods
3.1. Plant Material and Extraction
3.2. General Experimental Materials and Procedures
3.3. Evaluation of the Total Phenolic Content (TPC) of Ethanolic Extract
3.4. Evaluation of the Total Flavonoid Content (TFC) of Ethanolic Extract
3.5. Chromatographic Isolation of Phytoconstituents
3.6. Antioxidant Activities of the Isolated Compounds
3.6.1. ABTS Radical Scavenging
3.6.2. Ferric Reducing/Antioxidant Power (FRAP)
3.7. Anti-Aging Activities of the Isolated Compounds
3.7.1. Tyrosinase Inhibitory Assay
3.7.2. Hyaluronidase Inhibitory Assay
3.8. In Silico Molecular Docking Studies
3.8.1. Anti-Tyrosinase Activity
3.8.2. Anti-Hyaluronidase Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Abd El-Raouf, H.S. Taxonomic significance of leaves in family Aizoaceae. Saudi J. Biol. Sci. 2021, 28, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Bakr, R.O. A Comprehensive Review of the Aizoaceae Family: Phytochemical and Biological Studies. Nat. Prod. J. 2021, 11, 288–304. [Google Scholar] [CrossRef]
- Glavich, T. Beginner’s Guide to Faucaria. Cactus Succul. J. 2016, 88, 290–292. [Google Scholar] [CrossRef]
- Schwantes, G.; Higgins, V. Flowering Stones and Mid-Day Flowers: A Book for Plant and Nature Lovers on the Mesembryanthemaceae; E. Benn: London, UK, 1957. [Google Scholar]
- Graf, A.B. Pictorial Cyclopedia of Exotic Plants from Tropical and Near-Tropic Regions: Guide to Care of Plants Indoors, Horticultural Color Guide, Plant Geography; Roehr: Cincinnati, OH, USA, 1976. [Google Scholar]
- Zhu, Y.; Yu, X.; Ge, Q.; Li, J.; Wang, D.; Wei, Y.; Ouyang, Z. Antioxidant and anti-aging activities of polysaccharides from Cordyceps cicadae. Int. J. Biol. Macromol. 2020, 157, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Widowati, W.; Fauziah, N.; Herdiman, H.; Afni, M.; Afifah, E.; Kusuma, H.S.W.; Nufus, H.; Arumwardana, S.; Rihibiha, D.D. Antioxidant and anti aging assays of Oryza sativa extracts, vanillin and coumaric acid. J. Nat. Remedies 2016, 16, 88–99. [Google Scholar] [CrossRef]
- Younis, M.M.; Ayoub, I.M.; Mostafa, N.M.; El Hassab, M.A.; Eldehna, W.M.; Al-Rashood, S.T.; Eldahshan, O.A. GC/MS profiling, anti-collagenase, anti-elastase, anti-tyrosinase and anti-hyaluronidase activities of a Stenocarpus sinuatus leaves extract. Plants 2022, 11, 918. [Google Scholar] [CrossRef]
- Masaki, H. Role of antioxidants in the skin: Anti-aging effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Biswas, R.; Sharma, A.; Banerjee, S.; Biswas, S.; Katiyar, C. Validation of medicinal herbs for anti-tyrosinase potential. J. Herb. Med. 2018, 14, 1–16. [Google Scholar] [CrossRef]
- Fayad, S.; Morin, P.; Nehmé, R. Use of chromatographic and electrophoretic tools for assaying elastase, collagenase, hyaluronidase, and tyrosinase activity. J. Chromatogr. A 2017, 1529, 1–28. [Google Scholar] [CrossRef]
- Youssif, K.; Elshamy, A.; Rabeh, M.; Gabr, N.; Haggag, E. A Phytochemical and Biological Review on Plants of The family Aizoaceae. J. Adv. Pharm. Res. 2019, 3, 158–181. [Google Scholar] [CrossRef]
- Bazzicalupo, M.; Cornara, L.; Burlando, B.; Cascini, A.; Denaro, M.; Smeriglio, A.; Trombetta, D. Carpobrotus edulis (L.) NE Br. extract as a skin preserving agent: From traditional medicine to scientific validation. J. Integr. Med. 2021, 19, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Kalicharan, B.; Naidoo, Y.; van Staden, J. Ethnopharmacology and biological activities of the Aizoaceae. J. Ethnopharmacol. 2022, 303, 115988. [Google Scholar] [CrossRef] [PubMed]
- Äyräs, P.; Widén, C.J. NMR spectroscopy of naturally occurring phloroglucinol derivatives. Planta Medica 1978, 34, 144–152. [Google Scholar] [CrossRef]
- Foo, L.Y.; Karchesy, J.J. Polyphenolic glycosides from Douglas fir inner bark. Phytochemistry 1989, 28, 1237–1240. [Google Scholar]
- ERSÖZ, T.; HARPUT, Ü.Ş.; SARACOĞLU, İ.; ÇALIŞ, İ.; OGIHARA, Y. Phenolic compounds from Scutellaria pontica. Turk. J. Chem. 2002, 26, 581–588. [Google Scholar]
- Andersen, O.M.; Markham, K.R. Flavonoids: Chemistry, Biochemistry and Applications; CRC press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Beck, M.-A.; Häberlein, H. Flavonol glycosides from Eschscholtzia californica. Phytochemistry 1999, 50, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Afifi, N.I.; Moawad, A.S.; Hetta, M.H.; Mohammed, R.M. Phytochemical composition and antioxidant activity of two species related to family Arecaceae. Pharm. Sci. Asia 2022, 49, 43–50. [Google Scholar] [CrossRef]
- Lou, H.; Yamazaki, Y.; Sasaki, T.; Uchida, M.; Tanaka, H.; Oka, S. A-type proanthocyanidins from peanut skins. Phytochemistry 1999, 51, 297–308. [Google Scholar] [CrossRef]
- Alejo-Armijo, A.; Salido, S.a.; Altarejos, J.n. Synthesis of A-type proanthocyanidins and their analogues: A comprehensive review. J. Agric. Food Chem. 2020, 68, 8104–8118. [Google Scholar] [CrossRef]
- de Moura Martins, C.; de Morais, S.A.; Martins, M.M.; Cunha, L.C.; da Silva, C.V.; Teixeira, T.L.; Santiago, M.B.; de Aquino, F.J.; Nascimento, E.A.; Chang, R. Antifungal and cytotoxicity activities and new proanthocyanidins isolated from the barks of Inga laurina (Sw.) Willd. Phytochem. Lett. 2020, 40, 109–120. [Google Scholar] [CrossRef]
- Afifi, N.I.; Moawad, A.S.; Zaki, M.A.; Rateb, M.E.; Rashed, M.H.; Saleh, I.G.; Hetta, M.H.; Mohammed, R.M. Four new phenolics and antiparasitic secondary metabolites from Flacourtia rukam Zoll. & Mortizi. Nat. Prod. Res. 2022, 36, 3626–3637. [Google Scholar] [PubMed]
- Cronjé, A.; Burger, J.F.; Brandt, E.V.; Kolodziej, H.; Ferreira, D. Assessment of 3, 4trans and 3, 4-cis relative configurations in the a-series of (4, 8)-linked proanthocyanidins. Tetrahedron Lett. 1990, 31, 3789–3792. [Google Scholar] [CrossRef]
- Liochev, S.I. Reactive oxygen species and the free radical theory of aging. Free. Radic. Biol. Med. 2013, 60, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Hussain, F.; Kayani, H.U.R. Aging-Oxidative stress, antioxidants and computational modeling. Heliyon 2020, 6, e04107. [Google Scholar]
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S. Oxidative stress in human pathology and aging: Molecular mechanisms and perspectives. Cells 2022, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free. Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Zhou, H.-C.; Lin, Y.-M.; Li, Y.-Y.; Li, M.; Wei, S.-D.; Chai, W.-M.; Tam, N.F.-y. Antioxidant properties of polymeric proanthocyanidins from fruit stones and pericarps of Litchi chinensis Sonn. Food Res. Int. 2011, 44, 613–620. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, J.H. Antioxidant and anti-inflammatory activity of phloroglucinol from seaweeds. FASEB J. 2016, 30, 1174.12. [Google Scholar]
- Dede, E.; Genc, N.; Elmastas, M.; Aksit, H.; Erenler, R. Chemical constituents isolated from Rhododendron ungernii with antioxidant profile. Nat. Prod. J. 2019, 9, 238–243. [Google Scholar] [CrossRef]
- Xiao, Z.; He, L.; Hou, X.; Wei, J.; Ma, X.; Gao, Z.; Yuan, Y.; Xiao, J.; Li, P.; Yue, T. Relationships between structure and antioxidant capacity and activity of glycosylated flavonols. Foods 2021, 10, 849. [Google Scholar] [CrossRef]
- Mahmoud, A.A.; Al-Shihry, S.S.; Hegazy, M.-E.F. Biological activity of a phloroglucinol glucoside derivative from Conyza aegyptiaca. Z. Für Naturforschung C 2009, 64, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.-P.; Wu, H.; Wang, Y.; Weng, X.-C. Isolation of some compounds from nutmeg and their antioxidant activities. Czech J. Food Sci. 2012, 30, 164–170. [Google Scholar] [CrossRef]
- Ghalloo, B.A.; Khan, K.-u.-R.; Ahmad, S.; Aati, H.Y.; Al-Qahtani, J.H.; Ali, B.; Mukhtar, I.; Hussain, M.; Shahzad, M.N.; Ahmed, I. Phytochemical profiling, in vitro biological activities, and in silico molecular docking studies of Dracaena reflexa. Molecules 2022, 27, 913. [Google Scholar] [CrossRef] [PubMed]
- Waksmundzka-Hajnos, M.; Sherma, J.; Kowalska, T. Thin Layer Chromatography in Phytochemistry; CRC Press: Boca Raton, FL, USA, 2008. [Google Scholar]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Antioxidant and phytochemical activities of Amaranthus caudatus L. harvested from different soils at various growth stages. Sci. Rep. 2019, 9, 12965. [Google Scholar] [CrossRef] [PubMed]
- Bakar, M.F.A.; Mohamed, M.; Rahmat, A.; Fry, J. Phytochemicals and antioxidant activity of different parts of bambangan (Mangifera pajang) and tarap (Artocarpus odoratissimus). Food Chem. 2009, 113, 479–483. [Google Scholar] [CrossRef]
- Liangli, L.Y. Wheat Antioxidants; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Sánchez, C.S.; González, A.T.; García-Parrilla, M.; Granados, J.Q.; De La Serrana, H.L.G.; Martínez, M.L. Different radical scavenging tests in virgin olive oil and their relation to the total phenol content. Anal. Chim. Acta 2007, 593, 103–107. [Google Scholar] [CrossRef]
- Zafar, R.; Ullah, H.; Zahoor, M.; Sadiq, A. Isolation of bioactive compounds from Bergenia ciliata (haw.) Sternb rhizome and their antioxidant and anticholinesterase activities. BMC Complement. Altern. Med. 2019, 19, 296. [Google Scholar] [CrossRef]
- Ferreira, I.C.; Baptista, P.; Vilas-Boas, M.; Barros, L. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal: Individual cap and stipe activity. Food Chem. 2007, 100, 1511–1516. [Google Scholar] [CrossRef]
- El-Nashar, H.A.; El-Labbad, E.M.; Al-Azzawi, M.A.; Ashmawy, N.S. A new xanthone glycoside from Mangifera indica L.: Physicochemical properties and in vitro anti-skin aging activities. Molecules 2022, 27, 2609. [Google Scholar] [CrossRef]
- Ibrahim, N.; Abbas, H.; El-Sayed, N.S.; Gad, H.A. Rosmarinus officinalis L. hexane extract: Phytochemical analysis, nanoencapsulation, and in silico, in vitro, and in vivo anti-photoaging potential evaluation. Sci. Rep. 2022, 12, 13102. [Google Scholar] [CrossRef]
- Roselan, M.A.; Zakaria, N.; Faujan, N.H.; Latif, M.A.M.; Faudzi, S.M.M.; Ab Hadi, H.; Ashari, S.E. In vitro cytotoxicity assay, mushroom tyrosinase inhibitory activity and release analysis of kojic monooleate nanodelivery system and in silico molecular docking study against 2Y9X target enzyme. J. Drug Deliv. Sci. Technol. 2021, 66, 102764. [Google Scholar] [CrossRef]
- Sepehri, N.; Iraji, A.; Yavari, A.; Asgari, M.S.; Zamani, S.; Hosseini, S.; Bahadorikhalili, S.; Pirhadi, S.; Larijani, B.; Khoshneviszadeh, M. The natural-based optimization of kojic acid conjugated to different thio-quinazolinones as potential anti-melanogenesis agents with tyrosinase inhibitory activity. Bioorganic Med. Chem. 2021, 36, 116044. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.A.; AboulMagd, A.M.; Abdel-Rahman, H.M. Quinazoline-Schiff base conjugates: In Silico study and ADMET predictions as multi-target inhibitors of coronavirus (SARS-CoV-2) proteins. RSC Adv. 2020, 10, 34033–34045. [Google Scholar] [CrossRef] [PubMed]
- El-Hawary, S.S.; Mohammed, R.; Taher, M.A.; AbouZid, S.F.; Mansour, M.A.; Almahmoud, S.A.; Huwaimel, B.; Amin, E. Characterization of Promising Cytotoxic Metabolites from Tabebuia guayacan Hemsl.: Computational Prediction and In Vitro Testing. Plants 2022, 11, 888. [Google Scholar] [CrossRef]
- El-Hawary, S.S.; Mohammed, R.; Lithy, N.M.; AbouZid, S.F.; Mansour, M.A.; Almahmoud, S.A.; Huwaimel, B.; Amin, E. Digalloyl glycoside: A potential inhibitor of trypanosomal PFK from Euphorbia abyssinica JF Gmel. Plants 2022, 11, 173. [Google Scholar] [CrossRef]
- Abdel-Azziz, I.A.; Amin, N.H.; El-Saadi, M.T.; Abdel-Rahman, H.M. Design, synthesis and mechanistic studies of benzophenones hydrazone derivatives as cathepsin inhibitors. J. Mol. Struct. 2023, 1274, 134583. [Google Scholar] [CrossRef]
- Shaker, A.M.; Shahin, M.I.; AboulMagd, A.M.; Aleem, S.A.A.; Abdel-Rahman, H.M.; Abou El Ella, D.A. Novel 1, 3-diaryl pyrazole derivatives bearing methylsulfonyl moiety: Design, synthesis, molecular docking and dynamics, with dual activities as anti-inflammatory and anticancer agents through selectively targeting COX-2. Bioorganic Chem. 2022, 129, 106143. [Google Scholar] [CrossRef]
- Altyar, A.E.; Ashour, M.L.; Youssef, F.S. Premna odorata: Seasonal metabolic variation in the essential oil composition of its leaf and verification of its anti-ageing potential via in vitro assays and molecular modelling. Biomolecules 2020, 10, 879. [Google Scholar] [CrossRef]
- Juszczak, A.M.; Marijan, M.; Jakupović, L.; Tomczykowa, M.; Tomczyk, M.; Zovko Končić, M. Glycerol and Natural Deep Eutectic Solvents Extraction for Preparation of Luteolin-Rich Jasione montana Extracts with Cosmeceutical Activity. Metabolites 2023, 13, 32. [Google Scholar] [CrossRef]
- Gębalski, J.; Graczyk, F.; Załuski, D. Paving the way towards effective plant-based inhibitors of hyaluronidase and tyrosinase: A critical review on a structure–activity relationship. J. Enzym. Inhib. Med. Chem. 2022, 37, 1120–1195. [Google Scholar] [CrossRef]
- Meyer-Almes, F.-J. Repurposing approved drugs as potential inhibitors of 3CL-protease of SARS-CoV-2: Virtual screening and structure based drug design. Comput. Biol. Chem. 2020, 88, 107351. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δH (mult, J) | δC | HMBC (H→C) |
|---|---|---|---|
| Flavanol moiety | |||
| 2 | ---- | 100.3 | ---- |
| 3 | 4.05 (d, 3.6) | 68.08 | C-4a |
| 4 | 4.64 (d, 4) | 29.5 | C-4a, C-2, C-2′, C-3′ |
| 4a | ---- | 104.5 | ---- |
| 5 | ---- | 155.5 | ---- |
| 6 | 6.03(d, 2.4) | 98.6 | C-4a |
| 7 | ---- | 156.2 | ---- |
| 8 | 6.10 (d, 2.4) | 96.9 | ---- |
| 8a | ---- | 158.6 | ----- |
| 9 | ---- | 132.1 | ----- |
| 10 | 7.13 (d, 2.4) | 115.65 | C-2, C-9, C-11, C-12, C-14 |
| 11 | ---- | 146.7 | ---- |
| 12 | ---- | 145.7 | ---- |
| 13 | 6.81 (d, 8.4) | 115.67 | C-2, C-11, C-12, C-14 |
| 14 | 7.01 (dd, 2, 8.4) | 119.8 | C-2, C-9, C-10, C-12, C-13 |
| Phloroglucinol moiety | |||
| 2′ | ---- | 108.1 | |
| 3′ | ---- | 154.5 | |
| 4′ | 6.08 (d, 2) | 98.1 | C-2′, C-5′ |
| 5′ | ---- | 154.9 | |
| 6′ | 6.23 (d, 2.4) | 96.8 | C-1′, C-2′ |
| 1′ | ---- | 158.1 | ---- |
| β-glucopyranoside moiety | |||
| 1″ | 4.95 (d, 7.6) | 101.9 | C-5′ |
| 2″ | 3.604 (dd, 1.6, 7.2) | 74.6 | |
| 3″ | 3.53 (m) | 77.3 | |
| 4″ | 3.46 (d, 4.8) | 71.2 | |
| 5″ | 3.46 (d, 4.8) | 78.2 | |
| 6″ | 3.73 (m, H-6′ a) | 62.3 | |
| 3.92 (m, H-6′ b) | |||
| Compound No. | Compound Name | IC50 Value (µg/mL) | |
|---|---|---|---|
| ABTS Assay | FRAP Assay | ||
| 1 | Phloroglucinol | 6.44 ± 0.47 | 12.89 ± 0.93 |
| 2 | Phlorin | 43.40 ± 3.18 | 64.52 ± 4.15 |
| 3 | Isorhamnetin 3-O-rutinoside | 12.24 ± 0.61 | 25.24 ± 2.03 |
| 4 | (-) Catechin-(2→1′,4→2′)-phloroglucinol | 4.11 ± 0.32 | 7.36 ± 0.57 |
| 5 | Isorhamnetin 3-O-[α-rhamnopyranosyl-(1→4)-α-rhamnopyranosyl-(1→6)-β-glucopyranoside] | 18.19 ± 2.94 | 35.04 ± 2.89 |
| 6 | β-sitosterol | 557.46 ± 19.76 | ND |
| Positive standard (Ascorbic acid) | 10.67 ± 0.85 | 20.86± 1.28 | |
| Tyrosinase (PDB ID: 2Y9X) | ||||
|---|---|---|---|---|
| Compound | Affinity (kcal/mol) | Distance (in Å) from Main Residue | Type of Interaction | |
| Phlorin | −10.935 | 2.61 | Cu400 | Metal acceptor |
| 2.55 | Cu401 | Metal acceptor | ||
| 2.50 | His85 | H-bond acceptor | ||
| 2.82 | Asn260 | H-bond donor | ||
| 2.74 | Met280 | H-bond donor | ||
| 4.93 | His244 | Hydrophobic | ||
| Isorhamnetin 3-O-rutinoside | −12.154 | 2.12 | Cu400 | Metal acceptor |
| 2.26 | Cu401 | Metal acceptor | ||
| 2.36 | His85 | H-bond acceptor | ||
| 2.97 | Met280 | H-bond donor | ||
| 2.27 | Val283 | H-bond acceptor | ||
| 2.53 | Asn81 | H-bond acceptor | ||
| 3.14 | His263 | Hydrophobic | ||
| 3.45 | Gly281 | Hydrophobic | ||
| kojic acid | −9.385 | 2.96 | Cu400 | Metal acceptor |
| 2.63 | Cu401 | Metal acceptor | ||
| 2.34 | His85 | H-bond acceptor | ||
| 2.90 | Asn260 | H-bond acceptor | ||
| 2.78 | Met280 | H-bond donor | ||
| 3.89 | His263 | Hydrophobic | ||
| 3.39 | Val283 | H-bond acceptor | ||
| Hyaluronidase (PDB ID: 1FCV) | ||||
| Compound | Affinity (kcal/mol) | Distance (in Å) from Main Residue | Type of Interaction | |
| Phlorin | −14.979 | 1.91 | Asp111 | H-bond donor |
| 2.34 | Ser304 | H-bond acceptor | ||
| 1.93 | Glu113 | H-bond donor | ||
| 2.98 | Glu113 | H-bond donor | ||
| 4.11 | Trp301 | Hydrophobic | ||
| 2.51 | Tyr184 | H-bond acceptor | ||
| Isorhamnetin 3-O-rutinoside | −16.578 | 2.15 | Asp111 | H-bond donor |
| 2.89 | Ser304 | H-bond acceptor | ||
| 2.36 | Ser303 | H-bond acceptor | ||
| 2.41 | Ser303 | H-bond acceptor | ||
| 1.99 | Glu113 | Electrostatic | ||
| 3.14 | Trp301 | Hydrophobic | ||
| 3.54 | Tyr55 | Hydrophobic | ||
| 3.22 | Asp305 | Hydrophobic | ||
| Luteolin | −12.404 | 2.66 | Asp111 | H-bond donor |
| 2.00 | Ser304 | H-bond acceptor | ||
| 3.57 | Glu113 | Hydrophobic | ||
| 4.74 | Trp301 | Hydrophobic | ||
| 4.77 | Tyr55 | Hydrophobic | ||
| 2.45 | Asp56 | H-bond acceptor | ||
| 1.95 | Asp305 | H-bond donor | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, H.S.; Abouzeid, H.; Mansour, M.A.; Owis, A.I.; Amin, E.; Darwish, H.W.; Alanazi, A.S.; Naguib, I.A.; Afifi, N. Antioxidant and Anti-Aging Phytoconstituents from Faucaria tuberculosa: In Vitro and In Silico Studies. Molecules 2023, 28, 6895. https://doi.org/10.3390/molecules28196895
Ahmed HS, Abouzeid H, Mansour MA, Owis AI, Amin E, Darwish HW, Alanazi AS, Naguib IA, Afifi N. Antioxidant and Anti-Aging Phytoconstituents from Faucaria tuberculosa: In Vitro and In Silico Studies. Molecules. 2023; 28(19):6895. https://doi.org/10.3390/molecules28196895
Chicago/Turabian StyleAhmed, Hayam S., Hala Abouzeid, Mostafa A. Mansour, Asmaa I. Owis, Elham Amin, Hany W. Darwish, Ashwag S. Alanazi, Ibrahim A. Naguib, and Naglaa Afifi. 2023. "Antioxidant and Anti-Aging Phytoconstituents from Faucaria tuberculosa: In Vitro and In Silico Studies" Molecules 28, no. 19: 6895. https://doi.org/10.3390/molecules28196895
APA StyleAhmed, H. S., Abouzeid, H., Mansour, M. A., Owis, A. I., Amin, E., Darwish, H. W., Alanazi, A. S., Naguib, I. A., & Afifi, N. (2023). Antioxidant and Anti-Aging Phytoconstituents from Faucaria tuberculosa: In Vitro and In Silico Studies. Molecules, 28(19), 6895. https://doi.org/10.3390/molecules28196895