
Unlocking the Therapeutic Potential of Stevia rebaudiana Bertoni: A Natural Antiglycating Agent and Non-Toxic Support for HDF Cell Health


 , , ,
, , ,
Abstract
:1. Introduction
2. Results
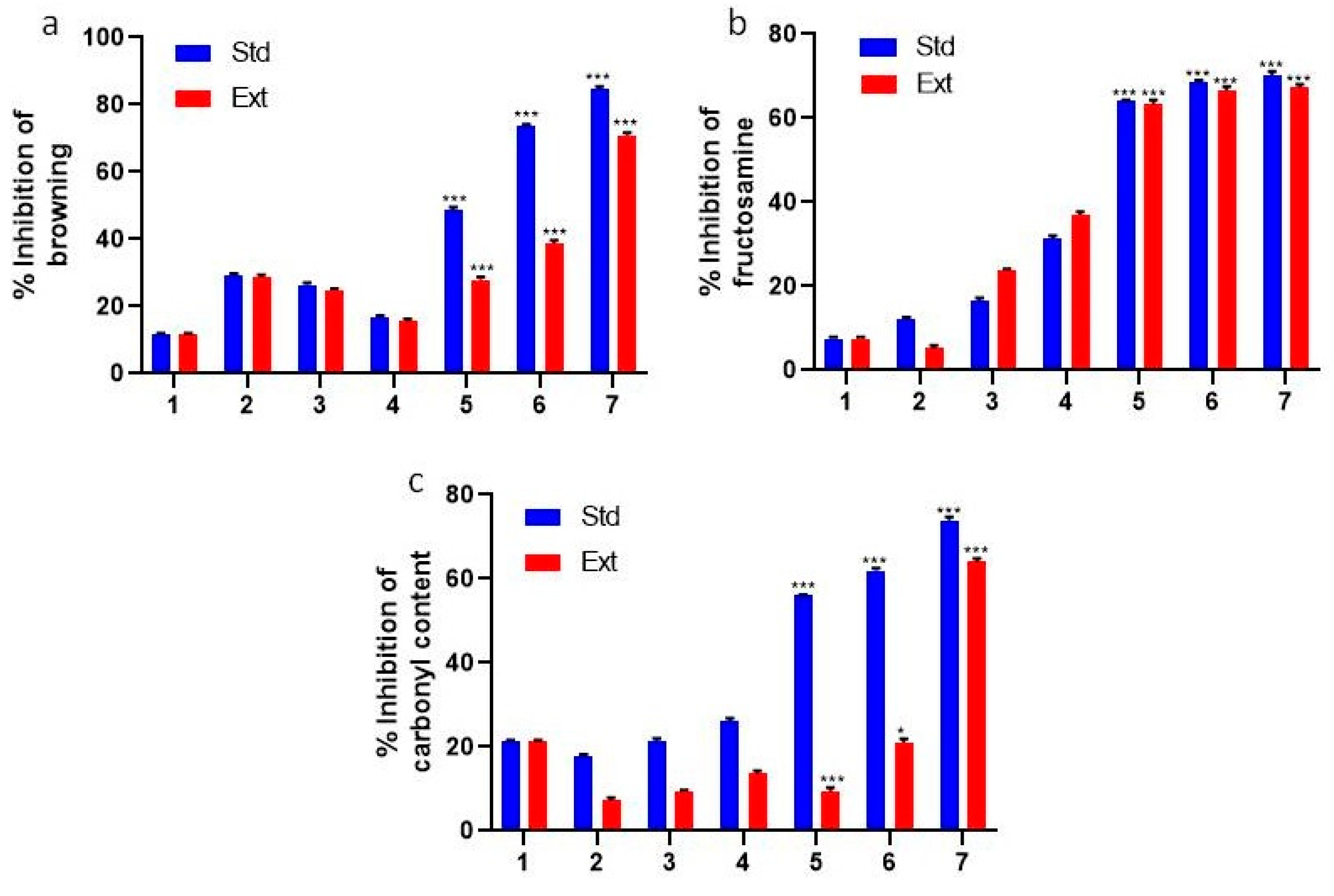
2.1. Measurement of Browning
2.2. Determination of Fructosamine
2.3. Determining the Carbonyl Content of Proteins
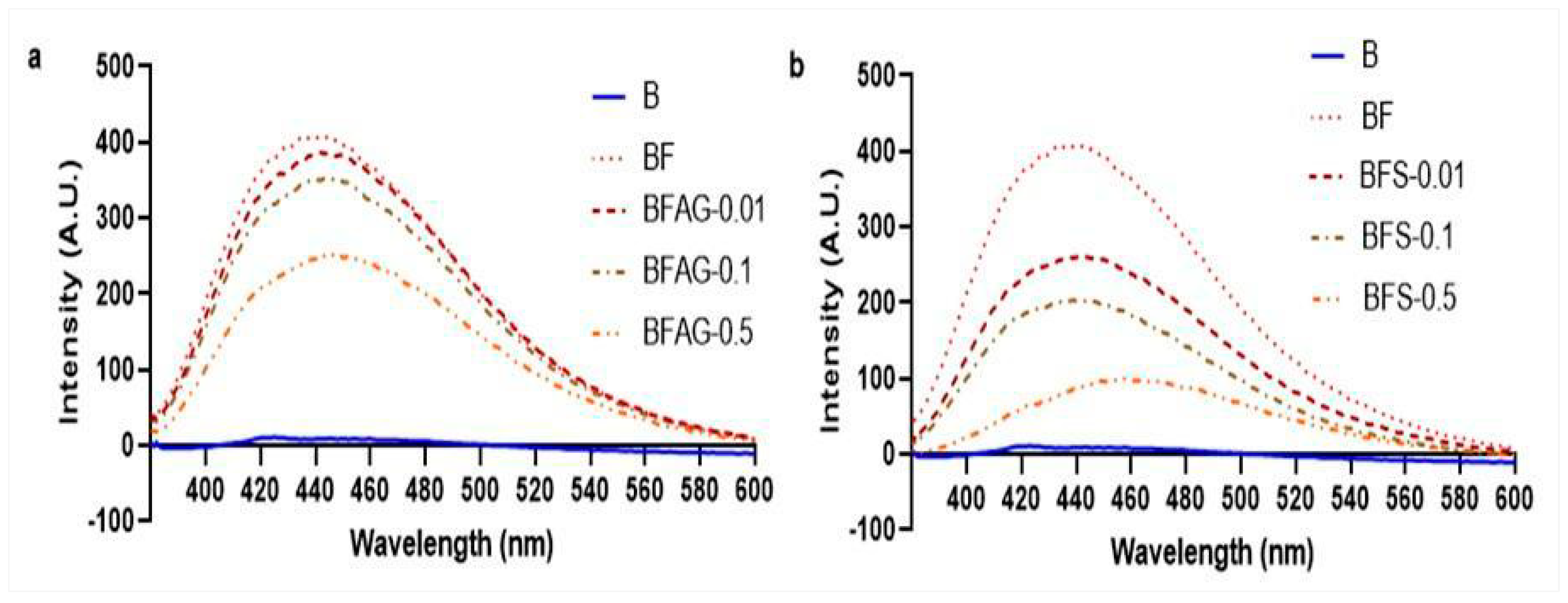
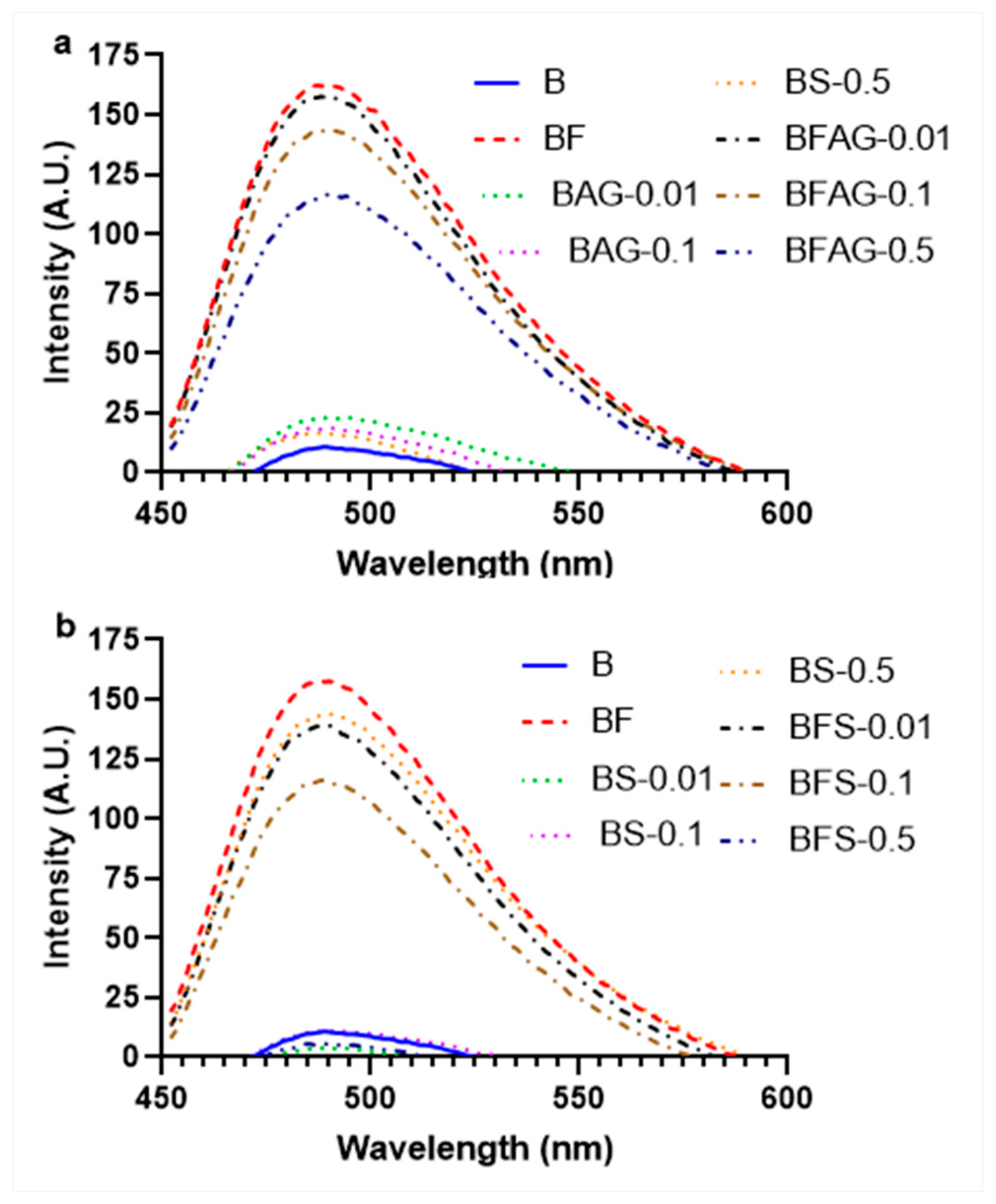
2.4. Measurement of Total AGEs and Individual AGEs
2.5. FTIR Investigation of Amide Bands
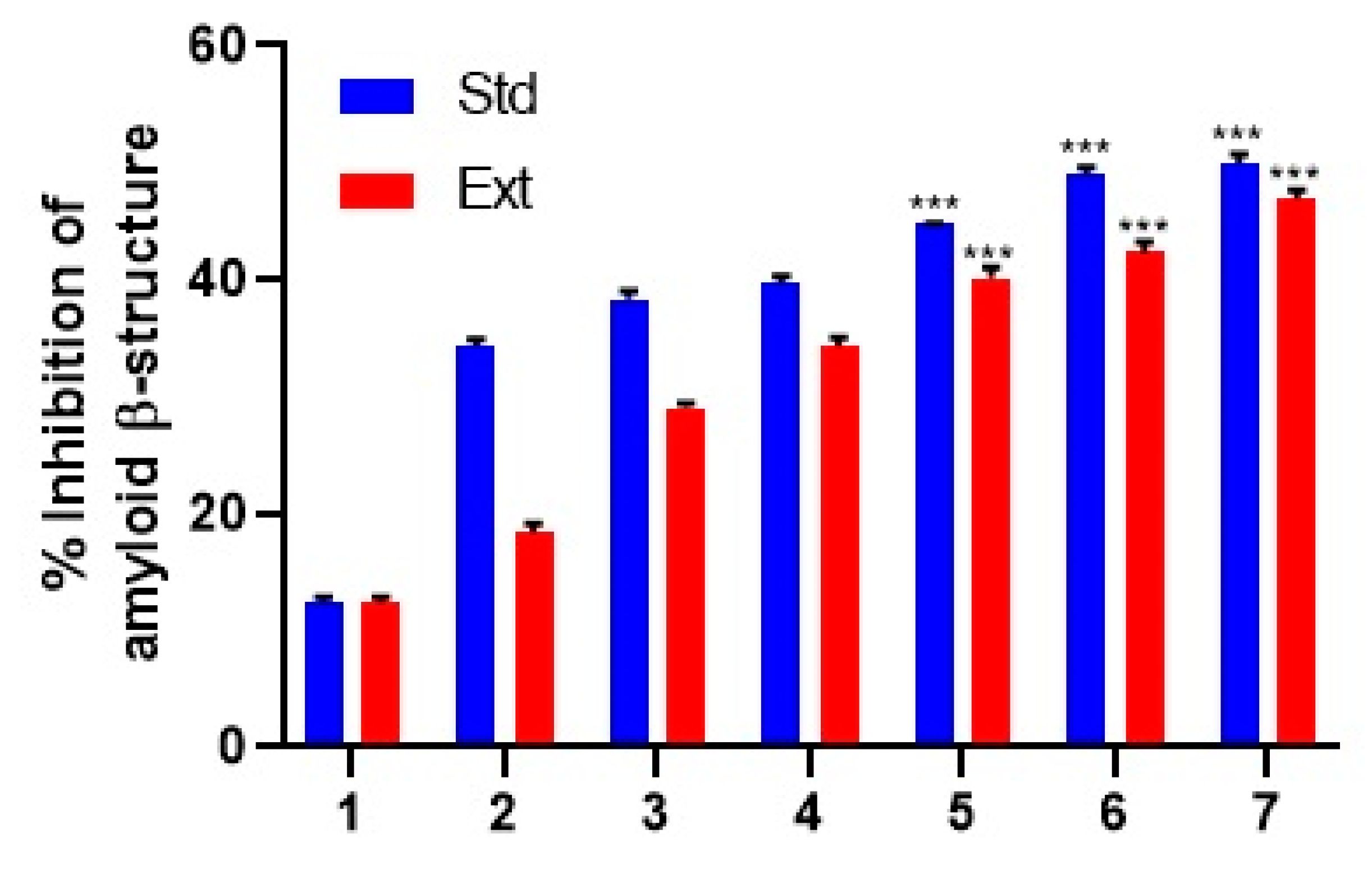
2.6. Determination of Amyloid β-Structure
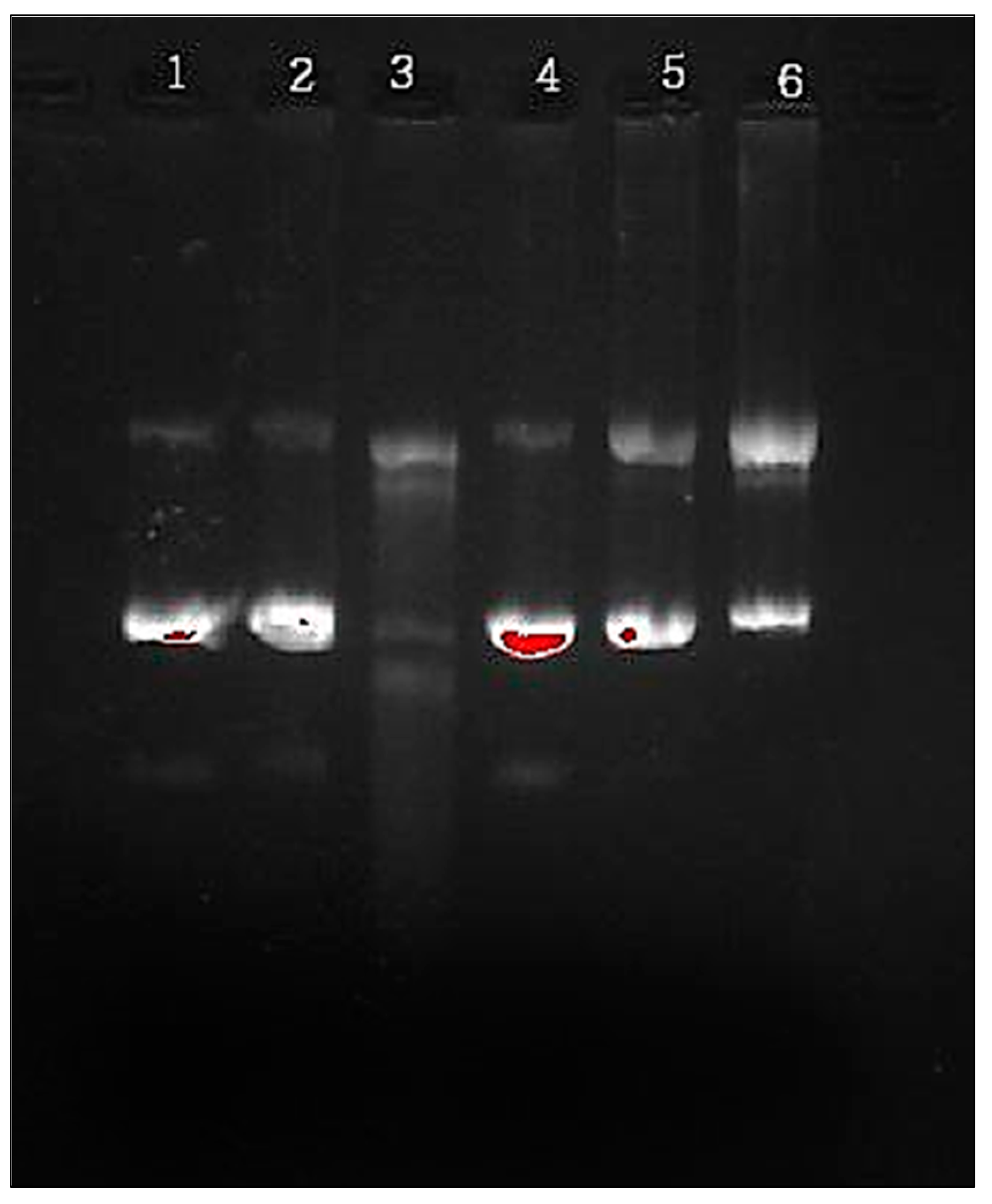
2.7. In Vitro DNA Damage Analysis
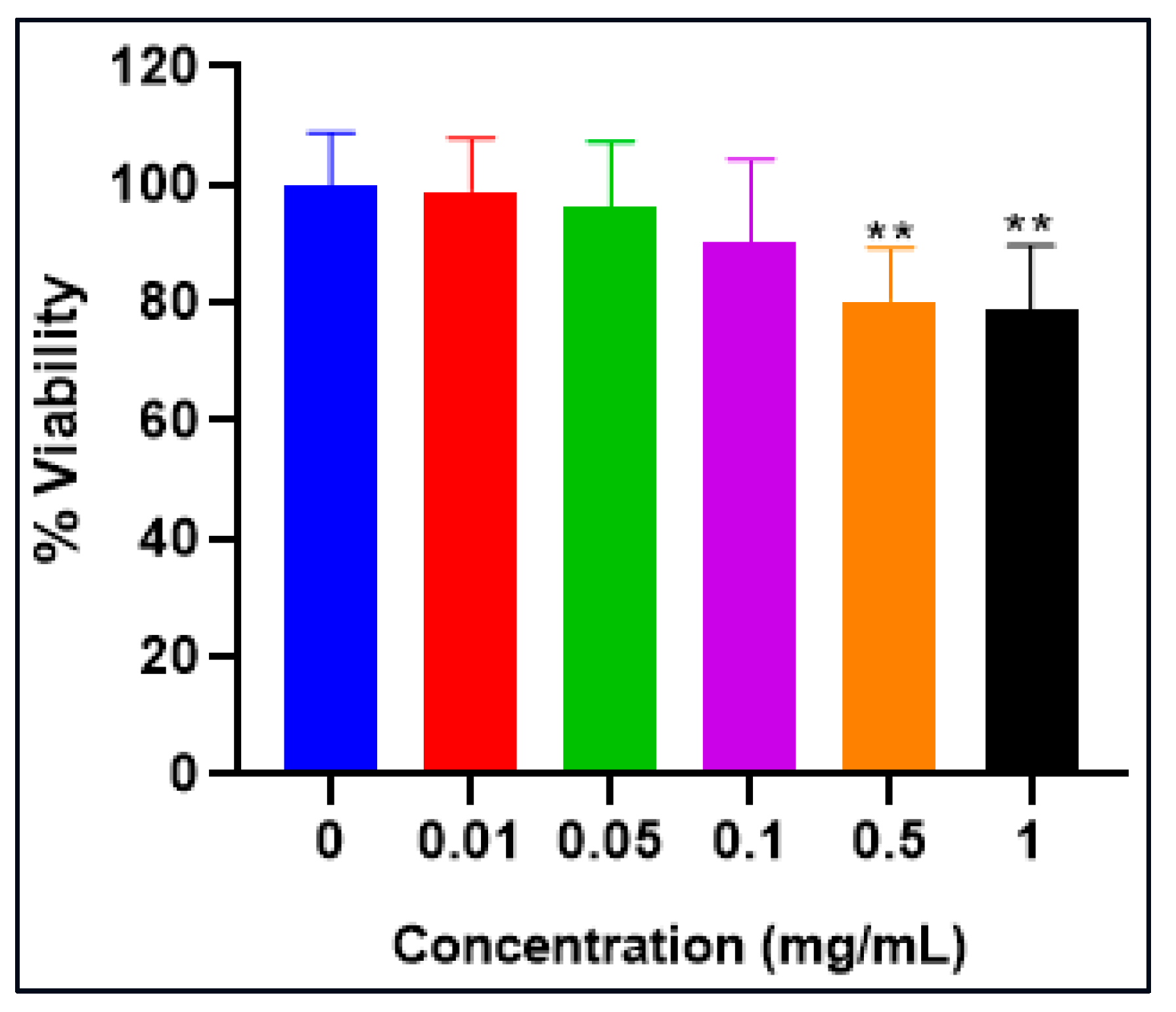
2.8. 3-[4,5-Dimethylthiazol-2-yl]-2, 5 Diphenyltetrazolium Bromide Assay (MTT ASSAY)
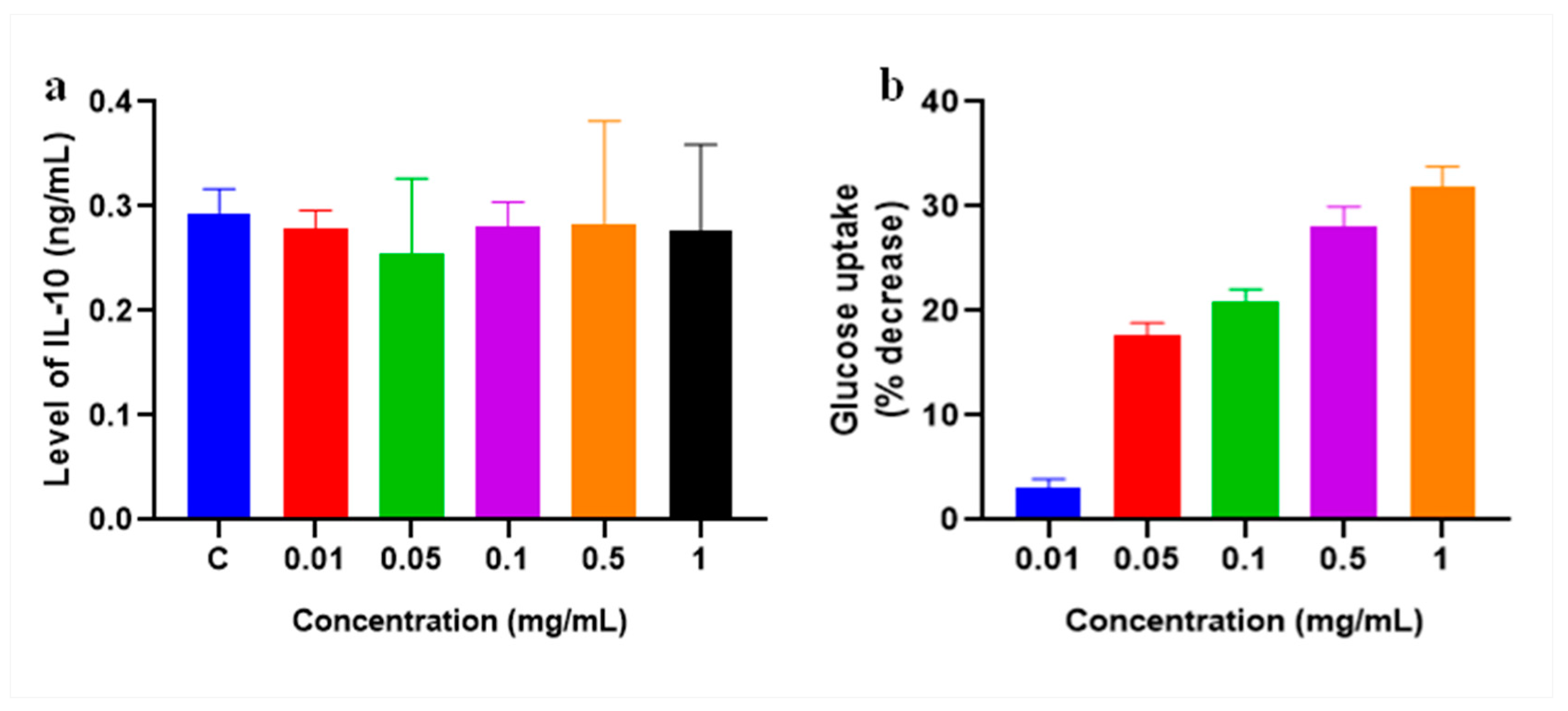
2.9. Production of Cytokine
2.10. Glucose Uptake Assay
3. Discussion
4. Materials and Methods
4.1. Material
4.1.1. Plant Material and Identification
4.1.2. Extraction of Stevia Plant Leaves
4.2. Antiglycation Assays
4.2.1. Albumin Glycation
4.2.2. Determining the Browning Level
4.2.3. Measurement of Fructosamine Content
4.2.4. Determination of Carbonyl Content
4.3. Measurement of Total AGEs and Individual AGEs
4.4. FTIR Study on AGE-Specific Amide Bands
4.5. Determination of Amyloid β-Structure
4.6. In Vitro Glycation of Plasmid DNA
4.7. MTT Assay
4.8. Phase Contrast Microscopy
4.9. Cytokine Production
4.10. Glucose Uptake Assay
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| AG | Aminoguanidine |
| AGEs | Advanced Glycation End Products |
| BSA | Bovine serum albumin |
| DNPH | 2,4-Dinitrophenylhydrazine |
| H2O2 | Hydrogen Peroxide |
| HDF | Human Dermal Fibroblast |
| MG | Methylglyoxal |
| NaN3 | Sodium azide |
| ROS | Reactive Oxygen Species |
| PEPCK | Phosphoenolpyruvate carboxykinase |
| ThT | Thioflavin T |
| mM | Milimolar |
| µL | Microliter |
| μM | Micromolar |
References
- Kikuchi, S.; Shinpo, K.; Takeuchi, M.; Yamagishi, S.; Makita, Z.; Sasaki, N.; Tashiro, K. Glycation—A sweet tempter for neuronal death. Brain Res. Rev. 2003, 41, 306–323. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N. Advanced glycation endproducts—Role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K. Studies on free radicals, antioxidants, and co-factors. Clin. Interv. Aging 2007, 2, 219. [Google Scholar] [PubMed]
- Sadowska-Bartosz, I.; Bartosz, G. Prevention of protein glycation by natural compounds. Molecules 2015, 20, 3309–3334. [Google Scholar] [CrossRef]
- Ahmad, S.; Khan, M.S.; Akhter, F.; Khan, M.S.; Khan, A.; Ashraf, J.M.; Pandey, R.P.; Shahab, U. Glycoxidation of biological macromolecules: A critical approach to halt the menace of glycation. Glycobiology 2014, 24, 979–990. [Google Scholar] [CrossRef]
- Grzegorczyk-Karolak, I.; Gołąb, K.; Gburek, J.; Wysokińska, H.; Matkowski, A. Inhibition of advanced glycation end-product formation and antioxidant activity by extracts and polyphenols from Scutellaria alpina L. and S. altissima L. Molecules 2016, 21, 739. [Google Scholar] [CrossRef]
- Banan, P.; Ali, A. Preventive effect of phenolic acids on in vitro glycation. Ann. Phytomedicine 2016, 5, 97–102. [Google Scholar] [CrossRef]
- Balyan, P.; Ola, M.S.; Alhomida, A.S.; Ali, A. D-Ribose-Induced Glycation and Its Attenuation by the Aqueous Extract of Nigella sativa Seeds. Medicina 2022, 58, 1816. [Google Scholar] [CrossRef]
- Poulsen, M.W.; Hedegaard, R.V.; Andersen, J.M.; de Courten, B.; Bügel, S.; Nielsen, J.; Skibsted, L.H.; Dragsted, L.O. Advanced glycation endproducts in food and their effects on health. Food Chem. Toxicol. 2013, 60, 10–37. [Google Scholar] [CrossRef]
- Thornalley, P.J. Use of aminoguanidine (Pimagedine) to prevent the formation of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 419, 31–40. [Google Scholar] [CrossRef]
- Nathan, D.M.; Buse, J.B.; Davidson, M.B.; Ferrannini, E.; Holman, R.R.; Sherwin, R.; Zinman, B. Medical management of hyperglycemia in type 2 diabetes: A consensus algorithm for the initiation and adjustment of therapy. Diabetes Care 2009, 32, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Hardy, O.T.; Czech, M.P.; Corvera, S. What causes the insulin resistance underlying obesity? Curr. Opin. Endocrinol. Diabetes Obes. 2012, 19, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, M.; le Roux, C.W.; Docherty, N.G. Morbidity and mortality associated with obesity. Ann. Transl. Med. 2017, 5, 161. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; Gothalwal, R. Herbal origins provision for non-enzymatic Glycation (NEGs) inhibition. Front. Med. Chem. Drug Discov. 2018, 2, 10-015. [Google Scholar]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and Other Phenolic Compounds from Medicinal Plants for Pharmaceutical and Medical Aspects: An Overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef]
- Martins, G.R.; Bronzel Junior, J.L.; Granero, F.O.; Figueiredo, C.C.M.; Silva, L.P.; Silva, R.M.G. Phytoconstituents, antioxidant and antiglycation activity of Chrysophyllum cainito L., Hancornia speciosa Gomes and Plinia glomerata Berg. fruits. An. Da Acad. Bras. De CiÊNcias 2023, 95, e20201853. [Google Scholar] [CrossRef]
- Rodwattanagul, S.; Nimlamool, W.; Okonogi, S. Antioxidant, antiglycation, and anti-inflammatory activities of Caesalpinia mimosoides. Drug Discov. Ther. 2023, 17, 114–123. [Google Scholar] [CrossRef]
- Rodrigues, W.D.; Cardoso, F.N.; Baviera, A.M.; Dos Santos, A.G. In Vitro Antiglycation Potential of Erva-Baleeira (Varronia curassavica Jacq.). Antioxidants 2023, 12, 522. [Google Scholar] [CrossRef]
- Fecka, I.; Bednarska, K.; Kowalczyk, A. In Vitro Antiglycation and Methylglyoxal Trapping Effect of Peppermint Leaf (Mentha× piperita L.) and Its Polyphenols. Molecules 2023, 28, 2865. [Google Scholar] [CrossRef]
- Ali, A.; Shahu, R.; Balyan, P.; Kumari, S.; Ghodmare, R.; Jobby, R.; Jha, P. Antioxidation and antiglycation properties of a natural sweetener: Stevia rebaudiana. Sugar Tech 2022, 563–575. [Google Scholar] [CrossRef]
- Borgo, J.; Laurella, L.C.; Martini, F.; Catalán CA, N.; Sülsen, V.P. Stevia genus: Phytochemistry and biological activities update. Molecules 2021, 26, 2733. [Google Scholar] [CrossRef] [PubMed]
- Rahbar, S.; Figarola, J.L. Novel inhibitors of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 419, 63–79. [Google Scholar] [CrossRef] [PubMed]
- Ramkissoon, J.S.; Mahomoodally, M.F.; Ahmed, N.; Subratty, A.H. Antioxidant and anti–glycation activities correlates with phenolic composition of tropical medicinal herbs. Asian Pac. J. Trop. Med. 2013, 6, 561–569. [Google Scholar] [CrossRef]
- Sero, L.; Sanguinet, L.; Blanchard, P.; Dang, B.T.; Morel, S.; Richomme, P.; Séraphin, D.; Derbré, S. Tuning a 96-well microtiter plate fluorescence-based assay to identify AGE inhibitors in crude plant extracts. Molecules 2013, 18, 14320–14339. [Google Scholar] [CrossRef] [PubMed]
- Rungratanawanich, W.; Qu, Y.; Wang, X.; Essa, M.M.; Song, B.J. Advanced glycation end products (AGEs) and other adducts in aging-related diseases and alcohol-mediated tissue injury. Exp. Mol. Med. 2021, 53, 168–188. [Google Scholar] [CrossRef] [PubMed]
- Biancalana, M.; Koide, S. Molecular mechanism of Thioflavin-T binding to amyloid fibrils. Biochim. Et Biophys. Acta BBA-Proteins Proteom. 2010, 1804, 1405–1412. [Google Scholar] [CrossRef]
- Makino, H.; Emi, H.; Yamaguchi, A.; Iritani, E.; Namiki, N.; Myojo, T.; Yamamoto, K. Environmental and safety issues with nanoparticles. In Nanoparticle Technology Handbook; Elsevier: Amsterdam, The Netherlands, 2008; pp. 385–417. [Google Scholar] [CrossRef]
- Nowicki, S.; Goldblum, R.M. Amniotic fluid and the fetal mucosal immune system. In Mucosal Immunology; Academic Press: Cambridge, MA, USA, 2015; pp. 2251–2268. [Google Scholar] [CrossRef]
- Clarke, D.D. Circulation and energy metabolism of the brain. In Basic Neurochemistry: Molecular, Cellular, and Medical Aspects; Lippincott-Raven: Philadelphia, PA, USA, 1999. [Google Scholar]
- Ajami, M.; Seyfi, M.; Hosseini, F.A.P.; Naseri, P.; Velayati, A.; Mahmoudnia, F.; Zahedirad, M.; Hajifaraji, M. Effects of Stevia on glycemic and lipid profile of type 2 diabetic patients: A randomized controlled trial. Avicenna J. Phytomedicine 2020, 10, 118. [Google Scholar]
- Rubab, U.; Kumar, D.; Farah, M.A.; Al-Anazi, K.M.; Ali, M.A.; Ali, A. Inhibitory roles of Nigella sativa seed extracts on in vitro glycation and aggregation. Phcog. Mag. 2021, 17, S220–S224. [Google Scholar]
- Eruygur, N.; Ayaz, F.; Bağci, Y.; Ayyildiz, H.F.; Çağil, E.M.; Malik, P.; Ali, A. Investigation of phenolic compounds, in vitro antioxidant and enzyme inhibition activities of methanol and aqueous extracts of different parts of Glaucosciadium cordifolium. Bot. Serbica 2022, 46, 239–252. [Google Scholar] [CrossRef]
- Cho, S.; Duong, V.A.; Mok, J.H.; Joo, M.; Park, J.M.; Lee, H. Enrichment and analysis of glycated proteins. Rev. Anal. Chem. 2022, 41, 83–97. [Google Scholar] [CrossRef]
- Nunthanawanich, P.; Sompong, W.; Sirikwanpong, S.; Mäkynen, K.; Adisakwattana, S.; Dahlan, W.; Ngamukote, S. Moringa oleifera aqueous leaf extract inhibits reducing monosaccharide-induced protein glycation and oxidation of bovine serum albumin. Springerplus 2016, 5, 1098. [Google Scholar] [CrossRef] [PubMed]
- Perera, H.K.I.; Premadasa, W.K.V.K.; Poongunran, J. α-glucosidase and glycation inhibitory effects of costus speciosus leaves. BMC Complement. Altern. Med. 2015, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- Adisakwattana, S.; Thilavech, T.; Sompong, W.; Pasukamonset, P. Interaction between ascorbic acid and gallic acid in a model of fructose-mediated protein glycation and oxidation. Electron. J. Biotechnol. 2017, 27, 32–36. [Google Scholar] [CrossRef]
- Jariyapamornkoon, N.; Yibchok-anun, S.; Adisakwattana, S. Inhibition of advanced glycation end products by red grape skin extract and its antioxidant activity. BMC Complement. Altern. Med. 2013, 13, 171. [Google Scholar] [CrossRef]
- Sajithlal, G.B.; Chithra, P.; Chandrakasan, G. The role of metal-catalyzed oxidation in the formation of advanced glycation end products: An in vitro study on collagen. Free. Radic. Biol. Med. 1998, 25, 265–269. [Google Scholar] [CrossRef]
- Ahmad, S.; Khan, H.; Shahab, U.; Rehman, S.; Rafi, Z.; Khan, M.Y.; Ansari, A.; Siddiqui, Z.; Ashraf, J.M.; Abdullah, S.M.; et al. Protein oxidation: An overview of metabolism of sulphur containing amino acid, cysteine. Front. Biosci. Sch. 2017, 9, 71–87. [Google Scholar] [CrossRef]
- Mohd Dom, N.S.; Yahaya, N.; Adam, Z.; Hamid, M. Antiglycation and antioxidant properties of Ficus deltoidea varieties. Evid.-Based Complement. Altern. Med. 2020, 2020, 6374632. [Google Scholar] [CrossRef]
- Wu, C.H.; Huang, S.M.; Lin, J.A.; Yen, G.C. Inhibition of advanced glycation endproduct formation by foodstuffs. Food Funct. 2011, 2, 224–234. [Google Scholar] [CrossRef]
- Kumar, D.; Shah, N.; Al-Anazi, K.M.; Farah, M.A.; Ali, M.A.; Lee, J.; Ali, A. Potential role of Tribulus terrestris fruit extract in inhibition of advanced glycation end products. Pharmacogn. Mag. 2022, 18, 341. [Google Scholar]
- Escutia-López, K.N.; González-Montoya, M.; Cruz-Ortiz, R.; Cano-Sampedro, E.; Rodríguez-Rivero, Y.; Sánchez-Pardo, M.E.; Mora-Escobedo, R. Effect of aqueous Stevia rebaudiana Bertoni extract in antioxidant and antiglycation capacity in vitro. Rev. Bio Cienc. 2020, 7, e875. [Google Scholar] [CrossRef]
- Awasthi, S.; Saraswathi, N.T. Silybin, a flavonolignan from milk thistle seeds, restrains the early and advanced glycation end product modification of albumin. RSC Adv. 2015, 5, 87660–87666. [Google Scholar]
- Brownlee, M. The pathological implications of protein glycation. Clin. Investig. Med. Med. Clin. Et Exp. 1995, 18, 275–281. [Google Scholar]
- Adisakwattana, S.; Thilavech, T.; Chusak, C. Mesona Chinensis Benth extract prevents AGE formation and protein oxidation against fructose-induced protein glycation in vitro. BMC Complement. Altern. Med. 2014, 14, 130. [Google Scholar] [CrossRef] [PubMed]
- Alvi, S.S.; Nabi, R.; Khan, M.; Akhter, F.; Ahmad, S.; Khan, M.S. Glycyrrhizic acid scavenges reactive carbonyl species and attenuates glycation-induced multiple protein modification: An in vitro and in silico study. Oxidative Med. Cell. Longev. 2021, 2021, 7086951. [Google Scholar] [CrossRef]
- Pawlukianiec, C.; Gryciuk, M.E.; Mil, K.M.; Żendzian-Piotrowska, M.; Zalewska, A.; Maciejczyk, M. A new insight into meloxicam: Assessment of antioxidant and anti-glycating activity in in vitro studies. Pharmaceuticals 2020, 13, 240. [Google Scholar] [CrossRef]
- Suji, G.; Sivakami, S. DNA damage by free radical production by aminoguanidine. Ann. N. Y. Acad. Sci. 2006, 1067, 191–199. [Google Scholar] [CrossRef]
- Kumar, D.; Ali, A. Antiglycation and antiaggregation potential of thymoquinone. Nat. Volatiles Essent. Oils 2019, 6, 25–33. [Google Scholar]
- Deve, S.A.; Sathish Kumar, T.; Kumaresan, K.; Rapheal, V.S. Extraction process optimization of polyphenols from Indian Citrus sinensis–as novel antiglycative agents in the management of diabetes mellitus. J. Diabetes Metab. Disord. 2014, 13, 1–10. [Google Scholar] [CrossRef]
- Mahapatra, D.K.; Bharti, S.K. Biologically Active Small Molecules Modern Applications and Therapeutic Perspectives, 1st ed.; Apple Academic Press: Palm Bay, FL, USA; CRC Press: Boca Raton, FL, USA, 2023; p. 2. [Google Scholar]
- Jayaraman, S.; Manoharan, M.S.; Illanchezian, S. In-vitro antimicrobial and antitumor activities of Stevia rebaudiana (Asteraceae) leaf extracts. Trop. J. Pharm. Res. 2008, 7, 1143–1149. [Google Scholar] [CrossRef]
- Rizzo, B.; Zambonin, L.; Angeloni, C.; Leoncini, E.; Vieceli Dalla Sega, F.; Prata, C.; Fiorentini, D.; Hrelia, S. Steviol glycosides modulate glucose transport in different cell types. Oxidative Med. Cell. Longev. 2013, 2013, 348169. [Google Scholar] [CrossRef]
- Kulkarni, A.S.; Karkhanis, D.; Barve, S.S.; Sharma, S. Antioxidant and cytotoxic study of extract of vegetative and flowering stages leaves Stevia rebaudiana. J. Pharmacogn. Phytochem. 2021, 10, 303–306. [Google Scholar]
- Hanna, D.H.; Osailan, R.; Ahmed, H.A. Stevia rebaudiana Methanolic Leaf Extract in Egypt: Phytochemical Analysis, Antioxidant, Antilipid Peroxidation, Antihemolytic, Antimetastatic, and Anticancer Properties. J. Food Biochem. 2023, 2023, 7161091. [Google Scholar] [CrossRef]
- Farid, A.; Hesham, M.; El-Dewak, M.; Amin, A. The hidden hazardous effects of Stevia and sucralose consumption in male and female albino mice in comparison to sucrose. Saudi Pharm. J. 2020, 28, 1290–1300. [Google Scholar] [CrossRef]
- Awney, H.A.; Massoud, M.I.; El-Maghrabi, S. Long-term feeding effects of stevioside sweetener on some toxicological parameters of growing male rats. J. Appl. Toxicol. 2011, 31, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Geeraert, B.; Crombe, F.; Hulsmans, M.; Benhabiles, N.; Geuns, J.M.; Holvoet, P. Stevioside inhibits atherosclerosis by improving insulin signaling and antioxidant defense in obese insulin-resistant mice. Int. J. Obes. 2010, 34, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Mohd-Radzman, N.H.; Ismail, W.I.W.; Jaapar, S.S.; Adam, Z.; Adam, A. Stevioside from Stevia rebaudiana Bertoni increases insulin sensitivity in 3T3-L1 adipocytes. Evid.-Based Complement. Altern. Med. 2013, 2013, 938081. [Google Scholar] [CrossRef] [PubMed]
- Bhasker, S.; Madhav, H.; Chinnamma, M. Molecular evidence of insulinomimetic property exhibited by steviol and stevioside in diabetes induced L6 and 3T3L1 cells. Phytomedicine 2015, 22, 1037–1044. [Google Scholar] [CrossRef]
- Elnaga, N.A.; Massoud, M.I.; Yousef, M.I.; Mohamed, H.H. Effect of Stevia sweetener consumption as non-caloric sweetening on body weight gain and biochemical’s parameters in overweight female rats. Ann. Agric. Sci. 2016, 61, 155–163. [Google Scholar] [CrossRef]
- Celiktas, O.Y.; Bedir, E.; Sukan, F.V. In vitro antioxidant activities of Rosmarinus officinalis extracts treated with supercritical carbon dioxide. Food Chem. 2007, 101, 1457–1464. [Google Scholar] [CrossRef]
- Woelwer-Rieck, U.; Lankes, C.; Wawrzun, A.; Wüst, M. Improved HPLC method for the evaluation of the major steviol glycosides in leaves of Stevia rebaudiana. Eur. Food Res. Technol. 2010, 231, 581–588. [Google Scholar] [CrossRef]
- Meeprom, A.; Sompong, W.; Chan, C.B.; Adisakwattana, S. Isoferulic acid, a new anti-glycation agent, inhibits fructose-and glucose-mediated protein glycation in vitro. Molecules 2013, 18, 6439–6454. [Google Scholar] [CrossRef] [PubMed]
- Thornalley, P.J.; Rabbani, N. Detection of oxidized and glycated proteins in clinical samples using mass spectrometry-a user’s perspective. Biochim. Et Biophys. Acta BBA-Gen. Subj. 2014, 1840, 818–829. [Google Scholar] [CrossRef] [PubMed]
- Nabi, R.; Alvi, S.S.; Khan, R.H.; Ahmad, S.; Ahmad, S.; Khan, M.S. Antiglycation study of HMG-R inhibitors and tocotrienol against glycated BSA and LDL: A comparative study. Int. J. Biol. Macromol. 2018, 116, 983–992. [Google Scholar] [CrossRef]
- Anandan, S.; Urooj, A. Bioactive compounds from Morus indica as inhibitors of advanced glycation end products. Indian J. Pharm. Sci. 2019, 81, 282–292. [Google Scholar] [CrossRef]
- Bouma, B.; Kroon-Batenburg, L.M.; Wu, Y.P.; Brünjes, B.; Posthuma, G.; Kranenburg, O.; de Groot, P.G.; Voest, E.E.; Gebbink, M.F. Glycation induces formation of amyloid cross-β structure in albumin. J. Biol. Chem. 2003, 278, 41810–41819. [Google Scholar] [CrossRef] [PubMed]
- Tupe, R.S.; Agte, V.V. Role of zinc along with ascorbic acid and folic acid during long-term in vitro albumin glycation. Br. J. Nutr. 2010, 103, 370–377. [Google Scholar] [CrossRef]
- Sathiyamoorthy, J.; Sudhakar, N. In vitro cytotoxicity and apoptotic assay in HT-29 cell line using Ficus hispida Linn: Leaves extract. Pharmacogn. Mag. 2017, 13 (Suppl. 4), S756. [Google Scholar]
- van de Venter, M.; Roux, S.; Bungu, L.C.; Louw, J.; Crouch, N.R.; Grace, O.M.; Maharaj, V.; Pillay, P.; Sewnarian, P.; Bhagwandin, N.; et al. Antidiabetic screening and scoring of 11 plants traditionally used in South Africa. J. Ethnopharmacol. 2008, 119, 81–86. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Inhibition | Primary Stage | Intermediate Stage | Advanced Stage | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Browning | Fructosamine | Carbonyl Content | Total AGEs | Individual AGEs | Congo Red | ThT | ||||
| Argpyrimidine | Vesperlysine | Pentosidine | Crossline | |||||||
| AG | 85 | 70 | 74 | 35 | 30 | 45 | 35 | 42 | 50 | 19 |
| Stevia | 71 | 67 | 64 | 81 | 82 | 81 | 80 | 70 | 47 | 31 |
| AGEs | Excitation Spectrum (nm) | Emission Spectrum (nm) |
|---|---|---|
| Argpyrimidine | 320 | 380 |
| Vesperlysine | 350 | 405 |
| Pentosidine | 335 | 385 |
| Crossline | 380 | 440 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahu, R.; Kumar, D.; Ali, A.; Tungare, K.; Al-Anazi, K.M.; Farah, M.A.; Jobby, R.; Jha, P. Unlocking the Therapeutic Potential of Stevia rebaudiana Bertoni: A Natural Antiglycating Agent and Non-Toxic Support for HDF Cell Health. Molecules 2023, 28, 6797. https://doi.org/10.3390/molecules28196797
Shahu R, Kumar D, Ali A, Tungare K, Al-Anazi KM, Farah MA, Jobby R, Jha P. Unlocking the Therapeutic Potential of Stevia rebaudiana Bertoni: A Natural Antiglycating Agent and Non-Toxic Support for HDF Cell Health. Molecules. 2023; 28(19):6797. https://doi.org/10.3390/molecules28196797
Chicago/Turabian StyleShahu, Rinkey, Dinesh Kumar, Ahmad Ali, Kanchanlata Tungare, Khalid Mashay Al-Anazi, Mohammad Abul Farah, Renitta Jobby, and Pamela Jha. 2023. "Unlocking the Therapeutic Potential of Stevia rebaudiana Bertoni: A Natural Antiglycating Agent and Non-Toxic Support for HDF Cell Health" Molecules 28, no. 19: 6797. https://doi.org/10.3390/molecules28196797
APA StyleShahu, R., Kumar, D., Ali, A., Tungare, K., Al-Anazi, K. M., Farah, M. A., Jobby, R., & Jha, P. (2023). Unlocking the Therapeutic Potential of Stevia rebaudiana Bertoni: A Natural Antiglycating Agent and Non-Toxic Support for HDF Cell Health. Molecules, 28(19), 6797. https://doi.org/10.3390/molecules28196797