Abstract
The gut–brain axis is an active area of research. Several representative diseases, including central nervous system disorders (Alzheimer’s disease, Parkinson’s disease, and depression), metabolic disorders (obesity-related diseases), and intestinal disorders (inflammatory bowel disease and dysbiosis), are associated with the dysfunctional gut–brain axis. Baicalin, a bioactive flavonoid extracted from Scutellaria baicalensis, is reported to exert various pharmacological effects. This narrative review summarizes the molecular mechanisms and potential targets of baicalin in disorders of the gut–brain axis. Baicalin protects the central nervous system through anti-neuroinflammatory and anti-neuronal apoptotic effects, suppresses obesity through anti-inflammatory and antioxidant effects, and alleviates intestinal disorders through regulatory effects on intestinal microorganisms and short-chain fatty acid production. The bioactivities of baicalin are mediated through the gut–brain axis. This review comprehensively summarizes the regulatory role of baicalin in gut–brain axis disorders, laying a foundation for future research, although further confirmatory basic research is required.
1. Introduction
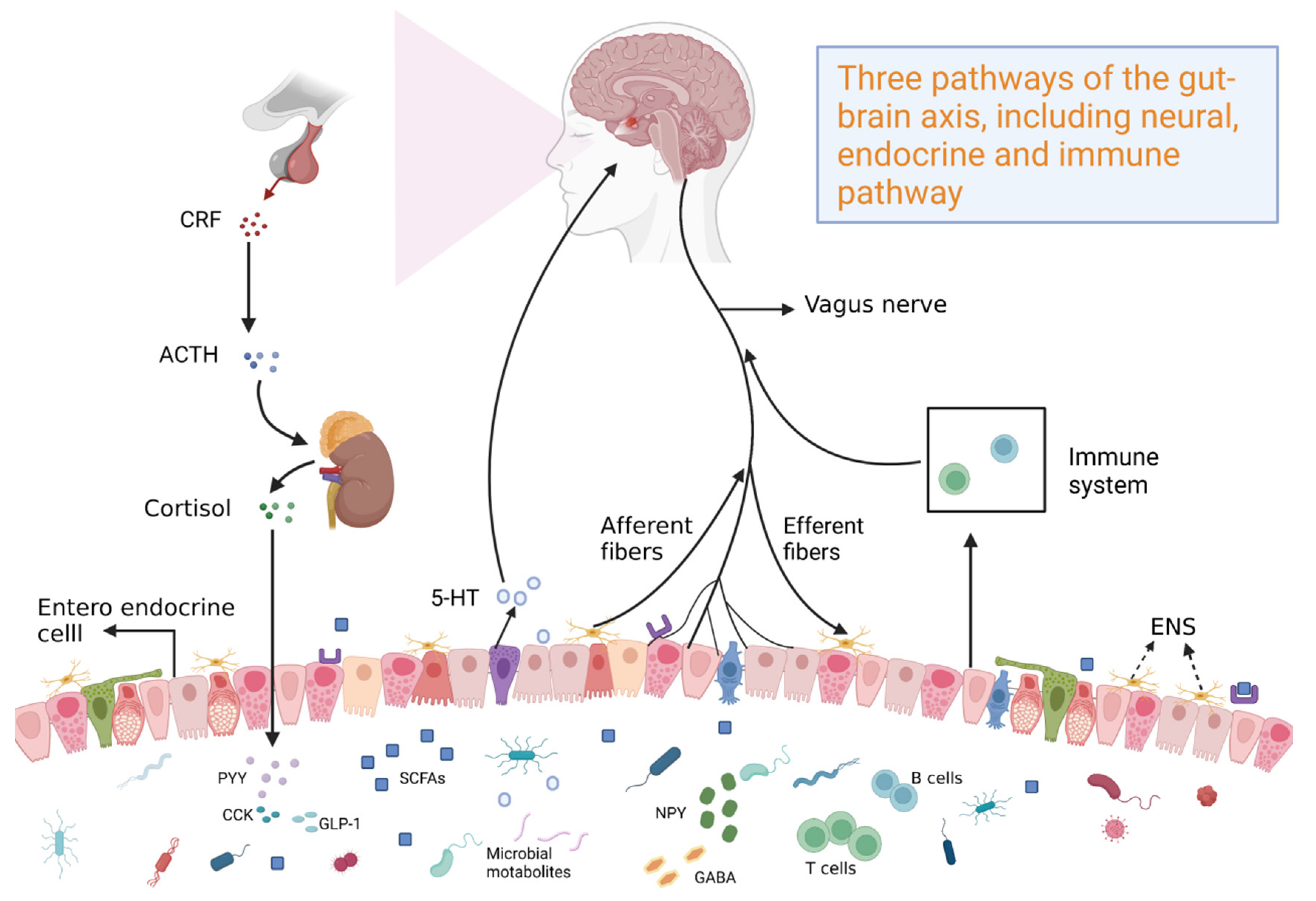
At the end of the twentieth century, research on neuroanatomy and the function and pathophysiology of gut–brain interaction rapidly progressed. Additionally, the field of neurogastroenterology and the concept of the gut–brain axis were established, which linked neuropsychiatry with gastroenterology. Theoretical understanding of the effects of the gut has markedly improved. The elucidation of the gut–brain axis revealed the bidirectional regulatory effects between the brain and the gut (Figure 1). In particular, the brain can project nerve impulses to affect intestinal microbiota and physiological function [1]. Conversely, intestinal microbiota can release metabolites, cytokines, and other signaling mediators to the central nervous system (CNS) through neuro-immune pathways, affecting the physiological function of the brain [2]. Currently, the gut–brain axis is suggested to be connected by the mutual activities of the CNS, the autonomic nervous system, and the enteric nervous system (ENS) [3]. The parasympathetic nerve is suggested to be a bridge connecting the CNS and the ENS. Thus, parasympathetic nerves, the CNS, and the ENS form the basic framework of the gut–brain axis [4,5]. However, previous studies have demonstrated the role of parasympathetic nerves in the regulation of the gut–brain axis [6,7]. The ENS is independent, as evidenced by its ability to regulate gastrointestinal functions without CNS innervation (e.g., gastrointestinal motility and secretion, intestinal blood flow, intestinal epithelial material transport, gastrointestinal immune responses, and regulation of inflammatory processes) [8]. Additionally, the ENS transmits gastrointestinal sensory signals out of the CNS and sympathetic ganglia through projection neurons [9]. The CNS occupies a dominant position, as evidenced by its regulatory effects on the ENS, mainly through gut microbiota, microbial products, and microbially produced metabolites in neuroinflammation and gastrointestinal diseases. The regulatory effect of the CNS on the gastrointestinal tract is mediated through the CNS-mediated integration of internal and external stimuli, which are transmitted to the enteric plexus through the nerve or neuroendocrine system or directly act on the gastrointestinal cells. Additionally, the hypothalamic–pituitary–adrenal (HPA) axis regulates the effects of stress on the gastrointestinal tract and gastrointestinal inflammation [10]. The gut–brain axis regulates gestational conditions, such as irritable bowel syndrome (IBS), inflammatory bowel disease (IBD), and gut microbiota composition. Among functional gastrointestinal diseases, IBS is the most prevalent disorder involving gut–brain interactions that are regulated by the gut–brain axis, which is related to intestinal motility and paresthesia, inflammation and immunity, and mental and psychological processes. These findings indicate the role of the gut–brain axis in the cross-talk between neurological disorders and intestinal disorders.
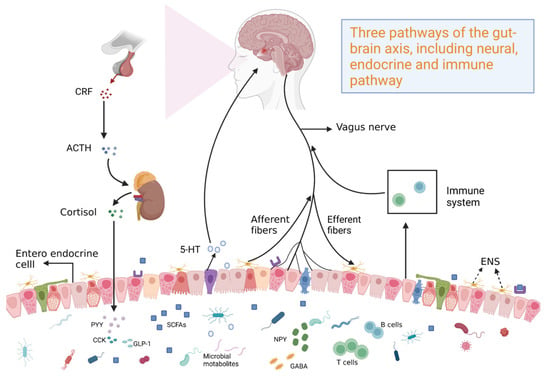
Figure 1.
Profiles of bidirectional regulation of the gut–brain axis.
To understand the gut–brain axis, the brain and the gut must be considered a whole system that interacts. The brain and the gastrointestinal system mutually interact through crisscrossed main and collateral channels of the whole body. In particular, the physiological function of the brain depends on the nourishment of qi and blood produced by the gastrointestinal system [11]. Additionally, the emotional activities determined by the brain are related to the function of the intestine. For example, the syndrome of liver qi stagnation and spleen deficiency in traditional Chinese medicine (TCM) is caused by excessive emotional pressure, which leads to the physiological imbalance of spleen and stomach functions and symptoms, such as abdominal distension, pain, and diarrhea [12]. Although ancient Chinese medicine did not have a specific concept of the gut–brain axis, holistic treatments linked the brain and the intestine emerged and different TCM therapies were developed to alleviate relevant diseases based on the gut–brain axis. Among the various active natural products of TCM, several components modulate neurological disorders and intestinal disorders through the regulation of the gut–brain axis.
Scutellaria baicalensis is a perennial herb of the Lamiaceae family. The dried roots are often used as a traditional Chinese herbal medicine called Huangqin [13]. These roots, which have been popular since ancient times, contain several bioactive components, such as baicalin, baicalein, baicalein II, and wogonin [14,15]. In TCM, Huangqin is commonly used to stop diarrhea, lower blood pressure, relieve insomnia, and treat respiratory infections [16]. Baicalin, a bioactive flavonoid extracted from Scutellaria baicalensis, is a typical multi-target, multi-channel agent with multiple pharmacological effects, including hepatoprotective, antitumor, antibacterial, anti-inflammatory, antidepressant, and antioxidant effects [17,18]. For example, Chung et al. demonstrated that baicalin inhibits breast cancer by suppressing transforming growth factor (TGF)-β1-mediated epithelial–mesenchymal transition through the upregulation of the nuclear factor (NF)-κB pathway [19]. The value of natural products in the development and utilization of new drugs has piqued the interest of the scientific community. Baicalin has great research value owing to its wide range of pharmacological activities [20]. A recent study demonstrated that baicalin promotes the interaction of the liver–gut axis by regulating GPBAR1, farnesoid X receptor, bile acids, and microbiota [20]. This review aimed to elucidate the effects of baicalin on the gut–brain axis.
In this review, studies focusing on the therapeutic effects of baicalin on neurological and intestinal disorders mediated through the gut–brain axis were retrieved from the PubMed, Cochrane Library Web of Science, and EMBASE databases over the last five years. Studies that lacked scientific value or had obvious errors were excluded. All articles were used to systematically analyze the pharmacological effects of baicalin on the gut–brain axis, providing references for future studies.
2. Effects of Baicalin Functions on the Gut–Brain Axis
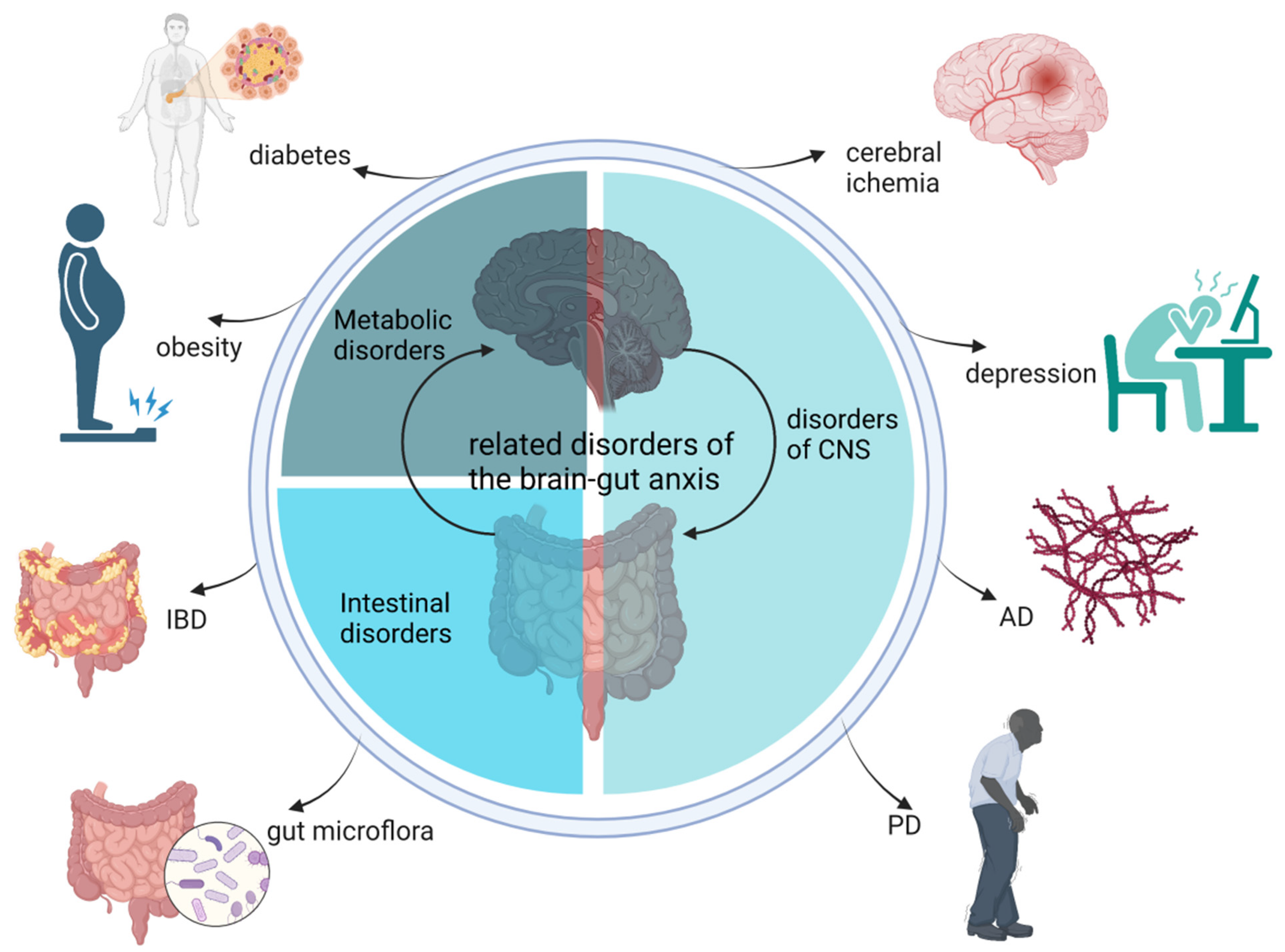
Several representative diseases, including CNS disorders (Alzheimer’s disease (AD), Parkinson’s disease (PD), and depression), metabolic disorders (obesity-related diseases), and intestinal disorders (IBD and regulation of the gut microbiota) are associated with the dysfunctional gut–brain axis (Figure 2).
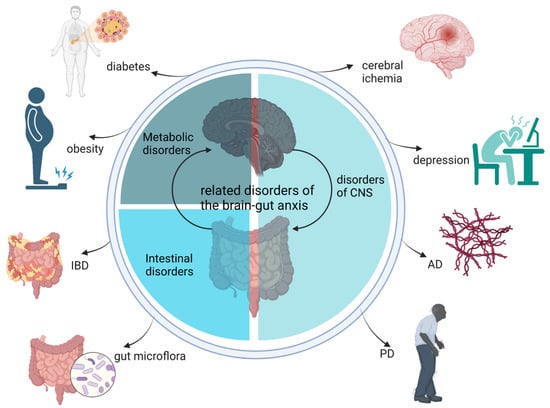
Figure 2.
Related disorders in the gut–brain axis. AD, Alzheimer’s disease; PD, Parkinson’s disease; IBD, inflammatory bowel disease.
2.1. Baicalin Alleviates CNS Disorders through the Gut–Brain Axis
The gut microbiota composition is dysregulated under stress conditions. This leads to the production of inflammatory mediators, such as IFN-γ, IL-1β, and IL-6, as well as the downregulation of short-chain fatty acids (SCFAs). These peripherally derived inflammatory factors may disrupt the blood–brain barrier (BBB) and migrate to the CNS, affecting the morphology of glial cells and microglia.
2.1.1. Baicalin Mitigates Depression through the Gut–Brain Axis
A recent study reported that in the prefrontal cortex of chronically stressed rats, an increase in NF-κB, NLRP3, and IL-1β were upregulated in the prefrontal cortex of chronically stressed rats, which was accompanied by microglial activation and astrocyte injury [21]. Changes in these glial cells affect brain networks involved in memory, learning, mood, and emotion regulation, which can result in the onset of depression or anxiety disorders [22]. In developed countries, depression, a serious mental disorder, is considered to be one of the main causes of disability [23]. Depression is mainly characterized by low mood, anhedonia, irritability, concentration difficulties, and changes in appetite and sleep [24,25]. Previous studies have demonstrated that depression is related to impaired psychological health, cognitive impairment, increased risk of suicidal behavior, and increased mortality [26,27]. The pathophysiology of depression has not been completely elucidated. The monoaminergic deficiency hypothesis is the most commonly used hypothesis to explain depressive symptoms. According to this hypothesis, the levels of serotonin, norepinephrine, and dopamine are downregulated in the synaptic cleft of patients with depression [28]. Inflammatory processes, including the upregulation of pro-inflammatory cytokine levels and microglial activation, may influence behavior and mood [29]. Recent studies have reported that baicalin alleviates depression by inhibiting neuroinflammation. Guo et al. demonstrated that baicalin (60 mg/kg bodyweight) downregulated the levels of IL-1β, interleukin-6 (IL-6), and tumor necrosis factor-α (TNF-α) levels, decreased TLR4, and increased upregulated the phosphorylation of phosphatidylinositol 3-kinase (PI3K), Akt1, and Foxo1 in the hippocampus of the chronic unpredictable mild stress (CUMS)-induced depression mouse model. Treatment with 100 μM baicalin reversed damages in BV2 microglial cell lines. These results suggest that baicalin mitigates neuroinflammation-induced depression by downregulating TLR4 expression through the PIK3R1/AKT1/FOXO1 pathway [30]. Zhang et al. reported that baicalin (20 and 40 mg/kg bodyweight) suppressed CUMS-induced hippocampal apoptosis and inflammatory cytokines (Casp1 and IL-1β) and upregulated DCXcx, Eno2, and Bdnf in the hippocampus. These results suggest that baicalin exerts neuroprotective effects on the rat model of depression by inhibiting the Gsk3b/Rela/NLRP3 pathway [31]. Intestinal microbiota promotes NLRP3-mediated neuroinflammation in the hippocampus and mediates chronic ethanol exposure-induced depressive-like behavior [32]. Additionally, baicalin (20 and 40 mg/kg bodyweight) significantly mitigated the CUMS-induced activation of the NLRP3 inflammasome and upregulation of pro-inflammatory cytokines (IL-1β and IL-6) in the prefrontal cortex of rats, suggesting that baicalin exerts an antidepressant effect by inhibiting NLRP3inflammasome activation in the rat prefrontal cortex through the regulation of intestinal flora [33]. One study demonstrated that baicalin (60 mg/kg bodyweight) significantly promoted neuronal differentiation, increased the number of Dcx+ cells, and mitigated the CUMS-induced downregulation of p-Akt1, Foxg1, and Fgf2. These results suggest that baicalin exerts antidepressant effects by promoting neuronal differentiation through the Akt1/Foxg1 pathway [34]. Fang et al. reported that baicalin (50 mg/kg bodyweight) alleviated inflammatory pain-related depressive symptoms through Akt1-mediated adult hippocampal neurogenesis and promoted the growth and differentiation of hippocampal neural stem cells [35]. This indicated that baicalin mitigates depression and alleviates the altered behavior and hippocampal inflammation by targeting the Akt1 pathway through the modulation of gut microbiota composition and metabolites [36]. Li et al. established a mouse model of diarrhea-predominant IBS (IBS-D) using chronic restraint stress plus Senna Alexandrina Mill decoction. The IBS-D mouse model was then treated with baicalin (0.935 mg/d). The symptoms of depression, irritability, loss of appetite, and varying degrees of loose stools were alleviated in the experimental group. Baicalin downregulated the colonic levels of 5-hydroxytryptamine (5-HT) and Vip and the serum levels of 5-HT and Chat. Additionally, baicalin downregulated the colonic expression of NF-κBRela and regulated the level of basophil granulocytes and leukomonocytes in the whole blood, suggesting that baicalin alleviates inflammation. This study also indicated the critical role of baicalin in modulating the abundance of intestinal flora (Bacteroidia, Deferribacteres, Verrucomicrobia, Candidatus_Saccharibacteria, and Cyanobacteria) [37]. Baicalin treatment (20 and 40 mg/kg bodyweight) modulated Sirt1 and downregulated the level of Rela acetylation in the hippocampus and hypothalamus and the levels of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) in mice with depression, suggesting that administration of baicalin may alleviate depression by modulating the Sirt1-NF-κB pathway [38]. SIRT1 is reported to be an important mediator of host-microbiome interactions and prevents intestinal inflammation by regulating the gut microbiota [39]. These findings suggest that baicalin exerts antidepressant effects by modulating the gut–brain axis, especially the gut microbes (Table 1).

Table 1.
Mechanisms of baicalin in gut–brain axis related disorders.
2.1.2. Baicalin Alleviates Cerebral Ischemia through the Gut–Brain Axis
Cerebral ischemia can damage the intestinal mucosal barrier and induce morphological damage in the intestinal mucosa, leading to intestinal dysbiosis and inflammation [60,61]. At month 12 post-cerebral ischemia, the serum levels of pro-inflammatory cytokines IFN-g, IL-6, and TNF-α are significantly upregulated. These pro-inflammatory cytokines from the intestinal tract can directly interact with each other, exacerbating pathological changes [62]. Cerebral ischemia, a global health issue caused by temporary or permanent, partial or total interruption of cerebral blood flow, is the second-leading cause of death and the third-leading cause of disability worldwide [63]. Based on the underlying neuropathology, stroke is classified as ischemic or hemorrhagic, with ischemic stroke accounting for 85% of all cases. Ischemic stroke is primarily caused by blockage of the middle cerebral artery, resulting in damage to the brain parenchyma in the affected area, followed by the induction of neuroinflammatory and immune responses [64]. Approximately 90% of stroke cases are reported to be related to behavioral factors, including malnutrition, low physical activity, and smoking, and metabolic factors, including diabetes, obesity, hyperlipidemia, and hypertension [65]. Brain damage from an ischemic stroke results from a complex series of neuropathological and neuropathological events, including excitotoxicity, oxidative stress, neuroinflammation, apoptosis, and amyloid production [66]. This section focuses on the correlation between the therapeutic effect of baicalin on cerebral ischemia and the brain–gut axis.
Liu et al. demonstrated that baicalin (50–100 mg/kg bodyweight) mitigated the cerebral ischemia and reperfusion-induced upregulation of trimethylamine (TMA) and TMA nitroxide and clusterin in middle cerebral artery occlusion (MCAO) mice. Additionally, treatment with baicalin (50 or 100 mg/kg bodyweight) significantly restored the level of intestinal flora, including the upregulation of the relative abundance of Halomonas_smyrnensis and the downregulation of the relative abundance of Parabacteroides_johnsonii and Bacteroides_uniformis in MCAO mice. The abundance of Citromicrobium_sp_WPS32 and Eubacterium_sp_CAG_86 was upregulated in MCAO mice treated with baicalin (100 mg/kg bodyweight), whereas that of Lactobacillus_plantarum was downregulated. Baicalin improved the cognition, memory, long-term potentiation, brain functional connectivity, and hippocampal neuronal plasticity of MACO mice. The intestinal flora of MACO mice was destroyed using broad-spectrum antibiotics. Treatment with antibiotics mitigated the neuroprotective effect of baicalin, which suggested that baicalin exerts neuroprotective effects in mice with cerebral ischemia-reperfusion injury by remodeling intestinal microorganisms [40]. Cerebral ischemia interrupts the transmission of GBA signals through the HPA axis [67]. Li demonstrated that baicalin (15 mg/kg bodyweight) mitigated the MCAO-induced neurological deficit and infarct volume by increasing the level of serum adrenocorticotropic hormone and the expression of the neuronal nucleus, glial fibrillary acidic protein, progesterone receptor, and serum progesterone in the ischemic penumbra in MCAO rats [41]. Hyperglycemia, which is an important risk factor for cerebral ischemia, promotes excessive activation of the HPA axis, leading to intestinal dysfunction [68,69]. Li et al. demonstrated that baicalin (100 mg/kg bodyweight) could regulate mitochondrial function through AMPK, alleviate hyperglycemia, and attenuate brain damage [42]. Although these two studies did not analyze GBA, we hypothesize that baicalin may regulate the HPA axis through different mechanisms and consequently affect the function of GBA and alleviate brain damage.
2.1.3. Baicalin Alleviates AD through the Gut–Brain Axis
In AD, alterations in gut microbial composition promote the release of lipopolysaccharide (LPS) and amyloid protein [70]. Amyloid beta (Aβ) hyperactivates microglia, stimulating the NF-κB signaling pathway and promoting the release of pro-inflammatory cytokines, reactive oxygen species (ROS), and reactive nitrogen species, leading to neuronal and glial cell death [71]. AD, a progressive multifactorial neurodegenerative disease, is characterized by severe deficits in memory, cognition, and motor functions, which result in decreased mental, behavioral, and functional activities and poor quality of life [72]. The total number of people living with AD is projected to reach 139 million in 2050, making it a global health challenge [73].
Cognitive dysfunction may be due to the deposition of extraneuronal Aβ plaques and intraneuronal tau tangles, leading to neuroinflammation, altered calcium homeostasis, vascular degeneration, and ultimately neuronal death [74]. Dietary flavonoids regulate the NF-κB signaling pathway, which is a potential therapeutic target for AD. Flavonoids, a subtype of polyphenols, modulate microbiota-related metabolism, improve neurological health, activate the anti-apoptotic survival signaling pathways, upregulate antioxidant gene expression, and alleviate Aβ pathology, exerting therapeutic effects on neuronal dysfunction and toxicity. Flavonoids and other sugars that are not absorbed in the small intestine are broken down by the intestinal microbiota into phenolic acids and other metabolites (such as galactose and uronic acid), which inhibit the growth of Ruminococcus gaunii, Bacteroides, and Lactobacillus. Harach et al. reported that the number of Bacteroidetes was downregulated in the intestine of APPS1 mice, while Aβ-induced pathology was alleviated in the brain of sterile amyloid precursor protein (APP) transgenic mice [75], which suggested that changes in the number of microbial strains may lead to amyloid protein deposition, triggering an immune response and consequently leading to AD. Additionally, flavonoids play an important role in the prevention of neuritis as evidenced by the inhibition of inflammatory cytokine (TNF-α and IL-1β) production in microglia and Nos2 and ROS production in glial cells and the downregulation of pro-inflammatory transcription factors, such as NF-κB, which are mediated through the regulation of the glial and neuronal signaling pathways [76]. Baicalin, a herbal flavonoid, was demonstrated to exert these effects in several studies. Jin et al. reported that baicalin (100 mg/kg bodyweight) alleviates spatial memory impairment in Aβ precursor protein/presenilin-1 (PS1) mice and effectively reduces the levels of pro-inflammatory factors (IL-6, TNF-α, IL-1β) and neuroinflammation-mediated neuronal apoptosis. These results suggested that baicalin can inhibit microglia-induced neuroinflammation, alleviate cognitive impairment, and protect neurons by inhibiting the activation of NLRP3 inflammasome and TLR4/NF-κB signaling pathway [43]. The accumulation and deposition of Aβ in senile plaques and cerebral blood vessels may contribute to the progressive neurodegeneration in AD. The oral administration of baicalin (30, 50, and 100 mg/kg bodyweight) attenuated Aβ1–42 protein-induced glial cell activation and upregulation of TNF-α and IL-6, suggesting that baicalin ameliorates Aβ1–42 protein-related pathology and cognitive dysfunction by exerting anti-inflammatory effects. Thus, baicalin is a potential therapeutic drug for AD [44]. Similarly, baicalin (50, 100, and 200 mg/kg bodyweight) treatment restored the activity of antioxidant enzymes (superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx)) and upregulated their gene expression, improving antioxidant capacity, inhibiting Bax/Bcl2 expression, cytochrome c release, and CASP9/CASP3 activation. Baicalin can effectively improve Aβ-induced learning and memory impairment, hippocampal damage, and neuronal apoptosis [45]. Additionally, baicalin (50 and 100 μM) inhibited the β-amyloid peptide (Aβ42)-induced proliferation of BV2 microglial cells, downregulated the expression of CD11b, decreased the chemotactic ability of BV2 cells, significantly inhibited the production of IL-6, TNF-α and NO, and suppressed Aβ-induced phosphorylation of Jak2 and STAT3, suggesting that baicalin can prevent Aβ-induced microglial activation by inhibiting the JAK2/STAT3 signaling pathway [46].
2.1.4. Baicalin Alleviates PD through the Gut–Brain Axis
In PD, gut dysbiosis stimulates the production of inflammatory cytokines and LPS, leading to intestinal epithelial damage and impaired barrier integrity [77]. Increased intestinal permeability leads to microbial translocation and the introduction of bacterial toxins and host-derived inflammatory cytokines (TNF-α, IL-6, and IL-10) into the bloodstream, directly contributing to the nervous system impairment owing to the disruption of the BBB integrity. The interaction provides a pathway [78], and these cytokines are significantly elevated in the serum of patients with PD, suggesting that the gut–brain axis may be associated with symptom severity and disease progression in PD [79].
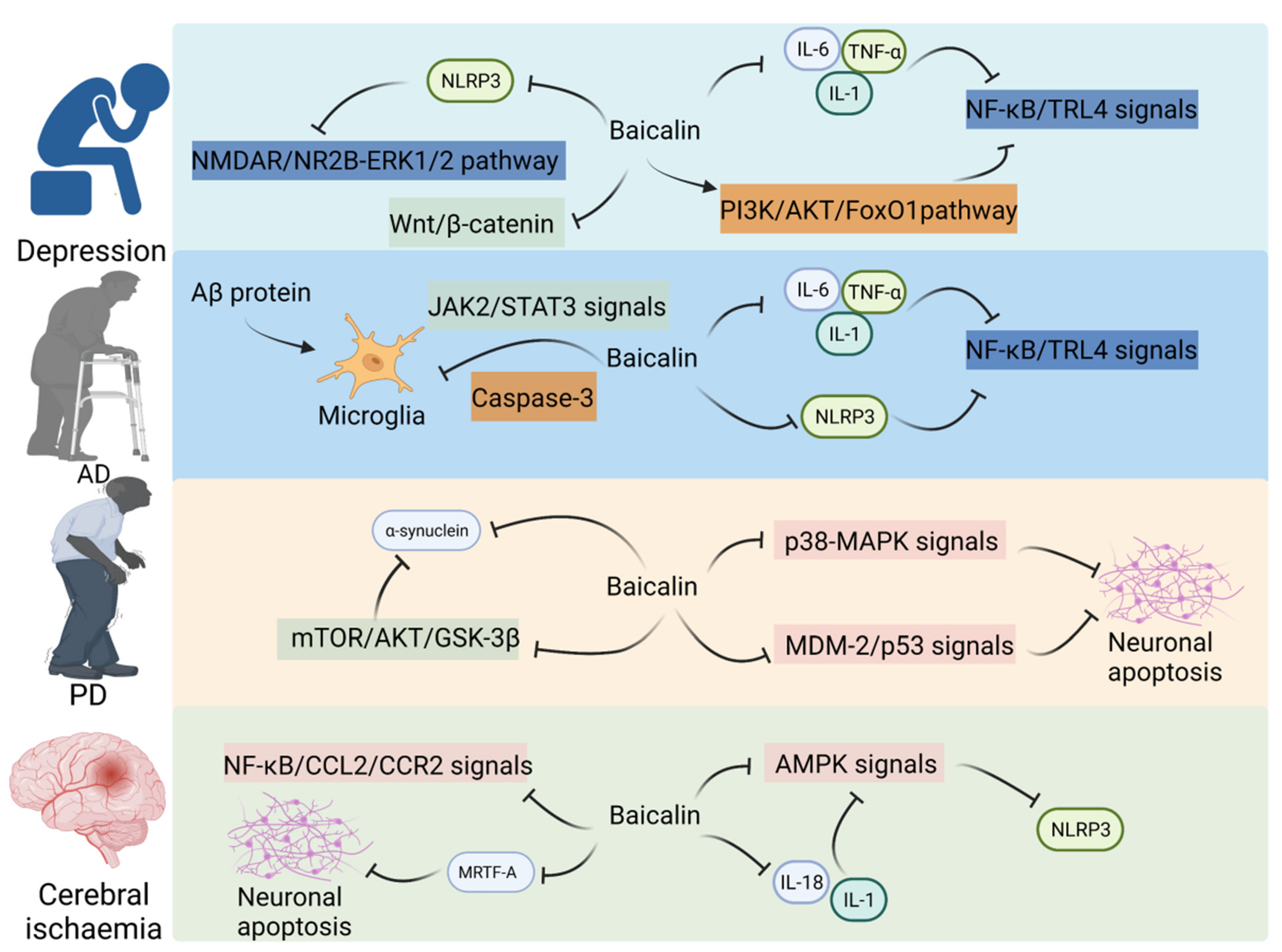
Dodiya et al. demonstrated that low-dose oral rotenone-induced intestinal hyper-permeability and/or microbial dysbiosis leads to pro-inflammatory intestinal milieu, aggravating PD phenotype in the PD mouse model, suggesting that the microbiota–gut–brain axis is involved in the pathogenesis of PD [80]. Lei et al. reported that baicalin (20 and 40 mg/kg bodyweight) protected dopaminergic (DA) neurons against ROS-induced damage and downregulated the expression of Cebpb and Snca, and alleviated the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced loss of substantia nigra DA neurons in PD mice. These results suggest that baicalin exerts therapeutic effects on PD by inhibiting the expression of Cebpb [47]. One study revealed that baicalin (25 mg/kg bodyweight) significantly mitigated 6-hydroxydopamine (6-OHDA)-induced neuronal apoptosis in PD rats and downregulated the expression of Snca, Mtor, Akt1, and Gsk3b in rats. These findings suggest that baicalin may inhibit the apoptosis of substantia nigra neurons in PD rats by suppressing the mTOR/Akt1/GSK-3β pathway [48]. Huang et al. revealed that the concentrations of some SCFAs, especially propionate, were significantly downregulated in the stool samples of patients with PD. Propionate administration significantly improved intestinal epithelial barrier function and motor behavior in the MPTP-induced PD mouse model through the Akt1 signaling pathway [81]. Additionally, baicalin mitigated 6-OHDA-induced cell damage and exerted therapeutic effects on PD by downregulating miR-192-5p and suppressing the PI3K/Akt1 and Mdm2/Tp53 signaling pathways [82] (Figure 3).
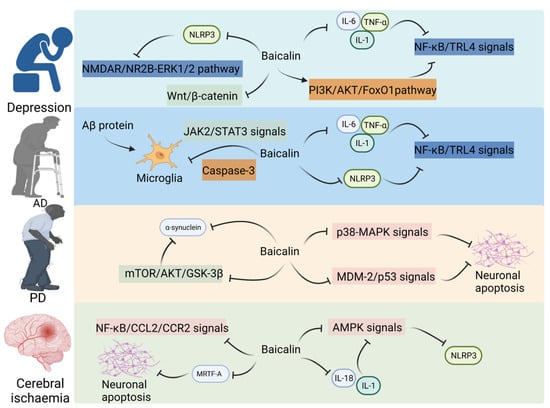
Figure 3.
Baicalin on CNS disorders in the gut–brain axis.
2.2. Therapeutic Effects of Baicalin on Metabolic Disorders
The intestinal microbiome plays an important role in the metabolism of undigested nutrients, fiber, and bile acid in the diet entering the large intestine [83,84]. The intestinal microbiota metabolizes these nutrients to release metabolites, such as SCFA, indole, and secondary bile acids. These metabolites interact with intestinal cells, promoting hormone secretion and regulating the balance of appetite homeostasis in the brain. Intestinal inflammation affects cells in the gut and gut microbiota, promoting the production and release of metabolites that act on the CNS. The disruption of gut microbiota can promote inflammatory reactions and exacerbate human metabolism, leading to metabolic disorders, such as non-alcoholic fatty liver disease and hyperlipidemia [28,85]. These results suggest that the dysregulation of the gut–brain axis promotes the development of diabetes, obesity, and other related metabolic disorders.
Hormones are involved in the regulation of food intake homeostasis. For example, gut hormones that promote appetite and anorexia, leptin signaling in adipose tissue and dopamine reward systems, and prefrontal suppression are closely related. These peripheral signals partially act through specific hypothalamic nuclei. Gut microbiota regulates the release of endocrine hormones in the gut through metabolism, altering the balance of appetite signals in the hypothalamus [86]. In the gastrointestinal tract, vagus nerve sensory neurons regulate gastric volume and intestinal contents. LGLE+ vagus nerve, especially the Glp1r+-type in the gastric myenteric layer, may be the first to transmit the gastric dilatation signal after eating to the brain stem [87]. Next, the ingested nutrients activate the intestinal endocrine cells, which respond to nutrients by releasing a large number of intestinal hormones, including glucagon-like peptide-1 (GLP-1), cholecystokinin (CCK), glucose-dependent insulin-stimulating peptide (GIP), and incretin, which can act on the CNS through the vagus nerve [88]. Additionally, these hormones regulate glucose metabolism and fat metabolism by acting on metabolism-related cells. For example, glucose-dependent insulinotropic peptides can act on pancreatic beta cells and adipocytes, promote adipocyte fat storage, and directly induce satiety through the hypothalamus [89]. GLP-1 can directly act on islet beta cells to accelerate insulin secretion and reduce glucagon secretion, lowering blood sugar [90].
Obesity has traditionally been defined as excess body fat leading to impaired health and is usually assessed in clinical practice based on body mass index [91]. The biology of obesity and energy balance has been extensively studied. However, the current understanding has not markedly delayed the obesity epidemic. Obesity, which is a global health burden, is a modifiable risk factor for cardiovascular disease and is not adequately addressed by lifestyle changes or pharmacotherapy when compared with hypertension, dyslipidemia, diabetes, and smoking [92]. In this section, the correlation between the therapeutic role of baicalin in metabolic disorders and the brain–gut axis has been examined (Figure 3).
Ju et al. demonstrated that baicalin effectively regulated the blood glucose metabolism of high-fat diet (HFD)-fed mice, including the downregulation of fasting blood glucose and the upregulation of postprandial blood glucose metabolism, suppressing fat accumulation and liver injury caused by long-term HFD consumption and alleviating dysregulated blood lipid metabolism. Baicalin significantly mitigated the HFD-induced downregulation of Ackermann bacteria, cocci, and rumen cocci abundances and the upregulation of odorous bacteria and Parabacillus abundances. The abundance of these bacteria in baicalin-treated HFD mice was similar to those in healthy control mice [58]. Ackermann bacteria, Ruminococcus, and Pseudomonas mainly produce acetic acid. Staphylococcus mainly produces propionic acid and butyric acid [93]. These SCFAs promote the metabolism, growth, and differentiation of intestinal cells, protect the intestinal mucosa, and alleviate local inflammatory reactions [94]. SCFAs interact with intestinal cells to produce GLP-1, CCK, GIP, and incretin. These hormones may act on the CNS through the vagus nerve and interact with specific receptors in the brain to modulate food intake and appetite. These results indicate that baicalin may modulate metabolic disorders by regulating the CNS through the modulation of intestinal microbiota. Lin et al. reported that 8 weeks of treatment with baicalin (400 mg/kg bodyweight) alleviated liver fat accumulation, dysregulated blood lipids, weight loss, and insulin sensitivity in the HFD-induced metabolic syndrome (MetS) mouse model through the regulation of citric acid cycle (tricarboxylic acid (TCA) cycle), alanine, aspartic acid, and glutamic acid metabolism, and glycerol phospholipid metabolism. Baicalin changed the composition and function of intestinal microbiota in MetS mice by regulating the TCA cycle and aminoacyl tRNA biosynthesis. In particular, baicalin exerts protective effects on the intestine by regulating the TCA cycle. Baicalin prevented the decline of the relative abundance of Bacteroides and the proportion of Bacteroides and Firmicutes in HFD-fed mice. Additionally, the change in succinic acid, an important product of the TCA cycle, was negatively correlated with the abundances of Bacteroides and Sutterella and positively correlated with the abundances of Myxosporidium. Treatment with succinic acid for 8 weeks significantly decreased the bodyweight and low-density lipoprotein-cholesterol levels of MetS mice. Hematoxylin and eosin revealed that baicalin downregulated fat accumulation. These results indicate that succinic acid exerts protective effects in MetS mice. Treatment with antibiotics to destroy the intestinal flora suppresses the protective effects of baicalin, indicating that baicalin exerts protective effects through the intestinal flora [49]. Although these two studies did not examine the CNS of mice, the effect of baicalin on the intestinal tract was apparent. The effects of many metabolites generated from the intestinal tract on the CNS are clear. Therefore, we predicted that baicalin regulates intestinal flora, modulates the CNS, and consequently exerts therapeutic effects on metabolic disorders.
2.3. Baicalin Alleviates Intestinal Disorders
Disturbances in the gut are correlated with metabolic disorders. Several studies have reported the correlation between IBD, neurodegenerative, and neuroinflammatory diseases [95,96]. Recent population-based studies suggest that patients with IBD are at a high risk of developing PD [97]. The vagus nerve exerts anti-inflammatory effects by inhibiting the cholinergic efferent fibers that release inflammatory cytokines from macrophages [98]. Thus, vagal nerve stimulation downregulates intestinal immune cells and clinical markers of inflammation in IBD [99]. In IBS, vagus nerve stimulation can alleviate constipation and abdominal pain and downregulate circulating IL-6 and TNF-α levels [100].
2.3.1. Baicalin Alleviates IBD through the Gut–Brain Axis
IBDs, a global healthcare problem with increasing incidence [101], including Crohn’s disease and ulcerative colitis (UC), are characterized by chronic relapse of intestinal inflammation [102]. Previous studies have suggested that IBD is caused by aberrant persistent immune responses against gut microbes that can be attributed to the genetic predisposition of individuals [103]. Although the etiology of IBD is largely unknown, it involves a complex interplay between genetic, environmental, or microbial factors and immune responses [104]. This section mainly discusses the correlation between the therapeutic effect of baicalin on UC and the brain–gut axis.
Zhu et al. demonstrated that baicalin (50 μM) suppressed LPS-induced M1 macrophage polarization and upregulation of TNF- α, IL-23, and interferon regulatory factor 5 (IRF5) in vitro, as well as upregulated the expression of IL10, ARG1, and IRF4. These factors alleviate intestinal inflammation. In vivo experiments revealed that baicalin (100 mg/kg bodyweight) inhibited the expression of IRF5 protein, promoted the expression of IRF4 protein, induced M2-type polarization of macrophages, and improved the disease activity index of dextran sodium sulfate (DSS)-induced colitis in mice [50,105]. The microglial expression of IRF5 and IRF4 was related to pro-inflammatory and anti-inflammatory responses, respectively, which contributed to the progression of poststroke inflammation and led to secondary neuronal damage. Additionally, the IRF5–IRF4 regulatory axis was suggested to be a crucial factor in microglial activation and a potential therapeutic target for neuroinflammation and ischemic stroke [106]. Baicalin (60 and 90 mg/kg bodyweight) can significantly increase the activities of SOD, CAT, and GPx in the colon tissue of UC rats and significantly downregulate the levels of malonaldehyde (MDA), IL-1β, myeloperoxidase, cleaved Casp3, cleaved Casp9, Bcl2/Bax, Cycs, and PEG2in the colon tissue of UC rats. These results suggest that baicalin exerts therapeutic effects on UC by inhibiting the IKK/IKB/NF-κB signaling pathway and apoptosis-related proteins [51]. However, Magdy et al. indicated that the NF-κB pathway alleviated glutamate accumulation and neuronal cell damage by regulating the expression of membrane proteins (connexins (Cxs), Panx1, and excitatory amino-acid transporters) in the brain tissue [107]. Baicalin (25, 50, and 100 mg/kg bodyweight) protected against trinitrobenzenesulfonic acid (TNBS)-induced UC in rats by inhibiting the upregulation of ROS and MDA in the colon tissue of rats, downregulation of the GSH and SOD levels, and modulation of Th17/T regulatory (Treg) cell balance, gut microbiota, and SCFAs [52]. These results suggest that baicalin alleviates UC-related inflammation and intestinal dysbiosis. Baicalin (316 µg/mL) significantly inhibited the release of TNF-α, IL-6, and IL-1β, promoted the expression of IL-10, and significantly suppressed LPS-induced apoptosis by blocking the PI3K/AKT signaling pathway [53]. Additionally, baicalin (50, 100, and 150 mg/kg bodyweight) downregulated the release of TNF-α, IL-1β, and IL-6 in the serum of DSS-induced chronic UC mice and suppressed IL-33 and NF-κB [54]. Similarly, treatment with baicalin (10 mg/kg bodyweight) significantly downregulated the levels of inflammatory mediators, including MPO activity, TNF-α, IL-1β, and Th1-related cytokines (IL-12 and IFN- c), decreased the number of Th17 cells, and suppressed the levels of Th17-related cytokines (IL-17 and IL-6) [55]. Similarly, baicalin can improve the inflammatory symptoms of induced colitis, downregulate the levels of pro-inflammatory mediators (TNF-α and IL-1β) in the colonic mucosa, and suppress the expression of Rela protein [56]. Additionally, baicalin can simultaneously downregulate the expression of migration inhibitory factors, including CCL2 and CCL20, and the number of macrophages in UC rats [57].
2.3.2. Baicalin Modulates Gut Microbes
Recent studies have demonstrated that gut microbiota, including bacteria, fungi, protozoa, and viruses, mediate gut–brain signaling. This has led to the emergence of the concept of a microbe–gut–brain axis [108]. The gut microbiota comprises trillions of microbes from thousands of species with 100 times more unique microbial genes than human genes. These microbes, which produce various metabolites that regulate other bacteria, are localized in the intestinal epithelium, bind to protein targets, and modulate the host functions through the intestinal barrier [109,110].
In the HFD-induced MetS mouse model, baicalin (200 mg/kg bodyweight/day) upregulated the number of SCFA-producing bacteria in the intestine, enhanced the metabolism of the intestinal cells, and alleviated MetS in mice. Additionally, hypertension can cause intestinal barrier damage [58]. For example, previous studies have investigated the therapeutic effect of baicalin (100 mg/kg bodyweight) on intestinal barrier injury in spontaneously hypertensive rats. The experimental outcomes indicated that baicalin administration increased the number of SCFA-producing bacteria and changed the intestinal microecosystem, leading to the alleviation of hypertension-induced intestinal barrier damage [59]. Zhu et al. demonstrated that baicalin (25–100 mg/kg bodyweight) exerted effective therapeutic effects on TNBS-induced UC by upregulating the number of SCFA-producing bacteria, especially butyrate-producing bacteria, such as Butyricimonas spp., Roseburia spp., Subdoligranulum spp., and Eubacterium spp. [52].
3. Discussion and Outlook
The discovery of the gut–brain axis has provided a new avenue to understand the pathogenesis of diseases. The disorders of the gut–brain axis can be mainly divided into CNS, metabolic, and intestinal disorders. Various studies have demonstrated that baicalin regulates the gut–brain axis. This narrative review provides a new perspective for understanding the therapeutic potential of baicalin in the gut–brain axis by comprehensively discussing the therapeutic role of baicalin in the disorders of the gut–brain axis.
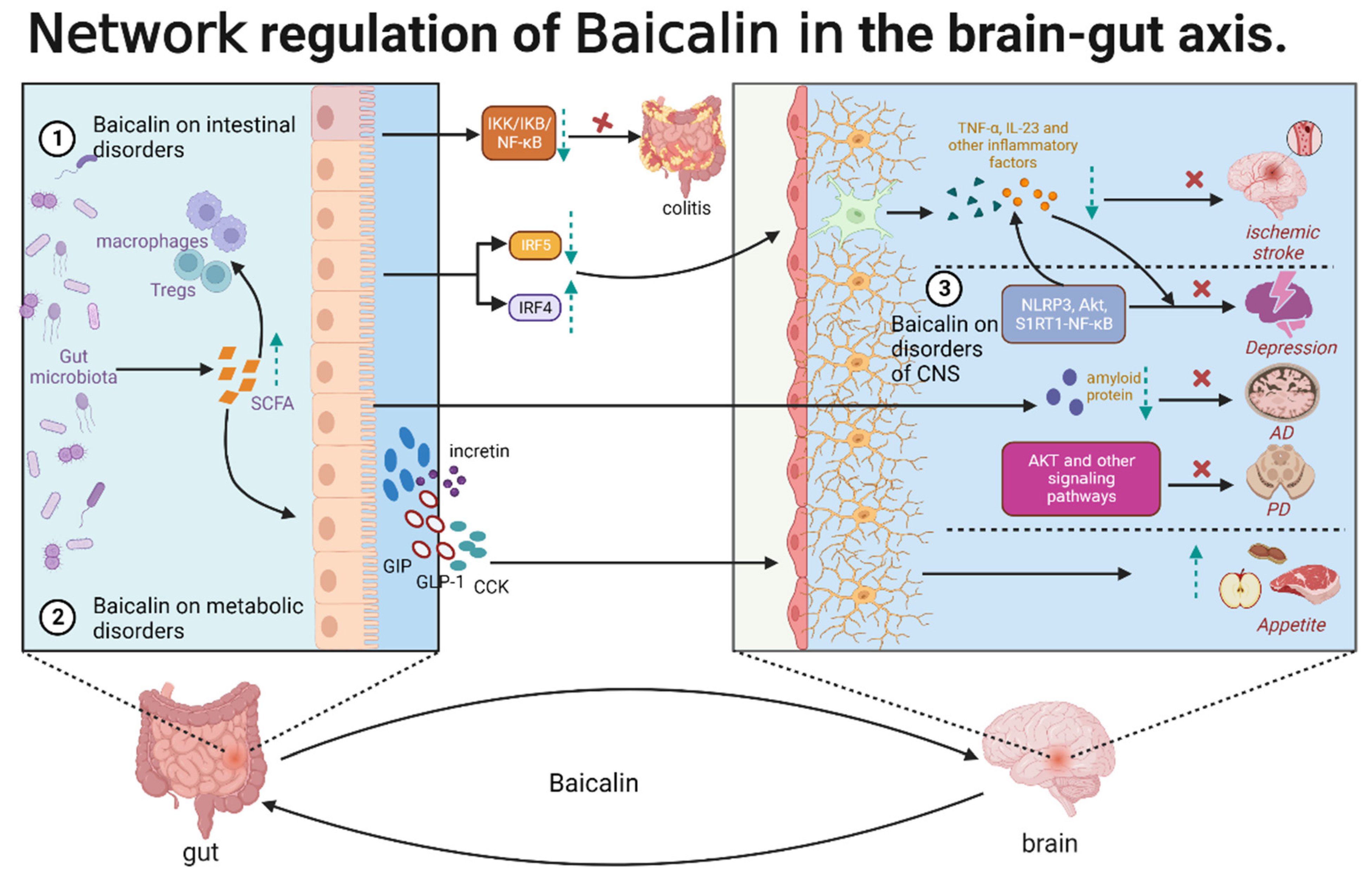
Baicalin exerts therapeutic effects on the gut–brain axis through the regulation of CNS, metabolic, and intestinal disorders. Additionally, baicalin exhibits multi-dimensional activities at the systemic, cellular, and molecular levels, forming a complex network relationship, which is the basis for the therapeutic effects of baicalin on various cerebral and intestinal diseases related to the gut–brain axis. In particular, baicalin indirectly regulates the components of refractory diseases that are inaccessible due to some physiological structures, such as BBB and intestinal wall barrier. In CNS disorders, baicalin exerts anti-neuroinflammatory and anti-neuronal apoptotic effects. Baicalin inhibits microglia activation by regulating the NLRP3, AKT1, and S1RT1-NF-κB signaling pathways and consequently alleviates neuroinflammation and depression. Additionally, baicalin can alleviate AD by regulating intestinal microbiota and decreasing the deposition of amyloid protein. Furthermore, baicalin inhibited neuronal apoptosis through AKT and other signaling pathways and alleviated PD and cerebral ischemia. In metabolic disorders, baicalin can balance intestinal flora composition, increase the production of SCFAs, and interact with intestinal cells to induce the production of GLP-1, CCK, GIP, and incretin. These hormones may act on the CNS through the vagus nerve to alter food intake and appetite. In intestinal diseases, the therapeutic effects of baicalin are multifaceted. Baicalin induces the gut microbiota to release SCFAs, which are involved in inflammation regulation by interacting with intestinal immune cells, such as macrophages and dendritic cells. Additionally, baicalin directly participates in intestinal inflammatory responses by inhibiting the IKK/IKB/NF-κB signaling pathway. Furthermore, baicalin may inhibit the activation of microglia through the IRF5–IRF4 regulatory axis, alleviating neuroinflammation and cerebral ischemia. These findings indicate that baicalin is involved in the regulation of the CNS, gut, and metabolism network through the gut–brain axis. This regulation may be mediated by gut microbes, glial cells, or gut immune cells (Figure 4). Intestinal flora can remove the glycoside on the 7-hydroxyl group of baicalin, resulting in the formation of baicalein. Thus, baicalin can exert therapeutic effects on intestinal diseases through its metabolites. Recent studies suggested a link between aberrant gut microbiota and post-traumatic stress disorder (PTSD) and indicated baicalin is a potential therapeutic target for PTSD [111]. Baicalein, a well-known precursor compound of baicalin, alleviated PTSD-induced anxiety and pain by modulating the serotonergic system and spinal δ-opioid receptors [112]. However, low bioavailability limits the clinical application of baicalin. New dosage forms or structural modifications, such as baicalin methyl ester and baicalin-7-methyl ether, must be developed to improve the pharmacokinetic parameters of baicalin and promote its clinical application.
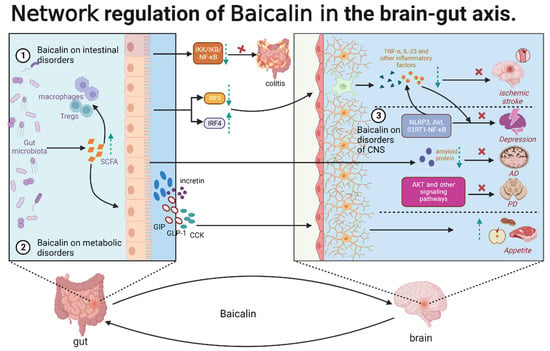
Figure 4.
The role of baicalin in the gut–brain axis.
Most studies included in this review focused on the therapeutic effect of baicalin on specific disease models. However, the therapeutic effects of baicalin have not been evaluated using related cellular studies. Microglia, astrocytes, macrophages, and dendritic cells play important regulatory roles in gut–brain axis disorders. Additionally, gut–brain axis disorders are closely related to other disorders, such as substance use disorders [113]. The gut microbiota and its secreted components, such as SCFAs, can further regulate CNS disorders. Therefore, the role of baicalin in the gut–brain axis may be a breakthrough finding.
Although several experimental studies have examined the pharmacological effects of baicalin on the gut–brain axis, limited numbers of clinical studies on baicalin have been performed. Thus, the clinical application of baicalin for the treatment of diseases may be delayed. This review has several limitations. For example, the therapeutic efficacy of baicalin may vary in different disease models. Additionally, key factors, such as clinical dose, dosage form, and mode of administration, varied between the studies. Therefore, baicalin must be comprehensively analyzed in different disease models before application in clinical trials to determine the most suitable model.
4. Conclusions
This review comprehensively discussed the therapeutic potential of baicalin in the gut–brain axis and elucidated its roles in CNS, metabolic, and intestinal disorders. Further studies are required to achieve clinical translation of baicalin. However, available evidence indicates the therapeutic value of baicalin in the disorders of the gut–brain axis.
Author Contributions
Q.H.: Conceptualization, Writing—Original Draft, Data Curation, Writing—Review and Editing; S.H.: Conceptualization, Writing—Original Draft; B.X.: Writing—Original Draft; Y.W.: Data Curation, Funding acquisition; J.W.: Data Curation, Funding acquisition; J.Z.: Writing—Review and Editing, Supervision, Funding acquisition; X.M.: Writing—Review and Editing, Supervision, Funding acquisition; F.W.: Writing—Review and Editing, Supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Science and Technology Project of Sichuan Province (grant nos. 2022YFS0399, 2023NSFSC0039, and 2023YFQ0016). “Hundred Talents Program” of the Hospital of the Chengdu University of Traditional Chinese Medicine (Grant No. 22-B09), Xinglin Scholar Research Promotion Project of Chengdu University of TCM (Grant nos. QJRC2022028 and QJJJ2022010).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available in the manuscript, and they are shown in the figures and tables.
Acknowledgments
The authors would like to thank the reviewers and authors of all references. The reviewer’s advice really made a great improvement to this paper. In addition, the figures were created by Biorender.com access on 6 June 2023, which the authors would like to thank.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Sample Availability
Not applicable.
Abbreviations
EMT: epithelial–mesenchymal transition; FXR: farnesoid X receptors; CNS: central nervous system; ENS: enteric nervous system; HPA: hypothalamic–pituitary–adrenal; IBS: irritable bowel syndrome; SCFAs: short-chain aliphatic hydrocarbons; BBB: blood–brain barrier; AD: Alzheimer’s disease; TCM: traditional Chinese medicine; LPS: lipopolysaccharide; PD: Parkinson’s disease; GLP-1: glucagon-like peptide-1; CCK: cholecystokinin; GIP: glucose-dependent insulinotropic peptide; IBD: inflammatory bowel disease; IBS: irritable bowel syndrome; IL-1β: interleukin-1β; IL-6: interleukin-6; TNF-α: tumor necrosis factor-α; PI3K: phosphatidylinositol 3-kinase; AKT: protein kinase B; FoxO1: factor forkhead box protein O 1; NAFLD: non-alcoholic fatty liver disease; NASH: non-alcoholic steatohepatitis; CUMS: chronic unpredictable mild stress; CMS: chronic mild stress; NLRP3: nucleotide-binding domain; NSE: neuron-specific enolase; BDNF: brain-derived neurotrophic factor; CEE: chronic ethanol exposure; DCX+: doublecortin; SIRT1: Sirtuin 1; GSK3β: glycogen synthase kinase-3; CREB: element-binding protein; MCAO: middle cerebral artery occlusion; TMA: trimethylamine; TMAO: trimethylamine nitroxide; NeuN: neuronal nuclei; GFAP: glial fibrillary acidic protein; ACTH: adrenocorticotropic hormone; MRTF-A: myocardin-related transcription factor-A; PS1: precursor protein/presenilin-1; Aβ: accumulation and deposition of beta-amyloid peptide; Aβ42: β-amyloid peptide; α-syn: α-synuclein; ROS: reactive oxygen species; BMI: body mass index; BAT: brown adipose tissue; WAT: white adipose tissue; UCP1: coupling protein 1; iBAT: interscapular brown adipose tissue; ATM: adipose tissue macrophages; IBD: inflammatory bowel disease; UC: ulcerative colitis; IRF5: interferon regulatory factor 5; LPS: lipopolysaccharide; MIF: macrophage migration inhibitory factor; BASO: basophil granulocyte; LYMPH: leukomonocyte; NF-κB: nuclear factor kappa-B; IBS-D: diarrhea-predominant irritable bowel syndrome; VIP: vasoactive intestinal polypeptide; CHAT: choline acety transferase; PTSD: post-traumatic stress disorder.
References
- Malagelada, J.R. The Brain-Gut Team. Dig. Dis. 2020, 38, 293–298. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaź, P. The role of microbiota-gut-brain axis in neuropsychiatric and neurological disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef]
- Pusceddu, M.M.; Del Bas, J.M. The role of the gut microbiota in the pathophysiology of mental and neurological disorders. Psychiatr. Genet. 2020, 30, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Bonaz, B.; Bazin, T.; Pellissier, S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front. Neurosci. 2018, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Breit, S.; Kupferberg, A.; Rogler, G.; Hasler, G. Vagus Nerve as Modulator of the Brain–Gut Axis in Psychiatric and Inflammatory Disorders. Front. Psychiatry 2018, 9, 44. [Google Scholar] [CrossRef]
- Chalazonitis, A.; Rao, M. Enteric nervous system manifestations of neurodegenerative disease. Brain Res. 2018, 1693, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Heiss, C.N.; Olofsson, L.E. The role of the gut microbiota in development, function and disorders of the central nervous system and the enteric nervous system. J. Neuroendocr. 2019, 31, e12684. [Google Scholar] [CrossRef]
- Agirman, G.; Yu, K.B.; Hsiao, E.Y. Signaling inflammation across the gut-brain axis. Science 2021, 374, 1087–1092. [Google Scholar] [CrossRef]
- Zhai, Z.; Su, P.-W.; Ma, L.-Y.; Yang, H.; Wang, T.; Fei, Z.-G.; Zhang, Y.-N.; Wang, Y.; Ma, K.; Han, B.-B.; et al. Progress on traditional Chinese medicine in treatment of ischemic stroke via the gut-brain axis. BioMedicine 2023, 157, 114056. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qiu, W.; Da, X.; Hou, Y.; Ma, Q.; Wang, T.; Zhou, X.; Song, M.; Bian, Q.; Chen, J. A combination of depression and liver Qi stagnation and spleen deficiency syndrome using a rat model. Anat. Rec. 2020, 303, 2154–2167. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Yang, J.; Cao, B.; Xue, Y.; Gao, P.; Liang, H.; Li, G. Growth years and post-harvest processing methods have critical roles on the contents of medicinal active ingredients of Scutellaria baicalensis. Ind. Crops Prod. 2020, 158, 112985. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.L.; Wang, S.; Kuang, Y.; Hu, Z.M.; Qiao, X.; Ye, M. A comprehensive review on phytochemistry, pharmacology, and flavonoid biosynthesis of Scutellaria baicalensis. Pharm. Biol. 2018, 56, 465–484. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi. (Lamiaceae): A review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J. Pharm. Pharmacol. 2019, 71, 1353–1369. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Li, P.; Liu, S.; Liu, Q.; Li, Y.; Sun, Y.; He, C.; Xiao, P. Traditional uses, ten-years research progress on phytochemistry and pharmacology, and clinical studies of the genus Scutellaria. J. Ethnopharmacol. 2021, 265, 113198. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Guan, Y.; Hu, W.; Xu, Z.; Ishfaq, M. An overview of pharmacological activities of baicalin and its aglycone baicalein: New insights into molecular mechanisms and signaling pathways. Iran. J. Basic. Med. Sci. 2022, 25, 14–26. [Google Scholar] [PubMed]
- Huang, T.; Liu, Y.; Zhang, C. Pharmacokinetics and Bioavailability Enhancement of Baicalin: A Review. Eur. J. Drug Metab. Pharmacokinet. 2018, 44, 159–168. [Google Scholar] [CrossRef]
- Chung, H.; Choi, H.S.; Seo, E.-K.; Kang, D.-H.; Oh, E.-S. Baicalin and baicalein inhibit transforming growth factor-β1-mediated epithelial-mesenchymal transition in human breast epithelial cells. Biochem. Biophys. Res. Commun. 2015, 458, 707–713. [Google Scholar] [CrossRef]
- Hu, Q.; Zhang, W.; Wu, Z.; Tian, X.; Xiang, J.; Li, L.; Li, Z.; Peng, X.; Wei, S.; Ma, X.; et al. Baicalin and the liver-gut system: Pharmacological bases explaining its therapeutic effects. Pharmacol. Res. 2021, 165, 105444. [Google Scholar] [CrossRef]
- Pan, Y.; Chen, X.Y.; Zhang, Q.Y.; Kong, L.D. Corrigendum to “Microglial NLRP3 inflammasome activation mediates IL-1beta-related inflammation in prefrontal cortex of depressive rats” [Brain Behav. Immun. 41 (2014) 90-100]. Brain Behav. Immun. 2021, 97, 455. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Zhang, J.; Li, X.; Liu, Y.; Wang, T.; Yan, Z.; Chen, J. Effects of Xiaoyaosan on Depressive-Like Behaviors in Rats with Chronic Unpredictable Mild Stress Through HPA Axis Induced Astrocytic Activities. Front. Psychiatry 2020, 11, 545823. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Sun, G.; Yang, F.; Guan, Z.; Zhang, Z.; Zhao, J.; Liu, Y.; Chu, L.; Pei, L. Baicalin regulates depression behavior in mice exposed to chronic mild stress via the Rac/LIMK/cofilin pathway. Biomed. Pharmacother. 2019, 116, 109054. [Google Scholar] [CrossRef]
- Hammen, C. Risk Factors for Depression: An Autobiographical Review. Annu. Rev. Clin. Psychol. 2018, 14, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Tolentino, J.C.; Schmidt, S.L. DSM-5 Criteria and Depression Severity: Implications for Clinical Practice. Front. Psychiatry 2018, 9, 450. [Google Scholar] [CrossRef]
- Pan, Z.; Park, C.; Brietzke, E.; Zuckerman, H.; Rong, C.; Mansur, R.B.; Fus, D.; Subramaniapillai, M.; Lee, Y.; McIntyre, R.S. Cognitive impairment in major depressive disorder. CNS Spectr. 2019, 24, 22–29. [Google Scholar] [CrossRef]
- Marwaha, S.; Palmer, E.; Suppes, T.; Cons, E.; Young, A.H.; Upthegrove, R. Novel and emerging treatments for major depression. Lancet 2023, 401, 141–153. [Google Scholar] [CrossRef]
- Jia, X.; Gao, Z.; Hu, H. Microglia in depression: Current perspectives. Sci. China Life Sci. 2021, 64, 911–925. [Google Scholar] [CrossRef]
- Carlessi, A.S.; Borba, L.A.; Zugno, A.I.; Quevedo, J.; Réus, G.Z. Gut microbiota–brain axis in depression: The role of neuroinflammation. Eur. J. Neurosci. 2021, 53, 222–235. [Google Scholar] [CrossRef]
- Guo, L.-T.; Wang, S.-Q.; Su, J.; Xu, L.-X.; Ji, Z.-Y.; Zhang, R.-Y.; Zhao, Q.-W.; Ma, Z.-Q.; Deng, X.-Y.; Ma, S.-P. Baicalin ameliorates neuroinflammation-induced depressive-like behavior through inhibition of toll-like receptor 4 expression via the PI3K/AKT/FoxO1 pathway. J. Neuroinflamm. 2019, 16, 95. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Zeng, M.J.; Zhou, L.P.; Li, Y.Q.; Zhao, F.; Shang, Z.Y.; Deng, X.Y.; Ma, Z.Q.; Fu, Q.; Ma, S.P.; et al. Baicalin exerts neuroprotective effects via inhibiting activation of GSK3β/NF-κB/NLRP3 signal pathway in a rat model of depression. Int. Immunopharmacol. 2018, 64, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Zhang, D.; Yu, H.; Yuan, H.; Shen, H.; Lan, X.; Liu, H.; Chen, X.; Meng, F.; Wu, X.; et al. Gut microbiota regulates chronic ethanol exposure-induced depressive-like behavior through hippocampal NLRP3-mediated neuroinflammation. Mol. Psychiatry 2023, 28, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, C. Baicalin ameliorates chronic unpredictable mild stress-induced depressive behavior: Involving the inhibition of NLRP3 inflammasome activation in rat prefrontal cortex. Int. Immunopharmacol. 2017, 48, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ma, Z.; Liu, K.; Li, Y.; Liu, D.; Xu, L.; Deng, X.; Qu, R.; Ma, Z.; Ma, S. Baicalin exerts antidepressant effects through Akt/FOXG1 pathway promoting neuronal differentiation and survival. Life Sci. 2019, 221, 241–248. [Google Scholar] [CrossRef]
- Fang, A.; Li, Y.; Wu, X.; Wu, B.; Zhang, Y. Baicalin attenuates inflammatory pain associated depressive symptoms via Akt-mediated adult hippocampal neurogenesis. Metab. Brain Dis. 2020, 35, 1085–1093. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Duan, W.; Zhao, Z.; Yang, L.; Wei, W.; Li, J.; Li, Y.; Yu, Y.; Dai, B.; et al. Shugan granule contributes to the improvement of depression-like behaviors in chronic restraint stress-stimulated rats by altering gut microbiota. CNS Neurosci. Ther. 2022, 28, 1409–1424. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cui, H.; Li, T.; Qi, J.; Chen, H.; Gao, F.; Tian, X.; Mu, Y.; He, R.; Lv, S.; et al. Synergistic Effect of Berberine-Based Chinese Medicine Assembled Nanostructures on Diarrhea-Predominant Irritable Bowel Syndrome In Vivo. Front. Pharmacol. 2020, 11, 1210. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, F.; Guan, X. Baicalin reverse depressive-like behaviors through regulation SIRT1-NF-kB signaling pathway in olfactory bulbectomized rats. Phytother. Res. 2019, 33, 1480–1489. [Google Scholar] [CrossRef]
- Wellman, A.S.; Metukuri, M.R.; Kazgan, N.; Xu, X.; Xu, Q.; Ren, N.S.; Czopik, A.; Shanahan, M.T.; Kang, A.; Chen, W.; et al. Intestinal Epithelial Sirtuin 1 Regulates Intestinal Inflammation During Aging in Mice by Altering the Intestinal Microbiota. Gastroenterology 2017, 153, 772–786. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, T.; Wang, Y.; Si, C.; Wang, X.; Wang, R.-T.; Lv, Z. Baicalin ameliorates neuropathology in repeated cerebral ischemia-reperfusion injury model mice by remodeling the gut microbiota. Aging 2020, 12, 3791–3806. [Google Scholar] [CrossRef]
- Li, H.; Wu, L.; Wang, D.; Guo, Y.; Liu, T.; Pan, Y. A protective effect of baicalin on cerebral ischemic rats is related to the improvement of serum progesterone level in serum. NeuroReport 2019, 30, 1121–1128. [Google Scholar] [CrossRef]
- Li, S.; Sun, X.; Xu, L.; Sun, R.; Ma, Z.; Deng, X.; Liu, B.; Fu, Q.; Qu, R.; Ma, S. Baicalin attenuates in vivo and in vitro hyperglycemia-exacerbated ischemia/reperfusion injury by regulating mitochondrial function in a manner dependent on AMPK. Eur. J. Pharmacol. 2017, 815, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Liu, M.Y.; Zhang, D.F.; Zhong, X.; Du, K.; Qian, P.; Yao, W.F.; Gao, H.; Wei, M.J. Baicalin mitigates cognitive impairment and protects neurons from microglia-mediated neuroinflammation via suppressing NLRP3 inflammasomes and TLR4/NF-κB signaling pathway. CNS Neurosci. Ther. 2019, 25, 575–590. [Google Scholar] [CrossRef]
- Chen, C.; Li, X.; Gao, P.; Tu, Y.; Zhao, M.; Li, J.; Zhang, S.; Liang, H. Baicalin attenuates Alzheimer-like pathological changes and memory deficits induced by amyloid β1–42 protein. Metab. Brain Dis. 2015, 30, 537–544. [Google Scholar] [CrossRef]
- Ding, H.; Wang, H.; Zhao, Y.; Sun, D.; Zhai, X. Protective Effects of Baicalin on Aβ1–42-Induced Learning and Memory Deficit, Oxidative Stress, and Apoptosis in Rat. Cell Mol. Neurobiol. 2015, 35, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Wang, C.; Chen, H.; Hu, Y.; Tian, L.; Pan, J.; Geng, M. Aβ-induced microglial cell activation is inhibited by baicalin through the JAK2/STAT3 signaling pathway. Int. J. Neurosci. 2014, 124, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Lei, K.; Shen, Y.; He, Y.; Zhang, L.; Zhang, J.; Tong, W.; Xu, Y.; Jin, L. Baicalin Represses C/EBPβ via Its Antioxidative Effect in Parkinson’s Disease. Oxidative Med. Cell. Longev. 2020, 2020, 8951907. [Google Scholar] [CrossRef]
- Zhai, H.; Kang, Z.; Zhang, H.; Ma, J.; Chen, G. Baicalin attenuated substantia nigra neuronal apoptosis in Parkinson’s disease rats via the mTOR/AKT/GSK-3β pathway. J. Integr. Neurosci. 2019, 18, 423–429. [Google Scholar] [PubMed]
- Lin, Y.; Wang, Z.-Y.; Wang, M.-J.; Jiang, Z.-M.; Qin, Y.-Q.; Huang, T.-Q.; Song, Y.; Liang, H.-T.; Liu, E.-H. Baicalin attenuate diet-induced metabolic syndrome by improving abnormal metabolism and gut microbiota. Eur. J. Pharmacol. 2022, 925, 174996. [Google Scholar] [CrossRef]
- Zhu, W.; Jin, Z.; Yu, J.; Liang, J.; Yang, Q.; Li, F.; Shi, X.; Zhu, X.; Zhang, X. Baicalin ameliorates experimental inflammatory bowel disease through polarization of macrophages to an M2 phenotype. Int. Immunopharmacol. 2016, 35, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Cheng, J.; Zhu, S.; Zhao, J.; Ye, Q.; Xu, Y.; Dong, H.; Zheng, X. Regulating effect of baicalin on IKK/IKB/NF-kB signaling pathway and apoptosis-related proteins in rats with ulcerative colitis. Int. Immunopharmacol. 2019, 73, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Xu, L.-Z.; Zhao, S.; Shen, Z.-F.; Shen, H.; Zhan, L.-B. Protective effect of baicalin on the regulation of Treg/Th17 balance, gut microbiota and short-chain fatty acids in rats with ulcerative colitis. Appl. Microbiol. Biotechnol. 2020, 104, 5449–5460. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Shen, H.; Gu, P.; Liu, Y.; Zhang, L.; Cheng, J. Baicalin alleviates TNBS-induced colitis by inhibiting PI3K/AKT pathway activation. Exp. Ther. Med. 2020, 20, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.L.; Zhang, S.; He, W.X.; Lu, J.L.; Xu, Y.J.; Yang, J.Y.; Liu, D. Baicalin may alleviate inflammatory infiltration in dextran sodium sulfate-induced chronic ulcerative colitis via inhibiting IL-33 expression. Life Sci. 2017, 186, 125–132. [Google Scholar] [CrossRef]
- Zou, Y.; Dai, S.X.; Chi, H.G.; Li, T.; He, Z.W.; Wang, J.; Ye, C.G.; Huang, G.L.; Zhao, B.; Li, W.Y.; et al. Baicalin attenuates TNBS-induced colitis in rats by modulating the Th17/Treg paradigm. Arch. Pharm. Res. 2015, 38, 1873–1887. [Google Scholar] [CrossRef]
- Cui, L.; Feng, L.; Zhang, Z.H.; Jia, X.B. The anti-inflammation effect of baicalin on experimental colitis through inhibiting TLR4/NF-κB pathway activation. Int. Immunopharmacol. 2014, 23, 294–303. [Google Scholar] [CrossRef]
- Dai, S.-X.; Zou, Y.; Feng, Y.-L.; Liu, H.-B.; Zheng, X.-B. Baicalin Down-regulates the Expression of Macrophage Migration Inhibitory Factor (MIF) Effectively for Rats with Ulcerative Colitis. Phytother. Res. 2012, 26, 498–504. [Google Scholar] [CrossRef]
- Ju, M.; Liu, Y.; Li, M.; Cheng, M.; Zhang, Y.; Deng, G.; Kang, X.; Liu, H. Baicalin improves intestinal microecology and abnormal metabolism induced by high-fat diet. Eur. J. Pharmacol. 2019, 857, 172457. [Google Scholar] [CrossRef]
- Wu, D.; Ding, L.; Tang, X.; Wang, W.; Chen, Y.; Zhang, T. Baicalin Protects Against Hypertension-Associated Intestinal Barrier Impairment in Part Through Enhanced Microbial Production of Short-Chain Fatty Acids. Front. Pharmacol. 2019, 10, 1271. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, S.; Kou, L.; Tang, C.; Huang, R.; Pei, Z.; Li, Z. Ischemic stroke damages the intestinal mucosa and induces alteration of the intestinal lymphocytes and CCL19 mRNA in rats. Neurosci. Lett. 2017, 658, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Song, Y.; Du, X.; Bai, L.; Hua, X.; Xiao, Y.; Yu, X. Intestinal barrier dysfunction following traumatic brain injury. Neurol. Sci. 2019, 40, 1105–1110. [Google Scholar] [CrossRef]
- Chen, Y.; Liang, J.; Ouyang, F.; Chen, X.; Lu, T.; Jiang, Z.; Li, J.; Li, Y.; Zeng, J. Persistence of Gut Microbiota Dysbiosis and Chronic Systemic Inflammation After Cerebral Infarction in Cynomolgus Monkeys. Front. Neurol. 2019, 10, 661. [Google Scholar] [CrossRef]
- Paul, S.; Candelario-Jalil, E. Emerging neuroprotective strategies for the treatment of ischemic stroke: An overview of clinical and preclinical studies. Exp. Neurol. 2021, 335, 113518. [Google Scholar] [CrossRef] [PubMed]
- Farina, M.; Vieira, L.E.; Buttari, B.; Profumo, E.; Saso, L. The Nrf2 Pathway in Ischemic Stroke: A Review. Molecules 2021, 26, 5001. [Google Scholar] [CrossRef] [PubMed]
- Diener, H.C.; Hankey, G.J. Primary and Secondary Prevention of Ischemic Stroke and Cerebral Hemorrhage: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 1804–1818. [Google Scholar] [CrossRef]
- Wei, X.; Huang, G.; Liu, J.; Ge, J.; Zhang, W.; Mei, Z. An update on the role of Hippo signaling pathway in ischemia-associated central nervous system diseases. Biomed. Pharmacother. 2023, 162, 114619. [Google Scholar] [CrossRef] [PubMed]
- Chidambaram, S.B.; Rathipriya, A.G.; Mahalakshmi, A.M.; Sharma, S.; Hediyal, T.A.; Ray, B.; Sunanda, T.; Rungratanawanich, W.; Kashyap, R.S.; Qoronfleh, M.W.; et al. The Influence of Gut Dysbiosis in the Pathogenesis and Management of Ischemic Stroke. Cells 2022, 11, 1239. [Google Scholar] [CrossRef] [PubMed]
- Chan, O.; Inouye, K.; Riddell, M.C.; Vranic, M.; Matthews, S.G. Diabetes and the hypothalamo-pituitary-adrenal (HPA) axis. Minerva Endocrinol. 2003, 28, 87–102. [Google Scholar]
- Li, M.; Wang, S.; Zhang, C.; Chi, C.; Liu, R.; Wang, T.; Fu, F. Escin alleviates stress-induced intestinal dysfunction to protect brain injury by regulating the gut-brain axis in ischemic stroke rats. Int. Immunopharmacol. 2023, 115, 109659. [Google Scholar] [CrossRef]
- Kesika, P.; Suganthy, N.; Sivamaruthi, B.S.; Chaiyasut, C. Role of gut-brain axis, gut microbial composition, and probiotic intervention in Alzheimer’s disease. Life Sci. 2021, 264, 118627. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Q.; Zheng, Q.; Wang, Q.; Zeng, Q.P. Biotic/Abiotic Stress-Driven Alzheimer’s Disease. Front. Cell Neurosci. 2016, 10, 269. [Google Scholar] [CrossRef]
- Eratne, D.; Loi, S.M.; Farrand, S.; Kelso, W.; Velakoulis, D.; Looi, J.C. Alzheimer’s disease: Clinical update on epidemiology, pathophysiology and diagnosis. Australas Psychiatry 2018, 26, 347–357. [Google Scholar] [CrossRef]
- International, World Alzheimer Report 2022. Life after Diagnosis: Navigating Treatment, Care and Support. 2022. Available online: https://www.alzint.org/u/ (accessed on 6 June 2023).
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Harach, T.; Marungruang, N.; Duthilleul, N.; Cheatham, V.; Mc Coy, K.D.; Frisoni, G.B.; Neher, J.J.; Fåk, F.; Jucker, M.; Lasser, T.; et al. Reduction of Abeta amyloid pathology in APPPS1 transgenic mice in the absence of gut microbiota. Sci. Rep. 2017, 7, 41802. [Google Scholar] [CrossRef] [PubMed]
- Megur, A.; Baltriukienė, D.; Bukelskienė, V.; Burokas, A. The Microbiota–Gut–Brain Axis and Alzheimer’s Disease: Neuroinflammation Is to Blame? Nutrients 2020, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Klann, E.M.; Dissanayake, U.; Gurrala, A.; Farrer, M.; Shukla, A.W.; Ramirez-Zamora, A.; Mai, V.; Vedam-Mai, V. The Gut-Brain Axis and Its Relation to Parkinson’s Disease: A Review. Front. Aging Neurosci. 2021, 13, 782082. [Google Scholar] [CrossRef]
- Visñuk, D.P.; de Giori, G.S.; LeBlanc, J.G.; de Moreno de LeBlanc, A. Neuroprotective effects associated with immune modulation by selected lactic acid bacteria in a Parkinson’s disease model. Nutrition 2020, 79–80, 110995. [Google Scholar] [CrossRef]
- Shannon, K.M. Gut-Derived Sterile Inflammation and Parkinson’s Disease. Front. Neurol. 2022, 13, 831090. [Google Scholar] [CrossRef] [PubMed]
- Dodiya, H.B.; Forsyth, C.B.; Voigt, R.M.; Engen, P.A.; Patel, J.; Shaikh, M.; Green, S.J.; Naqib, A.; Roy, A.; Kordower, J.H.; et al. Chronic stress-induced gut dysfunction exacerbates Parkinson’s disease phenotype and pathology in a rotenone-induced mouse model of Parkinson’s disease. Neurobiol. Dis. 2020, 135, 104352. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Shi, H.; Xu, Y.; Ji, L. The gut microbiota metabolite propionate ameliorates intestinal epithelial barrier dysfunction-mediated Parkinson’s disease via the AKT signaling pathway. Neuroreport 2021, 32, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.; Wu, Z.-Y.; Yang, X.-L.; Zhang, Q.; Gu, R.; Wang, Q.; Tian, T.; Yao, H.; Qu, X.; Tian, J.-Y. Neuroprotective effect and mechanism of baicalin on Parkinson’s disease model induced by 6-OHDA. Neuropsychiatr. Dis. Treat. 2019, 15, 3615–3625. [Google Scholar] [CrossRef] [PubMed]
- Plaisancié, P.; Dumoulin, V.; Chayvialle, J.-A.; Cuber, J.-C. Luminal glucagon-like peptide-1(7–36) amide-releasing factors in the isolated vascularly perfused rat colon. J. Endocrinol. 1995, 145, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Peredo-Lovillo, A.; Romero-Luna, H.; Jiménez-Fernández, M. Health promoting microbial metabolites produced by gut microbiota after prebiotics metabolism. Food Res. Int. 2020, 136, 109473. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-S.; Hu, Y.-Y. Intestinal Microecology: An Important Target for Chinese Medicine Treatment of Non-alcoholic Fatty Liver Disease. Chin. J. Integr. Med. 2020, 26, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.-J.; Tomasi, D.; Baler, R.D. Obesity and addiction: Neurobiological overlaps. Obes. Rev. 2013, 14, 2–18. [Google Scholar] [CrossRef] [PubMed]
- EWilliams, K.; Chang, R.B.; Strochlic, D.E.; Umans, B.D.; Lowell, B.B.; Liberles, S.D. Sensory Neurons that Detect Stretch and Nutrients in the Digestive System. Cell 2016, 166, 209–221. [Google Scholar] [CrossRef]
- Bai, L.; Mesgarzadeh, S.; Ramesh, K.S.; Huey, E.L.; Liu, Y.; Gray, L.A.; Aitken, T.J.; Chen, Y.; Beutler, L.R.; Ahn, J.S.; et al. Genetic Identification of Vagal Sensory Neurons That Control Feeding. Cell 2019, 179, 1129–1143.e23. [Google Scholar] [CrossRef]
- Khan, R.; Tomas, A.; Rutter, G.A. Effects on pancreatic Beta and other Islet cells of the glucose-dependent insulinotropic polypeptide. Peptides 2020, 125, 170201. [Google Scholar] [CrossRef]
- Drucker, D.J. Mechanisms of Action and Therapeutic Application of Glucagon-like Peptide-1. Cell Metab. 2018, 27, 740–756. [Google Scholar] [CrossRef]
- LMayoral, P.; Andrade, G.M.; Mayoral, E.P.; Huerta, T.H.; Canseco, S.P.; Canales, F.J.R.; Cabrera-Fuentes, H.A.; Cruz, M.M.; Santiago, A.D.P.; Alpuche, J.J.; et al. Obesity subtypes, related biomarkers & heterogeneity. Indian J. Med. Res. 2020, 151, 11–21. [Google Scholar]
- Hu, Q.; Chen, Y.; Deng, X.; Li, Y.; Ma, X.; Zeng, J.; Zhao, Y. Diabetic Nephropathy: Focusing on Pathological Signals, Clinical Treatment, and Dietary Regulation. Biomed. Pharmacother. 2023, 159, 114252. [Google Scholar] [CrossRef] [PubMed]
- Venegas, D.P.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yu, T.; Huang, X.; Bilotta, A.J.; Xu, L.; Lu, Y.; Sun, J.; Pan, F.; Zhou, J.; Zhang, W.; et al. Intestinal microbiota-derived short-chain fatty acids regulation of immune cell IL-22 production and gut immunity. Nat. Commun. 2020, 11, 4457. [Google Scholar] [CrossRef] [PubMed]
- D’antongiovanni, V.; Pellegrini, C.; Antonioli, L.; Ippolito, C.; Segnani, C.; Benvenuti, L.; D’amati, A.; Errede, M.; Virgintino, D.; Fornai, M.; et al. Enteric Glia and Brain Astroglia: Complex Communication in Health and Disease along the Gut-Brain Axis. Neuroscientist 2023, 10738584231163460. [Google Scholar] [CrossRef] [PubMed]
- Günther, C.; Rothhammer, V.; Karow, M.; Neurath, M.F.; Winner, B. The Gut-Brain Axis in Inflammatory Bowel Disease—Current and Future Perspectives. Int. J. Mol. Sci. 2021, 22, 8870. [Google Scholar] [CrossRef] [PubMed]
- Villumsen, M.; Aznar, S.; Pakkenberg, B.; Jess, T.; Brudek, T. Inflammatory bowel disease increases the risk of Parkinson’s disease: A Danish nationwide cohort study 1977–2014. Gut 2019, 68, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, D.; Ottani, A.; Altavilla, D.; Bazzani, C.; Squadrito, F.; Guarini, S. Melanocortins and the Cholinergic Anti-Inflammatory Pathway. Adv. Exp. Med. Biol. 2010, 681, 71–87. [Google Scholar] [CrossRef]
- Meroni, E.; Stakenborg, N.; Viola, M.F.; Boeckxstaens, G.E. Intestinal macrophages and their interaction with the enteric nervous system in health and inflammatory bowel disease. Acta Physiol. 2019, 225, e13163. [Google Scholar] [CrossRef]
- O’Brien, R.; Buckley, M.M.; O’Malley, D. Divergent effects of exendin-4 and interleukin-6 on rat colonic secretory and contractile activity are associated with changes in regional vagal afferent signaling. Neurogastroenterol. Motil. 2021, 33, e14160. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef]
- Guzzo, G.L.; Andrews, J.M.; Weyrich, L.S. The Neglected Gut Microbiome: Fungi, Protozoa, and Bacteriophages in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2022, 28, 1112–1122. [Google Scholar] [CrossRef]
- Rashed, R.; Valcheva, R.; Dieleman, L.A. Manipulation of Gut Microbiota as a Key Target for Crohn’s Disease. Front. Med. 2022, 9, 887044. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Chang, E.B. Inflammatory Bowel Diseases (IBD) and the Microbiome—Searching the Crime Scene for Clues. Gastroenterology 2021, 160, 524–537. [Google Scholar] [CrossRef]
- Lai, Y.-S.; Putra, R.B.D.S.; Aui, S.-P.; Chang, K.-T. M2C Polarization by Baicalin Enhances Efferocytosis via Upregulation of MERTK Receptor. Am. J. Chin. Med. 2018, 46, 1899–1914. [Google Scholar] [CrossRef] [PubMed]
- Al Mamun, A.; Chauhan, A.; Qi, S.; Ngwa, C.; Xu, Y.; Sharmeen, R.; Hazen, A.L.; Li, J.; Aronowski, J.A.; McCullough, L.D.; et al. Microglial IRF5-IRF4 regulatory axis regulates neuroinflammation after cerebral ischemia and impacts stroke outcomes. Proc. Natl. Acad. Sci. USA 2020, 117, 1742–1752. [Google Scholar] [CrossRef] [PubMed]
- Magdy, S.; Gamal, M.; Samir, N.F.; Rashed, L.; Aboulhoda, B.E.; Mohammed, H.S.; Sharawy, N. IκB kinase inhibition remodeled connexins, pannexin-1, and excitatory amino-acid transporters expressions to promote neuroprotection of galantamine and morphine. J. Cell Physiol. 2011, 236, 7516–7532. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xia, Y.; He, F.; Zhu, C.; Ren, W. Intestinal mycobiota in health and diseases: From a disrupted equilibrium to clinical opportunities. Microbiome 2021, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, P.; Capparelli, R.; Alifano, M.; Iannelli, A.; Iannelli, D. Gut Microbiota Host-Gene Interaction. Int. J. Mol. Sci. 2022, 23, 13717. [Google Scholar] [CrossRef] [PubMed]
- Worthington, J.; Reimann, F.; Gribble, F. Enteroendocrine cells-sensory sentinels of the intestinal environment and orchestrators of mucosal immunity. Mucosal Immunol. 2018, 11, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Laudani, S.; Torrisi, S.A.; Alboni, S.; Bastiaanssen, T.F.; Benatti, C.; Rivi, V.; Moloney, R.D.; Fuochi, V.; Furneri, P.M.; Drago, F.; et al. Gut microbiota alterations promote traumatic stress susceptibility associated with p-cresol-induced dopaminergic dysfunctions. Brain, Behav. Immun. 2023, 107, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Ruan, L.; Guan, K.; Wang, Y.; Gu, M.; Chen, Y.; Cai, L.; Ye, R.; Huang, Z.; Guo, A.; Su, Z.; et al. Baicalein exerts anxiolytic and antinociceptive effects in a mouse model of posttraumatic stress disorder: Involvement of the serotonergic system and spinal delta-opioid receptors. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2023, 122, 110689. [Google Scholar] [CrossRef] [PubMed]
- Baslam, A.; Aitbaba, A.; Hanchi, A.L.; Tazart, Z.; Aboufatima, R.; Soraa, N.; Ait-El-Mokhtar, M.; Boussaa, S.; Baslam, M.; Chait, A. Modulation of Gut Microbiome in Ecstasy/MDMA-Induced Behavioral and Biochemical Impairment in Rats and Potential of Post-Treatment with Anacyclus pyrethrum L. Aqueous Extract to Mitigate Adverse Effects. Int. J. Mol. Sci. 2023, 24, 9086. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).