
Glycosylation Contributes to Thermostability and Proteolytic Resistance of rFIP-nha (Nectria haematococca)

 , , ,
, , ,
Abstract
1. Introduction
2. Results and Discussion
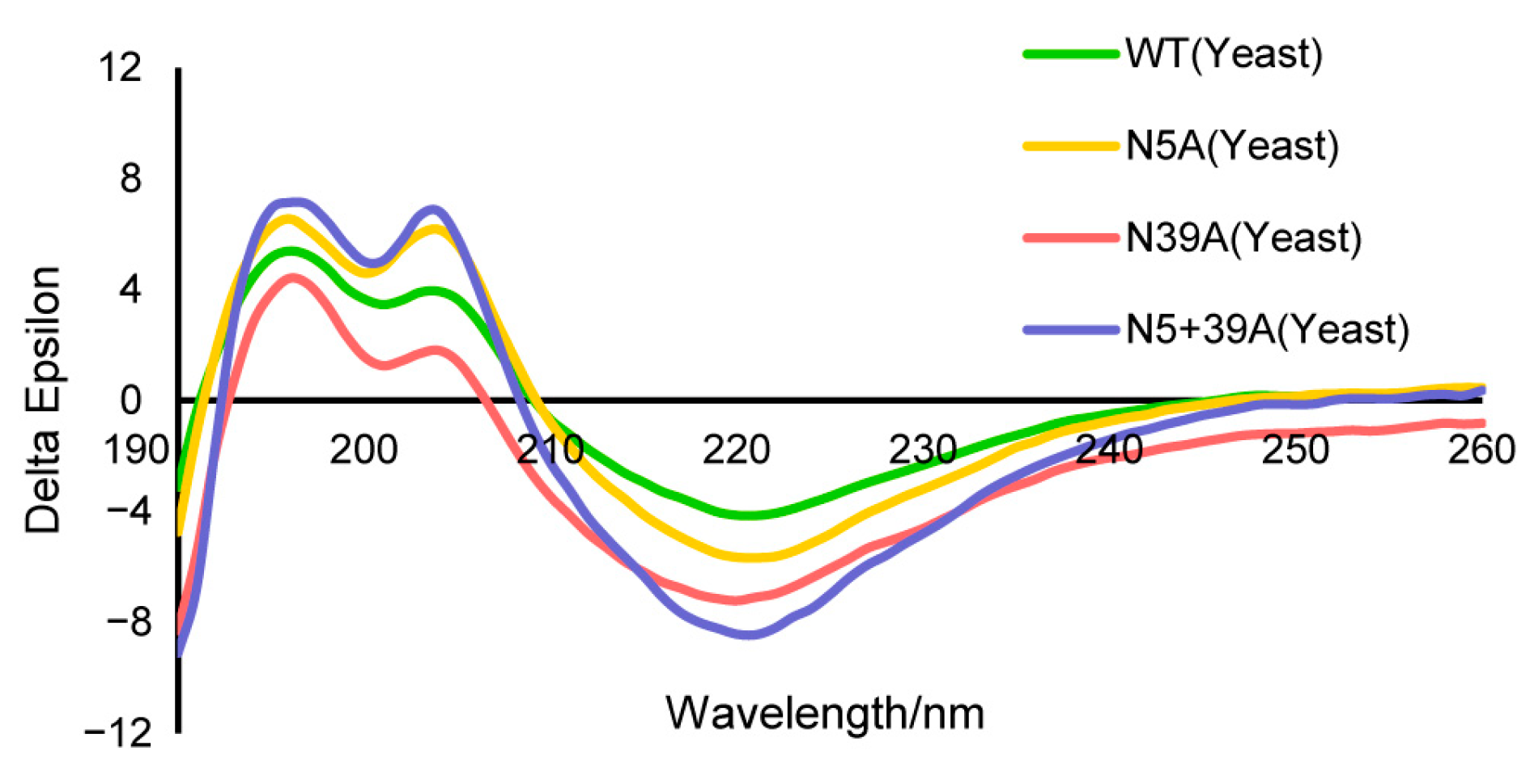
2.1. Biochemical Characteristics of Glycosylation Mutants of FIP-nha
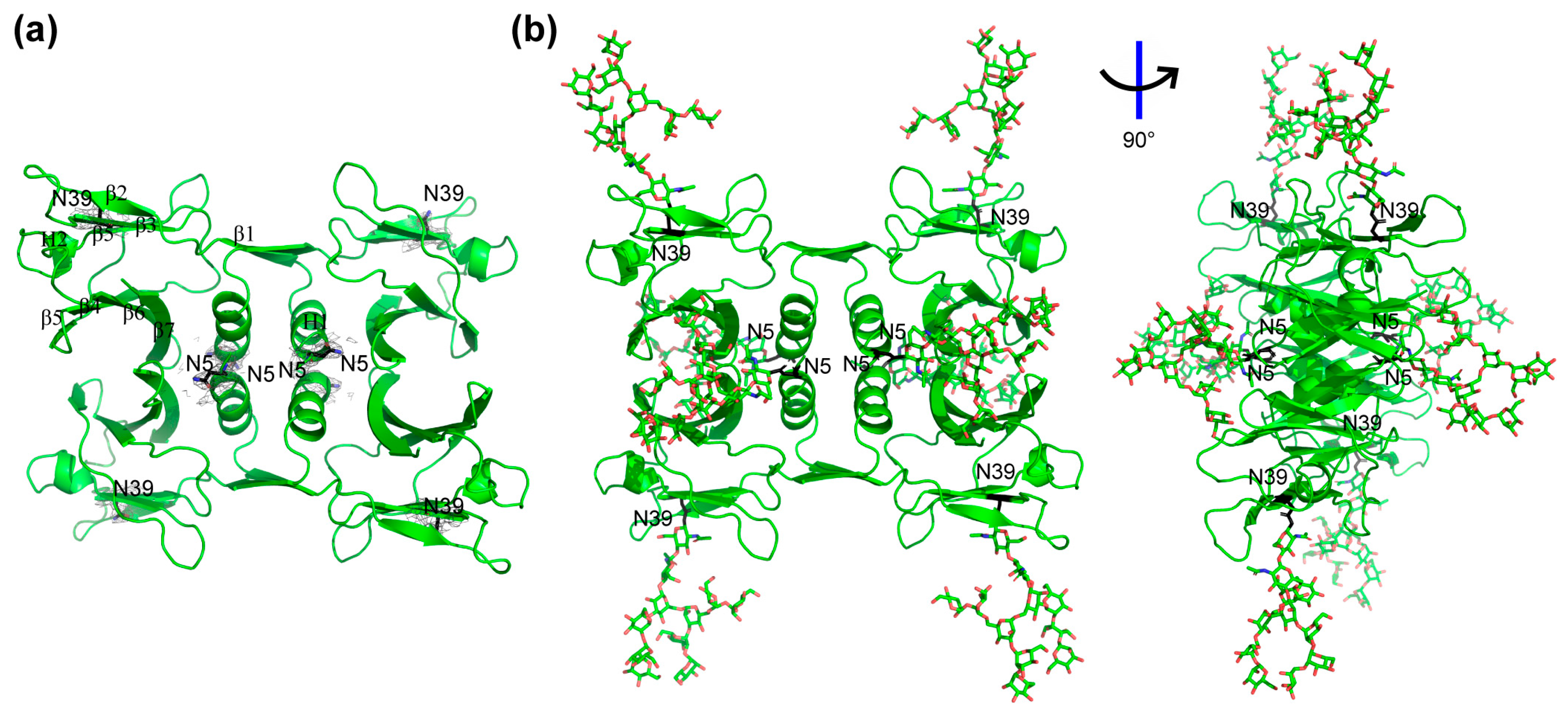
2.2. Crystal Structure Analysis
2.3. Thermostability of rFIP-nha and Its Variants
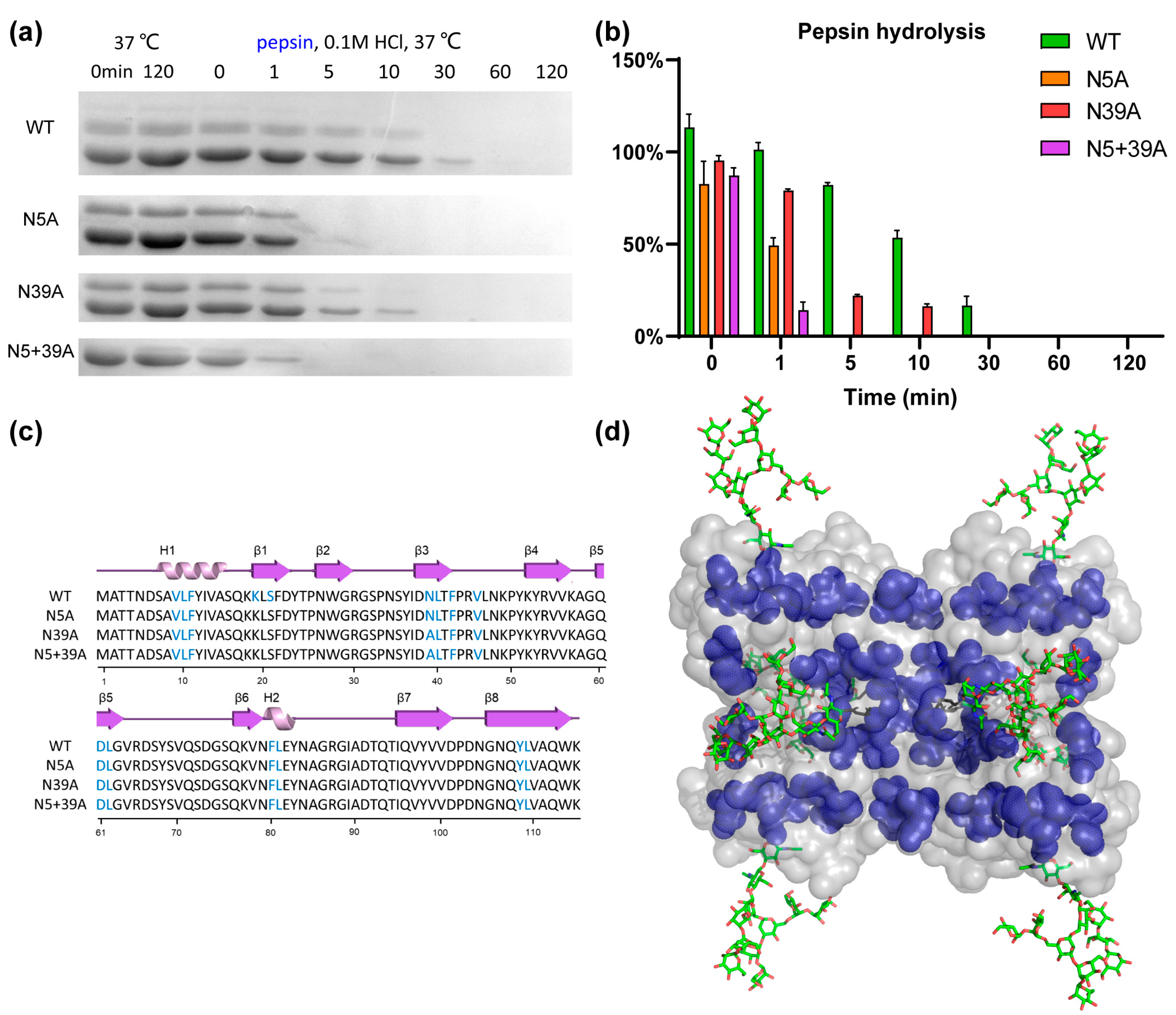
2.4. Proteolytic Resistance
2.5. Bioactivity of FIP-nha and Its Variants
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Gene and Mutagenesis Construction of FIP-nha
3.3. Protein Preparation
3.4. Oligomerization State Analysis
3.5. Thermostability Measurements
3.6. Protein Crystallization, X-ray Data Collection, Structure Determination and Refinement
3.7. Digestion Resistance Measurements
3.8. Bioactivity Measurements
3.9. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, D.F.; Gnad, F.; Wiśniewski, J.R.; Mann, M. Precision mapping of an in vivo N-glycoproteome reveals rigid topological and sequence constraints. Cell 2010, 141, 897–907. [Google Scholar] [CrossRef]
- Eichler, J. Protein glycosylation. Curr. Biol. 2019, 29, R229–R231. [Google Scholar] [CrossRef]
- Mammadova-Bach, E.; Jaeken, J.; Gudermann, T.; Braun, A. Platelets and Defective N-Glycosylation. Int. J. Mol. Sci. 2020, 21, 5630. [Google Scholar] [CrossRef]
- Esmail, S.; Manolson, M.F. Advances in understanding N-glycosylation structure, function, and regulation in health and disease. Eur. J. Cell Biol. 2021, 100, 151186. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-W.; Lim, S.-O.; Xia, W.; Lee, H.-H.; Chan, L.-C.; Kuo, C.-W.; Khoo, K.-H.; Chang, S.-S.; Cha, J.-H.; Kim, T.; et al. Glycosylation and stabilization of programmed death ligand-1 suppresses T-cell activity. Nat. Commun. 2016, 7, 12632. [Google Scholar] [CrossRef]
- Han, C.; Wang, Q.; Sun, Y.; Yang, R.; Liu, M.; Wang, S.; Liu, Y.; Zhou, L.; Li, D. Improvement of the catalytic activity and thermostability of a hyperthermostable endoglucanase by optimizing N-glycosylation sites. Biotechnol. Biofuels 2020, 13, 30. [Google Scholar] [CrossRef]
- Niu, C.; Luo, H.; Shi, P.; Huang, H.; Wang, Y.; Yang, P.; Yao, B. N-Glycosylation Improves the Pepsin Resistance of Histidine Acid Phosphatase Phytases by Enhancing Their Stability at Acidic pHs and Reducing Pepsin’s Accessibility to Its Cleavage Sites. Appl. Environ. Microbiol. 2016, 82, 1004–1014. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fan, C.; Zhang, L.; Ma, X. Glycosylation-dependent antitumor therapeutic monoclonal antibodies. Prog. Mol. Biol. Transl. Sci. 2019, 163, 471–485. [Google Scholar] [CrossRef]
- Jones, A.J.S.; Papac, D.I.; Chin, E.H.; Keck, R.; Baughman, S.A.; Lin, Y.S.; Kneer, J.; Battersby, J.E. Selective clearance of glycoforms of a complex glycoprotein pharmaceutical caused by terminal N-acetylglucosamine is similar in humans and cynomolgus monkeys. Glycobiology 2007, 17, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Erjavec, J.; Kos, J.; Ravnikar, M.; Dreo, T.; Sabotič, J. Proteins of higher fungi—From forest to application. Trends Biotechnol. 2012, 30, 259–273. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, Y.; Zhou, X. Fungal immunomodulatory proteins: Characteristic, potential antitumor activities and their molecular mechanisms. Drug Discov. Today 2018, 24, 307–314. [Google Scholar] [CrossRef]
- Ejike, U.C.; Chan, C.J.; Okechukwu, P.N.; Lim, R.L.H. New advances and potentials of fungal immunomodulatory proteins for therapeutic purposes. Crit. Rev. Biotechnol. 2020, 40, 1172–1190. [Google Scholar] [CrossRef]
- Liu, Y.; Bastiaan-Net, S.; Wichers, H. Current understanding of the structure and function of fungal immunomodulatory proteins. Front. Nutr. 2020, 7, 132. [Google Scholar] [CrossRef]
- Bastiaan-Net, S.; Chanput, W.; Hertz, A.; Zwittink, R.D.; Mes, J.J.; Wichers, H.J. Biochemical and functional characterization of recombinant fungal immunomodulatory proteins (rFIPs). Int. Immunopharmacol. 2013, 15, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bastiaan-Net, S.; Zhang, Y.; Hoppenbrouwers, T.; Xie, Y.; Wang, Y.; Wei, X.; Du, G.; Zhang, H.; Imam, K.M.D.S.U.; et al. Linking the thermostability of FIP-nha (Nectria haematococca) to its structural properties. Int. J. Biol. Macromol. 2022, 213, 555–564. [Google Scholar] [CrossRef]
- Chang, H.H.; Yeh, C.H.; Sheu, F. A novel immunomodulatory protein from Poria cocos induces Toll-like receptor 4-dependent activation within mouse peritoneal macrophages. J. Agric. Food Chem. 2009, 57, 6129–6139. [Google Scholar] [CrossRef] [PubMed]
- Sheu, F.; Chien, P.J.; Hsieh, K.Y.; Chin, K.L.; Huang, W.T.; Tsao, C.Y.; Chen, Y.F.; Cheng, H.C.; Chang, H.H. Purification, cloning, and functional characterization of a novel immunomodulatory protein from Antrodia camphorata (bitter mushroom) that exhibits TLR2-dependent NF-kappaB activation and M1 polarization within murine macrophages. J. Agric. Food Chem. 2009, 57, 4130–4141. [Google Scholar] [CrossRef]
- Wu, T.-Y.; Chen, H.-A.; Li, F.-Y.; Lin, C.-T.; Wu, C.-M.; Hsieh, F.-C.; Tzen, J.T.-C.; Hsieh, S.-K.; Ko, J.-L.; Jinn, T.-R. High-level expression, purification and production of the fungal immunomodulatory protein-gts in baculovirus-infected insect larva. Appl. Biochem. Biotechnol. 2013, 169, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Mesters, J.R.; Hilgenfeld, R. Protein Glycosylation, Sweet to Crystal Growth? Cryst. Growth Des. 2007, 7, 2251–2253. [Google Scholar] [CrossRef]
- Bohne-Lang, A.; von der Lieth, C.W. GlyProt: In silico glycosylation of proteins. Nucleic Acids Res. 2005, 33, W214–W219. [Google Scholar] [CrossRef] [PubMed]
- Macauley-Patrick, S.; Fazenda, M.L.; McNeil, B.; Harvey, L.M. Heterologous protein production using the Pichia pastoris expression system. Yeast 2005, 22, 249–270. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.Z.; Wang, X.; Wang, W.; Fu, Y.J.; Liang, A.H. Improving the thermostability of Escherichia coli phytase, appA, by enhancement of glycosylation. Biotechnol. Lett. 2013, 35, 1669–1676. [Google Scholar] [CrossRef]
- Ma, J.; Li, Q.; Tan, H.; Jiang, H.; Li, K.; Zhang, L.; Shi, Q.; Yin, H. Unique N-glycosylation of a recombinant exo-inulinase from Kluyveromyces cicerisporus and its effect on enzymatic activity and thermostability. J. Biol. Eng. 2019, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Shental-Bechor, D.; Levy, Y. Effect of glycosylation on protein folding: A close look at thermodynamic stabilization. Proc. Natl. Acad. Sci. USA 2008, 105, 8256–8261. [Google Scholar] [CrossRef]
- Kilgore, H.R.; Latham, A.P.; Ressler, V.T.; Zhang, B.; Raines, R.T. Structure and Dynamics of N-Glycosylated Human Ribonuclease 1. Biochemistry 2020, 59, 3148–3156. [Google Scholar] [CrossRef]
- Deng, Y.; Gruppen, H.; Wierenga, P.A. Comparison of Protein Hydrolysis Catalyzed by Bovine, Porcine, and Human Trypsins. J. Agric. Food Chem. 2018, 66, 4219–4232. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; He, Z.; Chen, J.; Liu, Y.; Ma, M.; Cai, Z. The impact of N-glycosylation on conformation and stability of immunoglobulin Y from egg yolk. Int. J. Biol. Macromol. 2017, 96, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Liu, X.; Li, Y.; Liu, D.; Kuang, Z.; Qian, C.; Yao, D. Rational design for the stability improvement of Armillariella tabescens β-mannanase MAN47 based on N-glycosylation modification. Enzym. Microb. Technol. 2017, 97, 82–89. [Google Scholar] [CrossRef]
- Ihara, S.; Miyoshi, E.; Nakahara, S.; Sakiyama, H.; Ihara, H.; Akinaga, A.; Honke, K.; Dickson, R.B.; Lin, C.-Y.; Taniguchi, N. Addition of beta1-6 GlcNAc branching to the oligosaccharide attached to Asn 772 in the serine protease domain of matriptase plays a pivotal role in its stability and resistance against trypsin. Glycobiology 2004, 14, 139–146. [Google Scholar] [CrossRef]
- Zou, S.; Huang, S.; Kaleem, I.; Li, C. N-Glycosylation enhances functional and structural stability of recombinant β-glucuronidase expressed in Pichia pastoris. J. Biotechnol. 2013, 164, 75–81. [Google Scholar] [CrossRef]
- Li, S.-Y.; Hou, L.-Z.; Gao, Y.-X.; Zhang, N.-N.; Fan, B.; Wang, F. FIP-nha, a fungal immunomodulatory protein from Nectria haematococca, induces apoptosis and autophagy in human gastric cancer cells via blocking the EGFR-mediated STAT3/Akt signaling pathway. Food Chem. Mol. Sci. 2022, 4, 100091. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, S.; Sun, L.; Liu, S.; Wang, F.; Wen, B.; Sun, L.; Fang, X.; Cha, Y.; Cao, H.; et al. Fungal immunomodulatory protein from Nectria haematococca suppresses growth of human lung adenocarcinoma by inhibiting the PI3K/Akt pathway. Int. J. Mol. Sci. 2018, 19, 3429. [Google Scholar] [CrossRef]
- Li, S.; Nie, Y.; Ding, Y.; Shi, L.; Tang, X. Recombinant expression of a novel fungal immunomodulatory protein with human tumor cell antiproliferative activity from Nectria haematococca. Int. J. Mol. Sci. 2014, 15, 17751–17764. [Google Scholar] [CrossRef]
- Oliveira, C.; Teixeira, J.A.; Domingues, L. Recombinant lectins: An array of tailor-made glycan-interaction biosynthetic tools. Crit. Rev. Biotechnol. 2013, 33, 66–80. [Google Scholar] [CrossRef]
- Oliveira, C.; Freitas, A.I.; Campos, N.; Saraiva, L.; Domingues, L. Cytotoxicity of Frutalin on Distinct Cancer Cells Is Independent of Its Glycosylation. Molecules 2021, 26, 4712. [Google Scholar] [CrossRef] [PubMed]
- Biswas, H.; Chattopadhyaya, R. Stability of Curcuma longa rhizome lectin: Role of N-linked glycosylation. Glycobiology 2016, 26, 410–426. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, Q.-Z.; Chen, X.; Mao, P.-W.; Jin, M.-Y.; Wu, Q.; Zhou, X.-W. N-Glycosylated Ganoderma lucidum immunomodulatory protein improved anti-inflammatory activity via inhibition of the p38 MAPK pathway. Food Funct. 2021, 12, 3393–3404. [Google Scholar] [CrossRef]
- Jacobs, M.R.; Grace, M.; Blumlein, A.; McManus, J.J. Differential scanning calorimetry to quantify heat-induced aggregation in concentrated protein solutions. Methods Mol. Biol. 2019, 2039, 117–129. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Meth. Enzymol. 1997, 276, 307–326. [Google Scholar] [CrossRef]
- Hough, M.A.; Wilson, K.S. From crystal to structure with CCP4. Acta Crystallogr. Sect. D Struct. Biol. 2018, 74 Pt 2, 67. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Grosse-Kunstleve, R.W.; Hung, L.W.; Ioerger, T.R.; McCoy, A.J.; Moriarty, N.W.; Read, R.J.; Sacchettini, J.C.; Sauter, N.K.; Terwilliger, T.C. PHENIX: Building new software for automated crystallographic structure determination. Acta Crystallogr. Sect. D 2002, 58 Pt 11, 1948–1954. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D 2004, 60 Pt 12, 2126–2132. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FIP-nha | Helix | Antiparallel | Parallel | Beta-Turn | Random Coil | Total Sum |
|---|---|---|---|---|---|---|
| WT | 6.3% | 48.5% | 3.7% | 16.5% | 29.3% | 104.3% |
| N5A | 6.4% | 48.1% | 3.7% | 16.5% | 29.4% | 104.1% |
| N39A | 6.4% | 46.9% | 3.7% | 16.7% | 29.8% | 103.6% |
| N5+39A | 6.4% | 46.8% | 3.7% | 16.6% | 30.0% | 103.5% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Hoppenbrouwers, T.; Wang, Y.; Xie, Y.; Wei, X.; Zhang, H.; Du, G.; Imam, K.M.S.U.; Wichers, H.; Li, Z.; et al. Glycosylation Contributes to Thermostability and Proteolytic Resistance of rFIP-nha (Nectria haematococca). Molecules 2023, 28, 6386. https://doi.org/10.3390/molecules28176386
Liu Y, Hoppenbrouwers T, Wang Y, Xie Y, Wei X, Zhang H, Du G, Imam KMSU, Wichers H, Li Z, et al. Glycosylation Contributes to Thermostability and Proteolytic Resistance of rFIP-nha (Nectria haematococca). Molecules. 2023; 28(17):6386. https://doi.org/10.3390/molecules28176386
Chicago/Turabian StyleLiu, Yusi, Tamara Hoppenbrouwers, Yulu Wang, Yingying Xie, Xue Wei, Haowen Zhang, Guoming Du, Khandader Md Sharif Uddin Imam, Harry Wichers, Zhen Li, and et al. 2023. "Glycosylation Contributes to Thermostability and Proteolytic Resistance of rFIP-nha (Nectria haematococca)" Molecules 28, no. 17: 6386. https://doi.org/10.3390/molecules28176386
APA StyleLiu, Y., Hoppenbrouwers, T., Wang, Y., Xie, Y., Wei, X., Zhang, H., Du, G., Imam, K. M. S. U., Wichers, H., Li, Z., & Bastiaan-Net, S. (2023). Glycosylation Contributes to Thermostability and Proteolytic Resistance of rFIP-nha (Nectria haematococca). Molecules, 28(17), 6386. https://doi.org/10.3390/molecules28176386