Elucidation of Pharmacological Mechanism Underlying the Anti-Alzheimer’s Disease Effects of Evodia rutaecarpa and Discovery of Novel Lead Molecules: An In Silico Study


Abstract
1. Introduction
2. Results
2.1. Active Compounds of WZY Collection and Toxicity Study
2.2. Target Prediction
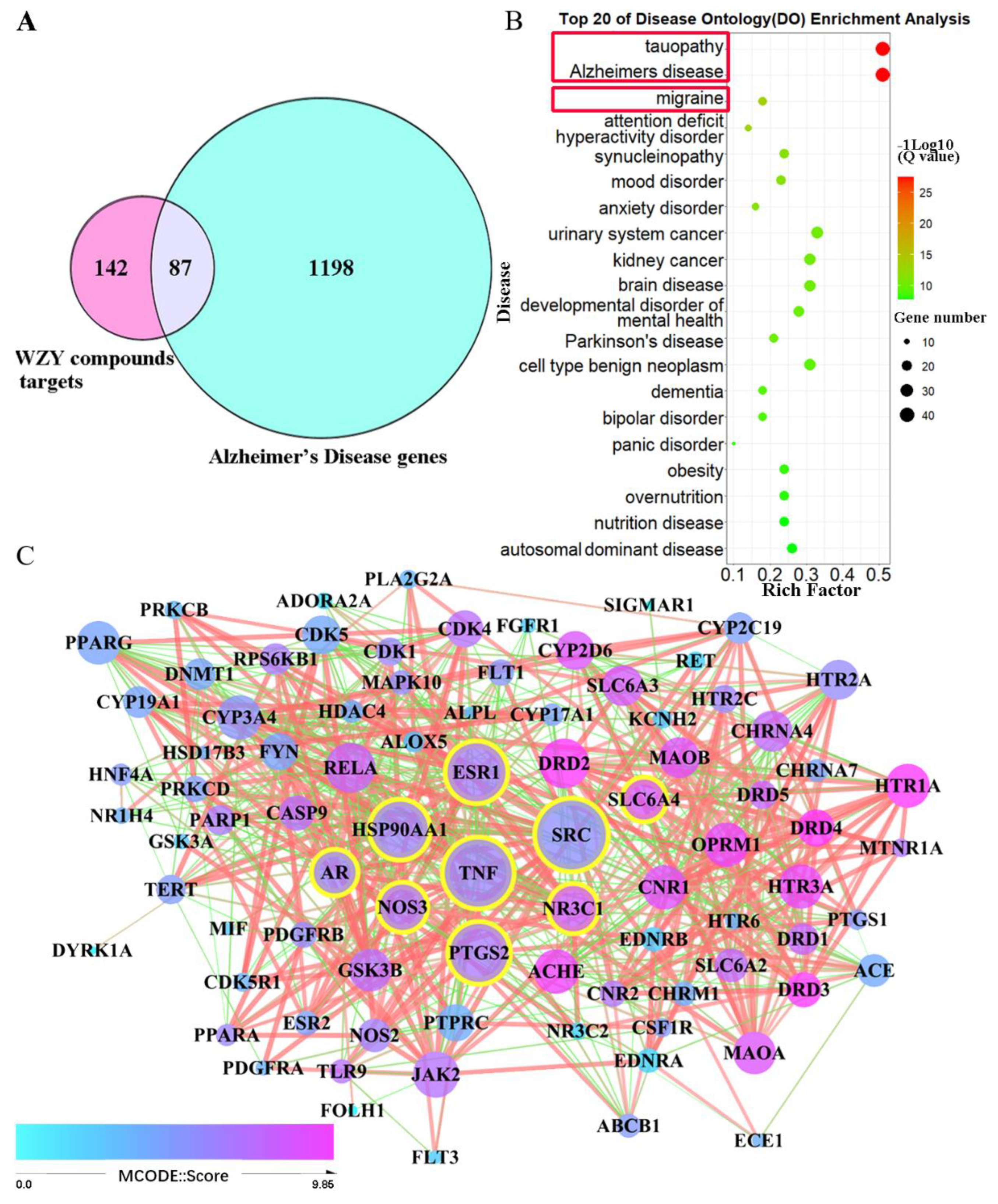
2.3. Alzheimer’s Disease, the Disease in Which Core 229 Target Genes Are Mainly Enriched
2.4. Neural Construction, Activities, and Pathways That Core 229 Target Genes Primarily Enriched
2.5. AD Target Genes’ Collection, Compound–AD Intersection, and PPI Network Construction
2.6. The 87 WZY–AD Intersection Genes Mainly Enriched in Alzheimer’s Disease
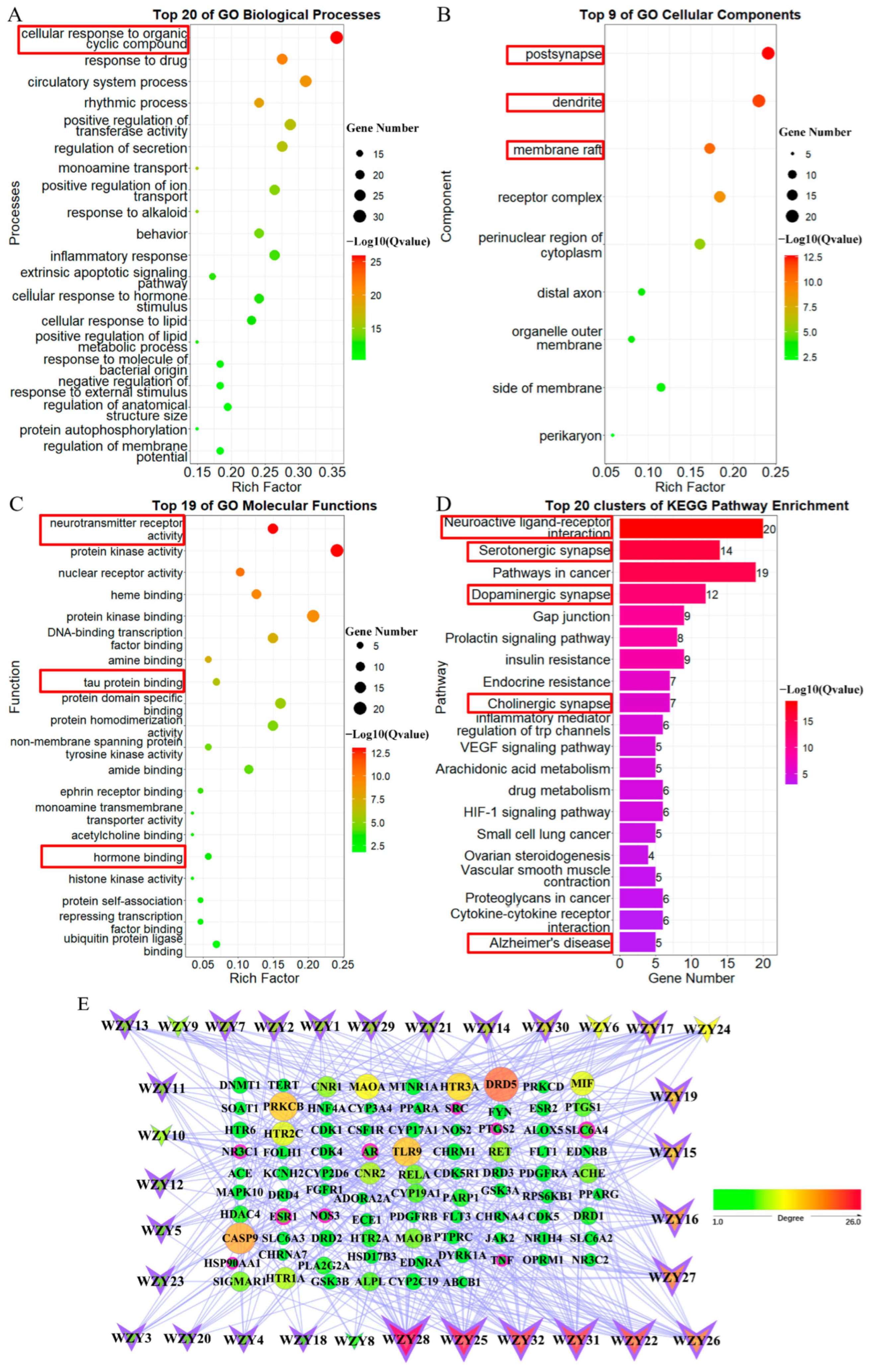
2.7. GO and KEGG Pathway Enrichment Analysis of 87 WZY–AD Intersection Genes
2.8. Network Construction of Compounds Targeted to WZY–AD Intersection Genes
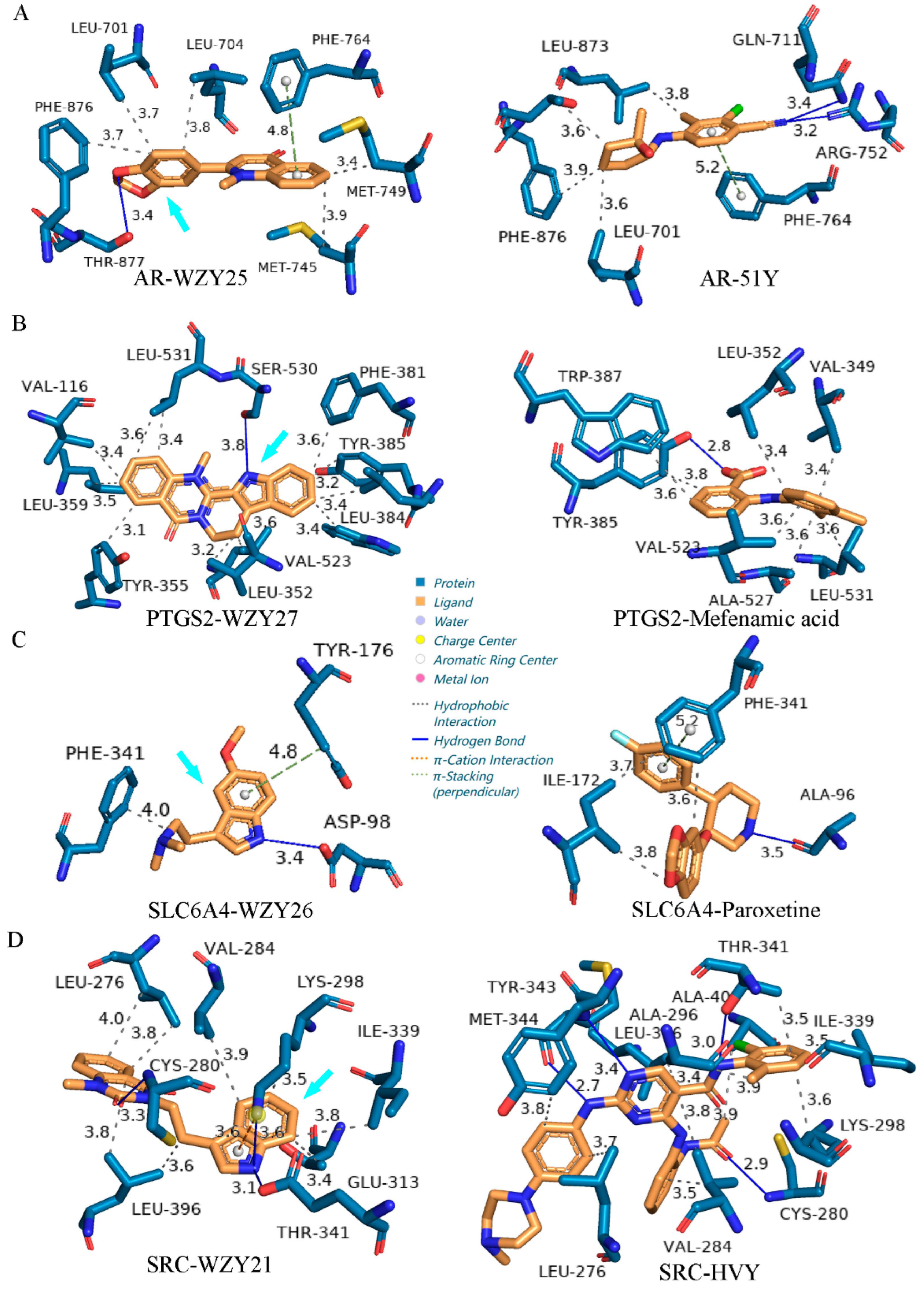
2.9. Molecular Docking
2.10. Protein–Ligand Interaction Comparison of Potential Compounds versus Positive Controls
3. Discussion
4. Materials and Methods
4.1. Compound Collection, ADMET Screening, and Toxicity Prediction
4.2. Target Prediction
4.3. PPI Networking
4.4. Disease Ontology Enrichment Analysis
4.5. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis
4.6. Compound–Target Networking
4.7. AD Target Screening and Identification of Target–AD Intersection Genes
4.8. Molecular Docking
4.9. Protein–Ligand Interaction Profiler (PLIP)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chetelat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- Long, J.M.; Holtzman, D.M. Alzheimer Disease: An Update on Pathobiology and Treatment Strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. Amyloid-Beta Solubility in the Treatment of Alzheimer’s Disease. N. Engl. J. Med. 2018, 378, 391–392. [Google Scholar] [CrossRef]
- Bloom, G.S. Amyloid-beta and tau: The trigger and bullet in Alzheimer disease pathogenesis. JAMA Neurol. 2014, 71, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Congdon, E.E.; Sigurdsson, E.M. Tau-targeting therapies for Alzheimer disease. Nat. Rev. Neurol. 2018, 14, 399–415. [Google Scholar] [CrossRef]
- Serrano-Pozo, A.; Das, S.; Hyman, B.T. APOE and Alzheimer’s disease: Advances in genetics, pathophysiology, and therapeutic approaches. Lancet Neurol. 2021, 20, 68–80. [Google Scholar] [CrossRef]
- Montagne, A.; Nation, D.A.; Sagare, A.P.; Barisano, G.; Sweeney, M.D.; Chakhoyan, A.; Pachicano, M.; Joe, E.; Nelson, A.R.; D’Orazio, L.M.; et al. APOE4 leads to blood-brain barrier dysfunction predicting cognitive decline. Nature 2020, 581, 71–76. [Google Scholar] [CrossRef]
- Cunnane, S.C.; Trushina, E.; Morland, C.; Prigione, A.; Casadesus, G.; Andrews, Z.B.; Beal, M.F.; Bergersen, L.H.; Brinton, R.D.; de la Monte, S.; et al. Brain energy rescue: An emerging therapeutic concept for neurodegenerative disorders of ageing. Nat. Rev. Drug Discov. 2020, 19, 609–633. [Google Scholar] [CrossRef]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: Where do we go from here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef]
- Burgaletto, C.; Munafo, A.; Di Benedetto, G.; De Francisci, C.; Caraci, F.; Di Mauro, R.; Bucolo, C.; Bernardini, R.; Cantarella, G. The immune system on the TRAIL of Alzheimer’s disease. J. Neuroinflammation 2020, 17, 298. [Google Scholar] [CrossRef]
- Honig, L.S.; Vellas, B.; Woodward, M.; Boada, M.; Bullock, R.; Borrie, M.; Hager, K.; Andreasen, N.; Scarpini, E.; Liu-Seifert, H.; et al. Trial of Solanezumab for Mild Dementia Due to Alzheimer’s Disease. N. Engl. J. Med. 2018, 378, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Salloway, S.; Sperling, R.; Fox, N.C.; Blennow, K.; Klunk, W.; Raskind, M.; Sabbagh, M.; Honig, L.S.; Porsteinsson, A.P.; Ferris, S.; et al. Two phase 3 trials of bapineuzumab in mild-to-moderate Alzheimer’s disease. N. Engl. J. Med. 2014, 370, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.; Lee, G.; Zhong, K.; Fonseca, J.; Taghva, K. Alzheimer’s disease drug development pipeline: 2021. Alzheimers Dement. 2021, 7, e12179. [Google Scholar] [CrossRef] [PubMed]
- Hung, S.Y.; Fu, W.M. Drug candidates in clinical trials for Alzheimer’s disease. J. Biomed. Sci. 2017, 24, 47. [Google Scholar] [CrossRef]
- Egan, M.F.; Kost, J.; Voss, T.; Mukai, Y.; Aisen, P.S.; Cummings, J.L.; Tariot, P.N.; Vellas, B.; van Dyck, C.H.; Boada, M.; et al. Randomized Trial of Verubecestat for Prodromal Alzheimer’s Disease. N. Engl. J. Med. 2019, 380, 1408–1420. [Google Scholar] [CrossRef]
- Li, W.; Sun, X.; Liu, B.; Zhang, L.; Fan, Z.; Ji, Y. Screening and identification of hepatotoxic component in Evodia rutaecarpa based on spectrum-effect relationship and UPLC-Q-TOFMS. Biomed. Chromatogr. 2016, 30, 1975–1983. [Google Scholar] [CrossRef]
- Wang, L. The Intestinal Metabolism, Absorption Distribution and Effects on Alzheimer’s Disease Mouse Models of Evodia Evodia Extract; Jilin Agricultural University: Changchun, China, 2019. [Google Scholar]
- Kumar, A.; Rajendran, V.; Sethumadhavan, R.; Purohit, R. In silico prediction of a disease-associated STIL mutant and its affect on the recruitment of centromere protein J (CENPJ). FEBS Open Bio. 2012, 2, 285–293. [Google Scholar] [CrossRef]
- Purohit, R.; Rajasekaran, R.; Sudandiradoss, C.; George Priya Doss, C.; Ramanathan, K.; Rao, S. Studies on flexibility and binding affinity of Asp25 of HIV-1 protease mutants. Int. J. Biol. Macromol. 2008, 42, 386–391. [Google Scholar] [CrossRef]
- Pei, H.; Ma, L.; Cao, Y.; Wang, F.; Li, Z.; Liu, N.; Liu, M.; Wei, Y.; Li, H. Traditional Chinese Medicine for Alzheimer’s Disease and Other Cognitive Impairment: A Review. Am. J. Chin. Med. 2020, 48, 487–511. [Google Scholar] [CrossRef]
- Yang, W.T.; Zheng, X.W.; Chen, S.; Shan, C.S.; Xu, Q.Q.; Zhu, J.Z.; Bao, X.Y.; Lin, Y.; Zheng, G.Q.; Wang, Y. Chinese herbal medicine for Alzheimer’s disease: Clinical evidence and possible mechanism of neurogenesis. Biochem. Pharmacol. 2017, 141, 143–155. [Google Scholar] [CrossRef]
- Ling, Y.; Hu, P.; Zhang, L.; Jin, H.; Chen, J.; Tao, Z.; Huang, L.; Ren, R. Identification and Structural Characterization of Acylgluconic Acids, Flavonol Glycosides, Limonoids and Alkaloids from the Fruits of Evodia Rutaecarpa by High Performance Liquid Chromatography Coupled to Electrospray Ionization and Quadrupole Time-of-Flight Mass Spectrometry. J. Chromatogr. Sci. 2016, 54, 1593–1604. [Google Scholar] [PubMed]
- Wang, X.X.; Zan, K.; Shi, S.P.; Zeng, K.W.; Jiang, Y.; Guan, Y.; Xiao, C.L.; Gao, H.Y.; Wu, L.J.; Tu, P.F. Quinolone alkaloids with antibacterial and cytotoxic activities from the fruits of Evodia rutaecarpa. Fitoterapia 2013, 89, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhang, Y.B.; Yang, X.W. Indoloquinazoline alkaloids from Euodia rutaecarpa and their cytotoxic activities. J. Asian Nat. Prod. Res. 2011, 13, 977–983. [Google Scholar] [CrossRef] [PubMed]
- Tanwar, G.; Mazumder, A.G.; Bhardwaj, V.; Kumari, S.; Bharti, R.; Yamini; Singh, D.; Das, P.; Purohit, R. Target identification, screening and in vivo evaluation of pyrrolone-fused benzosuberene compounds against human epilepsy using Zebrafish model of pentylenetetrazol-induced seizures. Sci. Rep. 2019, 9, 7904. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, V.; Purohit, R. Computational investigation on effect of mutations in PCNA resulting in structural perturbations and inhibition of mismatch repair pathway. J. Biomol. Struct. Dyn. 2020, 38, 1963–1974. [Google Scholar] [CrossRef] [PubMed]
- Kumar Bhardwaj, V.; Purohit, R.; Kumar, S. Himalayan bioactive molecules as potential entry inhibitors for the human immunodeficiency virus. Food Chem. 2021, 347, 128932. [Google Scholar] [CrossRef]
- Bhardwaj, V.K.; Oakley, A.; Purohit, R. Mechanistic behavior and subtle key events during DNA clamp opening and closing in T4 bacteriophage. Int. J. Biol. Macromol. 2022, 208, 11–19. [Google Scholar] [CrossRef]
- Saeed, A.; Vaught, G.M.; Gavardinas, K.; Matthews, D.; Green, J.E.; Losada, P.G.; Bullock, H.A.; Calvert, N.A.; Patel, N.J.; Sweetana, S.A.; et al. 2-Chloro-4-[[(1R,2R)-2-hydroxy-2-methyl-cyclopentyl]amino]-3-methyl-benzonitrile: A Transdermal Selective Androgen Receptor Modulator (SARM) for Muscle Atrophy. J. Med. Chem. 2016, 59, 750–755. [Google Scholar] [CrossRef]
- Orlando, B.J.; Malkowski, M.G. Substrate-Selective Inhibition of Cyclooxygeanse-2 by Fenamic Acid Derivatives Is Dependent on Peroxide Tone. J. Biol. Chem. 2016, 291, 15069–15081. [Google Scholar] [CrossRef]
- Mattson, R.J.; Catt, J.D.; Denhart, D.J.; Deskus, J.A.; Ditta, J.L.; Higgins, M.A.; Marcin, L.R.; Sloan, C.P.; Beno, B.R.; Gao, Q.; et al. Conformationally restricted homotryptamines. 2. Indole cyclopropylmethylamines as selective serotonin reuptake inhibitors. J. Med. Chem. 2005, 48, 6023–6034. [Google Scholar] [CrossRef]
- Du, G.; Rao, S.; Gurbani, D.; Henning, N.J.; Jiang, J.; Che, J.; Yang, A.; Ficarro, S.B.; Marto, J.A.; Aguirre, A.J.; et al. Structure-Based Design of a Potent and Selective Covalent Inhibitor for SRC Kinase That Targets a P-Loop Cysteine. J. Med. Chem. 2020, 63, 1624–1641. [Google Scholar] [CrossRef]
- Chou, C.H.; Yang, C.R. Neuroprotective Studies of Evodiamine in an Okadaic Acid-Induced Neurotoxicity. Int. J. Mol. Sci. 2021, 22, 5347. [Google Scholar] [CrossRef]
- Fang, Z.; Tang, Y.; Ying, J.; Tang, C.; Wang, Q. Traditional Chinese medicine for anti-Alzheimer’s disease: Berberine and evodiamine from Evodia rutaecarpa. Chin. Med. 2020, 15, 82. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B.; Harney, J.T.; Ahmed, B.A.; Jeffus, B.C.; Unal, R.; Mehta, J.L.; Kilic, F. Plasma serotonin levels and the platelet serotonin transporter. J. Neurochem. 2007, 102, 206–215. [Google Scholar] [CrossRef] [PubMed]
- van der Zande, J.J.; Joling, M.; Happach, I.G.; Vriend, C.; Scheltens, P.; Booij, J.; Lemstra, A.W. Serotonergic deficits in dementia with Lewy bodies with concomitant Alzheimer’s disease pathology: An (123)I-FP-CIT SPECT study. Neuroimage Clin. 2020, 25, 102062. [Google Scholar] [CrossRef]
- Metaxas, A.; Anzalone, M.; Vaitheeswaran, R.; Petersen, S.; Landau, A.M.; Finsen, B. Neuroinflammation and amyloid-beta 40 are associated with reduced serotonin transporter (SERT) activity in a transgenic model of familial Alzheimer’s disease. Alzheimers Res. Ther. 2019, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.S.; Barrett, F.S.; Joo, J.H.; Nassery, N.; Savonenko, A.; Sodums, D.J.; Marano, C.M.; Munro, C.A.; Brandt, J.; Kraut, M.A.; et al. Molecular imaging of serotonin degeneration in mild cognitive impairment. Neurobiol. Dis. 2017, 105, 33–41. [Google Scholar] [CrossRef]
- Ogilvie, A.D.; Battersby, S.; Bubb, V.J.; Fink, G.; Harmar, A.J.; Goodwim, G.M.; Smith, C.A. Polymorphism in serotonin transporter gene associated with susceptibility to major depression. Lancet 1996, 347, 731–733. [Google Scholar] [CrossRef] [PubMed]
- Lesch, K.P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Muller, C.R.; Hamer, D.H.; Murphy, D.L. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef]
- Hariri, A.R.; Mattay, V.S.; Tessitore, A.; Kolachana, B.; Fera, F.; Goldman, D.; Egan, M.F.; Weinberger, D.R. Serotonin transporter genetic variation and the response of the human amygdala. Science 2002, 297, 400–403. [Google Scholar] [CrossRef]
- Dhawan, G.; Combs, C.K. Inhibition of Src kinase activity attenuates amyloid associated microgliosis in a murine model of Alzheimer’s disease. J. Neuroinflammation 2012, 9, 117. [Google Scholar] [CrossRef] [PubMed]
- Mota, S.I.; Ferreira, I.L.; Valero, J.; Ferreiro, E.; Carvalho, A.L.; Oliveira, C.R.; Rego, A.C. Impaired Src signaling and post-synaptic actin polymerization in Alzheimer’s disease mice hippocampus--linking NMDA receptors and the reelin pathway. Exp. Neurol. 2014, 261, 698–709. [Google Scholar] [CrossRef] [PubMed]
- Socodato, R.; Portugal, C.C.; Canedo, T.; Rodrigues, A.; Almeida, T.O.; Henriques, J.F.; Vaz, S.H.; Magalhães, J.; Silva, C.M.; Baptista, F.I.; et al. Microglia Dysfunction Caused by the Loss of Rhoa Disrupts Neuronal Physiology and Leads to Neurodegeneration. Cell Rep. 2020, 31, 107796. [Google Scholar]
- McCusker, S.M.; Curran, M.D.; Dynan, K.B.; McCullagh, C.D.; Urquhart, D.D.; Middleton, D.; Patterson, C.C.; McIlroy, S.P.; Passmore, A.P. Association between polymorphism in regulatory region of gene encoding tumour necrosis factor alpha and risk of Alzheimer’s disease and vascular dementia: A case-control study. Lancet 2001, 357, 436–439. [Google Scholar] [PubMed]
- Rodriguez-Acevedo, A.J.; Maher, B.H.; Lea, R.A.; Benton, M.; Griffiths, L.R. Association of oestrogen-receptor gene (ESR1) polymorphisms with migraine in the large Norfolk Island pedigree. Cephalalgia 2013, 33, 1139–1147. [Google Scholar]
- An, X.; Fang, J.; Lin, Q.; Lu, C.; Ma, Q.; Qu, H. New evidence for involvement of ESR1 gene in susceptibility to Chinese migraine. J. Neurol. 2017, 264, 81–87. [Google Scholar] [CrossRef]
- Pike, C.J. Sex and the development of Alzheimer’s disease. J. Neurosci. Res. 2017, 95, 671–680. [Google Scholar] [CrossRef]
- Foster, T.C. Role of estrogen receptor alpha and beta expression and signaling on cognitive function during aging. Hippocampus 2012, 22, 656–669. [Google Scholar] [CrossRef]
- Barnes, L.L.; Wilson, R.S.; Bienias, J.L.; Schneider, J.A.; Evans, D.A.; Bennett, D.A. Sex differences in the clinical manifestations of Alzheimer disease pathology. Arch. Gen. Psychiatry 2005, 62, 685–691. [Google Scholar] [CrossRef]
- Uddin, M.S.; Rahman, M.M.; Jakaria, M.; Rahman, M.S.; Hossain, M.S.; Islam, A.; Ahmed, M.; Mathew, B.; Omar, U.M.; Barreto, G.E.; et al. Estrogen Signaling in Alzheimer’s Disease: Molecular Insights and Therapeutic Targets for Alzheimer’s Dementia. Mol. Neurobiol. 2020, 57, 2654–2670. [Google Scholar] [CrossRef]
- Xu, H.; Gouras, G.K.; Greenfield, J.P.; Vincent, B.; Naslund, J.; Mazzarelli, L.; Fried, G.; Jovanovic, J.N.; Seeger, M.; Relkin, N.R.; et al. Estrogen reduces neuronal generation of Alzheimer beta-amyloid peptides. Nat. Med. 1998, 4, 447–451. [Google Scholar]
- Pompili, A.; Arnone, B.; Gasbarri, A. Estrogens and memory in physiological and neuropathological conditions. Psychoneuroendocrinology 2012, 37, 1379–1396. [Google Scholar] [CrossRef]
- Correia, S.C.; Santos, R.X.; Cardoso, S.; Carvalho, C.; Santos, M.S.; Oliveira, C.R.; Moreira, P.I. Effects of estrogen in the brain: Is it a neuroprotective agent in Alzheimer’s disease? Curr. Aging Sci. 2010, 3, 113–126. [Google Scholar] [CrossRef]
- Lee, J.H.; Jiang, Y.; Han, D.H.; Shin, S.K.; Choi, W.H.; Lee, M.J. Targeting estrogen receptors for the treatment of Alzheimer’s disease. Mol. Neurobiol. 2014, 49, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Goodenough, S.; Schleusner, D.; Pietrzik, C.; Skutella, T.; Behl, C. Glycogen synthase kinase 3beta links neuroprotection by 17beta-estradiol to key Alzheimer processes. Neuroscience 2005, 132, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.G.; Wang, R.; Khan, M.; Mahesh, V.; Brann, D.W. Role of Dickkopf-1, an antagonist of the Wnt/beta-catenin signaling pathway, in estrogen-induced neuroprotection and attenuation of tau phosphorylation. J. Neurosci. 2008, 28, 8430–8441. [Google Scholar] [CrossRef]
- Russell, J.K.; Jones, C.K.; Newhouse, P.A. The Role of Estrogen in Brain and Cognitive Aging. Neurotherapeutics 2019, 16, 649–665. [Google Scholar] [CrossRef]
- Gonzalez, B.D.; Jim, H.S.; Booth-Jones, M.; Small, B.J.; Sutton, S.K.; Lin, H.Y.; Park, J.Y.; Spiess, P.E.; Fishman, M.N.; Jacobsen, P.B. Course and Predictors of Cognitive Function in Patients with Prostate Cancer Receiving Androgen-Deprivation Therapy: A Controlled Comparison. J. Clin. Oncol. 2015, 33, 2021–2027. [Google Scholar] [PubMed]
- Rosario, E.R.; Chang, L.; Stanczyk, F.Z.; Pike, C.J. Age-related testosterone depletion and the development of Alzheimer disease. JAMA 2004, 292, 1431–1432. [Google Scholar] [CrossRef] [PubMed]
- Hogervorst, E.; Combrinck, M.; Smith, A.D. Testosterone and gonadotropin levels in men with dementia. Neuro Endocrinol. Lett. 2003, 24, 203–208. [Google Scholar]
- Carcaillon, L.; Brailly-Tabard, S.; Ancelin, M.L.; Tzourio, C.; Foubert-Samier, A.; Dartigues, J.F.; Guiochon-Mantel, A.; Scarabin, P.Y. Low testosterone and the risk of dementia in elderly men: Impact of age and education. Alzheimers Dement. 2014, 10 (Suppl. S5), S306–S314. [Google Scholar] [PubMed]
- Akita, K.; Harada, K.; Ichihara, J.; Takata, N.; Takahashi, Y.; Saito, K. A novel selective androgen receptor modulator, NEP28, is efficacious in muscle and brain without serious side effects on prostate. Eur. J. Pharmacol. 2013, 720, 107–114. [Google Scholar] [PubMed]
- Christiansen, A.R.; Lipshultz, L.I.; Hotaling, J.M.; Pastuszak, A.W. Selective androgen receptor modulators: The future of androgen therapy? Transl. Androl. Urol. 2020, 9 (Suppl. S2), S135–S148. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.R.; Tan, M.S.; Xie, A.M.; Yu, J.T.; Tan, L. Heat shock protein 90 in Alzheimer’s disease. Biomed. Res. Int. 2014, 2014, 796869. [Google Scholar] [CrossRef]
- Bohush, A.; Bieganowski, P.; Filipek, A. Hsp90 and Its Co-Chaperones in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 4976. [Google Scholar] [CrossRef]
- Shelton, L.B.; Baker, J.D.; Zheng, D.; Sullivan, L.E.; Solanki, P.K.; Webster, J.M.; Sun, Z.; Sabbagh, J.J.; Nordhues, B.A.; Koren, J., 3rd; et al. Hsp90 activator Aha1 drives production of pathological tau aggregates. Proc. Natl. Acad. Sci. USA 2017, 114, 9707–9712. [Google Scholar] [CrossRef]
- Pedrazzoli, M.; Losurdo, M.; Paolone, G.; Medelin, M.; Jaupaj, L.; Cisterna, B.; Slanzi, A.; Malatesta, M.; Coco, S.; Buffelli, M. Glucocorticoid receptors modulate dendritic spine plasticity and microglia activity in an animal model of Alzheimer’s disease. Neurobiol. Dis. 2019, 132, 104568. [Google Scholar] [CrossRef]
- Canet, G.; Pineau, F.; Zussy, C.; Hernandez, C.; Hunt, H.; Chevallier, N.; Perrier, V.; Torrent, J.; Belanoff, J.K.; Meijer, O.C.; et al. Glucocorticoid receptors signaling impairment potentiates amyloid-β oligomers-induced pathology in an acute model of Alzheimer’s disease. FASEB J. 2020, 34, 1150–1168. [Google Scholar] [CrossRef]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform 2014, 6, 13. [Google Scholar] [CrossRef]
- Xu, H.Y.; Zhang, Y.Q.; Liu, Z.M.; Chen, T.; Lv, C.Y.; Tang, S.H.; Zhang, X.B.; Zhang, W.; Li, Z.Y.; Zhou, R.R.; et al. ETCM: An encyclopaedia of traditional Chinese medicine. Nucleic Acids Res. 2019, 47, D976–D982. [Google Scholar] [CrossRef]
- Yao, Z.J.; Dong, J.; Che, Y.J.; Zhu, M.F.; Wen, M.; Wang, N.N.; Wang, S.; Lu, A.P.; Cao, D.S. TargetNet: A web service for predicting potential drug-target interaction profiling via multi-target SAR models. J. Comput. Aided Mol. Des. 2016, 30, 413–424. [Google Scholar] [CrossRef] [PubMed]
- UniProt, C. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Yan, G.R.; He, Q.Y. DOSE: An R/Bioconductor package for disease ontology semantic and enrichment analysis. Bioinformatics 2015, 31, 608–609. [Google Scholar]
- Pinero, J.; Ramirez-Anguita, J.M.; Sauch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- The Gene Ontology, C. Expansion of the Gene Ontology knowledgebase and resources. Nucleic Acids Res. 2017, 45, D331–D338. [Google Scholar]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinformatics 2016, 54, 1301–13,033. [Google Scholar]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. Comparative Toxicogenomics Database (CTD): Update 2021. Nucleic Acids Res. 2021, 49, D1138–D1143. [Google Scholar] [CrossRef] [PubMed]
- Amberger, J.S.; Bocchini, C.A.; Schiettecatte, F.; Scott, A.F.; Hamosh, A. OMIM.org: Online Mendelian Inheritance in Man (OMIM(R)), an online catalog of human genes and genetic disorders. Nucleic Acids Res. 2015, 43, D789–D798. [Google Scholar] [PubMed]
- Gurbani, D.; Du, G.; Henning, N.J.; Rao, S.; Bera, A.K.; Zhang, T.; Gray, N.S.; Westover, K.D. Structure and Characterization of a Covalent Inhibitor of Src Kinase. Front. Mol. Biosci. 2020, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Do, H.T.; Li, H.; Chreifi, G.; Poulos, T.L.; Silverman, R.B. Optimization of Blood-Brain Barrier Permeability with Potent and Selective Human Neuronal Nitric Oxide Synthase Inhibitors Having a 2-Aminopyridine Scaffold. J. Med. Chem. 2019, 62, 2690–2707. [Google Scholar] [CrossRef]
- Hosfield, D.J.; Weber, S.; Li, N.S.; Sauvage, M.; Joiner, C.F.; Hancock, G.R.; Sullivan, E.A.; Ndukwe, E.; Han, R.; Cush, S.; et al. Stereospecific lasofoxifene derivatives reveal the interplay between estrogen receptor alpha stability and antagonistic activity in ESR1 mutant breast cancer cells. eLife 2022, 11, e72512. [Google Scholar] [CrossRef]
- Kreusch, A.; Han, S.; Brinker, A.; Zhou, V.; Choi, H.S.; He, Y.; Lesley, S.A.; Caldwell, J.; Gu, X.J. Crystal structures of human HSP90alpha-complexed with dihydroxyphenylpyrazoles. Bioorg. Med. Chem. Lett. 2005, 15, 1475–1478. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.Y.; Li, N.; Duan, J.J.; Jiang, B.; Lu, Z.; Ngu, K.; Tino, J.; Kopcho, L.M.; Lu, H.; Chen, J.; et al. Biologic-like In Vivo Efficacy with Small Molecule Inhibitors of TNFalpha Identified Using Scaffold Hopping and Structure-Based Drug Design Approaches. J. Med. Chem. 2020, 63, 15050–15071. [Google Scholar] [CrossRef]
- Rew, Y.; Du, X.; Eksterowicz, J.; Zhou, H.; Jahchan, N.; Zhu, L.; Yan, X.; Kawai, H.; McGee, L.R.; Medina, J.C.; et al. Discovery of a Potent and Selective Steroidal Glucocorticoid Receptor Antagonist (ORIC-101). J. Med. Chem. 2018, 61, 7767–7784. [Google Scholar] [CrossRef]
- Coleman, J.A.; Navratna, V.; Antermite, D.; Yang, D.; Bull, J.A.; Gouaux, E. Chemical and structural investigation of the paroxetine-human serotonin transporter complex. Elife 2020, 9, e56427. [Google Scholar] [CrossRef]
- El-Hachem, N.; Haibe-Kains, B.; Khalil, A.; Kobeissy, F.H.; Nemer, G. AutoDock and AutoDockTools for Protein-Ligand Docking: Beta-Site Amyloid Precursor Protein Cleaving Enzyme 1(BACE1) as a Case Study. Methods Mol. Biol. 2017, 1598, 391–403. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Compound Name | PubChem CID | MW |
|---|---|---|---|
| Quinolone Alkaloids | |||
| WZY1 | 1-methyl-2-nonyl-4-quinolone | 13967189 | 285.47 |
| WZY2 | 1-methyl-2-undecyl-4-quinolone | 5319811 | 313.53 |
| WZY3 | Evocarpine | 5317303 | 339.57 |
| WZY4 | 1-methyl-2-[(Z)-undec-6-enyl]-4-quinolone | 5319810 | 311.51 |
| WZY5 | 1-methyl-2-[(Z)-pentadec-10-enyl]-4-quinolone | 5319752 | 367.63 |
| WZY7 | 1-methyl-2-pentadecyl-4-quinolone | 5319753 | 369.65 |
| WZY20 | Hydroxyevodiamine | 71307457 | 319.39 |
| WZY23 | 1-methyl-2-[(Z)-5-undecenyl]-4(1H)-quinolone | 5319809 | 311.46 |
| WZY28 | Echinopsine | 6748 | 159.18 |
| WZY29 | Rhetsinine | 99652 | 319.36 |
| Indole Alkaloids | |||
| WZY11 | Evodiamine | 442088 | 303.39 |
| WZY12 | Evodiamide | 189454 | 307.43 |
| WZY13 | N-(2-methylaminobenzoyl) tryptamine | 5319506 | 293.4 |
| WZY16 | Rutaecarpine | 65752 | 287.34 |
| WZY17 | Dihydrorutaecarpine | N/A | 289.36 |
| WZY18 | Goshuyuamide I | 5317827 | 305.41 |
| WZY21 | Goshuyuamide II | 5317828 | 319.39 |
| WZY26 | 5-methoxy-N, N-dimethyltryptamine | 1832 | 218.29 |
| WZY27 | Dehydroevodiamine | 9817839 | 301.34 |
| WZY30 | 14-formyldihydrorutaecarpine | 5317369 | 317.34 |
| WZY32 | l-oxonoreleagnine | 87371 | 186.21 |
| Other Alkaloids | |||
| WZY14 | Fordimine | 5462442 | 256.38 |
| WZY15 | Evodione | 624052 | 292.36 |
| WZY19 | Berberine | 2353 | 336.39 |
| WZY22 | 2-hydroxy-3-formyl-7-methoxycarbazole | 189687 | 241.26 |
| WZY25 | Graveoline | 353825 | 279.29 |
| WZY31 | Kokusaginine | 10227 | 259.26 |
| Fatty acid/ester | |||
| WZY6 | Icosa-11,14,17-trienoic acid methyl ester | 5367326 | 320.57 |
| WZY24 | Goshuyic acid | 5312409 | 224.34 |
| Sterols | |||
| WZY8 | 24-methyl-31-norlanost-9(11)-enol | 5319735 | 428.82 |
| WZY9 | Beta-sitosterol | 86821 | 414.79 |
| WZY10 | Sitosterol | 222284 | 414.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Xu, J.; Guo, J.; Wang, Y.; Wang, Q. Elucidation of Pharmacological Mechanism Underlying the Anti-Alzheimer’s Disease Effects of Evodia rutaecarpa and Discovery of Novel Lead Molecules: An In Silico Study. Molecules 2023, 28, 5846. https://doi.org/10.3390/molecules28155846
Zhang L, Xu J, Guo J, Wang Y, Wang Q. Elucidation of Pharmacological Mechanism Underlying the Anti-Alzheimer’s Disease Effects of Evodia rutaecarpa and Discovery of Novel Lead Molecules: An In Silico Study. Molecules. 2023; 28(15):5846. https://doi.org/10.3390/molecules28155846
Chicago/Turabian StyleZhang, Lulu, Jia Xu, Jiejie Guo, Yun Wang, and Qinwen Wang. 2023. "Elucidation of Pharmacological Mechanism Underlying the Anti-Alzheimer’s Disease Effects of Evodia rutaecarpa and Discovery of Novel Lead Molecules: An In Silico Study" Molecules 28, no. 15: 5846. https://doi.org/10.3390/molecules28155846
APA StyleZhang, L., Xu, J., Guo, J., Wang, Y., & Wang, Q. (2023). Elucidation of Pharmacological Mechanism Underlying the Anti-Alzheimer’s Disease Effects of Evodia rutaecarpa and Discovery of Novel Lead Molecules: An In Silico Study. Molecules, 28(15), 5846. https://doi.org/10.3390/molecules28155846