
Coriandrum sativum and Its Utility in Psychiatric Disorders

 ,
,  ,
, 
 and
and
Abstract
1. Introduction
2. Methodology of Literature Research
3. Mental Disorders
3.1. Impact of Psychiatric Disorders on Society
3.2. Anxiety Disorders (AD)
3.3. Mood Disorders: Emphasis on Depression
3.4. Sleep Disorders
3.5. Epilepsy
4. Neurotransmitters Involved in Psychiatric Disorders
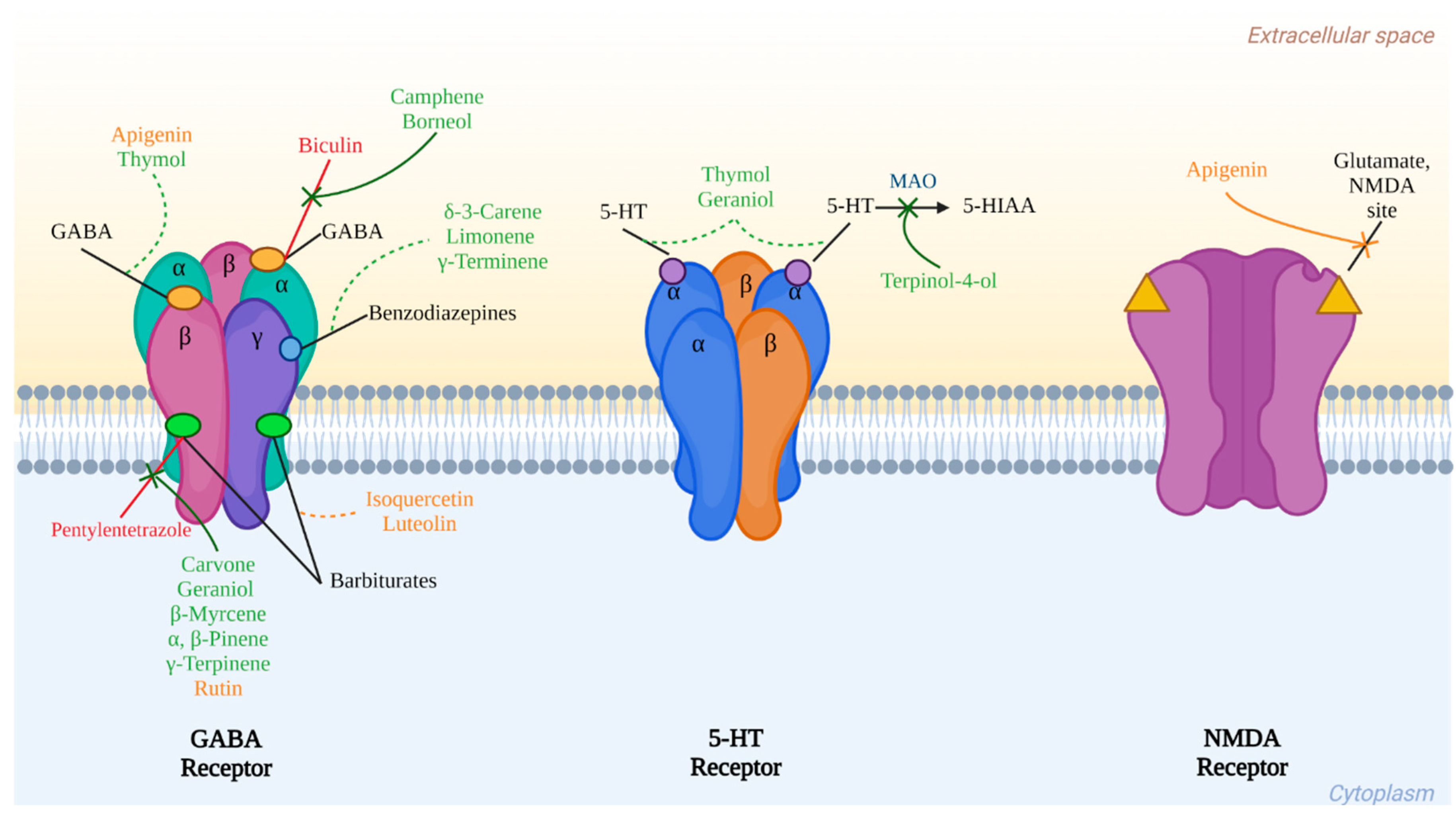
4.1. GABA and Glutamate
4.2. Serotonin: 5-hydroxytryptamine (5-HT)
5. Inflammation and Oxidative Stress Associated with AD, MDD, TS, and Epilepsy
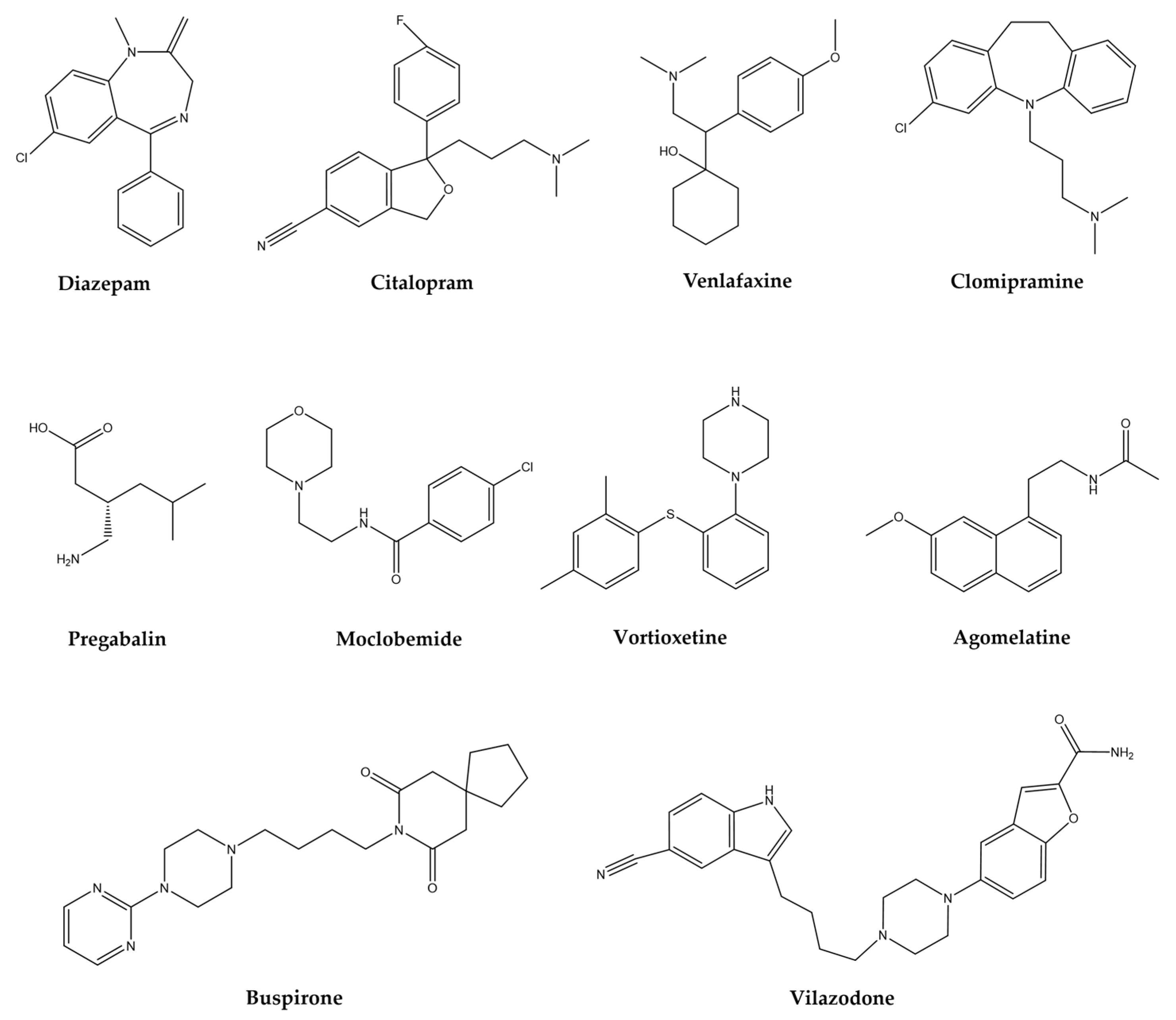
6. Treatments for AD, MDD, TS, and Epilepsy

{kind=link}
{kind=link}
| Group | Drugs | Mechanism of Action | Clinically | Reference | |||
|---|---|---|---|---|---|---|---|
| Anxiolytic | Antidepressant | Sedative | Antinociceptive | ||||
| BZD | Diazepam, clonazepam, alprazolam, chlordiazepoxide, clorazepate, lorazepam | GABAA receptor allosteric modulators | + | + | + | + | [58,59] |
| SSRIs | Citalopram, escitalopram, fluoxetine, fluvoxamine, paroxetine, sertraline | Selective serotonin reuptake inhibition | + | + | − | − | [60,61] |
| SNRIs | Duloxetine, venlafaxine | Serotonin and norepinephrine reuptake inhibition | + | + | − | + | [62] |
| TCAs | Clomipramine, imipramine, desipramine, nortriptyline | Reuptake of serotonin and norepinephrine blockage | − | + | − | + | [63,64] |
| CaM | Pregabalin | Decrease calcium influx | + | + | + | + | [65,66] |
| AZA | Buspirone | Acts on 5-HT-1A receptors for serotonin | + | − | − | − | [67,68] |
| MAOIs | Moclobemide | Reversible inhibitor of the enzyme MAO | + | + | − | − | [73] |
| SERTIs | Vortioxetine | Serotonin transporter inhibition | + | − | − | − | [69,70] |
| SERTIs | Vilazodone | SERT blocker; acts on 5-HT-1A receptors for serotonin | + | + | − | − | [70] |
| MMAs | Agomelatine | Melatonergic receptor agonist and 5-HT2C antagonist | + | + | − | − | [70] |
7. Coriandrum sativum L.
7.1. Botanical Description and Traditional Uses
7.2. Plant Ethnobotany on CNS Diseases in Mexico
8. Phytochemical Constituents of C. sativum and Its Activities in Psychiatric Disorders
8.1. Essential Oils
8.2. Extracts and Active Compounds on Nervous Disorders
9. Activities of Compounds from C. sativum over Mental Disorders
9.1. Essential Oil Terpenes
9.1.1. Linalool
9.1.2. Borneol
9.1.3. Camphene
9.1.4. δ-3-Carene
9.1.5. Carvone
9.1.6. Geraniol
9.1.7. Limonene
9.1.8. β-Myrcene
9.1.9. α- and β-Pinene
9.1.10. γ-Terminene
9.1.11. γ-Terpinene
9.1.12. Terpinen-4-ol
9.1.13. Thymol
9.2. Flavonoids
9.2.1. Apigenin
9.2.2. Daidzein
9.2.3. Isoquercetin
9.2.4. Luteolin
9.2.5. Pinocembrin
9.2.6. Quercetin
9.2.7. Rutin
| Phytochemical Group | Compound | Effect | Mechanism of Action | Reference |
|---|---|---|---|---|
| Terpenes | Borneol | Anticonvulsant and anxiolytic | GABAAR agonist | [100,101] |
| Camphene | Anxiolytic | GABAAR agonist | [100] | |
| δ-3-carene | Sleep-enhancing | Interaction with BZD receptors in GABAAR | [103] | |
| Carvone | Delay of induced seizure Sleep-enhancing | GABAAR agonist | [104] | |
| Geraniol | Delay of induced seizure Sedative and sleep-enhancing | GABAAR agonist Blocks 5-HT3A receptors | [106,107] | |
| Limonene | Antidepressant, anxiolytic and sedative | GABAAR agonist | [108,109,110] | |
| (−)-Linalool | Anticonvulsant and antidepressant | GluR interaction through 5-HT1A receptors | [97,99] | |
| β-Myrcene | Anticonvulsant, sedative, and sleep-enhancing | Metabolism interference of barbiturates by cytochrome P-450 enzyme activation | [111,112,113] | |
| α- and β- pinene | Anxiolytic and hypnotic Delay of induced seizure | Interaction in BZD receptors in GABAAR | [114,117] | |
| γ-Terminene | Sedative and sleep-enhancing | Interaction in BZD receptors in GABAAR | [118] | |
| γ-Terpinene | Sedative effect | Similar behavior as diazepam | [118] | |
| Thymol | Sedative effect | Partial agonist and positive modulator of 5-HT3A receptor | [118] | |
| Terpinen-4-ol | Antidepressant, sedative, sleep-enhancing Delay of induced seizure | GABAergic transmission modulation without involving BZD receptors | [104,119,120,121] | |
| Flavonoids | Apigenin | Anticonvulsant, antidepressant, anxiolytic, and sedative | GABAergic transmission modulation without involving BZD receptors Reduces alterations on the levels of 5-HT | [122,123,124,125,126,128,129] |
| Daidzein | Anxiolytic effect | --- | [129] | |
| Flavonoids | Isoquercetin | Antidepressant and sleep-enhancing | ---- | [130,131] |
| Luteolin | Antidepressant, anxiolytic, and sleep-enhancing | GABAergic transmission modulation without involving BZD receptors | [133,134] | |
| Pinocembrin | Antidepressant effect | ---- | [135] | |
| Quercetin | Anticonvulsant, antidepressant, anxiolytic, sleep-enhancing Memory improvement | Modulation of Glu receptor GABAA α5 receptors expression modulation Ionic inhibition current on 5-HT3R | [136,137,138,139,140] | |
| Rutin | Antidepressant, anxiolytic, and seizures severity attenuation Memory improvement | GABAergic transmission modulation without involving BZD receptors GABAA chloride channel blocking Serotonin availability increase in the synaptic cleft | [141,142,143,145,146] |
10. Metabolic Richness of C. sativum for the Design of Future Phytomedicine Based on Nanoparticles and Molecular Coupling Technologies
10.1. Molecular Docking
10.2. Nanoparticles
11. Discussion
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Mental Disorders. Available online: https://www.who.int/news-room/fact-sheets/detail/mental-disorders (accessed on 30 June 2021).
- World Health Organization. Mental Health Action Plan 2013–2020; World Health Organization: Geneva, Switzerland, 2013.
- Laribi, B.; Kouki, K.; M’Hamdi, M.; Bettaieb, T. Coriander (Coriandrum sativum L.) and its bioactive constituents. Fitoterapia 2015, 103, 9–26. [Google Scholar] [CrossRef]
- Instituto Nacional Indigenista. Cilantro. In Atlas de las Plantas de la Medicina Tradicional Mexicana Tomo I; Instituto Nacional Indigenista: Pátzcuaro, Mexico, 1994; pp. 443–444. [Google Scholar]
- Kasmaei, H.D.; Ghorbanifar, Z.; Zayeri, F.; Minaei, B.; Kamali, S.H.; Rezaeizadeh, H.; Amin, G.; Ghobadi, A.; Mirzaei, Z. Effects of coriandrum sativum syrup on migraine: A randomized, triple-blind, placebo-controlled trial. Iran. Red Crescent Med. J. 2016, 18, e20759. [Google Scholar] [CrossRef]
- Gastón, M.S.; Cid, M.P.; Vázquez, A.M.; Decarlini, M.F.; Demmel, G.I.; Rossi, L.I.; Aimar, M.L.; Salvatierra, N.A. Sedative effect of central administration of Coriandrum sativum essential oil and its major component linalool in neonatal chicks. Pharm. Biol. 2016, 54, 1954–1961. [Google Scholar] [CrossRef]
- Zenki, K.C.; de Souza, L.S.; Góis, A.M.; Lima, B.D.S.; Araújo, A.A.D.S.; Vieira, J.S.; Camargo, E.A.; Kalinine, E.; de Oliveira, D.L.; Walker, C.I.B. Coriandrum sativum Extract Prevents Alarm Substance-Induced Fear- and Anxiety-Like Responses in Adult Zebrafish. Zebrafish 2020, 17, 120–130. [Google Scholar] [CrossRef]
- Latha, K.; Rammohan, B.; Sunanda, B.P.V.; Uma Maheswari, M.S.; Mohan, S.K. Evaluation of anxiolytic activity of aqueous extract of Coriandrum sativum Linn. in mice: A preliminary experimental study. Pharmacogn. Res. 2015, 7, S47–S51. [Google Scholar] [CrossRef]
- Rakhshandeh, H.; Sadeghnia, H.R.; Ghorbani, A. Sleep-prolonging effect of Coriandrum sativum hydro-alcoholic extract in mice. Nat. Prod. Res. 2012, 26, 2095–2098. [Google Scholar] [CrossRef]
- Cioanca, O.; Hritcu, L.; Mihasan, M.; Trifan, A.; Hancianu, M. Inhalation of coriander volatile oil increased anxiolytic-antidepressant-like behaviors and decreased oxidative status in beta-amyloid (1–42) rat model of Alzheimer’s disease. Physiol. Behav. 2014, 131, 68–74. [Google Scholar] [CrossRef]
- Pourzaki, M.; Homayoun, M.; Sadeghi, S.; Seghatoleslam, M.; Hosseini, M.; Ebrahimzadeh Bideskan, A. Preventive effect of Coriandrum sativum on neuronal damages in pentylentetrazole-induced seizure in rats. Avicenna J. Phytomedicine 2017, 7, 116–128. [Google Scholar] [CrossRef]
- Anaeigoudari, A.; Hosseini, M.; Karami, R.; Vafaee, F.; Mohammadpour, T.; Ghorbani, A.; Sadeghnia, H.R. The effects of different fractions of Coriandrum sativum on pentylenetetrazole-induced seizures and brain tissues oxidative damage in rats. Avicenna J. Phytomedicine 2016, 6, 223–235. [Google Scholar] [CrossRef]
- Karami, R.; Hosseini, M.; Mohammadpour, T.; Ghorbani, A.; Sadeghnia, H.R.; Rakhshandeh, H.; Vafaee, F.; Esmaeilizadeh, M. Effects of hydroalcoholic extract of Coriandrum sativum on oxidative damage in pentylenetetrazole-induced seizures in rats. Iran. J. Neurol. 2015, 14, 59–66. [Google Scholar]
- World Health Organization. Health and Well-Being. Available online: https://www.who.int/data/gho/data/major-themes/health-and-well-being (accessed on 6 June 2021).
- World Health Organization. Salud Mental. Available online: https://www.who.int/es/news-room/facts-in-pictures/detail/mental-health (accessed on 30 June 2021).
- World Health Organization. Epilepsy: A Public Health Imperative: Summary; World Health Organization: Geneva, Switzerland, 2019.
- Bhaskar, S.; Hemavathy, D.; Prasad, S. Prevalence of chronic insomnia in adult patients and its correlation with medical comorbidities. J. Fam. Med. Prim. Care 2016, 5, 780. [Google Scholar] [CrossRef]
- Morrison, A.S.; Heimberg, R.G. Social anxiety and social anxiety disorder. Annu. Rev. Clin. Psychol. 2013, 9, 249–274. [Google Scholar] [CrossRef]
- Thibaut, F. Anxiety disorders: A review of current literature. Dialogues Clin. Neurosci. 2017, 19, 87–88. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 0-89042-555-8. [Google Scholar]
- James, M. DSM-5 Made Easy: The Clinician’s Guide to Diagnosis; The Guilford Press: New York, NY, USA, 2014. [Google Scholar]
- Kessler, R.C.; Ruscio, A.M.; Shear, K.; Wittchen, H.U. Epidemiology of Anxiety Disorders. Curr. Top. Behav. Neurosci. 2010, 2, 21–35. [Google Scholar]
- Baglioni, C.; Nanovska, S.; Regen, W.; Spiegelhalder, K.; Feige, B.; Nissen, C.; Reynolds, C.F.; Riemann, D. Sleep and mental disorders: A meta-analysis of polysomnographic research. Psychol. Bull. 2016, 142, 969–990. [Google Scholar] [CrossRef]
- Sateia, M.J. International classification of sleep disorders-third edition highlights and modifications. Chest 2014, 146, 1387–1394. [Google Scholar] [CrossRef]
- Rémi, J.; Pollmächer, T.; Spiegelhalder, K.; Trenkwalder, C.; Young, P. Sleep-Related Disorders in Neurology and Psychiatry. Dtsch. Arztebl. Int. 2019, 116, 681–688. [Google Scholar] [CrossRef]
- Sucher, N.J.; Carles, M.C. A pharmacological basis of herbal medicines for epilepsy. Epilepsy Behav. 2015, 52, 308–318. [Google Scholar] [CrossRef]
- Dika, H.; Nkola, R.; Iddi, S.; Magwiza, C.; Kongola, G. Limited care offered to people with epilepsy in Mwanza, Tanzania: Need for intervention. Pan Afr. Med. J. 2021, 38, 407. [Google Scholar] [CrossRef]
- Allen, M.J.; Sabir, S.; Sharma, S. GABA Receptor. Trends Pharmacol. Sci. 2023, 2, 62–64. [Google Scholar] [CrossRef]
- Köles, L.; Kató, E.; Hanuska, A.; Zádori, Z.S.; Al-Khrasani, M.; Zelles, T.; Rubini, P.; Illes, P. Modulation of excitatory neurotransmission by neuronal/glial signalling molecules: Interplay between purinergic and glutamatergic systems. Purinergic Signal. 2016, 12, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-S.; Huang, T.-H.; Lai, M.-C.; Huang, C.-W. The Role of Glutamate Receptors in Epilepsy. Biomedicines 2023, 11, 783. [Google Scholar] [CrossRef] [PubMed]
- Sarawagi, A.; Soni, N.D.; Patel, A.B. Glutamate and GABA Homeostasis and Neurometabolism in Major Depressive Disorder. Front. Psychiatry 2021, 12, 419. [Google Scholar] [CrossRef]
- Rudolph, U.; Knoflach, F. Beyond classical benzodiazepines: Novel therapeutic potential of GABA A receptor subtypes. Nat. Rev. Drug Discov. 2011, 10, 685–697. [Google Scholar] [CrossRef]
- Cryan, J.F.; Kaupmann, K. Don’t worry “B” happy!: A role for GABA B receptors in anxiety and depression. Trends Pharmacol. Sci. 2005, 26, 36–43. [Google Scholar] [CrossRef]
- Serretti, A.; Artioli, P.; De Ronchi, D. The 5-HT2C receptor as a target for mood disorders. Expert Opin. Ther. Targets 2004, 8, 15–23. [Google Scholar] [CrossRef]
- Hannon, J.; Hoyer, D. Molecular biology of 5-HT receptors. Behav. Brain Res. 2008, 195, 198–213. [Google Scholar] [CrossRef]
- Hen, R.; Nautiyal, K.M. Serotonin receptors in depression: From A to B. F1000Research 2017, 6, 123. [Google Scholar]
- Fakhfouri, G.; Rahimian, R.; Dyhrfjeld-Johnsen, J.; Zirak, M.R.; Beaulieu, J.M. 5-HT3 receptor antagonists in neurologic and neuropsychiatric disorders: The iceberg still lies beneath the surface. Pharmacol. Rev. 2019, 71, 383–412. [Google Scholar] [CrossRef]
- Yirmiya, R.; Goshen, I. Immune modulation of learning, memory, neural plasticity and neurogenesis. Brain. Behav. Immun. 2011, 25, 181–213. [Google Scholar] [CrossRef]
- Köhler, C.A.; Freitas, T.H.; Maes, M.; de Andrade, N.Q.; Liu, C.S.; Fernandes, B.S.; Stubbs, B.; Solmi, M.; Veronese, N.; Herrmann, N.; et al. Peripheral cytokine and chemokine alterations in depression: A meta-analysis of 82 studies. Acta Psychiatr. Scand. 2017, 135, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Michopoulos, V.; Powers, A.; Gillespie, C.F.; Ressler, K.J.; Jovanovic, T. Inflammation in Fear-and Anxiety-Based Disorders: PTSD, GAD, and beyond. Neuropsychopharmacology 2017, 42, 254–270. [Google Scholar] [CrossRef]
- Ichiyama, T.; Nishikawa, M.; Yoshitomi, T.; Hayashi, T.; Furukawa, S. Tumor necrosis factor-α, interleukin-1β and interleukin-6 in cerebrospinal fluid from children with prolonged febrile seizures: Comparison with acute encephalitis/encephalopathy. Neurology 1998, 50, 407–411. [Google Scholar] [CrossRef]
- Butler, T.; Li, Y.; Tsui, W.; Friedman, D.; Maoz, A.; Wang, X.; Harvey, P.; Tanzi, E.; Morim, S.; Kang, Y.; et al. Transient and chronic seizure-induced inflammation in human focal epilepsy. Epilepsia 2016, 57, e191–e194. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.R.; Opp, M.R. Sleep Health: Reciprocal Regulation of Sleep and Innate Immunity. Neuropsychopharmacology 2017, 42, 129–155. [Google Scholar] [CrossRef] [PubMed]
- Moldofsky, H.; Lue, F.A.; Davidson, J.R.; Gorczynski, R. Effects of sleep deprivation on human immune functions. FASEB J. 1989, 3, 1972–1977. [Google Scholar] [CrossRef]
- Stepanova, A.; Konrad, C.; Manfredi, G.; Springett, R.; Ten, V.; Galkin, A. The dependence of brain mitochondria reactive oxygen species production on oxygen level is linear, except when inhibited by antimycin A. J. Neurochem. 2019, 148, 731–745. [Google Scholar] [CrossRef]
- Souza, C.G.; Moreira, J.D.; Siqueira, I.R.; Pereira, A.G.; Rieger, D.K.; Souza, D.O.; Souza, T.M.; Portela, L.V.; Perry, M.L.S. Highly palatable diet consumption increases protein oxidation in rat frontal cortex and anxiety-like behavior. Life Sci. 2007, 81, 198–203. [Google Scholar] [CrossRef]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar] [CrossRef]
- Fedoce, A.d.G.; Ferreira, F.; Bota, R.G.; Bonet-Costa, V.; Sun, P.Y.; Davies, K.J.A. The role of oxidative stress in anxiety disorder: Cause or consequence? Free Radic. Res. 2018, 52, 737–750. [Google Scholar] [CrossRef]
- Liang, L.P.; Patel, M. Seizure-induced changes in mitochondrial redox status. Free Radic. Biol. Med. 2006, 40, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Ingold, I.; Berndt, C.; Schmitt, S.; Doll, S.; Poschmann, G.; Buday, K.; Roveri, A.; Peng, X.; Porto Freitas, F.; Seibt, T.; et al. Selenium Utilization by GPX4 Is Required to Prevent Hydroperoxide-Induced Ferroptosis. Cell 2018, 172, 409–422.e21. [Google Scholar] [CrossRef]
- Reimund, E. The free radical flux theory of sleep. Med. Hypotheses 1994, 43, 231–233. [Google Scholar] [CrossRef] [PubMed]
- D’Almeida, V.; Lobo, L.L.; Hipólide, D.C.; De Oliveira, A.C.; Nobrega, J.N.; Tufik, S. Sleep deprivation induces brain region-specific decreases in glutathione levels. Neuroreport 1998, 9, 2853–2856. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, L.; Gulyani, S.; Nienhuis, R.; Siegel, J.M. Sleep deprivation decreases superoxide dismutase activity in rat hippocampus and brainstem. Neuroreport 2002, 13, 1387–1390. [Google Scholar] [CrossRef]
- D’Almeida, V.; Hipólide, D.C.; Azzalis, L.A.; Lobo, L.L.; Junqueira, V.B.C.; Tufik, S. Absence of oxidative stress following paradoxical sleep deprivation in rats. Neurosci. Lett. 1997, 235, 25–28. [Google Scholar] [CrossRef]
- Kempf, A.; Song, S.M.; Talbot, C.B.; Miesenböck, G. A potassium channel β-subunit couples mitochondrial electron transport to sleep. Nature 2019, 568, 230–234. [Google Scholar] [CrossRef]
- Vaccaro, A.; Kaplan Dor, Y.; Nambara, K.; Pollina, E.A.; Lin, C.; Greenberg, M.E.; Rogulja, D. Sleep Loss Can Cause Death through Accumulation of Reactive Oxygen Species in the Gut. Cell 2020, 181, 1307–1328.e15. [Google Scholar] [CrossRef]
- Pandey, A.; Kar, S.K. Rapid Eye Movement sleep deprivation of rat generates ROS in the hepatocytes and makes them more susceptible to oxidative stress. Sleep Sci. 2018, 11, 245–253. [Google Scholar] [CrossRef]
- Edwards, Z.; Preuss, C.V. GABA Receptor Positive Allosteric Modulators; StatPearls Publishing: Tampa, FL, USA, 2020. [Google Scholar]
- Griffin, C.E.; Kaye, A.M.; Rivera Bueno, F.; Kaye, A.D. Benzodiazepine pharmacology and central nervous system-mediated effects. Ochsner J. 2013, 13, 214–223. [Google Scholar]
- Ramic, E.; Prasko, S.; Gavran, L.; Spahic, E. Assessment of the Antidepressant Side Effects Occurrence in Patients Treated in Primary Care. Mater. Socio Medica 2020, 32, 131. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.M. SSRI antidepressant medications: Adverse effects and tolerability. Prim. Care Companion J. Clin. Psychiatry 2001, 3, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Santarsieri, D.; Schwartz, T.L. Antidepressant efficacy and side-effect burden: A quick guide for clinicians. Drugs Context 2015, 4, 212290. [Google Scholar] [CrossRef] [PubMed]
- Cookson, J. Side-effects of antidepressants. Br. J. Psychiatry 1993, 163, 20–24. [Google Scholar] [CrossRef]
- Moncrieff, J. Persistent adverse effects of antidepressants. Epidemiol. Psychiatr. Sci. 2019, 29, e56. [Google Scholar] [CrossRef]
- Zaccara, G.; Gangemi, P.; Perucca, P.; Specchio, L. The adverse event profile of pregabalin: A systematic review and meta-analysis of randomized controlled trials. Epilepsia 2011, 52, 826–836. [Google Scholar] [CrossRef]
- Toth, C. Pregabalin: Latest safety evidence and clinical implications for the management of neuropathic pain. Ther. Adv. Drug Saf. 2014, 5, 38–56. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.P. Buspirone, a new approach to the treatment of anxiety. FASEB J. 1988, 2, 2445–2452. [Google Scholar] [CrossRef]
- Bandelow, B.; Michaelis, S.; Wedekind, D. Treatment of anxiety disorders. Dialogues Clin. Neurosci. 2017, 19, 93–107. [Google Scholar] [CrossRef]
- Long, J.A.D. Vortioxetine for Depression in Adults. Issues Ment. Health Nurs. 2019, 40, 819–820. [Google Scholar] [CrossRef]
- Koesters, M.; Ostuzzi, G.; Guaiana, G.; Breilmann, J.; Barbui, C. Vortioxetine for depression in adults. Cochrane Database Syst. Rev. 2017, 7, CD011520. [Google Scholar] [CrossRef] [PubMed]
- Badgujar, S.B.; Patel, V.V.; Bandivdekar, A.H. Foeniculum vulgare Mill: A review of its botany, phytochemistry, pharmacology, contemporary application, and toxicology. Biomed Res. Int. 2014, 2014, 842674. [Google Scholar] [CrossRef]
- Mara De Menezes Epifanio, N.; Rykiel Iglesias Cavalcanti, L.; Falcão Dos Santos, K.; Soares Coutinho Duarte, P.; Kachlicki, P.; Ozarowski, M.; Jorge Riger, C.; Siqueira De Almeida Chaves, D. Chemical characterization and in vivo antioxidant activity of parsley (Petroselinum crispum) aqueous extract. Food Funct. 2020, 11, 5346–5356. [Google Scholar] [CrossRef] [PubMed]
- Zwanzger, P. Treatment of anxiety disorders. MMW-Fortschr. Med. 2018, 160, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Servicio de Información Agroalimentaria y Pesquera México Exportó 64 mil 647 Toneladas de Cilantro, en 2017. Available online: https://www.gob.mx/siap/es/articulos/mexico-exporto-64-mil-647-toneladas-de-cilantro-en-2017?idiom=es (accessed on 1 July 2021).
- Ishikawa, T.; Kondo, K.; Kitajima, J. Water-soluble constituents of coriander. Chem. Pharm. Bull. 2003, 51, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Flores y Escalante, J. Breve Historia de la Comida Mexicana; Random House Mondadori: Ciudad de México, Mexico, 1994. [Google Scholar]
- Sahib, N.G.; Anwar, F.; Gilani, A.H.; Hamid, A.A.; Saari, N.; Alkharfy, K.M. Coriander (Coriandrum sativum L.): A potential source of high-value components for functional foods and nutraceuticals–A review. Phyther. Res. 2013, 27, 1439–1456. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Mandal, M. Coriander (Coriandrum sativum L.) essential oil: Chemistry and biological activity. Asian Pac. J. Trop. Biomed. 2015, 5, 421–428. [Google Scholar] [CrossRef]
- Silva, F.; Ferreira, S.; Queiroz, J.A.; Domingues, F.C. Coriander (Coriandrum sativum L.) essential oil: Its antibacterial activity and mode of action evaluated by flow cytometry. J. Med. Microbiol. 2011, 60, 1479–1486. [Google Scholar] [CrossRef]
- Can, E.; Kızak, V.; Can, Ş.S.; Özçiçek, E. Anesthetic Efficiency of Three Medicinal Plant Oils for Aquatic Species: Coriander Coriandrum sativum, Linaloe Tree Bursera delpechiana, and Lavender Lavandula hybrida. J. Aquat. Anim. Health 2019, 31, 266–273. [Google Scholar] [CrossRef]
- Caputo, L.; Souza, L.F.; Alloisio, S.; Cornara, L.; De Feo, V. Coriandrum sativum and Lavandula angustifolia essential oils: Chemical composition and activity on central nervous system. Int. J. Mol. Sci. 2016, 17, 1999. [Google Scholar] [CrossRef]
- Kazempor, S.F.; Langehbiz, S.V.; Hosseini, M.; Shafei, M.N.; Ghorbani, A.; Pourganji, M. The analgesic effects of different extracts of aerial parts of Coriandrum Sativum in mice. Int. J. Biomed. Sci. 2015, 11, 23–28. [Google Scholar]
- Heldwein, C.G.; Silva, L.d.L.; Gai, E.Z.; Roman, C.; Parodi, T.V.; E Bürger, M.; Baldisserotto, B.; Flores, É.M.d.M.; Heinzmann, B.M. S-(+)-Linalool from Lippia alba: Sedative and anesthetic for silver catfish (Rhamdia quelen). Vet. Anaesth. Analg. 2014, 41, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, K.E.; de Oliveira, F.R.; Barbosa, B.R.C.; Paraense, R.S.O.; Bannwart, C.M.; Pinheiro, B.G.; Botelho, A.d.S.; Muto, N.A.; do Amarante, C.B.; Hamoy, M.; et al. Aqueous Coriandrum sativum L. extract promotes neuroprotection against motor changes and oxidative damage in rat progeny after maternal exposure to methylmercury. Food Chem. Toxicol. 2019, 133, 110755. [Google Scholar] [CrossRef]
- Velaga, M.K.; Yallapragada, P.R.; Williams, D.; Rajanna, S.; Bettaiya, R. Hydroalcoholic seed extract of Coriandrum sativum (coriander) alleviates lead-induced oxidative stress in different regions of rat brain. Biol. Trace Elem. Res. 2014, 159, 351–363. [Google Scholar] [CrossRef]
- Gallo, M.; Ferracane, R.; Graziani, G.; Ritieni, A.; Fogliano, V. Microwave assisted extraction of phenolic compounds from four different spices. Molecules 2010, 15, 6365–6374. [Google Scholar] [CrossRef] [PubMed]
- Rahiman, S.; Tantry, B.A.; Kumar, A. Variation of antioxidant activity and phenolic content of some common home remedies with storage time. Afr. J. Tradit. Complement. Altern. Med. 2012, 10, 124–127. [Google Scholar] [CrossRef][Green Version]
- Wu, T.T.; Tsai, C.W.; Yao, H.T.; Lii, C.K.; Chen, H.W.; Wu, Y.L.; Chen, P.Y.; Liu, K.L. Suppressive effects of extracts from the aerial part of Coriandrum sativum L. on LPS-induced inflammatory responses inmurine RAW 264.7 macrophages. J. Sci. Food Agric. 2010, 90, 1846–1854. [Google Scholar] [CrossRef]
- Manville, R.W.; Abbott, G.W. Cilantro leaf harbors a potent potassium channel–activating anticonvulsant. FASEB J. 2019, 33, 11349–11363. [Google Scholar] [CrossRef]
- Zhang, C.R.; Dissanayake, A.A.; Kevseroǧlu, K.; Nair, M.G. Evaluation of coriander spice as a functional food by using in vitro bioassays. Food Chem. 2015, 167, 24–29. [Google Scholar] [CrossRef]
- Kozłowska, M.; Gruczyńska, E.; Ścibisz, I.; Rudzińska, M. Fatty acids and sterols composition, and antioxidant activity of oils extracted from plant seeds. Food Chem. 2016, 213, 450–456. [Google Scholar] [CrossRef]
- Begnami, A.F.; Spindola, H.M.; Ruiz, A.L.T.G.; de Carvalho, J.E.; Groppo, F.C.; Rehder, V.L.G. Antinociceptive and anti-edema properties of the ethyl acetate fraction obtained from extracts of Coriandrum sativum Linn. leaves. Biomed. Pharmacother. 2018, 103, 1617–1622. [Google Scholar] [CrossRef]
- Puthusseri, B.; Divya, P.; Lokesh, V.; Neelwarne, B. Salicylic acid-induced elicitation of folates in coriander (Coriandrum sativum L.) improves bioaccessibility and reduces pro-oxidant status. Food Chem. 2013, 136, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Emamghoreishi, M.; Khasaki, M.; Aazam, M.F. Coriandrum sativum: Evaluation of its anxiolytic effect in the elevated plus-maze. J. Ethnopharmacol. 2005, 96, 365–370. [Google Scholar] [CrossRef]
- Kunzemann, J.; Herrmann, K. Isolation and identification of flavon(ol)-O-glycosides in caraway (Carum carvi L.), fennel (Foeniculum vulgare Mill.), anise (Pimpinella anisum L.), and coriander (Coriandrum sativum L.), and of flavon-C-glycosides in anise. I. Phenolics of spices (author’s transl). Z. Lebensm. Unters. Forsch. 1977, 164, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Hussain, F.; Jahan, N.; Rahman, K.-U.; Sultana, B.; Jamil, S. Identification of hypotensive biofunctional compounds of Coriandrum sativum and evaluation of their Angiotensin-Converting Enzyme (ACE) inhibition potential. Oxidative Med. Cell. Longev. 2018, 2018, 4643736. [Google Scholar] [CrossRef] [PubMed]
- Elisabetsky, E.; Brum, L.F.S.; Souza, D.O. Anticonvulsant properties of linalool in glutamate-related seizure models. Phytomedicine 1999, 6, 107–113. [Google Scholar] [CrossRef]
- Coelho, V.; Mazzardo-Martins, L.; Martins, D.F.; Santos, A.R.S.; Da Silva Brum, L.F.; Picada, J.N.; Pereira, P. Neurobehavioral and genotoxic evaluation of (-)-linalool in mice. J. Nat. Med. 2013, 67, 876–880. [Google Scholar] [CrossRef]
- Guzmán-Gutiérrez, S.L.; Bonilla-Jaime, H.; Gómez-Cansino, R.; Reyes-Chilpa, R. Linalool and β-pinene exert their antidepressant-like activity through the monoaminergic pathway. Life Sci. 2015, 128, 24–29. [Google Scholar] [CrossRef]
- Umezu, T.; Nagano, K.; Ito, H.; Kosakai, K.; Sakaniwa, M.; Morita, M. Anticonflict effects of lavender oil and identification of its active constituents. Pharmacol. Biochem. Behav. 2006, 85, 713–721. [Google Scholar] [CrossRef]
- Cao, B.; Ni, H.Y.; Li, J.; Zhou, Y.; Bian, X.L.; Tao, Y.; Cai, C.Y.; Qin, C.; Wu, H.Y.; Chang, L.; et al. (+)-Borneol suppresses conditioned fear recall and anxiety-like behaviors in mice. Biochem. Biophys. Res. Commun. 2018, 495, 1588–1593. [Google Scholar] [CrossRef]
- Tambe, R.; Jain, P.; Patil, S.; Ghumatkar, P.; Sathaye, S. Antiepileptogenic effects of borneol in pentylenetetrazole-induced kindling in mice. Naunyn-Schmiedeb. Arch. Pharmacol. 2016, 389, 467–475. [Google Scholar] [CrossRef]
- Woo, J.; Yang, H.; Yoon, M.; Gadhe, C.G.; Pae, A.N.; Cho, S.; Justin Lee, C. 3-Carene, a phytoncide from pine tree has a sleep-enhancing effect by targeting the GABAA-benzodiazepine receptors. Exp. Neurobiol. 2019, 28, 593–601. [Google Scholar] [CrossRef]
- De Sousa, D.P.; Nóbrega, F.F.F.; De Morais, L.C.S.L.; De Almeida, R.N. Evaluation of the anticonvulsant activity of terpinen-4-ol. Z. Naturforsch.-Sect. C J. Biosci. 2009, 64, 1–5. [Google Scholar] [CrossRef]
- Sánchez-Borzone, M.; Delgado-Marín, L.; García, D.A. Inhibitory effects of carvone isomers on the GABAA receptor in primary cultures of rat cortical neurons. Chirality 2014, 26, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, K.A.A.L.; dos Santos, J.R.; Melo, T.C.d.S.; de Souza, M.F.; Santos, L.d.G.; de Gois, A.M.; Cintra, R.R.; Lins, L.C.R.F.; Ribeiro, A.M.; Marchioro, M. Depressant effect of geraniol on the central nervous system of rats: Behavior and ECoG power spectra. Biomed. J. 2018, 41, 298–305. [Google Scholar] [CrossRef]
- Ziemba, P.M.; Schreiner, B.S.P.; Flegel, C.; Herbrechter, R.; Stark, T.D.; Hofmann, T.; Hatt, H.; Werner, M.; Gisselmann, G. Activation and modulation of recombinantly expressed serotonin receptor type 3A by terpenes and pungent substances. Biochem. Biophys. Res. Commun. 2015, 467, 1090–1096. [Google Scholar] [CrossRef]
- De Almeida, A.A.C.; Costa, J.P.; De Carvalho, R.B.F.; De Sousa, D.P.; De Freitas, R.M. Evaluation of acute toxicity of a natural compound (+)-limonene epoxide and its anxiolytic-like action. Brain Res. 2012, 1448, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Lima, N.G.P.B.; De Sousa, D.P.; Pimenta, F.C.F.; Alves, M.F.; De Souza, F.S.; MacEdo, R.O.; Cardoso, R.B.; De Morais, L.C.S.L.; Melo Diniz, M.D.F.F.; De Almeida, R.N. Anxiolytic-like activity and GC-MS analysis of (R)-(+)-limonene fragrance, a natural compound found in foods and plants. Pharmacol. Biochem. Behav. 2013, 103, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Yang, Z.Y.; Fan, G.; Ren, J.N.; Yin, K.J.; Pan, S.Y. Antidepressant-like Effect of Citrus sinensis (L.) Osbeck Essential Oil and Its Main Component Limonene on Mice. J. Agric. Food Chem. 2019, 67, 13817–13828. [Google Scholar] [CrossRef]
- Freitas, J.C.B.R.; Presgrave, O.A.F.; Fingola, F.F.; Menezes, M.A.C.; Paumgartten, F.J.R. Effect of β-myrcene on pentobarbital sleeping time. Braz. J. Med. Biol. Res. 1993, 26, 519–523. [Google Scholar]
- Do Vale, T.G.; Furtado, E.C.; Santos, J.G.; Viana, G.S.B. Central effects of citral, myrcene and limonene, constituents of essential oil chemotypes from Lippia alba (mill.) N.E. Brown. Phytomedicine 2002, 9, 709–714. [Google Scholar] [CrossRef] [PubMed]
- De Barros Viana, G.S.; Do Vale, T.G.; Silva, C.M.M.; De Abreu Matos, F.J. Anticonvulsant activity of essential oils and active principles from chemotypes of Lippia alba (Mill.) N.E. Brown. Biol. Pharm. Bull. 2000, 23, 1314–1317. [Google Scholar] [CrossRef]
- Yang, H.; Woo, J.; Pae, A.N.; Um, M.Y.; Cho, N.C.; Park, K.D.; Yoon, M.; Kim, J.; Lee, C.J.; Cho, S. α-pinene, a major constituent of pine tree oils, enhances non-rapid eye movement sleep in mice through GABAA-benzodiazepine receptors. Mol. Pharmacol. 2016, 90, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Kasuya, H.; Okada, N.; Kubohara, M.; Satou, T.; Masuo, Y.; Koike, K. Expression of BDNF and TH mRNA in the brain following inhaled administration of α-Pinene. Phyther. Res. 2015, 29, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Zamyad, M.; Abbasnejad, M.; Esmaeili-Mahani, S.; Mostafavi, A.; Sheibani, V. The anticonvulsant effects of ducrosia anethifolia (boiss) essential oil are produced by its main component alpha-pinene in rats. Arq. Neuropsiquiatr. 2019, 77, 106–114. [Google Scholar] [CrossRef]
- Felipe, C.F.B.; Albuquerque, A.M.S.; de Pontes, J.L.X.; de Melo, J.Í.V.; Rodrigues, T.C.M.L.; de Sousa, A.M.P.; Monteiro, Á.B.; Ribeiro, A.E.d.S.; Lopes, J.P.; de Menezes, I.R.A.; et al. Comparative study of alpha- and beta-pinene effect on PTZ-induced convulsions in mice. Fundam. Clin. Pharmacol. 2019, 33, 181–190. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Takemoto, H.; Fu, Z.; Shimizu, E.; Kinjo, Y. Enhancement of pentobarbital-induced sleep by the vaporized essential oil of citrus keraji var. Kabuchii and its characteristic component, γ-Terpinene. Nat. Prod. Commun. 2016, 11, 1175–1178. [Google Scholar] [CrossRef]
- Ito, K.; Ito, M. The sedative effect of inhaled terpinolene in mice and its structure-activity relationships. J. Nat. Med. 2013, 67, 833–837. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Koch, W.; Czernicka, L.; Głowniak, K.; Asakawa, Y.; Umeyama, A.; Marzec, Z.; Kuzuhara, T. MAO-A inhibitory potential of terpene constituents from ginger rhizomes—A bioactivity guided fractionation. Molecules 2018, 23, 1301. [Google Scholar] [CrossRef]
- Nóbrega, F.F.F.; Salvadori, M.G.S.S.; Masson, C.J.; Mello, C.F.; Nascimento, T.S.; Leal-Cardoso, J.H.; De Sousa, D.P.; Almeida, R.N. Monoterpenoid terpinen-4-ol exhibits anticonvulsant activity in behavioural and electrophysiological studies. Oxidative Med. Cell. Longev. 2014, 2014, 703848. [Google Scholar] [CrossRef]
- Viola, H.; Wasowski, C.; Levi De Stein, M.; Wolfman, C.; Silveira, R.; Dajas, F.; Medina, J.H.; Paladini, A.C. Apigenin, a component of Matricaria recutita flowers, is a central benzodiazepine receptors-ligand with anxiolytic effects. Planta Med. 1995, 61, 213–216. [Google Scholar] [CrossRef]
- Zanoli, P.; Avallone, R.; Baraldi, M. Behavioral characterisation of the flavonoids apigenin and chrysin. Fitoterapia 2000, 71, S117–S123. [Google Scholar] [CrossRef]
- Campbell, E.L.; Chebib, M.; Johnston, G.A.R. The dietary flavonoids apigenin and (-)-epigallocatechin gallate enhance the positive modulation by diazepam of the activation by GABA of recombinant GABA A receptors. Biochem. Pharmacol. 2004, 68, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Losi, G.; Puia, G.; Garzon, G.; De Vuono, M.C.; Baraldi, M. Apigenin modulates GABAergic and glutamatergic transmission in cultured cortical neurons. Eur. J. Pharmacol. 2004, 502, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, T.; Yasuda, T.; Ueda, J.; Ohsawa, K. Antidepressant-like effects of apigenin and 2,4,5-trimethoxycinnamic acid from Perilla frutescens in the forced swimming test. Biol. Pharm. Bull. 2003, 26, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.; Guo, X.; Li, Y.; Yang, X.; Han, Y. Apigenin reverses depression-like behavior induced by chronic corticosterone treatment in mice. Eur. J. Pharmacol. 2016, 774, 50–54. [Google Scholar] [CrossRef]
- Yi, L.T.; Li, J.M.; Li, Y.C.; Pan, Y.; Xu, Q.; Kong, L.D. Antidepressant-like behavioral and neurochemical effects of the citrus-associated chemical apigenin. Life Sci. 2008, 82, 741–751. [Google Scholar] [CrossRef]
- Zeng, S.; Tai, F.; Zhai, P.; Yuan, A.; Jia, R.; Zhang, X. Effect of daidzein on anxiety, social behavior and spatial learning in male Balb/cJ mice. Pharmacol. Biochem. Behav. 2010, 96, 16–23. [Google Scholar] [CrossRef]
- Kang, T.H.; Jeong, S.J.; Kim, N.Y.; Higuchi, R.; Kim, Y.C. Sedative activity of two flavonol glycosides isolated from the flowers of Albizzia julibrissin Durazz. J. Ethnopharmacol. 2000, 71, 321–323. [Google Scholar] [CrossRef]
- Butterweck, V.; Jürgenliemk, G.; Nahrstedt, A.; Winterhoff, H. Flavonoids from Hypericum perforatum show antidepressant activity in the forced swimming test. Planta Med. 2000, 66, 3–6. [Google Scholar] [CrossRef]
- Raines, T.; Jones, P.; Moe, N.; Duncan, R.; McCall, S.; Ceremuga, T.E. Investigation of the Anxiolytic Effects of Luteolin, a Lemon Balm Flavonoid in the Male Sprague-Dawley Rat. AANA J. 2009, 77, 33–36. [Google Scholar] [PubMed]
- Coleta, M.; Campos, M.G.; Cotrim, M.D.; de Lima, T.C.M.; Cunha, A.P. da Assessment of luteolin (3′,4′,5,7-tetrahydroxyflavone) neuropharmacological activity. Behav. Brain Res. 2008, 189, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Ishisaka, M.; Kakefuda, K.; Yamauchi, M.; Tsuruma, K.; Shimazawa, M.; Tsuruta, A.; Hara, H. Luteolin shows an antidepressant-like effect via suppressing endoplasmic reticulum stress. Biol. Pharm. Bull. 2011, 34, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zheng, L.; Xu, L.; Tu, J.; Gu, X. Pinocembrin mitigates depressive-like behaviors induced by chronic unpredictable mild stress through ameliorating neuroinflammation and apoptosis. Mol. Med. 2020, 26, 53. [Google Scholar] [CrossRef]
- Kosari-Nasab, M.; Shokouhi, G.; Ghorbanihaghjo, A.; Mesgari-Abbasi, M.; Salari, A.A. Quercetin mitigates anxiety-like behavior and normalizes hypothalamus-pituitary-adrenal axis function in a mouse model of mild traumatic brain injury. Behav. Pharmacol. 2019, 30, 282–289. [Google Scholar] [CrossRef]
- Samad, N.; Saleem, A.; Yasmin, F.; Shehzad, M.A. Quercetin protects against stress-induced anxiety- and depression- like behavior and improves memory in male mice. Physiol. Res. 2018, 67, 795–808. [Google Scholar] [CrossRef]
- Nassiri-Asl, M.; Hajiali, F.; Taghiloo, M.; Abbasi, E.; Mohseni, F.; Yousefi, F. Comparison between the effects of quercetin on seizure threshold in acute and chronic seizure models. Toxicol. Ind. Health 2016, 32, 936–944. [Google Scholar] [CrossRef]
- Moghbelinejad, S.; Alizadeh, S.; Mohammadi, G.; Khodabandehloo, F.; Rashvand, Z.; Najafipour, R.; Nassiri-Asl, M. The effects of quercetin on the gene expression of the GABAA receptor α5 subunit gene in a mouse model of kainic acid-induced seizure. J. Physiol. Sci. 2017, 67, 339–343. [Google Scholar] [CrossRef]
- Lee, B.H.; Jeong, S.M.; Lee, J.H.; Kim, J.H.; Yoon, I.S.; Lee, J.H.; Choi, S.H.; Lee, S.M.; Chang, C.G.; Kim, H.C.; et al. Erratum: Quercetin inhibits the 5-hydroxytryptamine type 3 receptor-mediated ion current by interacting with pre-transmembrane domain I (Molecules and Cells 20, 1 (69–76)). Mol. Cells 2005, 20, 452. [Google Scholar]
- Nöldner, M.; Schötz, K. Rutin is essential for the antidepressant activity of Hypericum perforatum extracts in the forced swimming test. Planta Med. 2002, 68, 577–580. [Google Scholar] [CrossRef]
- Hernandez-Leon, A.; González-Trujano, M.E.; Fernández-Guasti, A. The anxiolytic-like effect of rutin in rats involves GABA A receptors in the basolateral amygdala. Behav. Pharmacol. 2017, 28, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Yusha’u, Y.; Muhammad, U.A.; Nze, M.; Egwuma, J.M.; Igomu, O.J.; Abdulkadir, M. Modulatory role of rutin supplement on Open Space Forced Swim Test Murine model of depression. Niger. J. Physiol. Sci. 2017, 32, 201–205. [Google Scholar] [PubMed]
- Du, B.; Tang, X.; Liu, F.; Zhang, C.; Zhao, G.; Ren, F.; Leng, X. Antidepressant-like effects of the hydroalcoholic extracts of Hemerocallis Citrina and its potential active components. BMC Complement. Altern. Med. 2014, 14, 326. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.G.; Bettio, L.E.B.; Cunha, M.P.; Santos, A.R.S.; Pizzolatti, M.G.; Brighente, I.M.C.; Rodrigues, A.L.S. Antidepressant-like effect of rutin isolated from the ethanolic extract from Schinus molle L. in mice: Evidence for the involvement of the serotonergic and noradrenergic systems. Eur. J. Pharmacol. 2008, 587, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Nassiri-Asl, M.; Mortazavi, S.R.; Samiee-Rad, F.; Zangivand, A.A.; Safdari, F.; Saroukhani, S.; Abbasi, E. The effects of rutin on the development of pentylenetetrazole kindling and memory retrieval in rats. Epilepsy Behav. 2010, 18, 50–53. [Google Scholar] [CrossRef]
- Mechchate, H.; de Oliveira, R.C.; Es-Safi, I.; Vasconcelos Mourão, E.M.; Bouhrim, M.; Kyrylchuk, A.; Soares Pontes, G.; Bousta, D.; Grafov, A. Antileukemic activity and molecular docking study of a polyphenolic extract from coriander seeds. Pharmaceuticals 2021, 14, 770. [Google Scholar] [CrossRef]
- Kajal, A.; Singh, R. Coriandrum sativum improve neuronal function via inhibition of oxidative/nitrosative stress and TNF-α in diabetic neuropathic rats. J. Ethnopharmacol. 2020, 263, 112959. [Google Scholar] [CrossRef]
- Kajal, A.; Singh, R. Coriandrum sativum seeds extract mitigate progression of diabetic nephropathy in experimental rats via AGEs inhibition. PLoS ONE 2019, 14, e0213147. [Google Scholar] [CrossRef] [PubMed]
- Kajal, A.; Singh, R. An allied approach for in vitro modulation of aldose reductase, sorbitol accumulation and advanced glycation end products by flavonoid rich extract of Coriandrum sativum L. seeds. Toxicol. Rep. 2018, 5, 800–807. [Google Scholar] [CrossRef]
- Ferrari, M. Cancer nanotechnology: Opportunities and challenges. Nat. Rev. Cancer 2005, 5, 161–171. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, X.; Chen, J. hua Biofabrication of AuNPs using Coriandrum sativum leaf extract and their antioxidant, analgesic activity. Sci. Total Environ. 2021, 767, 144914. [Google Scholar] [CrossRef] [PubMed]
- Alquraidi, A.O.; Mosa, K.A.; Ramamoorthy, K. Phytotoxic and genotoxic effects of copper nanoparticles in coriander (Coriandrum sativum—Apiaceae). Plants 2019, 8, 19. [Google Scholar] [CrossRef]
- Fatemi, H.; Esmaiel Pour, B.; Rizwan, M. Foliar application of silicon nanoparticles affected the growth, vitamin C, flavonoid, and antioxidant enzyme activities of coriander (Coriandrum sativum L.) plants grown in lead (Pb)-spiked soil. Environ. Sci. Pollut. Res. 2021, 28, 1417–1425. [Google Scholar] [CrossRef]
- Hu, J.; Wu, X.; Wu, F.; Chen, W.; White, J.C.; Yang, Y.; Wang, B.; Xing, B.; Tao, S.; Wang, X. Potential application of titanium dioxide nanoparticles to improve the nutritional quality of coriander (Coriandrum sativum L.). J. Hazard. Mater. 2020, 389, 121837. [Google Scholar] [CrossRef]
- Sathishkumar, P.; Preethi, J.; Vijayan, R.; Mohd Yusoff, A.R.; Ameen, F.; Suresh, S.; Balagurunathan, R.; Palvannan, T. Anti-acne, anti-dandruff and anti-breast cancer efficacy of green synthesised silver nanoparticles using Coriandrum sativum leaf extract. J. Photochem. Photobiol. B Biol. 2016, 163, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.A.; Manaa, E.G.; Osama, N.; Aborehab, N.M.; Ragab, M.F.; Haggag, Y.A.; Ibrahim, M.T.; Hamdan, D.I. Coriander (Coriandrum sativum L.) essential oil and oil-loaded nano-formulations as an anti-aging potentiality via TGFβ/SMAD pathway. Sci. Rep. 2022, 12, 6578. [Google Scholar] [CrossRef] [PubMed]


| Pathology | Pharmacological Model | Biological Targets | C. sativum Compounds | Results | Reference |
|---|---|---|---|---|---|
| Leukemia | Chronic (K562) and acute (HL60) myeloid leukemia cell lines and on normal Vero cell line | ABL kinase ABL1 Bcl2 FLT3 | Epicatechin gallate, epicatechin, catechin rutin, vanillic acid | Activity on receptor: ABL kinase: similar effect for epicatechin and rutin ABL1: epicatechin gallate > rutin Bcl2: epicatechin gallate > vanillic acid FLT3: catechin > vanillic acid | [147] |
| Diabetic neuropathy | Diabetic neuropathy induced by streptozotocin | TNF-α | Quercetin, kaempferol, rutin | Quercetin> rutin> kaempferol | [148] |
| Diabetic nephropathy | Diabetic nephropathy in streptozotocin nicotinamide induced type 2 diabetes model | Receptors of AGEs | Linalool | Suitable binding interaction with amino acid residues of RAGEs (Dock score = −2.098 kcal/mol) | [149] |
| Diabetic nephropathy | Diabetic nephropathy induced by streptozotocin | KCNQ2/KCNQ3 | (E)-2-dodecenal | Binding in a juxtaposed zone between residues on the KCNQ S5 transmembrane segment and S4-5 linker | [89] |
| Hyperglycemia | In vitro methods for aldose reductase (ALR) enzyme inhibition, antiglycation activity, and sorbitol accumulation inhibition | ALR, receptors of AGEs, and sorbitol dehydrogenase | Flavonoid-rich seeds extract: Quercetin, rutin, kaempferol, (+)-catechin, luteolin, rhamnetin, and apigenin | Rutin, (+)-catechin, and rhamnetin exerted a suitable binding with all receptors with a docking score up to −4.94 for sorbitol dehydrogenase, and mostly hydrogen bonding and hydrophobic interactions were found with ALR enzyme receptors | [150] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santibáñez, A.; Jiménez-Ferrer, E.; Angulo-Bejarano, P.I.; Sharma, A.; Herrera-Ruiz, M. Coriandrum sativum and Its Utility in Psychiatric Disorders. Molecules 2023, 28, 5314. https://doi.org/10.3390/molecules28145314
Santibáñez A, Jiménez-Ferrer E, Angulo-Bejarano PI, Sharma A, Herrera-Ruiz M. Coriandrum sativum and Its Utility in Psychiatric Disorders. Molecules. 2023; 28(14):5314. https://doi.org/10.3390/molecules28145314
Chicago/Turabian StyleSantibáñez, Anislada, Enrique Jiménez-Ferrer, Paola Isabel Angulo-Bejarano, Ashutosh Sharma, and Maribel Herrera-Ruiz. 2023. "Coriandrum sativum and Its Utility in Psychiatric Disorders" Molecules 28, no. 14: 5314. https://doi.org/10.3390/molecules28145314
APA StyleSantibáñez, A., Jiménez-Ferrer, E., Angulo-Bejarano, P. I., Sharma, A., & Herrera-Ruiz, M. (2023). Coriandrum sativum and Its Utility in Psychiatric Disorders. Molecules, 28(14), 5314. https://doi.org/10.3390/molecules28145314