
Integrated Transcriptomic and Metabolomic Analyses Reveal the Effects of Grafting on Special Metabolites of Acanthopanax senticosus Leaves

Abstract
1. Introduction
2. Results
2.1. LC-MS/MS-Based Metabolomics Reveals Metabolite Changes in GSCL vs. SCL
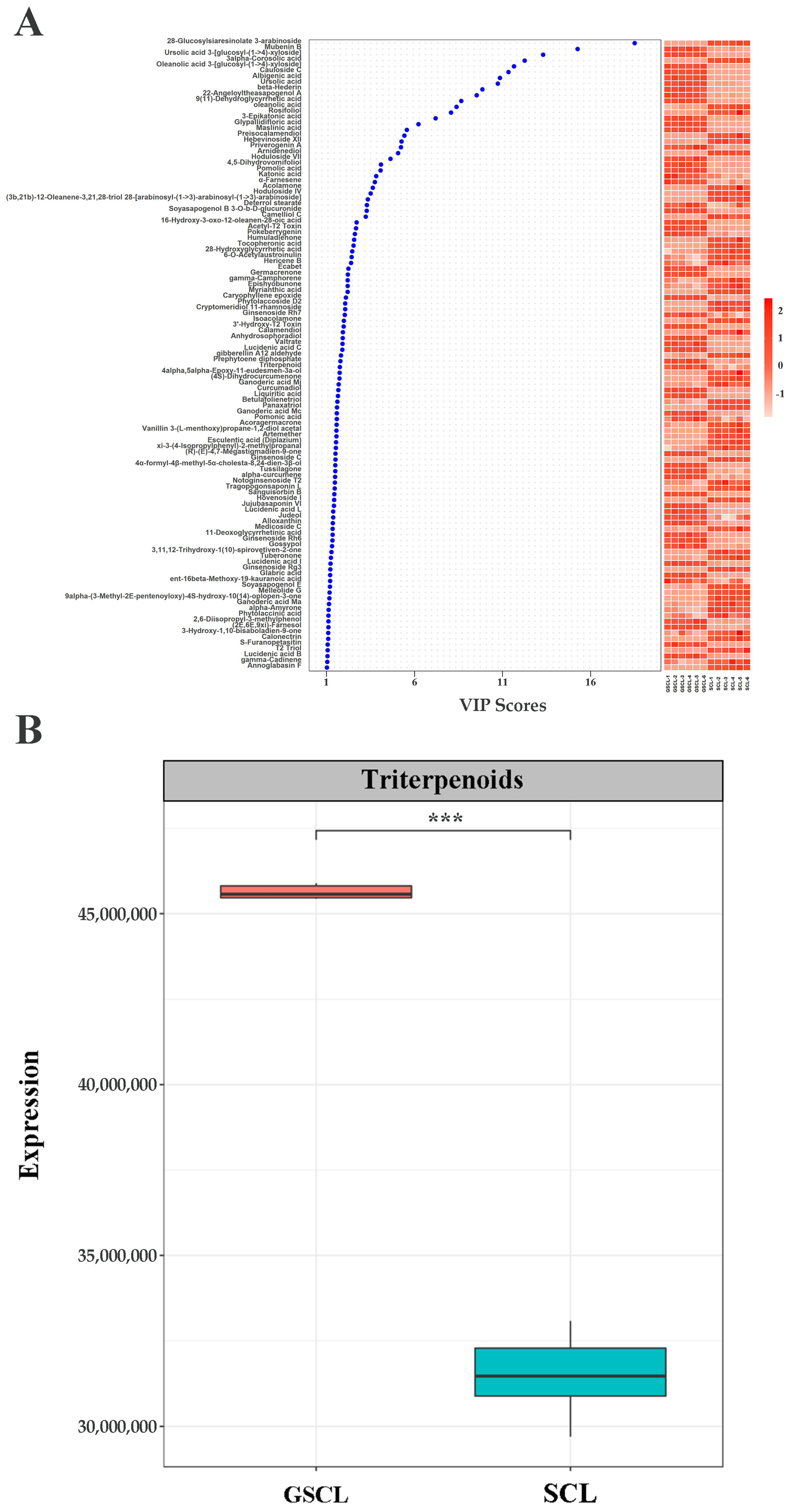
2.2. Special Differential Metabolite Analysis in GSCL vs. SCL
2.3. De Novo Assembly, Analysis, and Functional Annotation of RNA-seq Data
2.4. GO and KEGG Enrichment Analysis of Differential Expression Genes in GSCL vs. SCL
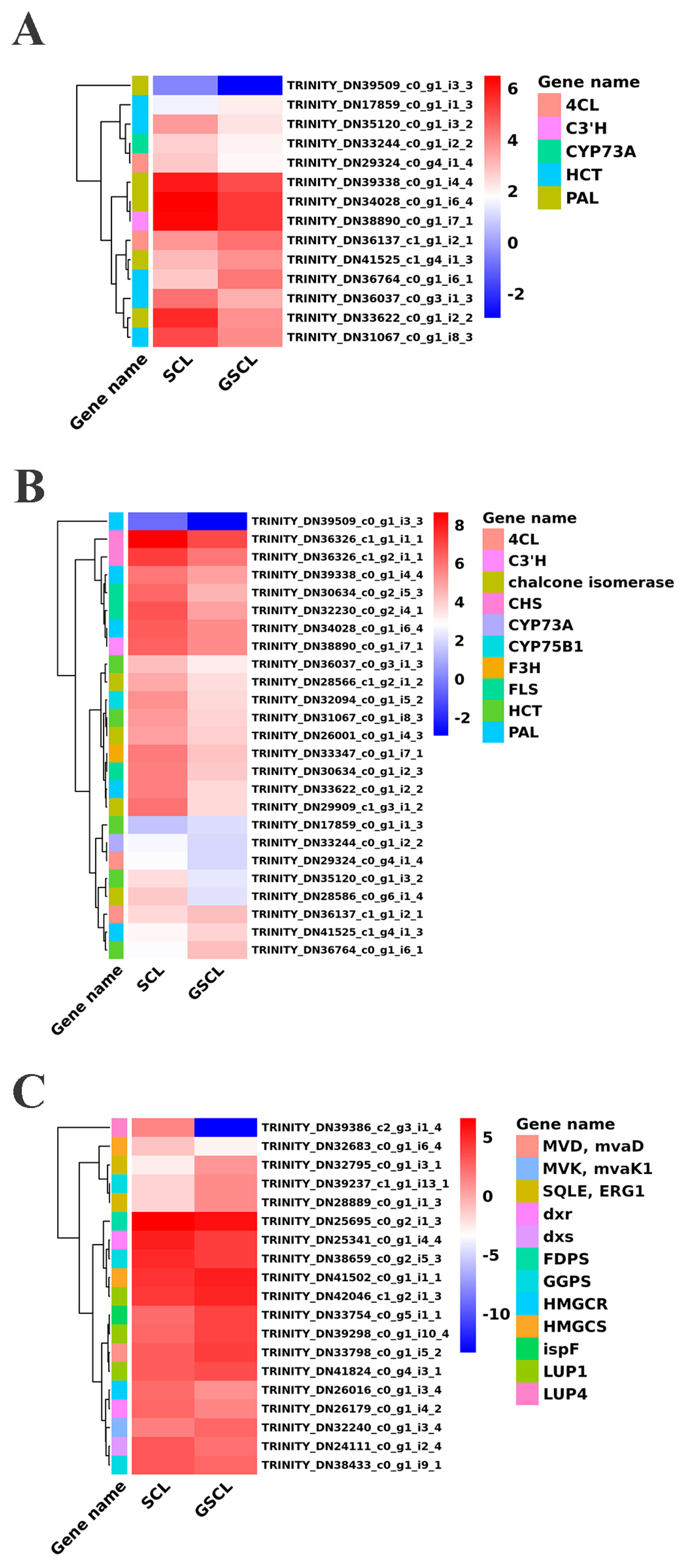
2.5. Differential Expression Genes in Special Metabolic Pathways in GSCL vs. SCL
2.6. Association Analysis Target to Special Metabolites
2.7. qRT-PCR Validation
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Sample Extraction and Metabolite Profiling
4.3. Multivariate Statistical Analysis and Metabolite Identification
4.4. RNA Extraction, cDNA Library Construction and IIIumina Sequencing
4.5. De Novo Transcriptome Assembly and Functional Annotation
4.6. Screening and Enrichment of Differentially Expressed Genes
4.7. Quantitative Real-Time PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Wang, K.W.; Yan, A. Protective effects of total flavonoids of Acanthopanax senticosus on brain tissue of neonatal rats with hypoxic-ischemic brain damage. Chin. J. Clin. Pharmacol. 2020, 36, 624–627. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Wang, R.J.; Shi, L.Q.; Liu, Z.Q.; Liu, Z.Y. Study on the therapeutic material basis and effect of Acanthopanax senticosus (Rupr. et Maxim.) Harms leaves in the treatment of ischemic stroke by PK-PD analysis based on online microdialysis-LC-MS/MS method. Food Funct. 2020, 11, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Wang, Q.; Li, Z.F.; Feng, Y.; Li, Y.; Yang, S.L.; Feng, Y.L. Different Metabolites in the Roots, Seeds, and Leaves of Acanthopanax senticosus and Their Role in Alleviating Oxidative Stress. J. Anal. Methods Chem. 2021, 26, 662–669. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.P.; Wu, D.; Sun, Y.P.; Zhao, H.Q.; Wang, Y.Y.; Zhang, W.S.; Su, F.Z.; Yang, B.Y.; Wang, Q.H.; Kuang, H.X. Comprehensive Analysis of Eleutherococcus senticosus (Rupr. & Maxim.) Maxim. Leaves Based on UPLC-MS/MS: Separation and Rapid Qualitative and Quantitative Analysis. Front. Pharmacol. 2022, 13, 865586. [Google Scholar] [CrossRef]
- Guo, M.Q.; Song, F.R.; Liu, Z.Q.; Liu, S.Y. Characterization of triterpenoidic saponin mixture in crude extracts from leaves of Acanthopanax senticosus Harms by saponin structural correlation and mass spectrometry. Anal. Chim. Acta 2006, 557, 198–203. [Google Scholar] [CrossRef]
- Jin, Z.X.; Xu, S.Y.; Sun, S.Q.; Zhou, Q. Analysis of Acanthopanax senticosus harms for different parts using fourier transform infrared spectroscopy. Spectrosc. Spect. Anal. 2008, 28, 2859–2863. [Google Scholar] [CrossRef]
- Liu, M.Y.; Li, N.N.; Shan, S.; Shi, Y.D.; Zhu, Y.B.; Lu, W. Acanthopanax senticosus Polysaccharide Enhances the Pathogen Resistance of Radiation-Damaged Caenorhabditis elegans through Intestinal p38 MAPK-SKN-1/ATF-7 Pathway and Stress Response. Int. J. Mol. Sci. 2022, 23, 5034. [Google Scholar] [CrossRef]
- Liu, H.D.; Yang, L.; Wan, C.L.; Li, Z.N.; Yan, G.L.; Han, Y.; Sun, H.; Wang, X.J. Exploring potential mechanism of ciwujia tablets for insomnia by UPLC-Q-TOF-MS/MS, network pharmacology, and experimental validation. Front. Pharmacol. 2022, 13, 990996. [Google Scholar] [CrossRef]
- Shang, H.H.; Wang, M.; Liu, Y.; Zheng, Y.N.; Bai, R.; Liao, M.L. Determination of protocatechuic acid, syringing, chlorogenic acid, eleutheroside E, and isofraxidin in Acanthopanax senticosus from various of habitats by RP-HPLC. Drugs Clin. 2018, 33, 1324–1328. [Google Scholar]
- Fu, X.; Liu, D.J.; Guo, L.L.; Shen, J.; Zhang, H.M.; Li, L.L. Ultrasonic-Assisted Enzyme Extraction and Antioxidant Activity of Chlorogenic Acid from Trollius chinensis. Mol. Plant Breed. 2023, 2, 1–12. [Google Scholar]
- Liu, G.L.; Wei, X.E.; Wu, Y.; Chen, C.Y.; Cai, W.J. Simultaneous determination of syringin, chlorogenic acid, gardenia glycosides and salidroside content by high performance liquid chromatography. J. Food Saf. Qual. 2017, 8, 4847–4853. [Google Scholar]
- Wang, R.J. Metabolomics-Based Study on the Mechanism of Eleutherococcus senticosus (Rupr. & Maxim.) Maxim. Leaves in the Treatment of Ischemic Stroke. Ph.D. Thesis, Jilin University, Changchun, China, 2021. [Google Scholar] [CrossRef]
- Gao, S.H.; Shan, B.Y.; Xu, Y.; Bao, H.W. Simultaneous Determination of Ferulic Acid and Quercetin in Wujia Shenghua Capsules by HPLC. J. Chang. Norm. Univ. 2018, 37, 112–116. [Google Scholar]
- Li, Q.L.; Danzeng, L.Z.; Liang, L.L.; Chen, Z.L.; Wang, X.X.; Liu, H. Effect of quercetin on cerebral ischemia-reperfusion injury involved and its mechanism. Acta Nutr. Sin. 2023, 45, 181–187. [Google Scholar] [CrossRef]
- Liu, Y. Adaptive Evolution of Key Enzyme Genes in Plant Triterpene Synthesis Pathway. Master’s Thesis, Guang Xi Medical University, Nanning, China, 2013. [Google Scholar]
- Jin, J.L.; Lee, S.; Lee, Y.Y.; Kim, J.M.; Heo, J.E.; Yun-Choi, H.S. Platelet anti-aggregating triterpenoids from the leaves of Acanthopanax senticosus and the fruits of A. sessiliflorus. Planta Med. 2004, 70, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.W.; Han, J.Y.; Fan, Y.B.; Tai, Y.L.; Zhu, B.Y.; Lu, M.Q.; Wang, R.J.; Wan, X.C.; Zhang, Z.Z. Uncovering tea-specific secondary metabolism using transcriptomic and metabolomic analyses in grafts of Camellia sinensis and C. oleifera. Tree Genet. Genomes 2018, 14, 23. [Google Scholar] [CrossRef]
- Xu, D.B.; Yuan, H.W.; Tong, Y.F.; Zhao, L.; Qiu, L.L.; Guo, W.B.; Shen, C.J.; Liu, H.J.; Yan, D.L.; Zheng, B.S. Comparative proteomic analysis of the graft unions in hickory (Carya cathayensis) provides insights into response mechanisms to grafting process. Front. Plant Sci. 2017, 8, 676. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable grafting: The implications of a growing agronomic imperative for vegetable fruit quality and nutritive value. Front. Plant Sci. 2017, 8, 741. [Google Scholar] [CrossRef]
- Habran, A.; Commisso, M.; Helwi, P.; Hilbert, G.; Negri, S.; Ollat, N.; Gomes, E.; Leeuwen, C.V.; Guzzo, F.; Delrot, S. Rootstocks/scion/nitrogen interactions affect secondary metabolism in the grape berry. Front. Plant Sci. 2016, 7, 1134. [Google Scholar] [CrossRef]
- Duan, X.; Bi, H.G.; Li, T.; Wu, G.X.; Li, Q.; Ai, X.Z. Root characteristics of grafted peppers and their resistance to Fusarium solani. Biol. Plant. 2017, 61, 579–586. [Google Scholar] [CrossRef]
- Proietti, S.; Rouphael, Y.; Colla, G.; Cardarelli, M.; Agazio, M.D.; Zacchini, M.; Rea, E.; Moscatello, S.; Battistelli, A. Fruit quality of mini-watermelon as affected by grafting and irrigation regimes. J. Sci. Food Agric. 2008, 88, 1107–1114. [Google Scholar] [CrossRef]
- Zhou, Y.C.; Liu, Y.; Zheng, W.Z.; Gao, J.; Kong, S.; Lv, Y.C. Analysis of Metabolite Difference between Scion and Scion Female Parent of Compound Tea Plant. Acta Bot. Boreal.-Occident. Sin. 2022, 42, 1654–1662. [Google Scholar]
- Cookson, S.J.; Clemente Moreno, M.J.; Hevin, C.; Nyamba Mendome, L.Z.; Delrot, S.; Trossat-Magnin, C.; Ollat, N. Graft union formation in grapevine induces transcriptional changes related to cell wall modification, wounding, hormonesignalling, and secondary metabolism. J. Exp. Bot. 2013, 64, 2997–3008. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Lewinsohn, E. Convergent evolution in plant specialized metabolism. Annu. Rev. Plant Biol. 2011, 62, 549–566. [Google Scholar] [CrossRef]
- De Luca, V.; Satim, V.; Atsumi, S.M.; Yu, F. Mining the biodiversity of plants: A revolution in the making. Science 2012, 336, 1658–1661. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y.; Lu, M.Q.; Zhu, B.Y.; Wang, R.J.; Wan, X.C.; Deng, W.W.; Zhang, Z.Z. Integrated transcriptomic and phytochemical analyses provide insights into characteristic metabolites variation in leaves of 1-year-old grafted tea (Camellia sinensis). Tree Genet. Genomes 2019, 15, 58. [Google Scholar] [CrossRef]
- Hoffffmann, L.; Maury, S.; Martz, F.; Geoffroy, P.; Legrand, M. Purifification, cloning, and properties of an acyltransferase controlling shikimate and quinate ester intermediates in phenylpropanoid metabolism. J. Biol. Chem. 2003, 278, 95–103. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Shakya, R.; Sengoda, V.G.; Munyaneza, J.E.; Swamy, P.; Navarre, D.A. Synthesis and regulation of chlorogenic acid in potato: Rerouting phenylpropanoid flflux in HQT-silenced lines. Plant Biotechnol. J. 2015, 13, 551–564. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Zheng, K.; Xie, M.; Feng, K.; Jawdy, S.S.; Gunter, L.E.; Ranjan, P.; Singan, V.R.; Engle, N.; et al. Genomewide association studies and expression-based quantitative trait loci analyses reveal roles of HCT2 in caffffeoylquinic acid biosynthesis and its regulation by defense-responsive transcription factors in Populus. N. Phytol. 2018, 220, 502–516. [Google Scholar] [CrossRef]
- Chen, X.D.; Cai, W.J.; Xia, J.; Yu, H.M.; Wang, Q.L.; Pang, F.H.; Zhao, M.Z. Metabolomic and transcriptomic analyses reveal that blue light promotes chlorogenic acid synthesis in strawberry. J. Agric. Food Chem. 2020, 68, 12485–12492. [Google Scholar] [CrossRef]
- Wen, H.; Jiang, X.; Wang, W.; Wu, M.; Bai, H.; Wu, C.; Shen, L. Comparative transcriptome analysis of candidate genes involved in chlorogenic acidbiosynthesis during fruit development in three pear varieties of Xinjiang Uygur Autonomous Region. J. Zhejiang Univ.-Sci. B (Biomed. Biotechnol.) 2022, 23, 345–356. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Loke, K.K.; Rahnamaie-Tajadod, R.; Yeoh, C.C.; Goh, H.H.; Mohamed-Hussein, Z.A.; Zainal, Z.; Ismail, I.; Noor, N.M. Transcriptome analysis of Polygonum minus reveals candidate genes involved in important secondary metabolic pathways of phenylpropanoids and flflavonoids. PeerJ 2017, 5, e2938. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.B.; Noel, J.P. The chalcone synthase superfamily of type III polyketide synthases. Nat. Prod. Rep. 2003, 20, 79–110. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Grotewold, E. The genetics and biochemistry of flfloral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef]
- Mahajan, M.; Joshi, R.; Gulati, A.; Yadav, S.K. Increase in flavan-3-ols by silencing flavonol synthase mRNA affffects the transcript expression and activity levels of antioxidant enzymes in tobacco. Plant Biol. 2012, 14, 725–733. [Google Scholar] [CrossRef]
- Xia, H.; Zhang, X.F.; Shen, Y.Q.; Guo, Y.Q.; Wang, T.; Wang, J.; Lin, L.J.; Deng, H.H.; Deng, Q.X.; Xu, K.F.; et al. Comparative analysis of flavonoids in white and red table grape cultivars during ripening by widely targeted metabolome and transcript levels. J. Food Sci. 2022, 87, 1650–1661. [Google Scholar] [CrossRef]
- Yuan, F.; Yin, X.; Zhao, K.H.; Lan, X.Z. Transcriptome and Metabolome Analyses of Codonopsis convolvulacea Kurz Tuber, Stem, and Leaf Reveal the Presence of Important Metabolites and Key Pathways Controlling Their Biosynthesis. Front Genet. 2022, 13, 884224. [Google Scholar] [CrossRef]
- Wang, Y.S.; Xu, Y.J.; Gao, L.P.; Yu, O.; Wang, X.Z.; He, X.J.; Jiang, X.L.; Liu, Y.Y.; Xia, T. Functional analysis of flavonoid 3′, 5′-hydroxylase from tea plant (Camellia sinensis): Critical role in the accumulation of catechins. BMC Plant Biol. 2014, 14, 347. [Google Scholar] [CrossRef]
- Xu, X.S.; Zhang, F.S.; Qin, X.M. Progress of triterpene saponin biosynthesis pathway and key enzymes. Mod. Tradit. Chin. Med. Mater. Med.-World Sci. Technol. 2014, 16, 2440–2448. [Google Scholar] [CrossRef]
- Xia, Q.F. Key Genes Cloning and Expression Analysis in Pathway of Terpenoids Metabolism of Blumea balsamifera L.DC. Master’s Thesis, Guizhou University, Guiyang, China, 2017. [Google Scholar]
- Zhang, G.X.; Wu, Y.G.; Muhammad, Z.U.; Yang, Y.Z.; Yu, J.; Zhang, J.F.; Yang, D.M. cDNA cloning, prokaryotic expression and functional analysis of 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGCR) in Pogostemon cablin. Protein Expr. Purif. 2019, 163, 105454. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zeng, Q.P. Genetic manipulation of terpenoid biosynthesis. China Biotechnol. 2006, 60–64. [Google Scholar] [CrossRef]
- Abe, I.; Rohmer, M.; Prestwich, G.D. Enzymatic cyclization of squalene and oxidosqualene to sterols and triterpenes. Chem. Rev. 1993, 93, 2189. [Google Scholar] [CrossRef]
- Robert, A.H.; Joseph, D.C. Triterpenoids. Nat. Prod. Rep. 2008, 35, 1294–1329. [Google Scholar] [CrossRef]
- Qi, X.Q.; Bakht, S.; Qin, B.; Leggett, M.; Hemmings, A.; Mellon, F.; Eagles, J.; Werck-Reichhart, D.; Schaller, H.; Lesot, A.; et al. A different function for a member of an ancient and highly conserved cytochrome P450 family: From essential sterols to plant defense. Proc. Natl. Acad. Sci. USA 2007, 103, 18848–18853. [Google Scholar] [CrossRef]
- Ranjith, K.; Ilango, R.V.J. Impact of grafting methods, scion materials and number of scions on graft success, vigour and flowering of top worked plants in tea (Camellia spp.). Sci. Hortic. 2017, 220, 139–146. [Google Scholar] [CrossRef]
- Ruan, C.J.; Mopper, S. High-crown grafting to increase low yields in Camellia oleifera. J. Hortic. Sci. Biotechnol. 2017, 92, 439–444. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Pachter, L. Streaming fragment assignment for real-time analysis of sequencing experiments. Nat. Methods 2012, 10, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level-The DESeq Package; European Molecular Biology Laboratory (EMBL): Heidelberg, Germany, 2013. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Special Differential Metabolite | GSCL | SCL | VIP | p-Values |
|---|---|---|---|---|
| syringin | 501,175.96 ± 87,306 | 546,293.52 ± 224,004 | 0.6022 | 0.6556 |
| isofraxidin | 11,210.14 ± 2332 | 17,301.89 ± 1466 | 0.5450 | 0.0003 |
| chlorogenic | 1,856,490.76 ± 385,710 | 507,624.08 ± 40,181 | 8.4377 | <0.0001 |
| quercetin | 675,766.18 ± 74,694 | 914,886.84 ± 68,981 | 3.4413 | 0.0002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Deng, K.; Ai, J.; Wang, Y.; Wang, Y.; Ren, Y.; Zhang, N. Integrated Transcriptomic and Metabolomic Analyses Reveal the Effects of Grafting on Special Metabolites of Acanthopanax senticosus Leaves. Molecules 2023, 28, 4877. https://doi.org/10.3390/molecules28124877
Wang Q, Deng K, Ai J, Wang Y, Wang Y, Ren Y, Zhang N. Integrated Transcriptomic and Metabolomic Analyses Reveal the Effects of Grafting on Special Metabolites of Acanthopanax senticosus Leaves. Molecules. 2023; 28(12):4877. https://doi.org/10.3390/molecules28124877
Chicago/Turabian StyleWang, Qi, Kedan Deng, Jun Ai, Yingping Wang, Yougui Wang, Yueying Ren, and Nanqi Zhang. 2023. "Integrated Transcriptomic and Metabolomic Analyses Reveal the Effects of Grafting on Special Metabolites of Acanthopanax senticosus Leaves" Molecules 28, no. 12: 4877. https://doi.org/10.3390/molecules28124877
APA StyleWang, Q., Deng, K., Ai, J., Wang, Y., Wang, Y., Ren, Y., & Zhang, N. (2023). Integrated Transcriptomic and Metabolomic Analyses Reveal the Effects of Grafting on Special Metabolites of Acanthopanax senticosus Leaves. Molecules, 28(12), 4877. https://doi.org/10.3390/molecules28124877