
Analysis of Floral Color Differences between Different Ecological Conditions of Clematis tangutica (Maxim.) Korsh


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
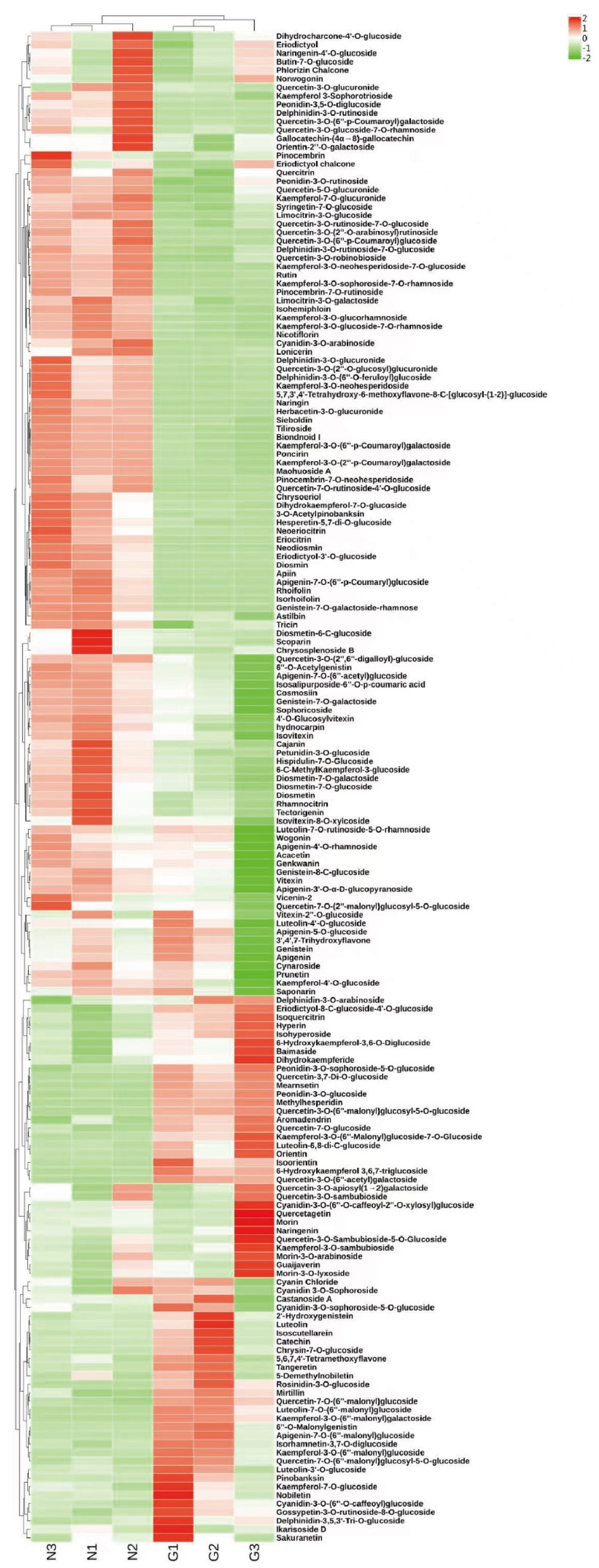
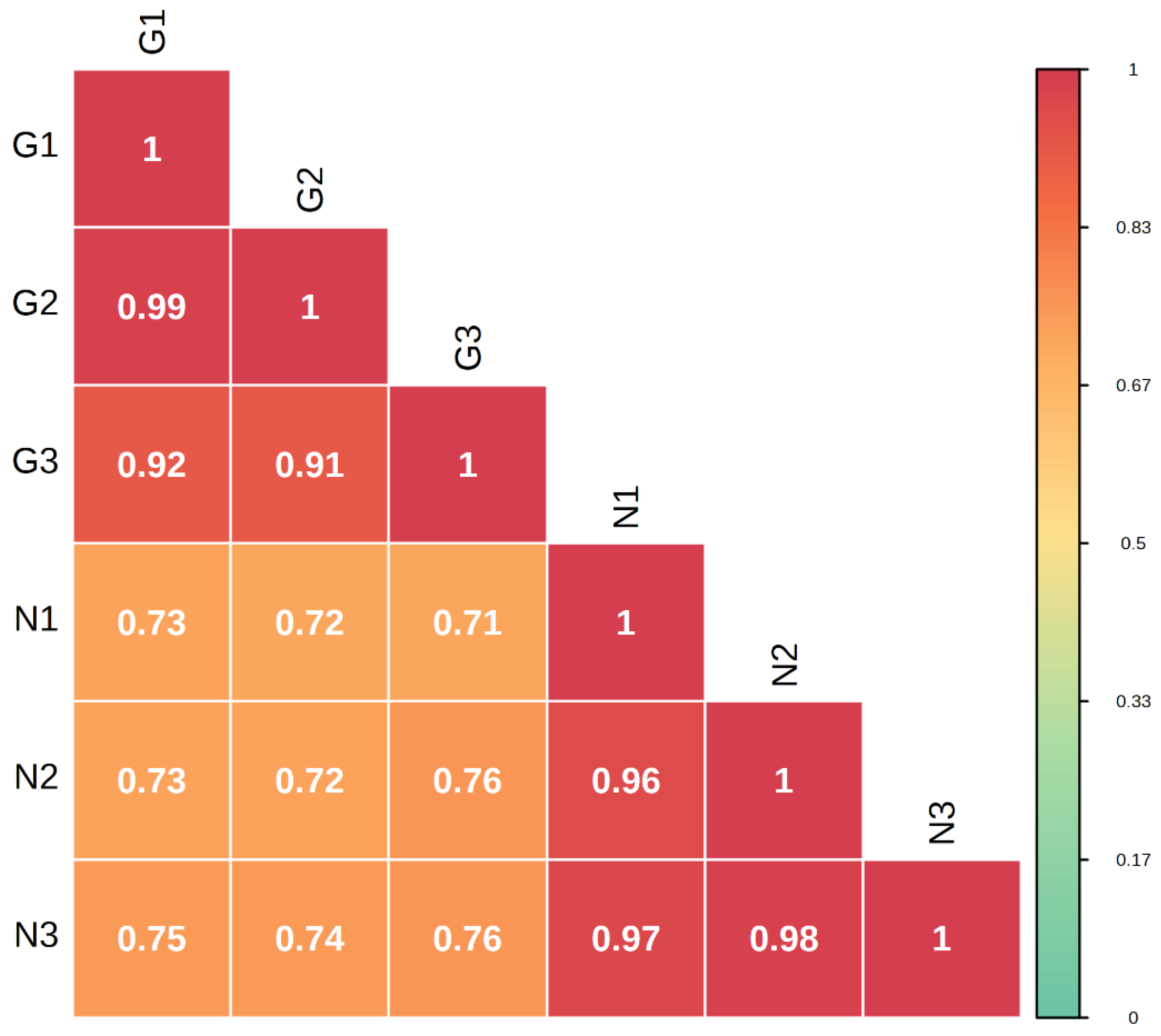
2.1. Analysis of Metabolites in the Flowers of C. tangutica (Maxim.) Korsh
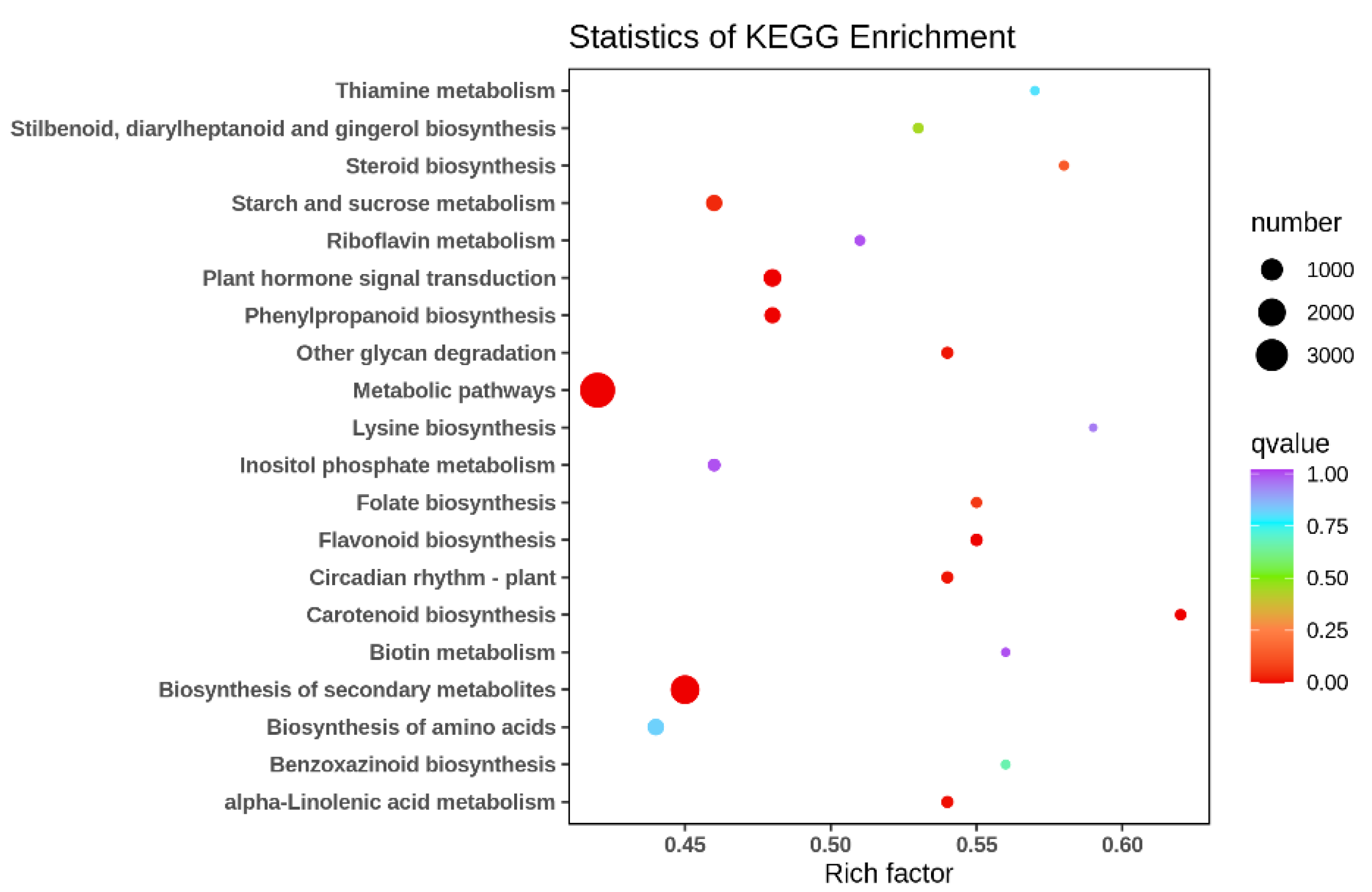
2.2. Differential Genes in Clematis tangutica (Maxim.) Korsh. Flowers in Different Ecological Environments
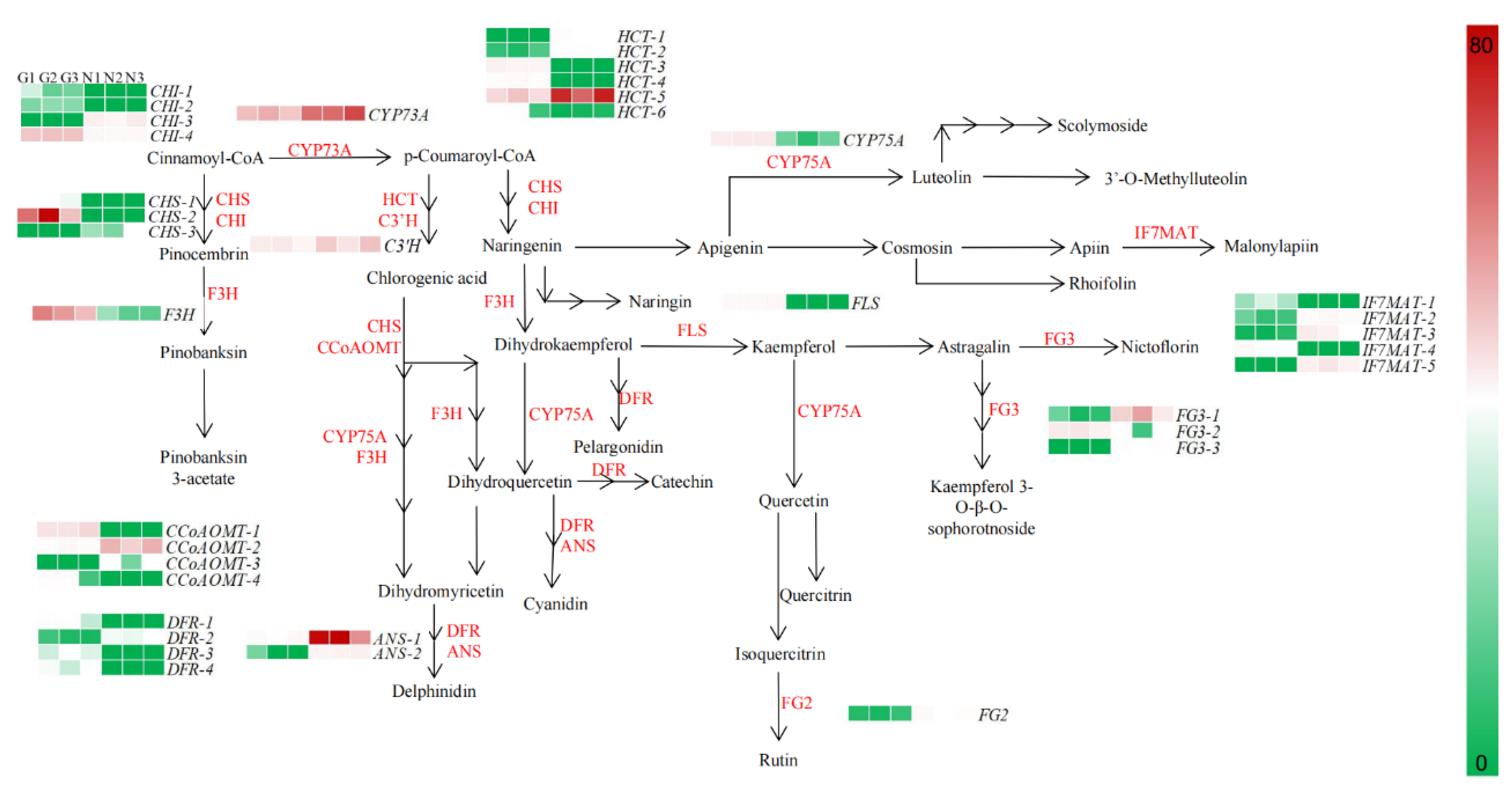
2.3. Expression Patterns of Genes Expressed in Relation to the Flavonoid Synthesis Pathway
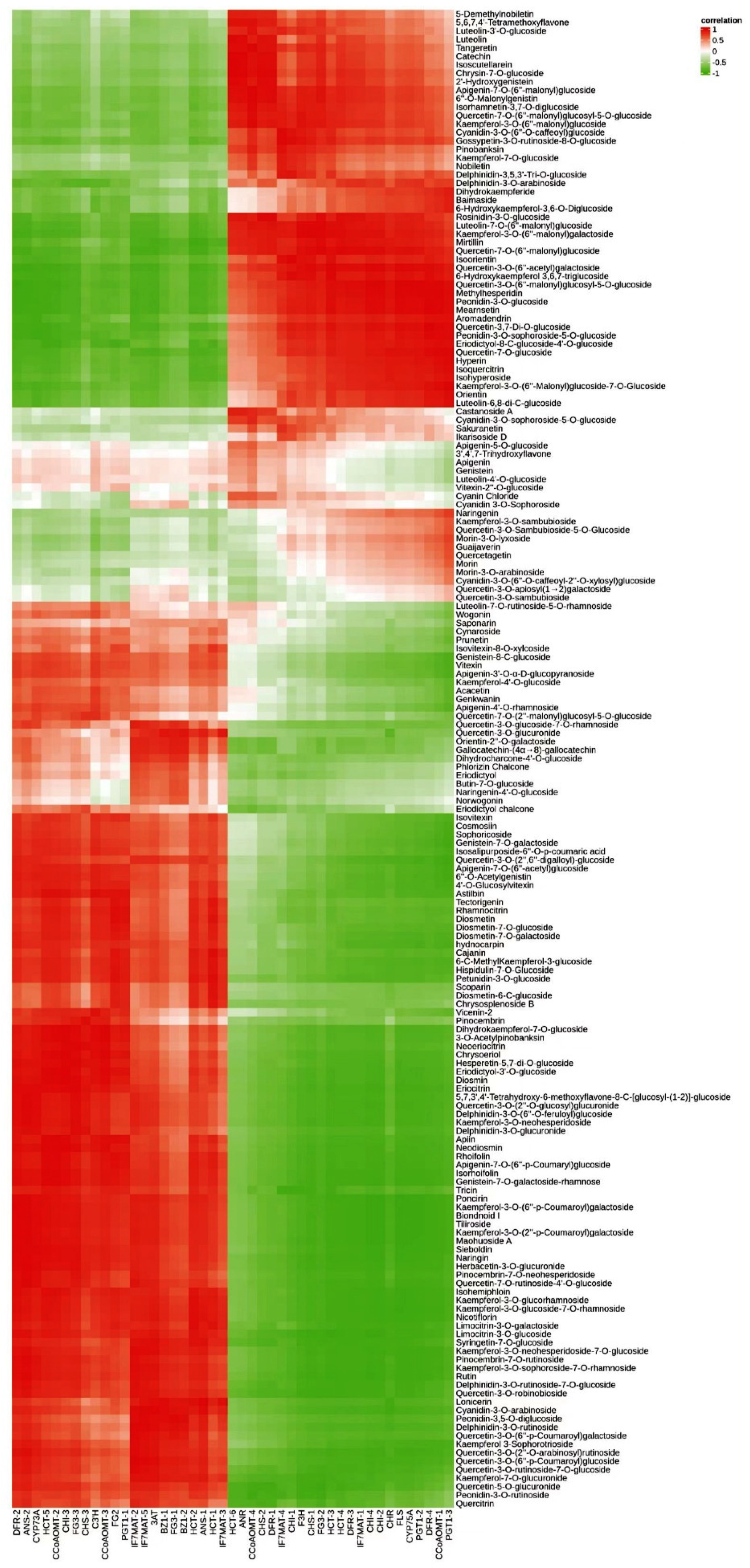
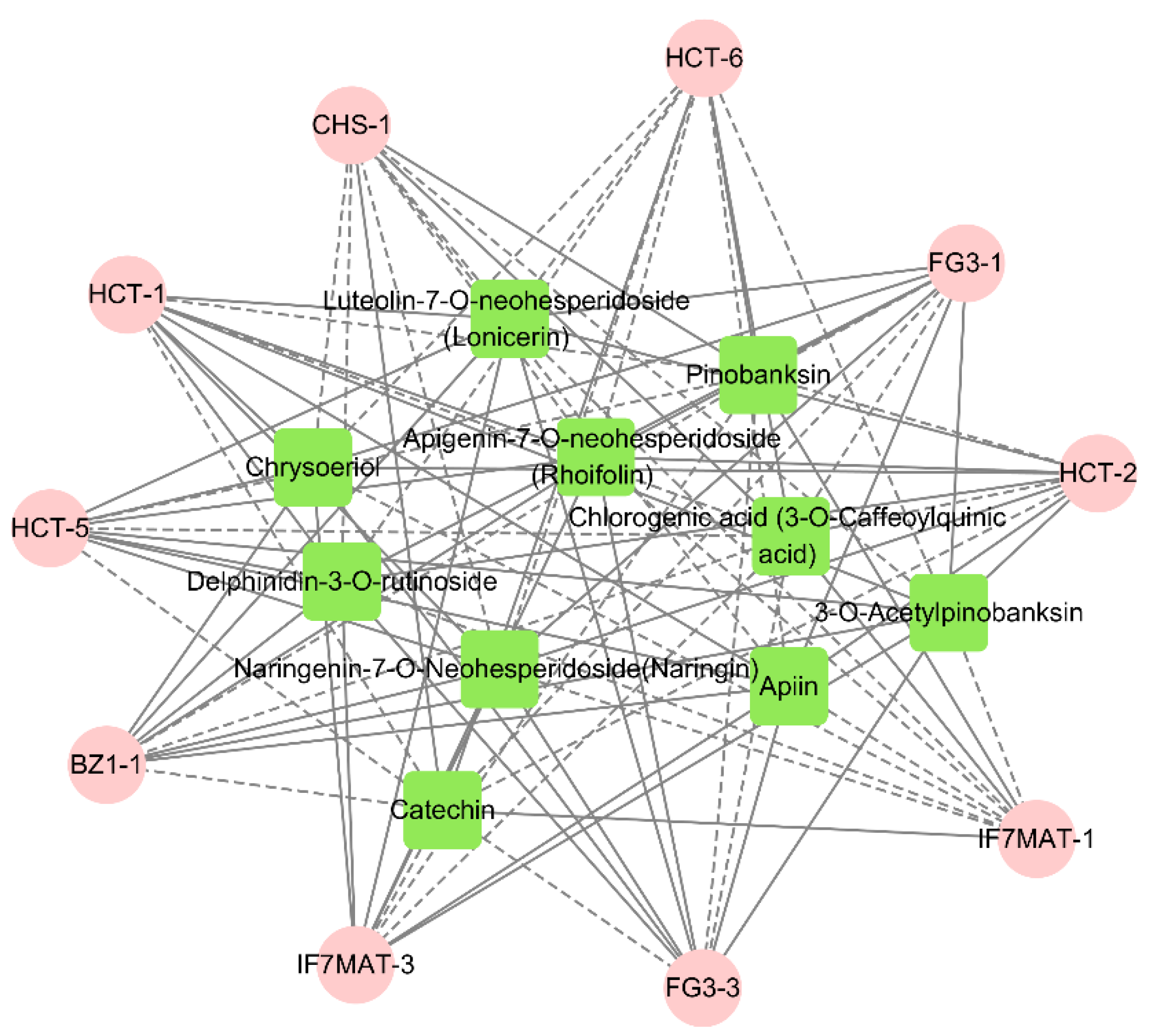
2.4. Correlation between Flavonoid Content and Flavonoid Synthesis Gene Expression
3. Discussion
4. Materials and Methods
4.1. Plant Material
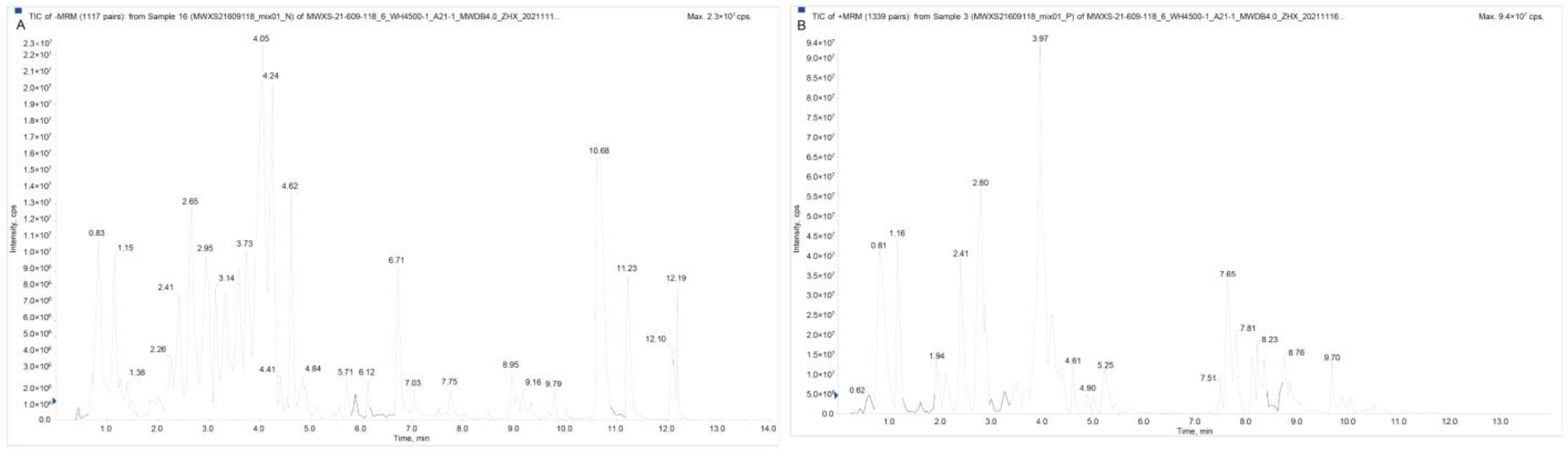
4.2. Extraction and Analysis of Flavonoid Metabolites
4.3. RNA Extraction and Sequencing
4.4. Transcriptomic Data Analysis
4.5. Correlation Analysis and Correlation Network Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, W.; Zhou, N.; Wang, Y.; Sun, S.; Zhang, Y.; Ju, Z.; Yi, Y. Characterization and functional analysis of RdDFR1 regulation on flower color formation in Rhododendron delavayi. Plant Physiol. Biochem. 2021, 169, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Halevas, E.G.; Avgoulas, D.I.; Katsipis, G.; Pantazaki, A.A. Flavonoid-liposomes formulations: Physico-chemical characteristics, biological activities and therapeutic applications. Eur. J. Med. Chem. Rep. 2022, 5, 100059. [Google Scholar] [CrossRef]
- Chen, S.; Liu, J.; Dong, G.; Zhang, X.; Liu, Y.; Sun, W.; Liu, A. Flavonoids and caffeoylquinic acids in Chrysanthemum morifolium Ramat flowers: A potentially rich source of bioactive compounds. Food Chem. 2021, 344, 128733. [Google Scholar] [CrossRef] [PubMed]
- Schlangen, K.; Miosic, S.; Halbwirth, H. Allelic variants from Dahlia variabilis encode flavonoid 3′-hydroxylases with functional differences in chalcone 3-hydroxylase activity. Arch. Biochem. Biophys. 2010, 494, 40–45. [Google Scholar] [CrossRef]
- Schmitzer, V.; Veberic, R.; Stampar, F. Prohexadione-Ca application modifies flavonoid composition and color characteristics of rose (Rosa hybrida L.) flowers. Sci. Hortic. 2012, 146, 14–20. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, C.; Huang, Z.; Lyu, L.; Li, J.; Li, W.; Wu, W. The color difference of rubus fruits is closely related to the composition of flavonoids including anthocyanins. LWT 2021, 149, 111825. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhou, W.; Chen, Y.; Li, Z.; Song, X.; Wang, J.; Tian, D.; Niu, J. Metabolite analysis in Nymphaea ‘Blue Bird’ petals reveal the roles of flavonoids in color formation, stress amelioration, and bee orientation. Plant Sci. 2021, 312, 111025. [Google Scholar] [CrossRef]
- Chen, S.-M.; Li, C.-H.; Zhu, X.-R.; Deng, Y.-M.; Sun, W.; Wang, L.-S.; Chen, F.-D.; Zhang, Z. The identification of flavonoids and the expression of genes of anthocyanin biosynthesis in the chrysanthemum flowers. Biol. Plant. 2012, 56, 458–464. [Google Scholar] [CrossRef]
- He, Q.; Shen, Y.; Wang, M.; Huang, M.; Yang, R.; Zhu, S.; Wang, L.; Xu, Y.; Wu, R. Natural variation in petal color in Lycoris longituba revealed by anthocyanin components. PLoS ONE 2011, 6, e22098. [Google Scholar] [CrossRef]
- Zhao, D.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant Sci. 2015, 261. [Google Scholar] [CrossRef]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef]
- Wallace, T.C.; Giusti, M.M. Anthocyanins. Adv. Nutr. 2015, 6, 620–622. [Google Scholar] [CrossRef]
- Verweij, W.; Spelt, C.; Di Sansebastiano, G.-P.; Vermeer, J.; Reale, L.; Ferranti, F.; Koes, R.; Quattrocchio, F. An H+ P-ATPase on the tonoplast determines vacuolar pH and flower colour. Nat. Cell Biol. 2008, 10, 1456–1462. [Google Scholar] [CrossRef]
- Yari, A.; Rashnoo, S. Optimization of a new method for extraction of cyanidin chloride and pelargonidin chloride anthocyanins with magnetic solid phase extraction and determination in fruit samples by HPLC with central composite design. J. Chromatogr. B 2017, 1067, 38–44. [Google Scholar] [CrossRef]
- Yang, L.; Rong-Rong, C.; Ji-Li, F.; Ke, Y. Total anthocyanins and cyanidin-3-O-glucoside contents and antioxidant activities of purified extracts from eight different pigmented plants. Pharmacogn. Mag. 2019, 15, 124. [Google Scholar]
- Kato, M.; Tani, T.; Terahara, N.; Tsuda, T. PLoS ONE 2015, 10, e0126157. [CrossRef]
- Toki, K.; Kawanishi, H. Survey of peonidin glycosides in garden snapdragons and changes in anthocyanin contents during floral development. J. Jpn. Soc. Hortic. Sci. 1992, 60, 989–995. [Google Scholar] [CrossRef]
- Catalano, G.; Fossen, T.; Andersen, Ø.M. Petunidin 3-O-α-rhamnopyranoside-5-O-β-glucopyranoside and other anthocyanins from flowers of Vicia villosa. J. Agric. Food Chem. 1998, 46, 4568–4570. [Google Scholar] [CrossRef]
- Peixoto, F.; Senna Gouvêa, A.; de Araújo Santiago, M.; de Sá Velosos Martins, Z.; Galhardo Borguini, R.; de Oliveira Godoy, R. Characterization and bioaccessibility of anthocyanins from blueberry (Vaccinium corymbosum L.) after simulated gastro-intestinal digestion: A positive effect on malvidin derivatives. Fruits 2018, 73, 101–109. [Google Scholar] [CrossRef]
- Tanaka, Y.; Brugliera, F.; Chandler, S. Recent progress of flower colour modification by biotechnology. Int. J. Mol. Sci. 2009, 10, 5350–5369. [Google Scholar] [CrossRef] [PubMed]
- Koes, R.E.; Quattrocchio, F.; Mol, J.N. The flavonoid biosynthetic pathway in plants: Function and evolution. BioEssays 1994, 16, 123–132. [Google Scholar] [CrossRef]
- Quattrocchio, F.; Baudry, A.; Lepiniec, L.; Grotewold, E. The regulation of flavonoid biosynthesis. In The Science of Flavonoids; Springer: Berlin/Heidelberg, Germany, 2006; pp. 97–122. [Google Scholar]
- Wan, L.; Lei, Y.; Yan, L.; Liu, Y.; Pandey, M.K.; Wan, X.; Varshney, R.K.; Fang, J.; Liao, B. Transcriptome and metabolome reveal redirection of flavonoids in a white testa peanut mutant. BMC Plant Biol. 2020, 20, 161. [Google Scholar] [CrossRef] [PubMed]
- Forkmann, G. Flavonoids as flower pigments: The formation of the natural spectrum and its extension by genetic engineering. Plant Breed. 1991, 106, 1–26. [Google Scholar] [CrossRef]
- Isaak, C.K.; Petkau, J.C.; Blewett, H.; Karmin, O.; Siow, Y.L. Lingonberry anthocyanins protect cardiac cells from oxidative-stress-induced apoptosis. Can. J. Physiol. Pharmacol. 2017, 95, 904–910. [Google Scholar] [CrossRef]
- Jung, J.; Shin, M.; Jeong, N.; Hwang, D. Antioxidant and anti-inflammatory activities of ethanol extract of Clematis trichotoma Nakai. Korean J. Clin. Lab. Sci. 2021, 53, 165–173. [Google Scholar] [CrossRef]
- Wei, Y.; Chen, T.; Wang, S.; Shen, C.; Yan, S.; Song, Z.; Liu, Y.; Yang, F.; Hai, P.; Li, Y. Separation of a new triterpenoid saponin together with six known ones from Clematis tangutica (Maxim.) Korsh and evaluation of their cytotoxic activities. Nat. Prod. Res. 2021, 1–8. [Google Scholar] [CrossRef]
- Colquhoun, T.A.; Clark, D.G. Unraveling the regulation of floral fragrance biosynthesis. Plant Signal. Behav. 2011, 6, 378–381. [Google Scholar] [CrossRef]
- Jiang, T.; Zhang, M.; Wen, C.; Xie, X.; Tian, W.; Wen, S.; Lu, R.; Liu, L. Integrated metabolomic and transcriptomic analysis of the anthocyanin regulatory networks in Salvia miltiorrhiza Bge. flowers. BMC Plant Biol. 2020, 20, 349. [Google Scholar] [CrossRef]
- Tatsuzawa, F.; Yoshikoshi, A.; Takehara, A.; Suzuki, S. Flavonoids from the flowers of Adenium obesum (Forssk.) Roem. & Schult. Mandevilla sanderi (Hemsl.) Woodson, and Nerium oleander L. (Apocynaceae). Biochem. Syst. Ecol. 2021, 99, 104347. [Google Scholar]
- Jiang, L.; Yanase, E.; Mori, T.; Kurata, K.; Toyama, M.; Tsuchiya, A.; Yamauchi, K.; Mitsunaga, T.; Iwahashi, H.; Takahashi, J. Relationship between flavonoid structure and reactive oxygen species generation upon ultraviolet and X-ray irradiation. J. Photochem. Photobiol. A Chem. 2019, 384, 112044. [Google Scholar] [CrossRef]
- Gras, C.C.; Bogner, H.; Carle, R.; Schweiggert, R.M. Effect of genuine non-anthocyanin phenolics and chlorogenic acid on color and stability of black carrot (Daucus carota ssp. sativus var. atrorubens Alef.) anthocyanins. Food Res. Int. 2016, 85, 291–300. [Google Scholar] [CrossRef]
- Markham, K.R.; Gould, K.S.; Ryan, K.G. Cytoplasmic accumulation of flavonoids in flower petals and its relevance to yellow flower colouration. Phytochemistry 2001, 58, 403–413. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, D.; Liu, F.; Liu, Z.; Wang, X.; Yang, Y.; Li, S.; Li, H.; Tian, D.; Wang, L. Quercetin-derivatives paint the yellow petals of American lotus (Nelumbo lutea) and enzymatic basis for their accumulation. Hortic. Plant J. 2022; in press. [Google Scholar] [CrossRef]
- Yang, Y.; Li, B.; Feng, C.; Wu, Q.; Wang, Q.; Li, S.; Yu, X.; Wang, L. Chemical mechanism of flower color microvariation in Paeonia with yellow flowers. Hortic. Plant J. 2020, 6, 179–190. [Google Scholar] [CrossRef]
- Qiao, Q.; Si, F.; Wu, C.; Wang, J.; Zhang, A.; Tao, J.; Zhang, L.; Liu, Y.; Feng, Z. Transcriptome sequencing and flavonoid metabolism analysis in the leaves of three different cultivars of Acer truncatum. Plant Physiol. Biochem. 2022, 171, 1–13. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Zhang, M.; Wang, Y.; Deng, X.; Sun, H.; Yang, D.; Xu, L.; Song, H.; Yang, M. Color fading in lotus (Nelumbo nucifera) petals is manipulated both by anthocyanin biosynthesis reduction and active degradation. Plant Physiol. Biochem. 2022, 179, 100–107. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, X.; Chen, Y.; Gao, J.; Shi, Q.; Tian, L.; Cao, L. Combined Metabolome and Transcriptome Analyses Reveal the Flavonoids Changes and Biosynthesis Mechanisms in Different Organs of Hibiseu manihot L. Front. Plant Sci. 2022, 13, 817378. [Google Scholar] [CrossRef]
- Roriz, C.L.; Heleno, S.A.; Carocho, M.; Rodrigues, P.; Pinela, J.; Dias, M.I.; Fernandes, I.P.; Barreiro, M.F.; Morales, P.; Barros, L.; et al. Betacyanins from Gomphrena globosa L. flowers: Incorporation in cookies as natural colouring agents. Food Chem. 2020, 329, 127178. [Google Scholar] [CrossRef]
- López-Hortas, L.; Falqué, E.; Domínguez, H.; Torres, M.D. Microwave hydrodiffusion and gravity versus conventional distillation for Acacia dealbata flowers. Recovery of bioactive extracts for cosmetic purposes. J. Clean. Prod. 2020, 274, 123143. [Google Scholar] [CrossRef]
- Manzoor, M.; Singh, J.; Gani, A.; Noor, N. Valorization of natural colors as health-promoting bioactive compounds: Phytochemical profile, extraction techniques, and pharmacological perspectives. Food Chem. 2021, 362, 130141. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Rong, H.; Ye, Y.; Ni, Z.; Xu, M.; Zhang, W.; Xu, L.-A. Transcriptomic analysis of flower color variation in the ornamental crabapple (Malus spp.) half-sib family through Illumina and PacBio Sequel sequencing. Plant Physiol. Biochem. 2020, 149, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Tomitaka, M.; Uchihara, S.; Goto, A.; Sasaki, T. Species richness and flower color diversity determine aesthetic preferences of natural-park and urban-park visitors for plant communities. Environ. Sustain. Indic. 2021, 11, 100130. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.-Y.; Dillies, M.-A. SARTools: A DESeq2-and EdgeR-based R pipeline for comprehensive differential analysis of RNA-Seq data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Wang, G.; Li, J.; Li, J.; Sun, X. Analysis of Floral Color Differences between Different Ecological Conditions of Clematis tangutica (Maxim.) Korsh. Molecules 2023, 28, 462. https://doi.org/10.3390/molecules28010462
Guo X, Wang G, Li J, Li J, Sun X. Analysis of Floral Color Differences between Different Ecological Conditions of Clematis tangutica (Maxim.) Korsh. Molecules. 2023; 28(1):462. https://doi.org/10.3390/molecules28010462
Chicago/Turabian StyleGuo, Xiaozhu, Gui Wang, Juan Li, Jiang Li, and Xuemei Sun. 2023. "Analysis of Floral Color Differences between Different Ecological Conditions of Clematis tangutica (Maxim.) Korsh" Molecules 28, no. 1: 462. https://doi.org/10.3390/molecules28010462
APA StyleGuo, X., Wang, G., Li, J., Li, J., & Sun, X. (2023). Analysis of Floral Color Differences between Different Ecological Conditions of Clematis tangutica (Maxim.) Korsh. Molecules, 28(1), 462. https://doi.org/10.3390/molecules28010462