
Atractylodin Induces Apoptosis and Inhibits the Migration of A549 Lung Cancer Cells by Regulating ROS-Mediated Signaling Pathways

Abstract
1. Introduction
2. Results
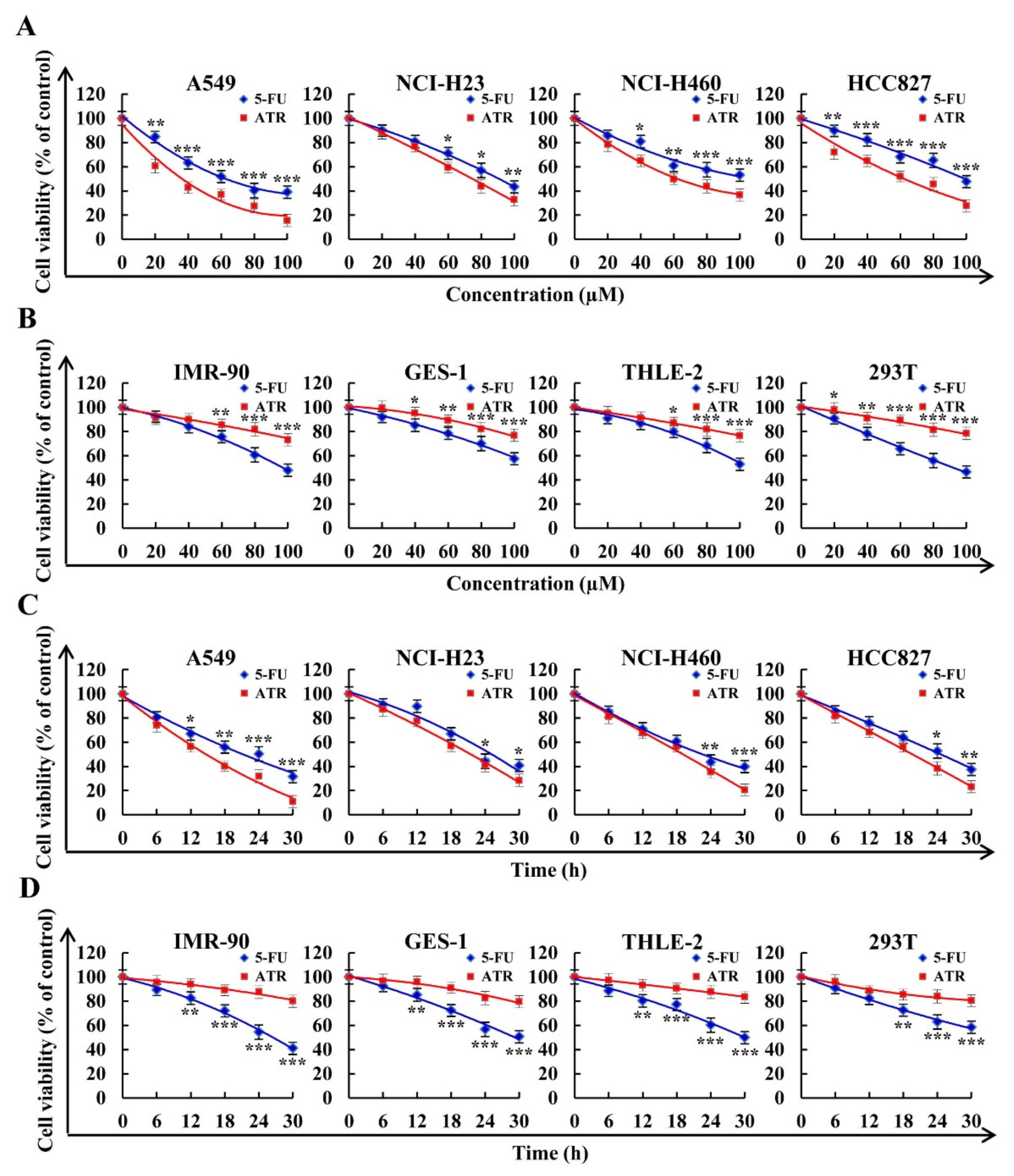
2.1. Effects of ATR on Cytotoxic Activity
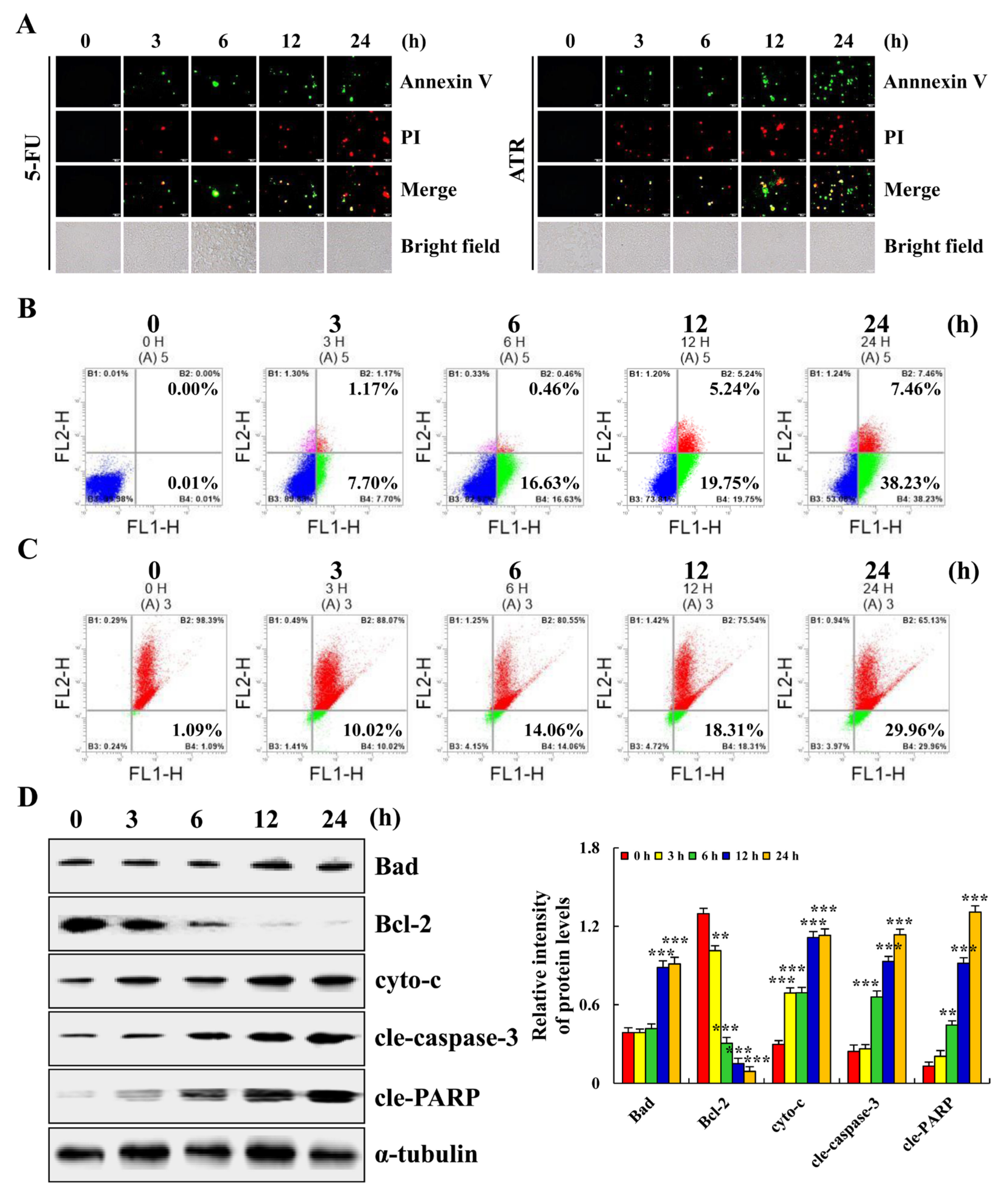
2.2. ATR Induces A549 Cell Apoptosis
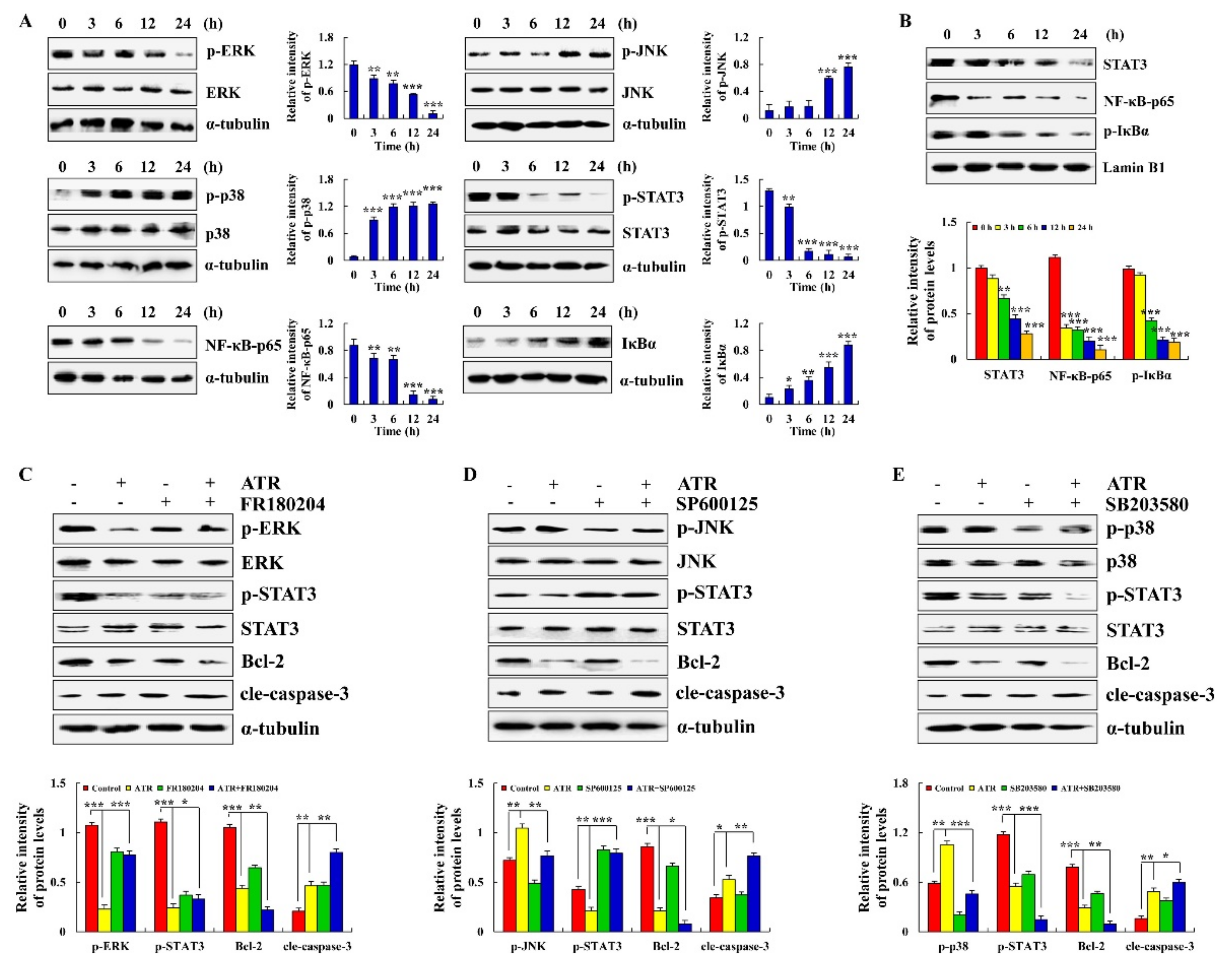
2.3. ATR Mediates MAPK, STAT3, and NF-κB Signaling Pathways to Induce A549 Cell Apoptosis
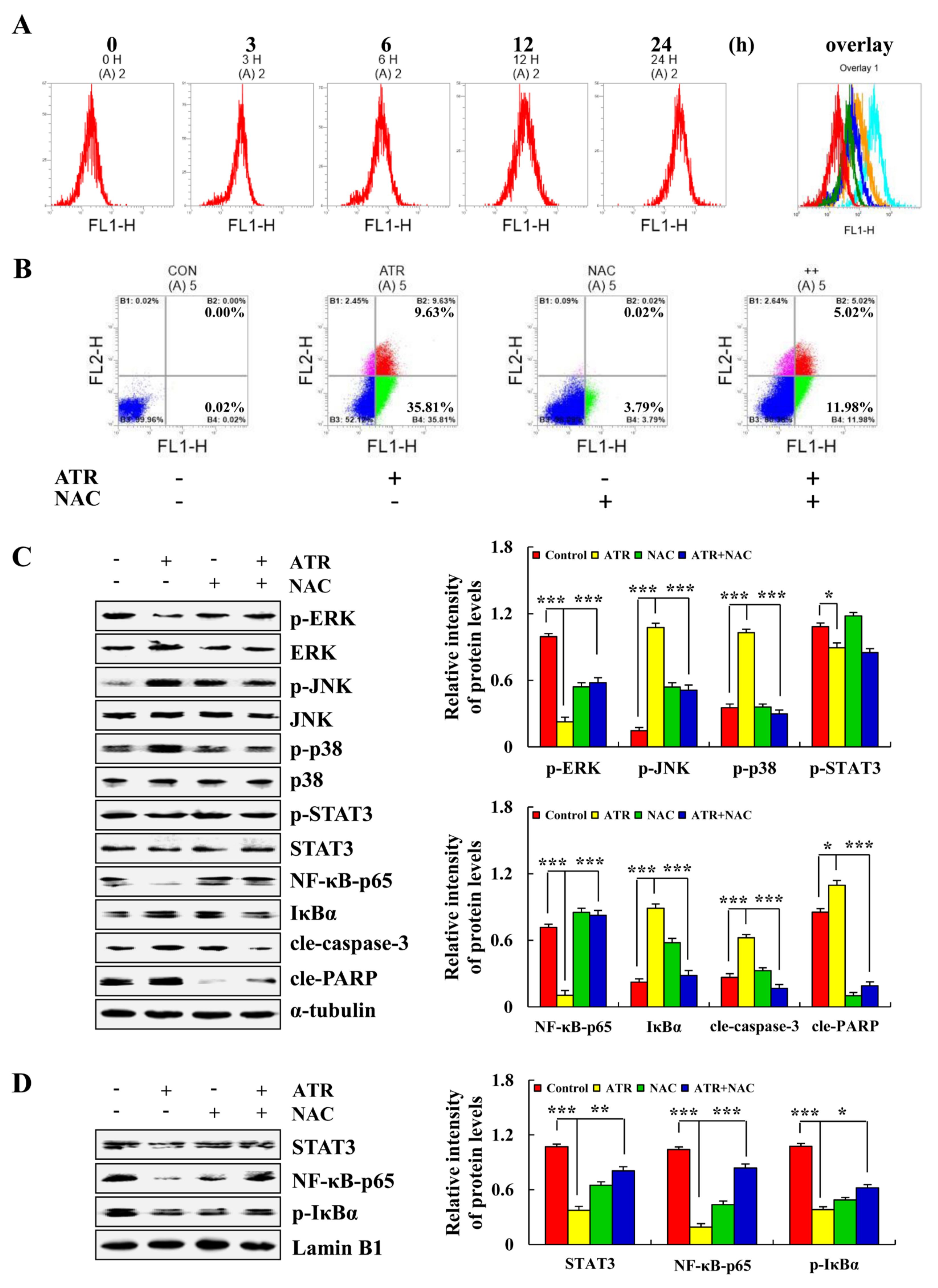
2.4. ATR Regulates MAPK, STAT3, and NF-κB Signaling Pathways by Upregulating ROS
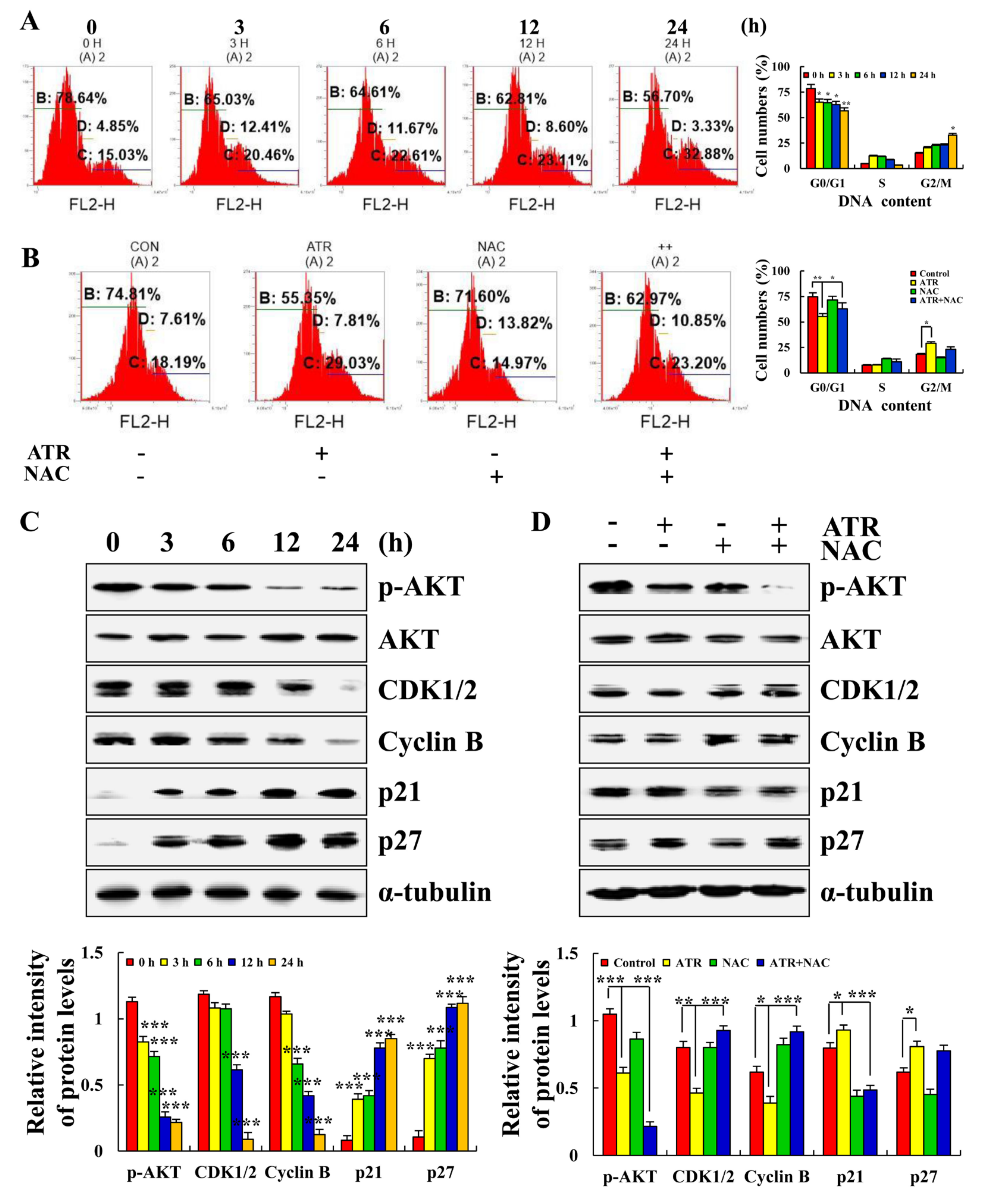
2.5. ATR Arrests A549 Cell Cycle
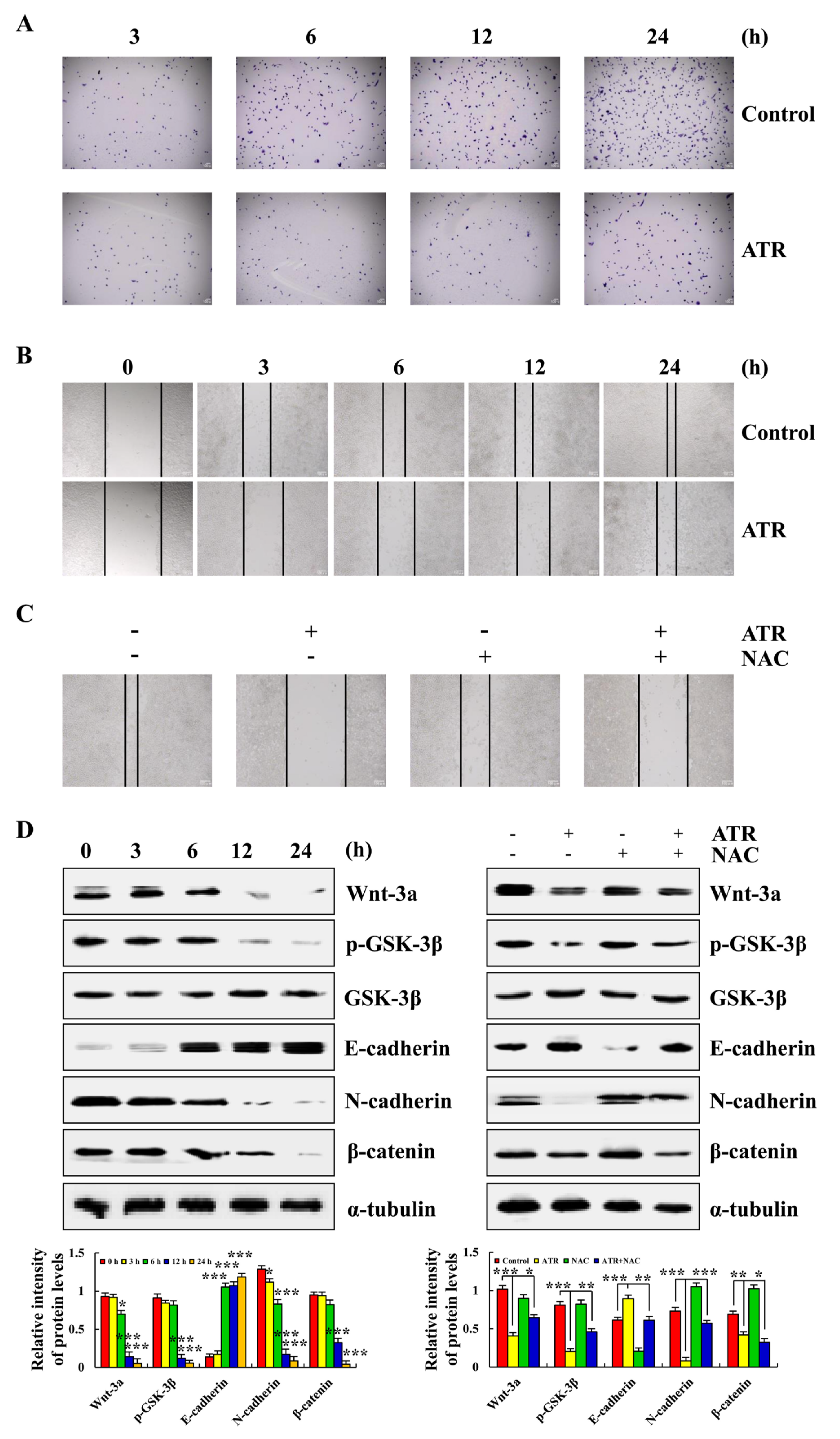
2.6. ATR Inhibits A549 Cell Migration
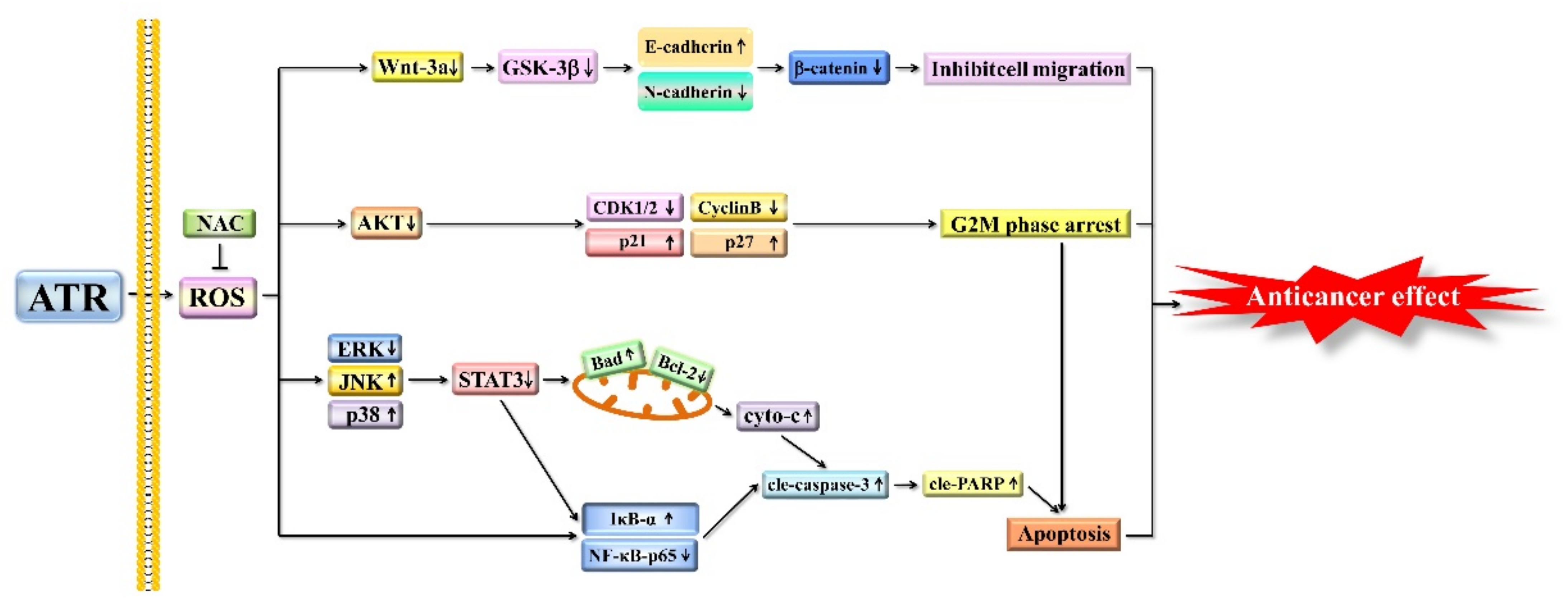
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Lines and Culture
4.3. Cytotoxic Assay
4.4. Cell Apoptosis Assay
4.5. MMP Assay
4.6. Nuclear Protein Extraction
4.7. Western Blot Analysis
4.8. ROS Accumulation Analysis
4.9. Cell Cycle Analysis
4.10. Cell Migration Analysis
4.11. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bolgova, L.S.; Tuganova, T.N.; Alekseenko, O.I.; Litvinets, O.M.; Suprun, G.A.; Ponomarenko, A.A. Histogenesis of central lung cancer: Cytological investigation. Exp. Oncol. 2020, 42, 310–313. [Google Scholar] [PubMed]
- Barta, J.A.; Powell, C.A.; Wisnivesky, J.P. Global Epidemiology of Lung Cancer. Ann. Glob. Health 2019, 85, 8. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Sun, L.; Li, J.; Yu, H.; Lin, H.; Yu, T.; Zhao, F.; Zhu, M.; Liu, L.; Geng, Q.; et al. RNA-binding protein KHSRP promotes tumor growth and metastasis in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 2019, 38, 478. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.T.; Zhang, T.; Lu, X.; Wang, R.Z. B7-H3 promotes metastasis, proliferation, and epithelial-mesenchymal transition in lung adenocarcinoma. Onco. Targets Ther. 2018, 11, 4693–4700. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.M.; Wu, S.Z.; Yang, S.H.; Wu, C.C.; Wang, C.Y.; Huang, B.M. FGF9/FGFR1 promotes cell proliferation, epithelial-mesenchymal transition, M2 macrophage infiltration and liver metastasis of lung cancer. Transl. Oncol. 2021, 14, 101208. [Google Scholar] [CrossRef]
- Wadowska, K.; Bil-Lula, I.; Trembecki, Ł.; Śliwińska-Mossoń, M. Genetic Markers in Lung Cancer Diagnosis: A Review. Int. J. Mol. Sci. 2020, 21, 4569. [Google Scholar] [CrossRef]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef]
- Wei, R.; Zhao, Y.; Wang, J.; Yang, X.; Li, S.; Wang, Y.; Yang, X.; Fei, J.; Hao, X.; Zhao, Y.; et al. Tagitinin C induces ferroptosis through PERK-Nrf2-HO-1 signaling pathway in colorectal cancer cells. Int. J. Biol. Sci. 2021, 17, 2703–2717. [Google Scholar] [CrossRef]
- Thomas-Schoemann, A.; Batteux, F.; Mongaret, C.; Nicco, C.; Chéreau, C.; Annereau, M.; Dauphin, A.; Goldwasser, F.; Weill, B.; Lemare, F.; et al. Arsenic trioxide exerts antitumor activity through regulatory T cell depletion mediated by oxidative stress in a murine model of colon cancer. J. Immunol. 2012, 189, 5171–5177. [Google Scholar] [CrossRef]
- Jiang, B.; Le, L.; Pan, H.; Hu, K.; Xu, L.; Xiao, P. Dihydromyricetin ameliorates the oxidative stress response induced by methylglyoxal via the AMPK/GLUT4 signaling pathway in PC12 cells. Brain Res. Bull. 2014, 109, 117–126. [Google Scholar] [CrossRef]
- Lake, D.; Corrêa, S.A.; Müller, J. Negative feedback regulation of the ERK1/2 MAPK pathway. Cell Mol. Life Sci. 2016, 73, 4397–4413. [Google Scholar] [CrossRef] [PubMed]
- Hodge, D.R.; Hurt, E.M.; Farrar, W.L. The role of IL-6 and STAT3 in inflammation and cancer. Eur. J. Cancer 2005, 41, 2502–2512. [Google Scholar] [CrossRef] [PubMed]
- Markov, A.V.; Zenkova, M.A.; Logashenko, E.B. Modulation of Tumour-Related Signaling Pathways by Natural Pentacyclic Triterpenoids and their Semisynthetic Derivatives. Curr. Med. Chem. 2017, 24, 1277–1320. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.T.; Mandal, J.P.; Wu, W.S. Regulation on tumor metastasis by Raf kinase inhibitory protein: New insight with reactive oxygen species signaling. Tzu Chi Med. J. 2021, 33, 332–338. [Google Scholar] [PubMed]
- Ehlting, C.; Ronkina, N.; Böhmer, O.; Albrecht, U.; Bode, K.A.; Lang, K.S.; Kotlyarov, A.; Radzioch, D.; Gaestel, M.; Häussinger, D.; et al. Distinct functions of the mitogen-activated protein kinase-activated protein (MAPKAP) kinases MK2 and MK3: MK2 mediates lipopolysaccharide-induced signal transducers and activators of transcription 3 (STAT3) activation by preventing negative regulatory effects of MK3. J. Biol. Chem. 2011, 286, 24113–24124. [Google Scholar]
- Van de Merbel, A.F.; van der Horst, G.; Buijs, J.T.; van der Pluijm, G. Protocols for Migration and Invasion Studies in Prostate Cancer. Methods Mol. Biol. 2018, 1786, 67–79. [Google Scholar]
- Najafi, S.M.A.A. The Canonical Wnt Signaling (Wnt/β-Catenin Pathway): A Potential Target for Cancer Prevention and Therapy. Iran BioMed. J. 2020, 24, 269–280. [Google Scholar] [CrossRef]
- Takahashi-Yanaga, F.; Sasaguri, T. Drug development targeting the glycogen synthase kinase-3beta (GSK-3beta)-mediated signal transduction pathway: Inhibitors of the Wnt/beta-catenin signaling pathway as novel anticancer drugs. J. Pharmacol. Sci. 2009, 109, 179–183. [Google Scholar] [CrossRef]
- Gupta, A.; Verma, A.; Mishra, A.K.; Wadhwa, G.; Sharma, S.K.; Jain, C.K. The Wnt pathway: Emerging anticancer strategies. Recent Pat. Endocr. Metab. Immune Drug Discov. 2013, 7, 138–147. [Google Scholar] [CrossRef]
- Sakanaka, C.; Sun, T.Q.; Williams, L.T. New steps in the Wnt/beta-catenin signal transduction pathway. Recent Prog. Horm. Res. 2000, 55, 225–236. [Google Scholar]
- Wong, S.H.M.; Fang, C.M.; Chuah, L.H.; Leong, C.O.; Ngai, S.C. E-cadherin: Its dysregulation in carcinogenesis and clinical implications. Crit. Rev. Oncol. Hematol. 2018, 121, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Liu, Z.Y.; Wang, P.Y.; Li, Y.J.; Wang, R.R.; Xie, S.Y. Nanoplatform-based natural products co-delivery system to surmount cancer multidrug-resistant. J. Control Release 2021, 336, 396–409. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.H.; Mao, J.W.; Tan, X.L. Research progress on the source, production, and anti-cancer mechanisms of paclitaxel. Chin. J. Nat. Med. 2020, 18, 890–897. [Google Scholar] [CrossRef]
- Mishra, A.; Dey, S. Molecular Docking Studies of a Cyclic Octapeptide-Cyclosaplin from Sandalwood. Biomolecules 2019, 9, 740. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Lin, X.; Liu, C.; Ke, C.; Zhou, Z.; Xu, K.; Cao, G.; Liu, Y. Atractylodin Attenuates Dextran Sulfate Sodium-Induced Colitis by Alleviating Gut Microbiota Dysbiosis and Inhibiting Inflammatory Response Through the MAPK Pathway. Front. Pharmacol. 2021, 12, 665376. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Ji, X.; Chen, G.; An, B. Atractylodin ameliorates lipopolysaccharide and d-galactosamine-induced acute liver failure via the suppression of inflammation and oxidative stress. Int. ImmunoPharmacol. 2019, 72, 348–357. [Google Scholar] [CrossRef]
- Bai, Y.; Zhao, Y.H.; Xu, J.Y.; Yu, X.Z.; Hu, Y.X.; Zhao, Z.Q. Atractylodin Induces Myosin Light Chain Phosphorylation and Promotes Gastric Emptying through Ghrelin Receptor. Evid. Based Complement. Altern. Med. 2017, 2017, 2186798. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, W.H.; Wang, Y.W.; Zhu, X.J.; Xiao, Y.J.; Gao, Q.W.; Li, W.; Chen, L. Positive inotropic effect of atractylodin in normal rats and its underlying mechanism. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2020, 36, 408–413. (In Chinese) [Google Scholar]
- Tshering, G.; Pimtong, W.; Plengsuriyakarn, T.; Na-Bangchang, K. Anti-angiogenic effects of beta-eudesMol. and atractylodin in developing zebrafish embryos. Comp. BioChem. Physiol. C Toxicol. Pharmacol. 2021, 243, 108980. [Google Scholar] [CrossRef]
- Muhamad, N.; Plengsuriyakarn, T.; Chittasupho, C.; Na-Bangchang, K. The Potential of Atractylodin-Loaded PLGA Nanoparticles as Chemotherapeutic for Cholangiocarcinoma. Asian Pac. J. Cancer Prev. 2020, 21, 935–941. [Google Scholar] [CrossRef]
- Shao, C.; Hu, J.P.; Yan, J.C.; Shao, S.H. Rhizoma atractylodis element influence on human colon cancer LS174T cells proliferation. J. Jiangsu Univ. 2016, 26, 480–483. [Google Scholar]
- Acharya, B.; Chaijaroenkul, W.; Na-Bangchang, K. Atractylodin inhibited the migration and induced autophagy in cholangiocarcinoma cells via PI3K/AKT/mTOR and p38MAPK signalling pathways. J. Pharm. Pharmacol. 2021, 73, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Fang, D.; Liang, T.; Pang, H.; Nong, Y.; Tang, L.; Yang, Z.; Lu, C.; Han, X.; Zhao, S.; et al. Atractylodin may induce ferroptosis of human hepatocellular carcinoma cells. Ann. Transl. Med. 2021, 9, 1535. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.W.; Zhang, X.; Lin, S.C.; Lin, Y.C.; Li, C.H.; Akhrymuk, I.; Lin, S.H.; Lin, C.C. Atractylodin Suppresses TGF-β-Mediated Epithelial-Mesenchymal Transition in Alveolar Epithelial Cells and Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice. Int. J. Mol. Sci. 2021, 22, 11152. [Google Scholar] [CrossRef]
- Kaczanowski, S. Apoptosis: Its origin, history, maintenance and the medical implications for cancer and aging. Phys. Biol. 2016, 13, 031001. [Google Scholar] [CrossRef]
- Park, J.H.; Seo, Y.H.; Jang, J.H.; Jeong, C.H.; Lee, S.; Park, B. Asiatic acid attenuates methamphetamine-induced neuroinflammation and neurotoxicity through blocking of NF-kB/STAT3/ERK and mitochondria-mediated apoptosis pathway. J. Neuroinflamm. 2017, 14, 240. [Google Scholar] [CrossRef]
- Fan, T.J.; Han, L.H.; Cong, R.S.; Liang, J. Caspase family proteases and apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef]
- Marsden, V.S.; Ekert, P.G.; Van Delft, M.; Vaux, D.L.; Adams, J.M.; Strasser, A. Bcl-2-regulated apoptosis and cytochrome c release can occur independently of both caspase-2 and caspase-9. J. Cell Biol. 2004, 165, 775–780. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, X. Cytochrome C-mediated apoptosis. Annu. Rev. BioChem. 2004, 73, 87–106. [Google Scholar] [CrossRef]
- Xu, W.T.; Shen, G.N.; Li, T.Z.; Zhang, Y.; Zhang, T.; Xue, H.; Zuo, W.B.; Li, Y.N.; Zhang, D.J.; Jin, C.H. Isoorientin induces the apoptosis and cell cycle arrest of A549 human lung cancer cells via the ROS-regulated MAPK, STAT3 and NF-κB signaling pathways. Int. J. Oncol. 2020, 57, 550–561. [Google Scholar] [CrossRef]
- Luo, Y.H.; Wang, C.; Xu, W.T.; Zhang, Y.; Zhang, T.; Xue, H.; Li, Y.N.; Fu, Z.R.; Wang, Y.; Jin, C.H. 18β-Glycyrrhetinic Acid Has Anti-Cancer Effects via Inducing Apoptosis and G2/M Cell Cycle Arrest, and Inhibiting Migration of A549 Lung Cancer Cells. Onco. Targets Ther. 2021, 14, 5131–5144. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Q.; Song, H.J.; Liu, Y.X.; Wang, Q.M. Dehydrogenation of N-Heterocycles by Superoxide Ion Generated through Single-Electron Transfer. Chemistry 2018, 24, 2065–2069. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Knaus, U.G. Oxidants in Physiological Processes. Handb. Exp. Pharmacol. 2021, 264, 27–47. [Google Scholar] [PubMed]
- Palma, F.R.; He, C.; Danes, J.M.; Paviani, V.; Coelho, D.R.; Gantner, B.N.; Bonini, M.G. Mitochondrial Superoxide Dismutase: What the Established, the Intriguing, and the Novel Reveal About a Key Cellular Redox Switch. Antioxid Redox Signal 2020, 32, 701–714. [Google Scholar] [CrossRef]
- Kirkland, R.A.; Franklin, J.L. Bax and caspases regulate increased production of mitochondria-derived reactive species in neuronal apoptosis: LACK of A role for depletion of cytochrome c from the mitochondrial electron transport chain. BioChem. Biophys. Rep. 2015, 4, 158–168. [Google Scholar] [CrossRef][Green Version]
- Murray, J.M.; Carr, A.M. Integrating DNA damage repair with the cell cycle. Curr. Opin. Cell Biol. 2018, 52, 120–125. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, Y.; Shi, Q.; Yu, Q.; Liu, C.; Feng, J.; Deng, J.; Evers, B.M.; Zhou, B.P.; Wu, Y. Stabilization of the transcription factors slug and twist by the deubiquitinase dub3 is a key requirement for tumor metastasis. Oncotarget 2017, 8, 75127–75140. [Google Scholar] [CrossRef]
- Mathema, V.B.; Chaijaroenkul, W.; Na-Bangchang, K. Cytotoxic activity and molecular targets of atractylodin in cholangiocarcinoma cells. J. Pharm. Pharmacol. 2019, 71, 185–195. [Google Scholar] [CrossRef]
- Walsh, F.S.; Barton, C.H.; Putt, W.; Moore, S.E.; Kelsell, D.; Spurr, N.; Goodfellow, P.N. N-cadherin gene maps to human chromosome 18 and is not linked to the E-cadherin gene. J. Neurochem. 1990, 55, 805–812. [Google Scholar] [CrossRef]
- Mărgineanu, E.; Cotrutz, C.E.; Cotrutz, C. Correlation between E-cadherin abnormal expressions in different types of cancer and the process of metastasis. Rev. Med. Chir. Soc. Med. Nat. Iasi 2008, 112, 432–436. [Google Scholar] [PubMed]
- Rosivatz, E.; Becker, I.; Bamba, M.; Schott, C.; Diebold, J.; Mayr, D.; Höfler, H.; Becker, K.F. Neoexpression of N-cadherin in E-cadherin positive colon cancers. Int. J. Cancer 2004, 111, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Wang, Y.; Zheng, Y.W.; Fei, L.R.; Shen, H.Y.; Li, Z.H.; Huang, W.J.; Yu, J.H.; Xu, H.T. Overexpression of Nemo-like Kinase Promotes the Proliferation and Invasion of Lung Cancer Cells and Indicates Poor Prognosis. Curr. Cancer Drug Targets 2019, 19, 674–680. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | 5-FU (µM) | ATR (µM) |
|---|---|---|
| A549 | 64.24 ± 2.58 | 37.92 ± 1.59 |
| NCI-N23 | 90.21 ± 1.44 | 76.88 ± 2.21 |
| NCI-H460 | 108.36 ± 2.59 | 63.27 ± 1.48 |
| HCC827 | 99.62 ± 2.41 | 61.05 ± 1.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Li, S.-M.; Li, Y.-N.; Cao, J.-L.; Xue, H.; Wang, C.; Jin, C.-H. Atractylodin Induces Apoptosis and Inhibits the Migration of A549 Lung Cancer Cells by Regulating ROS-Mediated Signaling Pathways. Molecules 2022, 27, 2946. https://doi.org/10.3390/molecules27092946
Zhang T, Li S-M, Li Y-N, Cao J-L, Xue H, Wang C, Jin C-H. Atractylodin Induces Apoptosis and Inhibits the Migration of A549 Lung Cancer Cells by Regulating ROS-Mediated Signaling Pathways. Molecules. 2022; 27(9):2946. https://doi.org/10.3390/molecules27092946
Chicago/Turabian StyleZhang, Tong, Shu-Mei Li, Yan-Nan Li, Jing-Long Cao, Hui Xue, Chang Wang, and Cheng-Hao Jin. 2022. "Atractylodin Induces Apoptosis and Inhibits the Migration of A549 Lung Cancer Cells by Regulating ROS-Mediated Signaling Pathways" Molecules 27, no. 9: 2946. https://doi.org/10.3390/molecules27092946
APA StyleZhang, T., Li, S.-M., Li, Y.-N., Cao, J.-L., Xue, H., Wang, C., & Jin, C.-H. (2022). Atractylodin Induces Apoptosis and Inhibits the Migration of A549 Lung Cancer Cells by Regulating ROS-Mediated Signaling Pathways. Molecules, 27(9), 2946. https://doi.org/10.3390/molecules27092946