
Bioactivity of Two Polyphenols Quercetin and Fisetin against Human Gastric Adenocarcinoma AGS Cells as Affected by Two Coexisting Proteins


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Kits
2.2. Cell Culture
2.3. Assay of Cell Viability
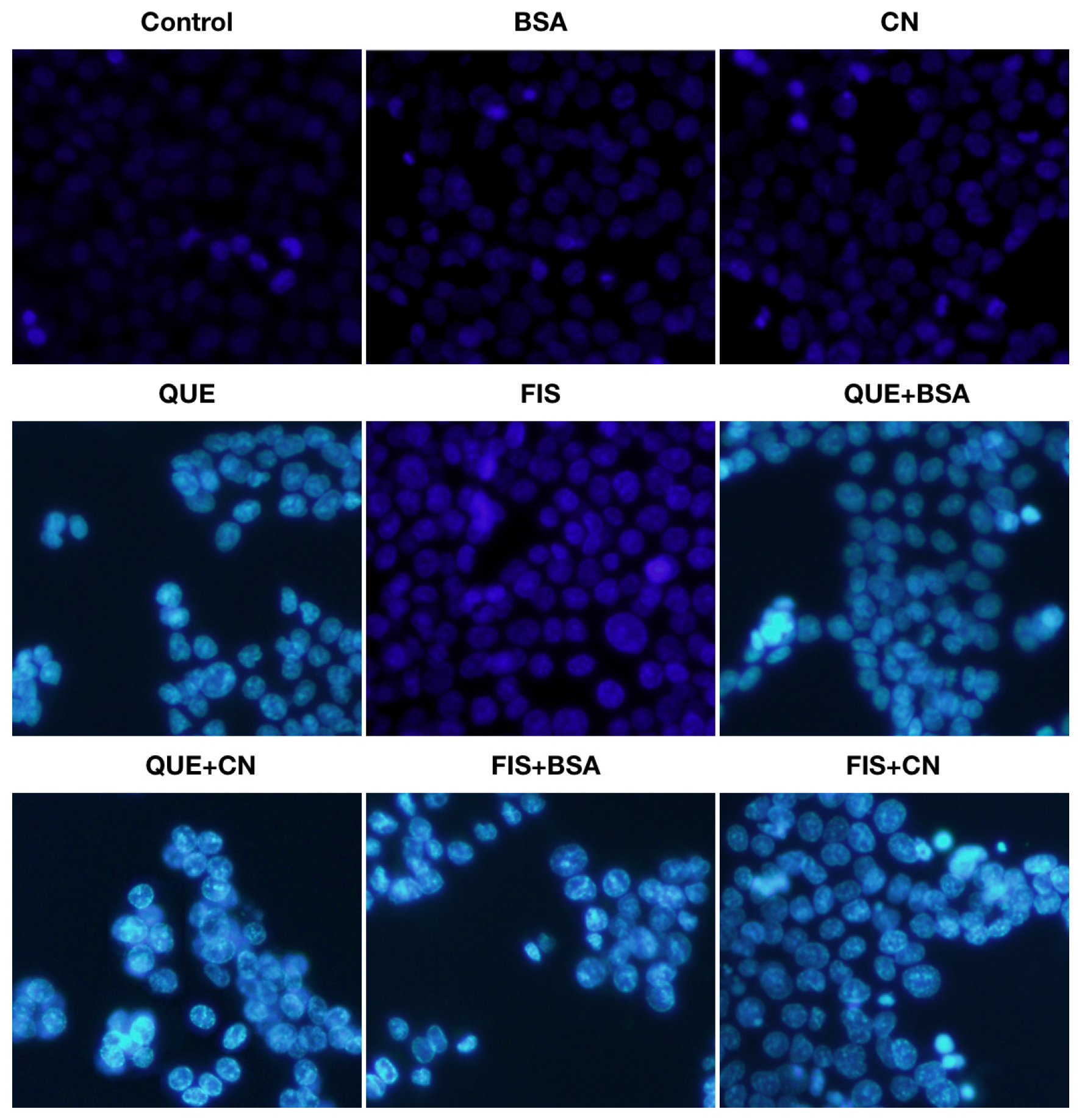
2.4. Morphology Assessment
2.5. Measurement of Mitochondrial Membrane Potential
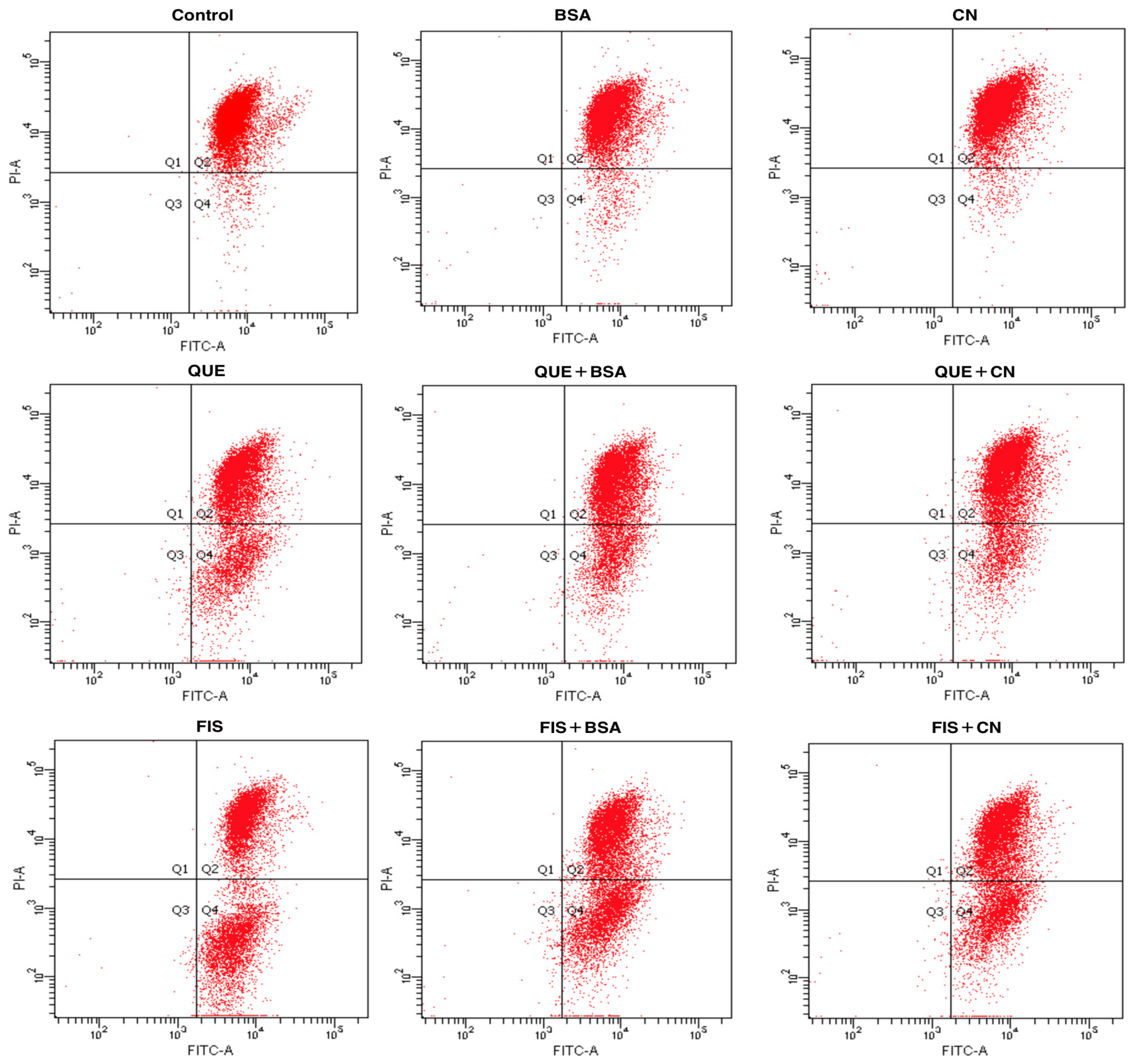
2.6. Cell Apoptosis Assay
2.7. Measurement of Intracellular ROS
2.8. Statistical Analysis
3. Results
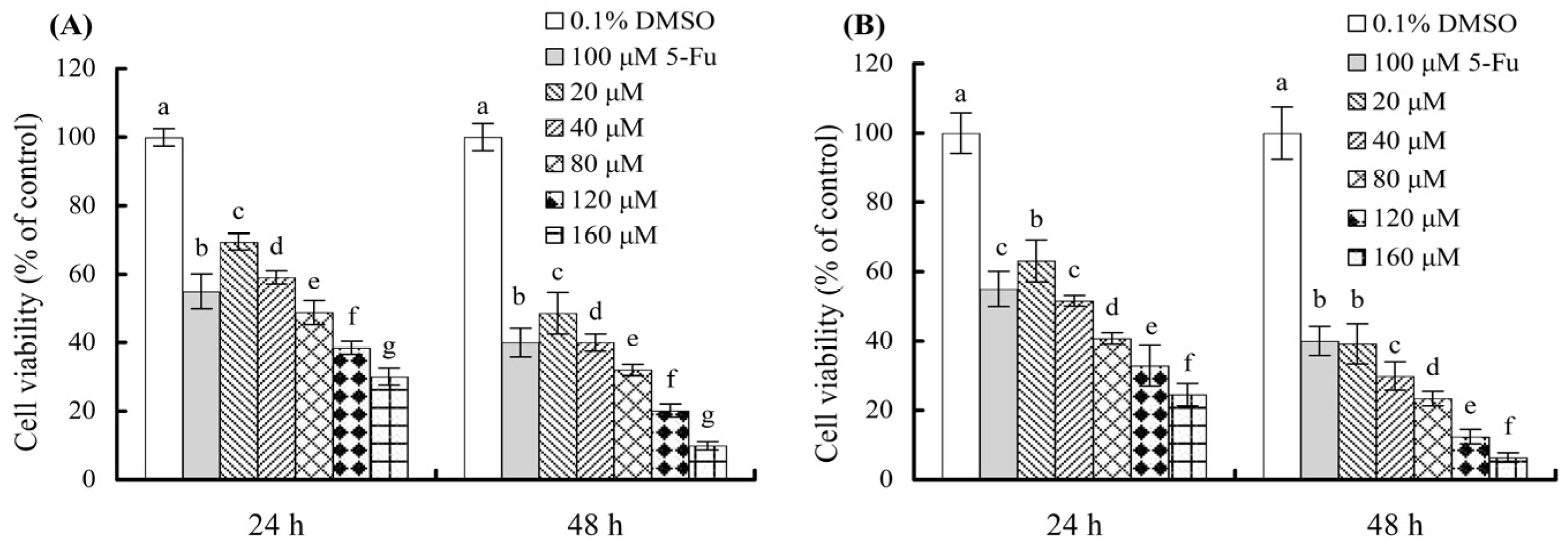
3.1. Growth Inhibition of the Polyphenols on the Cells
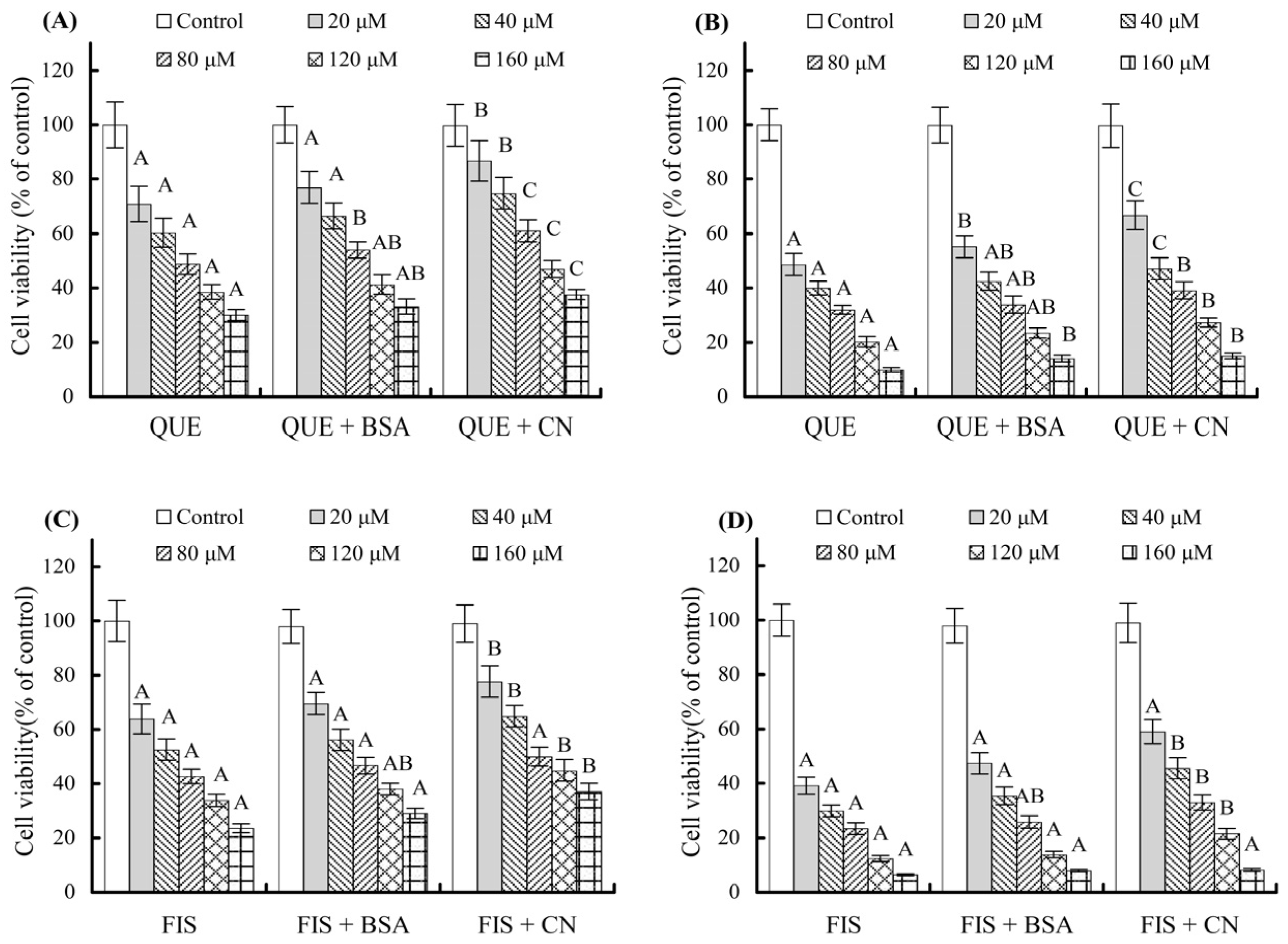
3.2. Growth Inhibition of the Polyphenols on the Cells as Affected by the Coexisting Proteins
3.3. MMP Loss of the Cells as Affected by the Two Polyphenols and Coexisting Proteins
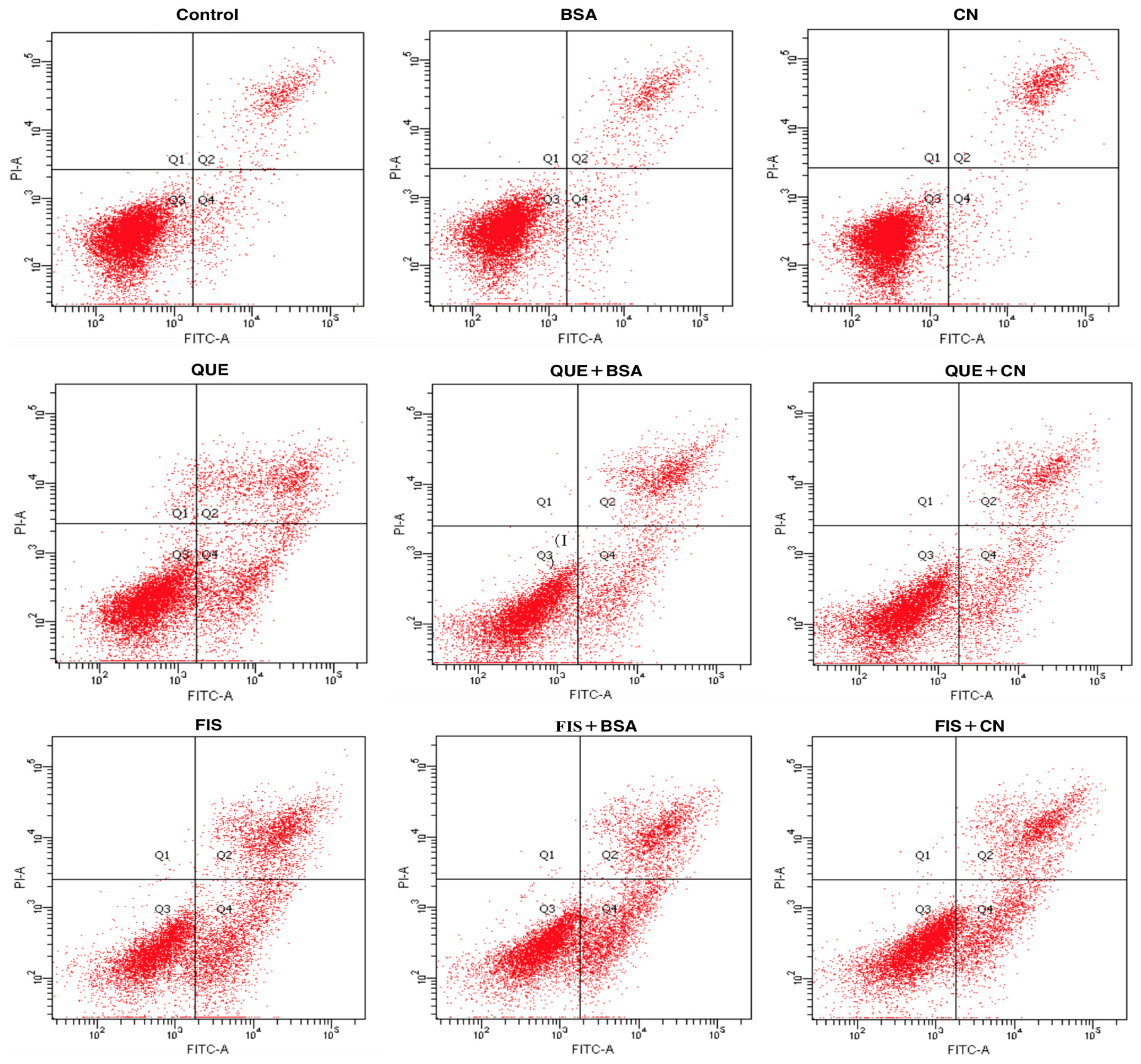
3.4. The Two Proteins Attenuated Apoptosis Induction of the Two Polyphenols in AGS Cells
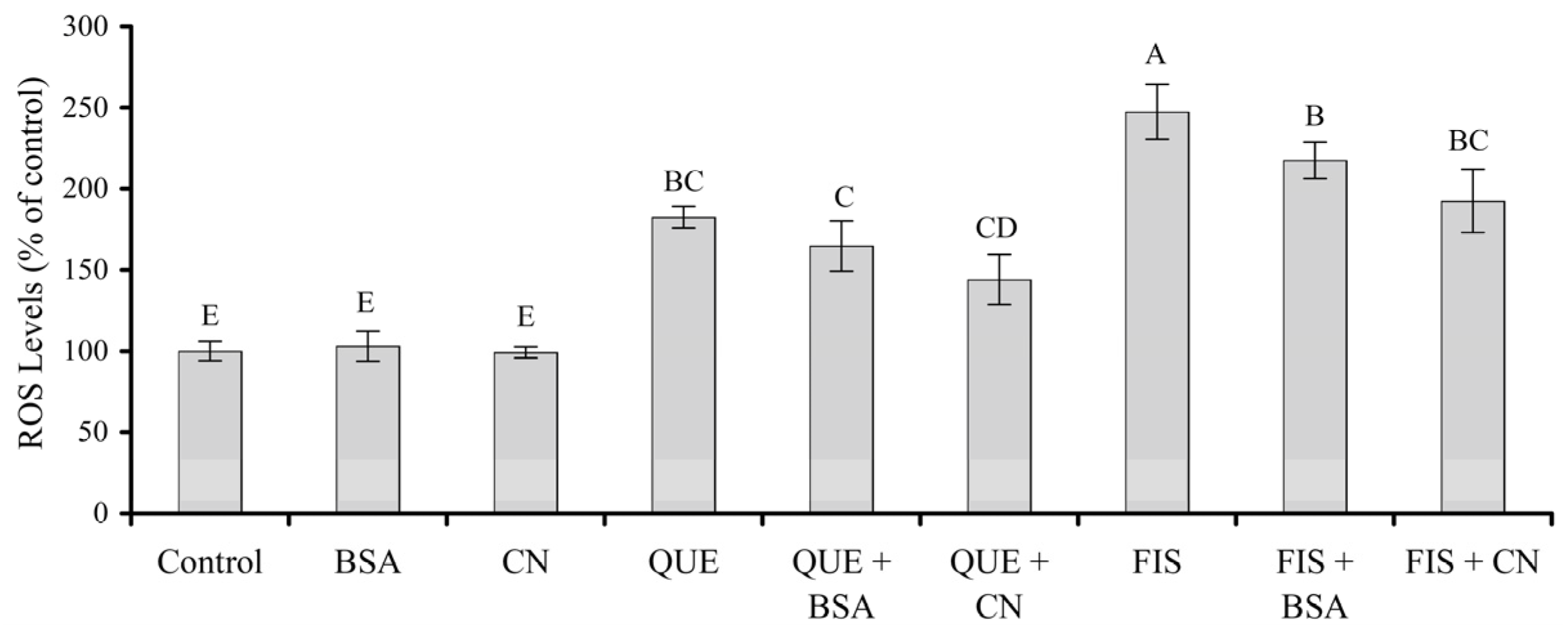
3.5. Prooxidation of the Two Polyphenols in the Cells as Affected by the Two Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bartolomé, B.; Estrella, I.; Hernandez, M.T. Interaction of low molecular weight phenolics with proteins (BSA). J. Food Sci. 2010, 65, 617–621. [Google Scholar] [CrossRef]
- Kosińska, A.; Karamać, M.; Penkacik, K.; Urbalewicz, A.; Amarowicz, R. Interactions between tannins and proteins isolated from broad bean seeds (Vicia faba Major) yield soluble and non-soluble complexes. Eur. Food Res. Technol. 2011, 233, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Wagdy, S.M.; Taha, F.S. Relation between binding of proteins with phenolics at different pH values in peanut meal. World Appl. Sci. J. 2014, 32, 207–213. [Google Scholar]
- Xiao, J.B.; Huo, J.L.; Yang, F.; Chen, X.Q. Noncovalent interaction of dietary polyphenols with bovine hemoglobin in vitro: Molecular structure/property-affinity relationship aspects. J. Agric. Food Chem. 2011, 59, 8484–8490. [Google Scholar] [CrossRef] [PubMed]
- Gallo, M.; Vinci, G.; Graziani, G.; Simone, C.D.; Ferranti, P. The interaction of cocoa polyphenols with milk proteins studied by proteomic techniques. Food Res. Int. 2013, 54, 406–415. [Google Scholar] [CrossRef]
- Gouda, M.; Moustafa, A.; Hussein, L.; Hamza, M. Three week dietary intervention using apricots, pomegranate juice or/and fermented sour sobya and impact on biomarkers of antioxidative activity, oxidative stress and erythrocytic glutathione transferase activity among adults. Nutr. J. 2016, 15, e52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazo, M.K.; Petrov, N.A.; Sarkisyan, V.A.; Kochetkova, A.A. The interaction of polyphenols with food proteins: Prospects for diet therapy of metabolic syndrome and type 2 diabetes. Problemy Endokrinologii 2018, 64, 252–257. [Google Scholar] [CrossRef]
- Cai, S.Q.; Zhang, Q.; Zhao, X.H.; Shi, J. The in vitro anti-inflammatory activities of galangin and quercetin towards the LPS-injured rat intestinal epithelial (IEC-6) cells as affected by heat treatment. Molecules 2021, 26, 7495. [Google Scholar] [CrossRef] [PubMed]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends. Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef]
- Su, Q.; Mei, P.; Zhang, Y.; Xu, W.; Yang, X. Quercetin induces bladder cancer cells apoptosis by activation of AMPK signaling pathway. Am. J. Cancer Res. 2016, 6, 498–508. [Google Scholar]
- Seo, H.S.; Ku, J.; Choi, H.S.; Choi, Y.; Woo, J.K.; Kim, M.; Kim, I.; Na, C.; Hur, H.; Jang, B. Quercetin induces caspase-dependent extrinsic apoptosis through inhibition of signal transducer and activator of transcription 3 signaling in HER2-overexpressing BT-474 breast cancer cells. Oncol. Rep. 2016, 36, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, S. Fisetin inhibits the growth and migration in the A549 human lung cancer cell line via the ERK1/2 pathway. Exp. Ther. Med. 2017, 15, 2667–2673. [Google Scholar] [CrossRef] [Green Version]
- Min, K.J.; Ju-Ock, N.; Taeg, K. Fisetin Induces apoptosis through p53-mediated up-regulation of DR5 expression in human renal carcinoma caki cells. Molecules 2017, 22, 1285. [Google Scholar] [CrossRef] [Green Version]
- Ekström, A.M.; Serafini, M.; Nyrén, O.; Wolk, A.; Bosetti, C.; Bellocco, R. Dietary quercetin intake and risk of gastric cancer: Results from a population-based study in Sweden. Ann. Oncol. 2011, 22, 438–443. [Google Scholar] [CrossRef]
- Yan, W.X.; Chen, S.H.; Zhao, Y.Y.; Ye, X.Y. Fisetin inhibits the proliferation of gastric cancer cells and induces apoptosis through suppression of ERK 1/2 activation. Oncol. Lett. 2018, 15, 8442–8446. [Google Scholar] [CrossRef]
- Sabarwal, A.; Agarwal, R.; Singh, R.P. Fisetin inhibits cellular proliferation and induces mitochondria-dependent apoptosis in human gastric cancer cells. Mol. Carcinog. 2016, 56, 499–514. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zhang, N.; Zhao, X.H. The non-covalent interaction between two polyphenols and caseinate as affected by two types of enzymatic protein crosslinking. Food Chem. 2021, 364, e130375. [Google Scholar] [CrossRef]
- Al-Hanish, A.; Stanic-Vucinic, D.; Mihailovic, J.; Prodic, I.; Minic, S.; Stojadinovic, M.; Radibratovic, M.; Milcic, M.; Cirkovic Velickovic, T. Noncovalent interactions of bovine α-lactalbumin with green tea polyphenol, epigalocatechin-3-gallate. Food Hydrocoll. 2016, 61, 241–250. [Google Scholar] [CrossRef]
- Tapal, A.; Tiku, P.K. Complexation of curcumin with soy protein isolate and its implications on solubility and stability of curcumin. Food Chem. 2012, 130, 960–965. [Google Scholar] [CrossRef]
- Namasivayam, S.K.R.; Raj, L.F.A.A.; Robin, A.T.G.; Chandra, J.H.; Bharani, R.S.A. Optimal synthesis of biocompatible bovine serum nanoparticles-incorporated quercetin (BSA NPS-QT) nano drug conjugate for the controlled release and improved anti oxidative activity. Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 478–487. [Google Scholar]
- Dubeau, S.; Samson, G.; Tajmir-Riahi, H.A. Dual effect of milk on the antioxidant capacity of green, Darjeeling, and English breakfast teas. Food Chem. 2010, 122, 539–545. [Google Scholar] [CrossRef]
- Guri, A.; Haratifar, S.; Corredig, M. Bioefficacy of tea catechins associated with milk caseins tested using different in vitro digestion models. Food Digest. 2014, 5, 8–18. [Google Scholar] [CrossRef]
- Gou, Y.; Zhang, Z.; Qi, J.; Liang, S.; Zhou, Z. Folate-functionalized human serum albumin carrier for anticancer copper (II) complexes derived from natural plumbagin. J. Inorg. Biochem. 2015, 153, 13–22. [Google Scholar] [CrossRef]
- Xiao, X.; Yi, G.; Yong, G.; Zhao, L.; Chu, J.; Zhou, J.; Cai, X.; Zhang, H.; Xu, L.; Nie, Y. Osteopontin contributes to TGF-β1 mediated hepatic stellate cell activation. Digest. Dis. Sci. 2012, 57, 2883–2891. [Google Scholar] [CrossRef]
- Kristiani, E.B.K.; Nugroho, L.H.; Moeljopawiro, S.; Widyarini, S. The cytotoxicity of mekai (Albertisia papuana Becc.) root extract on breast cancer cell lines T47D and Vero cell lines. AIP Conf. Proc. 2016, 1744, e020016. [Google Scholar]
- Smiley, S.T.; Reers, M.; Mottola-Hartshorn, C.; Mei, L.; Chen, A.; Smith, T.W.; Steele, G.D.; Chen, A.L.B. Intracellular heterogeneity in mitochondrial membrane potentials revealed by a J-aggregate-forming lipophilic cation JC-1. Proc. Natl. Acad. Sci. USA 1991, 88, 3671–3675. [Google Scholar] [CrossRef] [Green Version]
- Span, L.F.R.; Pennings, A.H.M.; Vierwinden, G.; Boezeman, J.B.M.; Raymakers, R.A.P.; Witte, T.D. The dynamic process of apoptosis analyzed by flow cytometry using Annexin-V/Propidium iodide and a modified in situ end labeling technique. Cytometry 2002, 47, 24–31. [Google Scholar] [CrossRef]
- Aranda, A.; Sequedo, L.; Tolosa, L.; Quintás, G.; Gombau, L. Dichloro-dihydro-fluorescein diacetate (DCFH-DA) assay: A quantitative method for oxidative stress assessment of nanoparticle-treated cells. Toxicol. In Vitro 2013, 27, 954–963. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, W.; Soladoye, O.P. Towards innovative food processing of flavonoid compounds: Insights into stability and bioactivity. LWT Food Sci. Technol. 2021, 150, e111968. [Google Scholar] [CrossRef]
- Bi, H.; Tang, L.; Gao, X.; Jia, J.; Lv, H. Spectroscopic analysis on the binding interaction between tetracycline hydrochloride and bovine proteins β-casein, α-lactalbumin. J. Lumin. 2016, 178, 72–83. [Google Scholar] [CrossRef]
- Arts, M.J.T.J.; Haenen, G.R.M.M.; Voss, H.P.; Bast, A. Masking of antioxidant capacity by the interaction of flavonoids with protein. Food Chem. Toxicol. 2001, 39, 787–791. [Google Scholar] [CrossRef]
- Geng, R.; Ma, L.; Liu, L.; Xie, Y. Influence of bovine serum albumin-flavonoid interaction on the antioxidant activity of dietary flavonoids: New evidence from electrochemical quantification. Molecules 2018, 24, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhao, X.H. Degradation kinetics of fisetin and quercetin in solutions affected by medium pH, temperature and coexisted proteins. J. Serb. Chem. Soc. 2016, 81, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Wang, S.; Feng, B.; Jiang, B.; Miao, M. Interaction between soybean protein and tea polyphenols under high pressure. Food Chem. 2019, 277, 632–638. [Google Scholar] [CrossRef]
- Hoskin, R.T.; Xiong, J.; Esposito, D.A.; Lila, M.A. Blueberry polyphenol-protein food ingredients: The impact of spray drying on the in vitro antioxidant activity, anti-inflammatory markers, glucose metabolism and fibroblast migration. Food Chem. 2019, 280, 187–194. [Google Scholar] [CrossRef]
- Von Staszewski, M.; Jagus, R.J.; Pilosof, A.M.R. Influence of green tea polyphenols on the colloidal stability and gelation of WPC. Food Hydrocoll. 2011, 25, 1077–1084. [Google Scholar] [CrossRef]
- Xiao, J.B.; Mao, F.F.; Yang, F.; Zhao, Y.L.; Zhang, C.; Yamamoto, K. Interaction of dietary polyphenols with bovine milk proteins: Molecular structure-affinity relationship and influencing bioactivity aspects. Mol. Nutr. Food Res. 2011, 55, 1637–1645. [Google Scholar] [CrossRef]
- Ma, C.M.; Zhao, J.R.; Zhao, X.H. The non-covalent interacting forces and scavenging activities to three free radicals involved in the caseinate-flavonol (kaempferol and quercetin) complexes. J. Food Meas. Charact. 2021, 16, 114–125. [Google Scholar] [CrossRef]
- Ma, C.M.; Zhao, X.H. Depicting the non-covalent interaction of whey proteins with galangin or genistein by the multi-spectroscopic techniques and molecular docking. Foods 2019, 8, 360. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.M.; Zhao, X.H. The non-covalent interaction and in vitro radical scavenging activities of the caseinate-galangin and caseinate-genistein complexes. Antioxidants 2019, 8, 354. [Google Scholar] [CrossRef] [Green Version]
- Farkye, N.Y.; Shah, N. Milk proteins. In Applied Food Protein Chemistry; Ustunol, Z., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015; pp. 427–458. [Google Scholar]
- Hasni, I.; Bourassa, P.; Hamdani, S.; Samson, G.; Carpentier, R.; Tajmir-Riahi, H.A. Interaction of milk α- and β-caseins with tea polyphenols. Food Chem. 2011, 126, 630–639. [Google Scholar] [CrossRef]
- Pawar, S.R.; Jangam, S.; Waghmare, S. Anti-cancer herbal drugs: An overview. J. Drug Deliv. Ther. 2018, 8, 48–58. [Google Scholar]
- Altundağ, E.M.; Kasacı, T.; Yılmaz, A.M.; Karademir, B.; Koçtürk, S.; Taga, Y.; Yalçın, A.S. Quercetin-induced cell death in human papillary thyroid cancer (B-CPAP) cells. J. Thyroid. Res. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Ma, X.M.; Li, Q.W.; Yang, Y.; Xu, X.L.; Sun, J.Q.; Yu, M.W.; Cao, K.X.; Yang, L.; Yang, G.W.; et al. Anti-cancer effects of fisetin on mammary carcinoma cells via regulation of the PI3K/Akt/mTOR pathway: In vitro and in vivo studies. Int J. Mol. Med. 2018, 42, 811–820. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, M.H.; Tsai, J.P.; Yang, S.F.; Chiou, H.F.; Lin, C.L.; Hsieh, Y.H.; Chang, H.R. Fisetin suppresses the proliferation and metastasis of renal cell carcinoma through upregulation of MEK/ERK-targeting CTSS and ADAM9. Cells 2019, 8, 948. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, H. Structure-dependent membrane interaction of flavonoids associated with their bioactivity. Food Chem. 2010, 120, 1089–1096. [Google Scholar] [CrossRef]
- Fan, J.; Zhao, X.H.; Li, T.J. Heat treatment of galangin and kaempferol inhibits their benefits to improve barrier function in rat intestinal epithelial cells. J. Nutr. Biochem. 2021, 87, e108517. [Google Scholar] [CrossRef]
- Fan, J.; Li, T.J.; Zhao, X.H. Barrier-promoting efficiency of two bioactive flavonols quercetin and myricetin on rat intestinal epithelial (IEC-6) cells via suppressing Rho activation. RSC Adv. 2020, 10, 27249–27258. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Time | Quercetin | Fisetin | ||
|---|---|---|---|---|
| Without Proteins | With Proteins | Without Proteins | With Proteins | |
| 24 h | 64.2 ± 4.8 | 69.3 ± 3.2 (BSA) 79.6 ± 2.6 (CN) | 43.4 ± 3.7 | 48.7 ± 4.4 (BSA) 52.1 ± 2.8 (CN) |
| 48 h | 21.4 ± 1.9 | 25.8 ± 1.4 (BSA) 31.4 ± 3.6 (CN) | 12.8 ± 1.1 | 17.1 ± 4.9 (BSA) 22.4 ± 2.5 (CN) |
| Cell Group | Q3 (%) | Q2 + Q4 (%) |
|---|---|---|
| 0.1% DMSO | 88.7 | 10.2 |
| BSA | 88.9 | 11.1 |
| Casein | 88.7 | 11.3 |
| Quercetin | 66.5 | 32.3 |
| Quercetin + BSA | 74.5 | 25.5 |
| Quercetin + CN | 71.2 | 28.7 |
| Fisetin | 49.9 | 50.0 |
| Fisetin + BSA | 54.9 | 45.0 |
| Fisetin + CN | 61.9 | 38.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, B.; Wang, J.; Zhao, X.-H. Bioactivity of Two Polyphenols Quercetin and Fisetin against Human Gastric Adenocarcinoma AGS Cells as Affected by Two Coexisting Proteins. Molecules 2022, 27, 2877. https://doi.org/10.3390/molecules27092877
Wang B, Wang J, Zhao X-H. Bioactivity of Two Polyphenols Quercetin and Fisetin against Human Gastric Adenocarcinoma AGS Cells as Affected by Two Coexisting Proteins. Molecules. 2022; 27(9):2877. https://doi.org/10.3390/molecules27092877
Chicago/Turabian StyleWang, Bo, Jing Wang, and Xin-Huai Zhao. 2022. "Bioactivity of Two Polyphenols Quercetin and Fisetin against Human Gastric Adenocarcinoma AGS Cells as Affected by Two Coexisting Proteins" Molecules 27, no. 9: 2877. https://doi.org/10.3390/molecules27092877
APA StyleWang, B., Wang, J., & Zhao, X.-H. (2022). Bioactivity of Two Polyphenols Quercetin and Fisetin against Human Gastric Adenocarcinoma AGS Cells as Affected by Two Coexisting Proteins. Molecules, 27(9), 2877. https://doi.org/10.3390/molecules27092877