
Identification of Potential Anti-Neuroinflammatory Inhibitors from Antarctic Fungal Strain Aspergillus sp. SF-7402 via Regulating the NF-κB Signaling Pathway in Microglia

 , , and
, , and 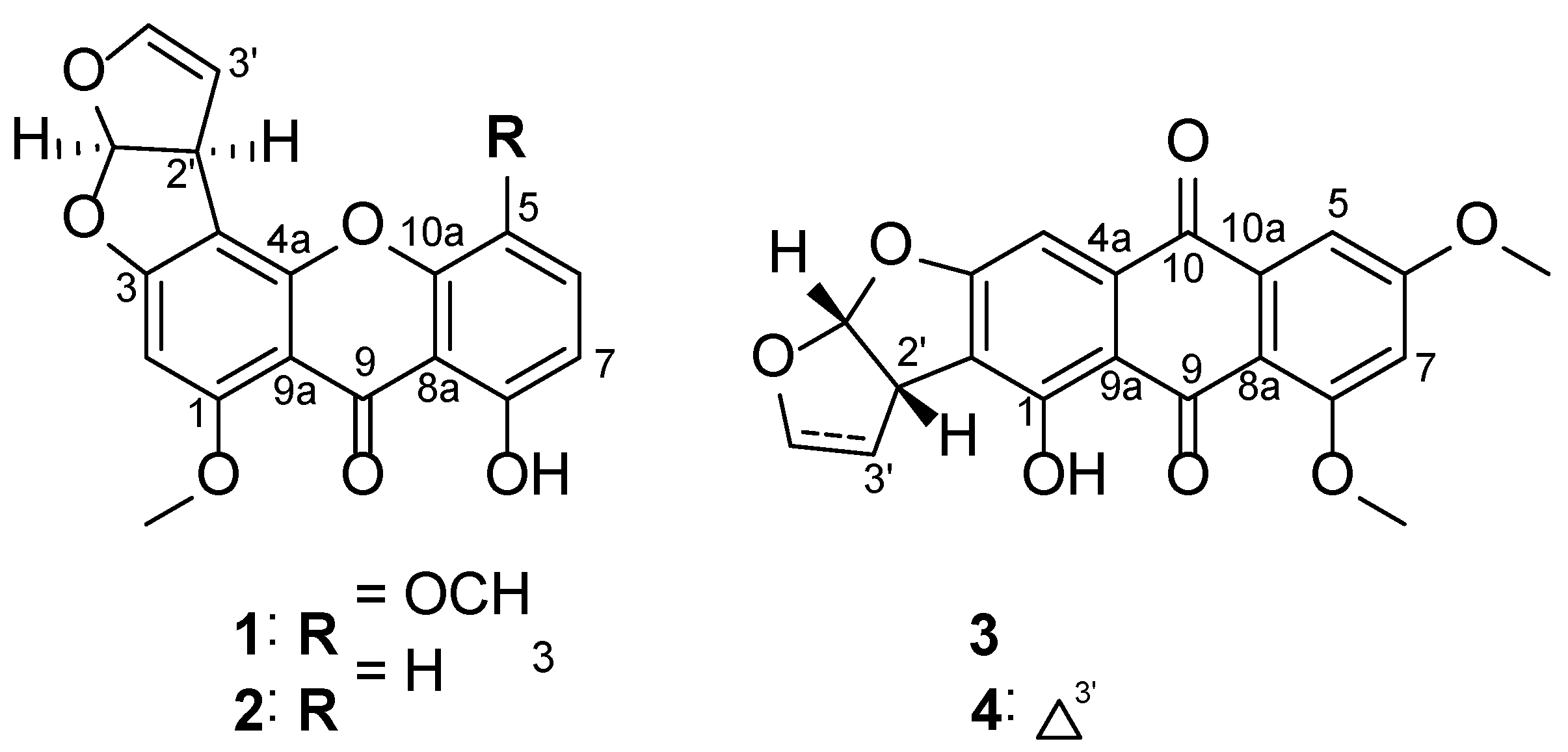{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Fungal Material and Fermentation
2.3. Extraction and Isolation of Metabolites
2.3.1. 5-Methoxysterigmatocystin (1)
2.3.2. Sterigmatocystin (2)
2.3.3. Aversin (3)
2.3.4. 6,8-O-dimethylversicolorin A (4)
2.4. Cell Culture and Reagents
2.5. Cell Viability Assay
2.6. Measurement of Nitrite Oxide (NO) Generation
2.7. Assays for IL-6 and TNF-α Production
2.8. Determination of Prostaglandin E2 (PGE2) Levels
2.9. Analysis of iNOS and COX-2 Protein Expression
2.10. Analysis of NF-κB (p65) Expressions
2.11. Statistical Analysis
3. Results
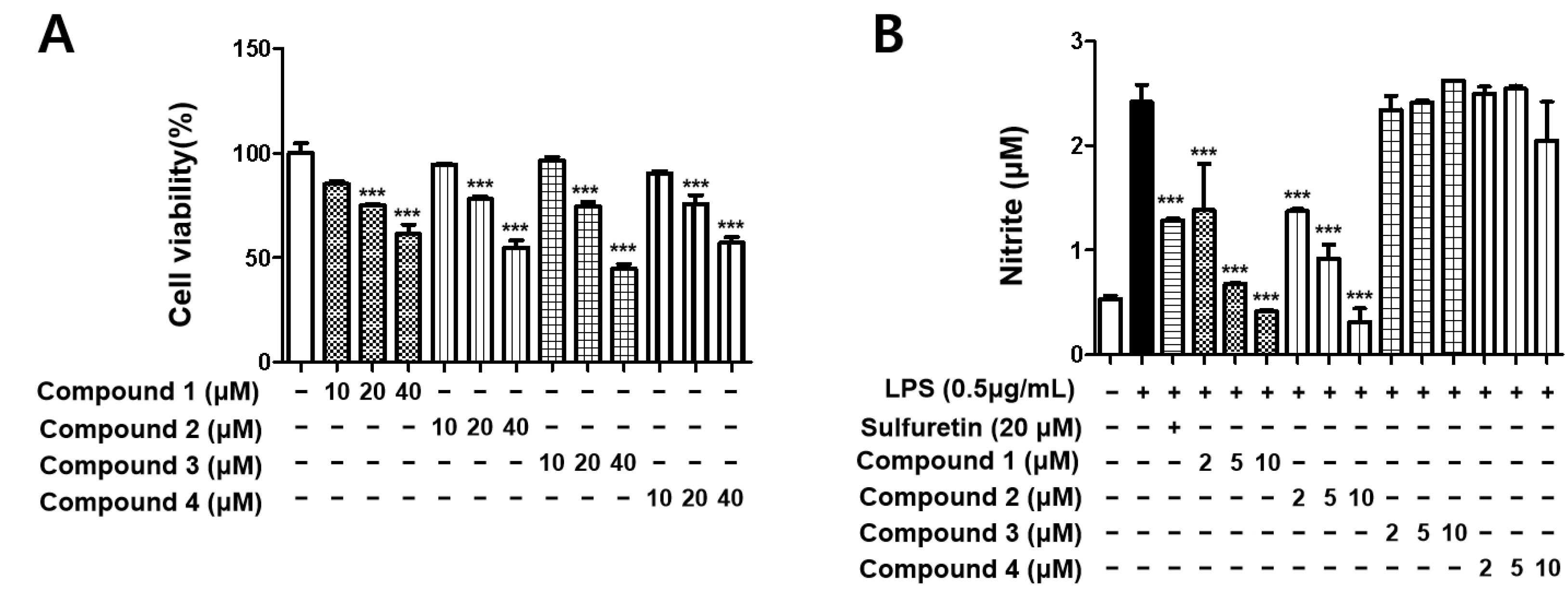
3.1. Structure Elucidation and Effects of Isolated Metabolites on BV2 Cells Viability
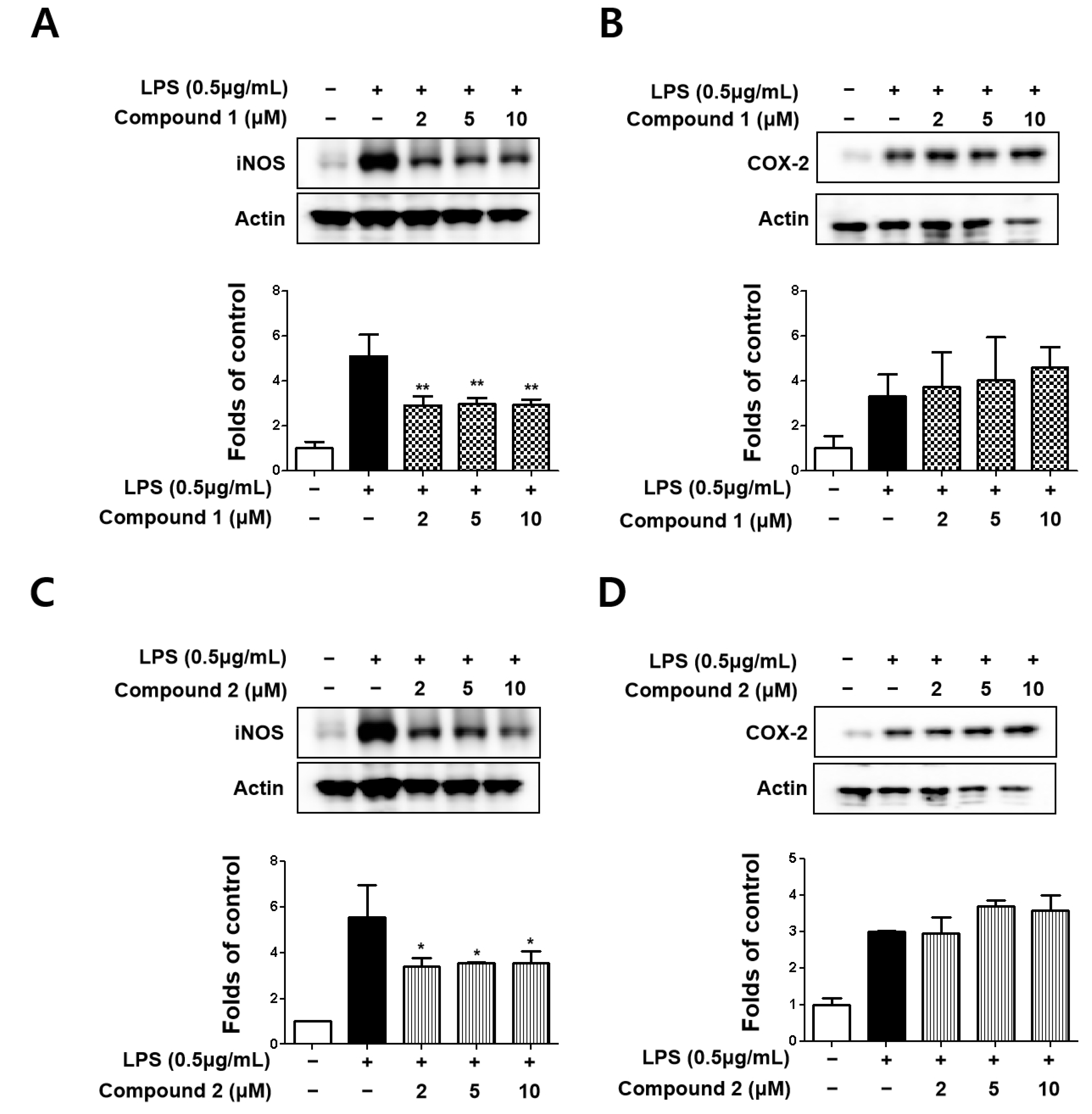
3.2. Sterigmatocystins Inhibited LPS-induced Expression of iNOS and COX-2 Proteins in BV2 Cells
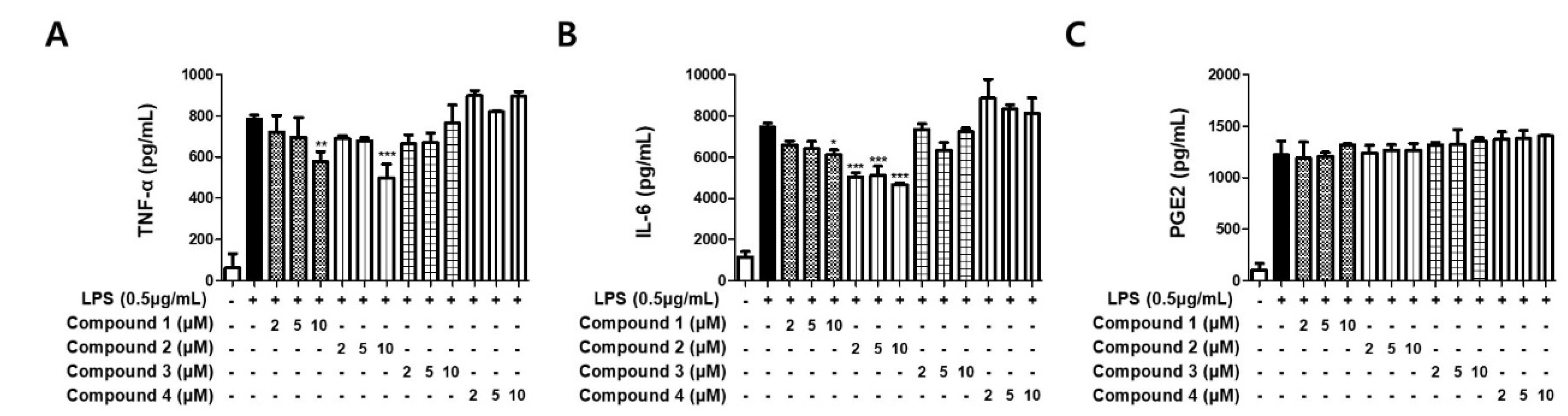
3.3. Sterigmatocystins Inhibited the LPS-induced Production of Pro-inflammatory Cytokines in BV2 Cells
3.4. Sterigmatocystins Inhibited the LPS-induced Activation of NF-κB Pathway in BV2 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Catorce, M.N.; Gevorkian, G. LPS-induced murine neuroinflammation model: Main features and suitability for pre-clinical assessment of nutraceuticals. Curr. Neuropharmacol. 2016, 14, 155–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disabato, D.; Quan, N.; Godbout, J.P. Neuroinflammation: The devil is in the details. J. Neurochem. 2016, 139, 136–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.W.; Zhang, X.; Huang, W.J. Role of neuroinflammation in neurodegerative diseases. Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streit, W.J. Microglia and neuroprotection: Implications for Alzheimer’s disease. Brain Res. Rev. 2005, 48, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Hernandez, S.; Blanco, J.; Garcia-Vallvé, S.; Pujadas, G.; Ojeda-Montes, M.J.; Gimeno, A.; Arola, L.; Minghetti, L.; Beltrán-Debón, R.; Mulero, M. Anti-inflammatory and immunomodulatory effects of the Grifola frondosa natural compound o-orsellinaldehyde on LPS-challenged murine primary glial cells. Roles of NF-κB and MAPK. Pharmaceutics 2021, 13, 806. [Google Scholar] [CrossRef]
- Kim, K.W.; Kim, H.J.; Sohn, J.H.; Yim, J.H.; Kim, Y.C.; Oh, H. Anti-neuroinflammatory effects of 6,8,1′-tri-O-methylaverantin, a metabolite from a marine-derived fungal strain Aspergillus sp., via upregulation of heme oxygenase-1 in lipopolysaccharide-activated microglia. Neurochem. Int. 2018, 113, 8–22. [Google Scholar]
- Shih, R.H.; Wang, C.Y.; Yang, C.M. NF-kappaB signaling pathways in neurological inflammation: A mini review. Front. Mol. Neurosci. 2015, 8, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Li, Y.L.; Zhao, F.C. Secondary metabolites from polar organisms. Mar. Drugs 2017, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Kim, H.J.; Cao, T.Q.; Liu, Z.; Lee, H.; Ko, W.; Kim, Y.C.; Sohn, J.H.; Kim, T.K.; Yim, J.H.; et al. Anti-inflammatory effects of metabolites from Antarctic fungal strain Pleosporales sp. SF-7343 in HaCaT human keratinocytes. Int. J. Mol. Sci. 2021, 22, 9674. [Google Scholar]
- Cong, B.; Yin, X.; Deng, A.; Shen, J.; Tian, Y.; Wang, S.; Yang, H. Diversity of cultivable microbes from soil of the Fildes Peninsula, Antarctica, and their potential application. Front. Microbiol. 2020, 11, 570836. [Google Scholar] [CrossRef]
- Ko, W.; Lee, H.; Kim, N.; Jo, H.G.; Woo, E.R.; Lee, K.; Han, Y.S.; Park, S.R.; Ahn, G.; Cheong, S.H.; et al. The anti-oxidative and anti-neuroinflammatory effects of Sargassum horneri by heme oxygenase-1 induction in BV2 and HT22 cells. Antioxidants 2021, 10, 859. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Liu, Z.; Yoon, C.S.; Dong, L.; Ko, W.; Woo, E.R.; Lee, D.S. Anti-neuroinflammatory and anti-inflammatory activities of phenylheptatriyne isolated from the flowers of Coreopsis lanceolata L. via NF-κB inhibition and HO-1 expression in BV2 and RAW264.7 cells. Int. J. Mol. Sci. 2021, 22, 7482. [Google Scholar] [PubMed]
- Shao, C.; She, Z.; Guo, Z.; Peng, H.; Cai, X.; Zhou, S.; Gu, Y.; Lin, Y. 1H and 13C NMR assignments for two anthraquinones and two xanthones from the mangrove fungus (ZSUH-36). Magn. Reson. Chem. 2007, 45, 434–438. [Google Scholar] [CrossRef]
- Hatsuda, Y.; Hamasaki, T.; Ishida, M.; Kiyama, Y. 6,8-O-Dimethyalversicolorin A, a new metabolite from Aspergillus versicolor. Agr. Biol. Chem. 1971, 35, 444. [Google Scholar]
- Jin, M.H.; Chen, D.Q.; Jin, Y.H.; Han, Y.H.; Sun, H.N.; Kwon, T. Hispidin inhibits LPS-induced nitric oxide production in BV-2 microglial cells via ROS-dependent MAPK signaling. Exp. Ther. Med. 2021, 22, 970. [Google Scholar] [CrossRef]
- Fulop, G.A.; Ahire, C.; Csipo, T.; Tarantini, S.; Kiss, T.; Balasubramanian, P.; Yabluchanskiy, A.; Farkas, E.; Toth, A.; Nyúl-Tóth, Á.; et al. Cerebral venous congestion promotes blood-brain barrier disruption and neuroinflammation, impairing cognitive function in mice. Geroscience 2019, 41, 575–589. [Google Scholar] [CrossRef]
- Wang, X.; Yu, H.; Wang, C.; Liu, Y.; Zhang, G. Chronic ethanol exposure induces neuroinflammation in h4 cells through tlr3/nf-κb pathway and anxiety-like behavior in male c57bl/6 mice. Toxicology 2020, 446, 152625. [Google Scholar] [CrossRef]
- Hung, W.; Ho, C.; Pan, M. Targeting the nlrp3 inflammasome in neuroinflammation: Health promoting effects of dietary phytochemicals in neurological disorders. Mol. Nutr. Food. Res. 2020, 64, e1900550. [Google Scholar] [CrossRef]
- Tripathi, A.; Whitehead, C.; Surrao, K.; Pillai, A.; Pillai, A. Type 1 interferon mediates chronic stress-induced neuroinflammation and behavioral deficits via complement component 3-dependent pathway. Mol. Psychiatry 2021, 26, 3043–3059. [Google Scholar] [CrossRef]
- Li, Z.; Korhonen, E.A.; Merlini, A.; Strauss, J.; Alitalo, K. Angiopoietin-2 blockade ameliorates autoimmune neuroinflammation by inhibiting leukocyte recruitment into the cns. J. Clin. Investig. 2020, 130, 1977–1990. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Wang, H.; Wang, T.; Luo, C.; Wang, Z.; Zhang, M.; Chen, X.; Tao, L. Platelet regulates neuroinflammation and restores blood–brain barrier integrity in a mouse model of traumatic brain injury. J. Neurochem. 2020, 154, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Yu, Y.Y.; Dai, J.J.; Zhang, Y.T.; Hu, W.P.; Zheng, L.; Shi, D.Y. New polyketides from the Antarctic fungus Pseudogymnoascus sp. HSX2#-11. Mar. Drugs 2021, 19, 168. [Google Scholar]
- Ha, T.M.; Kim, D.C.; Sohn, J.H.; Yim, J.H.; Oh, H. Anti-inflammatory and protein tyrosine phosphatase 1B inhibitory metabolites from the Antarctic marine-derived fungal strain Penicillium glabrum SF-7123. Mar. Drugs 2020, 18, 247. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.Q.; Tran, M.H.; Kim, J.A.; Tran, P.T.; Lee, J.H.; Woo, M.H.; Lee, H.K.; Min, B.S. Inhibitory effects of compounds from Styrax obassia on NO production. Bioorg. Med. Chem. Lett. 2015, 25, 5087–5091. [Google Scholar] [CrossRef] [PubMed]
- Hur, G.M.; Ryu, Y.S.; Yun, H.Y.; Jeon, B.H.; Kim, Y.M.; Seok, J.H.; Lee, J.H. Hepatic ischemia/reperfusion in rats induces 584 iNOS gene transcription by activation of NF-kappaB. Biochem. Biophys. Res. Commun. 1999, 261, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Colton, C.A.; Snell, J.; Chernyshev, O.; Gilbert, D.L. Induction of superoxide anion and nitric oxide production in cultured microglia. Ann. N. Y. Acad. Sci. 2002, 40, 140–155. [Google Scholar]
- Chun, K.S.; Surh, Y.J. Signal transduction pathways regulating cyclooxygenase-2 expression: Potential molecular targets for chemoprevention. Biochem. Pharmacol. 2004, 68, 1089–1100. [Google Scholar] [CrossRef]
- Kleinert, H.; Pautz, A.; Linker, K.; Schwarz, P.M. Regulation of the expression of inducible nitric oxide synthase. Eur. J. Pharmacol. 2004, 500, 255–266. [Google Scholar] [CrossRef]
- Luo, J.; Huang, B.; Zhang, Z.; Liu, M.; Luo, T. Delayed treatment of propofol inhibits lipopolysaccharide-induced inflammation in microglia through the PI3K/PKB pathway. Neuroreport 2018, 29, 839–845. [Google Scholar] [CrossRef]
- Depino, A.; Ferrari, C.; Godoy, M.C.P.; Tarelli, R.; Pitossi, F.J. Differential effects of interleukin-1β on neurotoxicity, cytokine induction and glial reaction in specific brain regions. J. Neuroimmunol. 2005, 168, 96–110. [Google Scholar] [CrossRef]
- Savarin, C.; Hinton, D.R.; Valentin-Torres, A.; Chen, Z.; Trapp, B.D.; Bergmann, C.C.; Stohlman, S.A. Astrocyte response to IFN-γ limits IL-6-mediated microglia activation and progressive autoimmune encephalomyelitis. J. Neuroinflamm. 2015, 12, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, C.L.; Chi, C.W.; Liu, T.Y. The anti-inflammatory potential of berberine in vitro and in vivo. Cancer Lett. 2004, 203, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.S.; Rai, S.N.; Birla, H.; Zahra, W.; Rathore, A.S.; Singh, S.P. NF-κB-mediated neuroinflammation in Parkinson’s disease and potential therapeutic effect of polyphenols. Neurotox. Res. 2020, 37, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Lanzillotta, A.; Porrini, V.; Bellucci, A.; Benarese, M.; Branca, C.; Parrella, E.; Spano, P.F.; Pizzi, M. NF-κB in innate neuroprotection and age-related neurodegenerative diseases. Front. Neurol. 2015, 6, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltschmidt, B.; Widera, D.; Kaltschmidt, C. Signaling via NF-κB in the nervous system. Biochim. Biophys. Acta 2005, 1745, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.M.; Li, H.; Hong, J.; Cho, H.Y.; Bae, K.S.; Kim, M.A.; Kim, D.K.; Jung, J.H. Bioactive metabolites from the sponge-derived fungus Aspergillus versicolor. Arch. Pharm. Res. 2010, 33, 231–235. [Google Scholar] [CrossRef]
- Han, X.; Tang, X.; Luo, X.; Sun, C.; Liu, K.; Zhang, Y.; Li, P.; Li, G. Isolation and identification of three new sterigmatocystin derivatives from the fungus Aspergillus versicolor guided by molecular networking approach. Chem. Biodivers. 2020, 17, e2000208. [Google Scholar] [CrossRef]
- Song, F.; Ren, B.; Chen, C.; Yu, K.; Liu, X.; Zhang, Y.; Yang, N.; He, H.; Liu, X.; Dai, H.; et al. Three new sterigmatocystin analogues from marine-derived fungus Aspergillus versicolor MF359. Appl. Microbiol. Biotechnol. 2014, 98, 3753–3758. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, T.Q.; Liu, Z.; Dong, L.; Lee, H.; Ko, W.; Vinh, L.B.; Tuan, N.Q.; Kim, Y.-C.; Sohn, J.H.; Yim, J.H.; et al. Identification of Potential Anti-Neuroinflammatory Inhibitors from Antarctic Fungal Strain Aspergillus sp. SF-7402 via Regulating the NF-κB Signaling Pathway in Microglia. Molecules 2022, 27, 2851. https://doi.org/10.3390/molecules27092851
Cao TQ, Liu Z, Dong L, Lee H, Ko W, Vinh LB, Tuan NQ, Kim Y-C, Sohn JH, Yim JH, et al. Identification of Potential Anti-Neuroinflammatory Inhibitors from Antarctic Fungal Strain Aspergillus sp. SF-7402 via Regulating the NF-κB Signaling Pathway in Microglia. Molecules. 2022; 27(9):2851. https://doi.org/10.3390/molecules27092851
Chicago/Turabian StyleCao, Thao Quyen, Zhiming Liu, Linsha Dong, Hwan Lee, Wonmin Ko, Le Ba Vinh, Nguyen Quoc Tuan, Youn-Chul Kim, Jae Hak Sohn, Joung Han Yim, and et al. 2022. "Identification of Potential Anti-Neuroinflammatory Inhibitors from Antarctic Fungal Strain Aspergillus sp. SF-7402 via Regulating the NF-κB Signaling Pathway in Microglia" Molecules 27, no. 9: 2851. https://doi.org/10.3390/molecules27092851
APA StyleCao, T. Q., Liu, Z., Dong, L., Lee, H., Ko, W., Vinh, L. B., Tuan, N. Q., Kim, Y.-C., Sohn, J. H., Yim, J. H., Lee, D.-S., & Oh, H. (2022). Identification of Potential Anti-Neuroinflammatory Inhibitors from Antarctic Fungal Strain Aspergillus sp. SF-7402 via Regulating the NF-κB Signaling Pathway in Microglia. Molecules, 27(9), 2851. https://doi.org/10.3390/molecules27092851