
Mechanistic Insights into the Ameliorating Effect of Melanogenesis of Psoralen Derivatives in B16F10 Melanoma Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
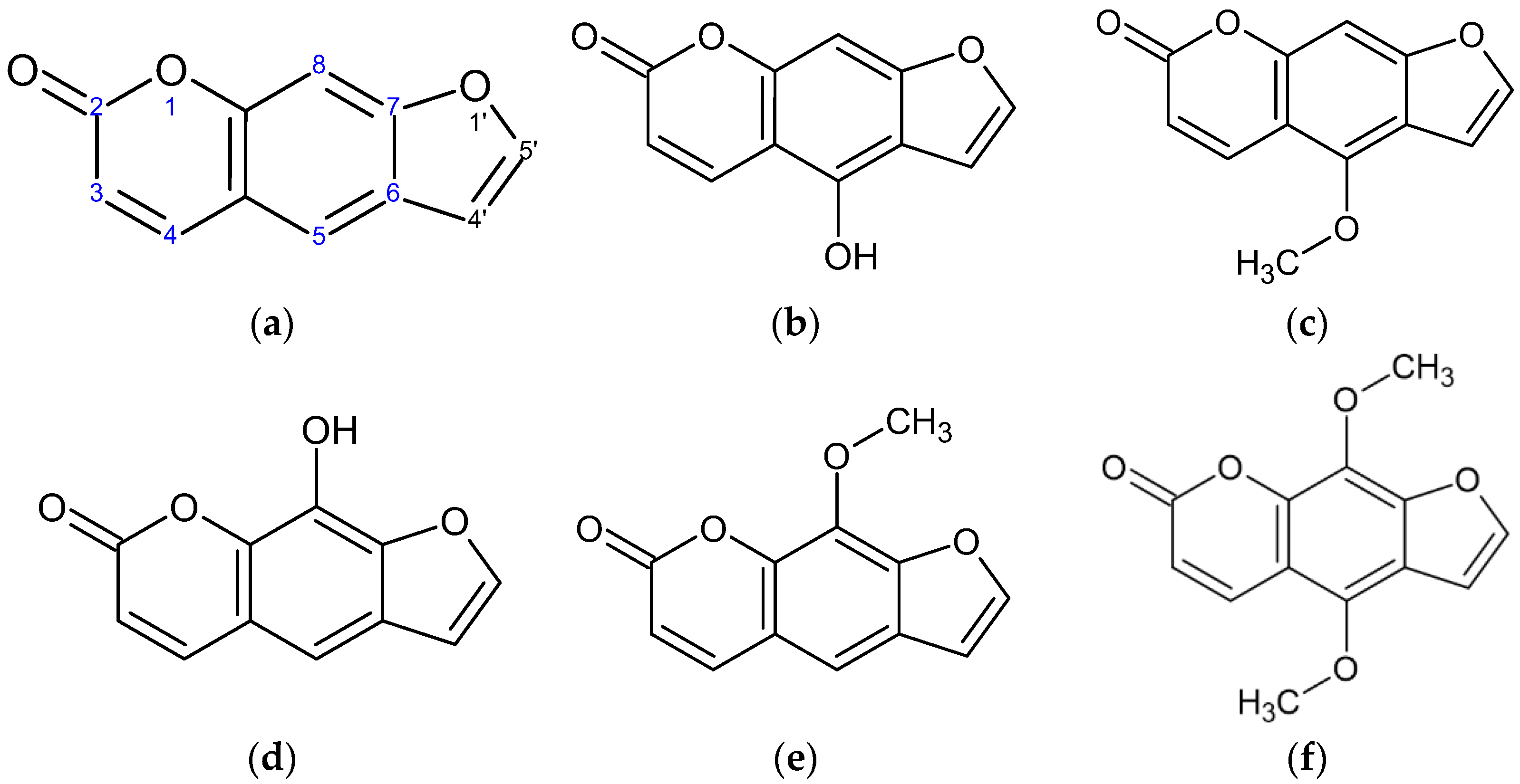
:1. Introduction
2. Results
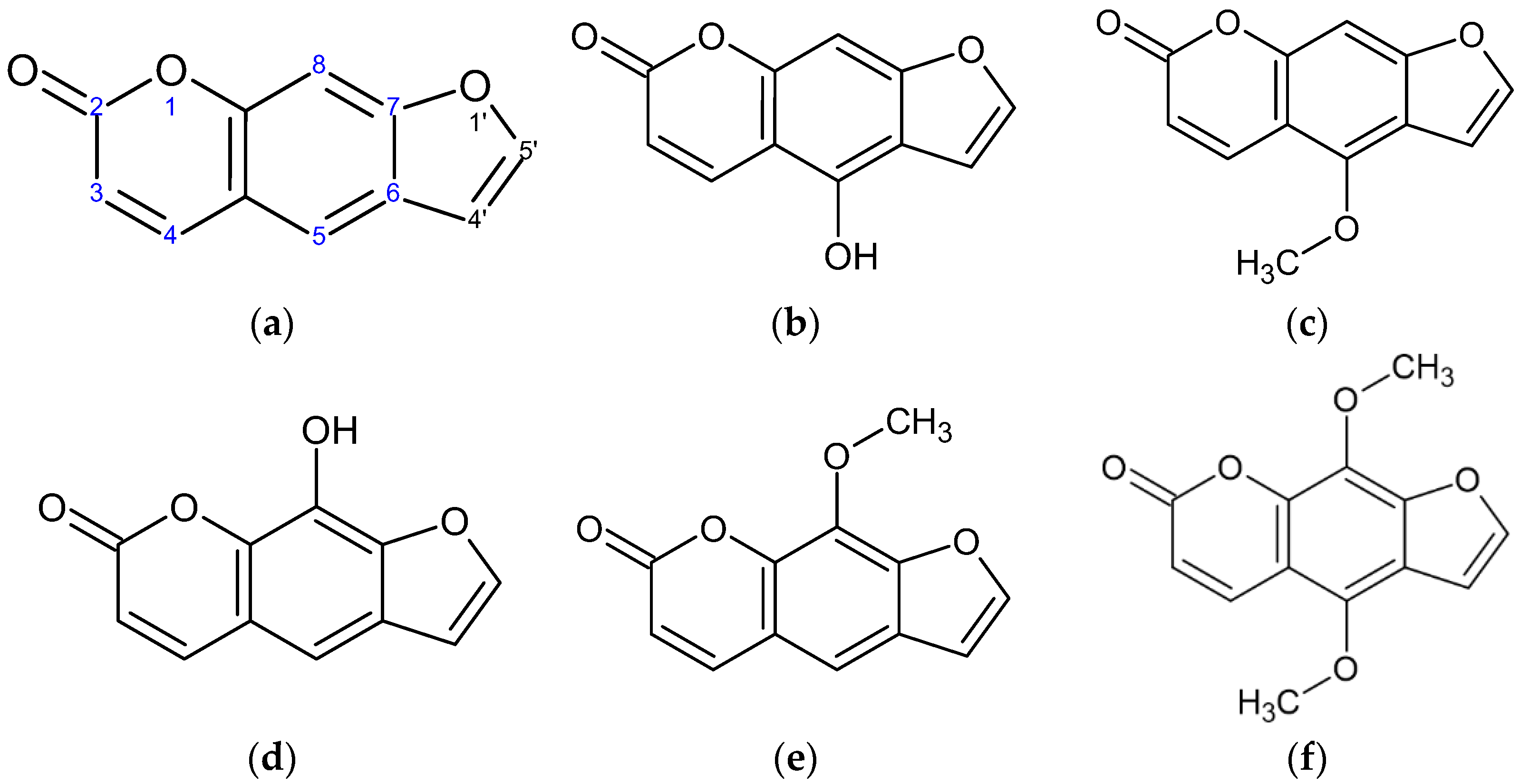
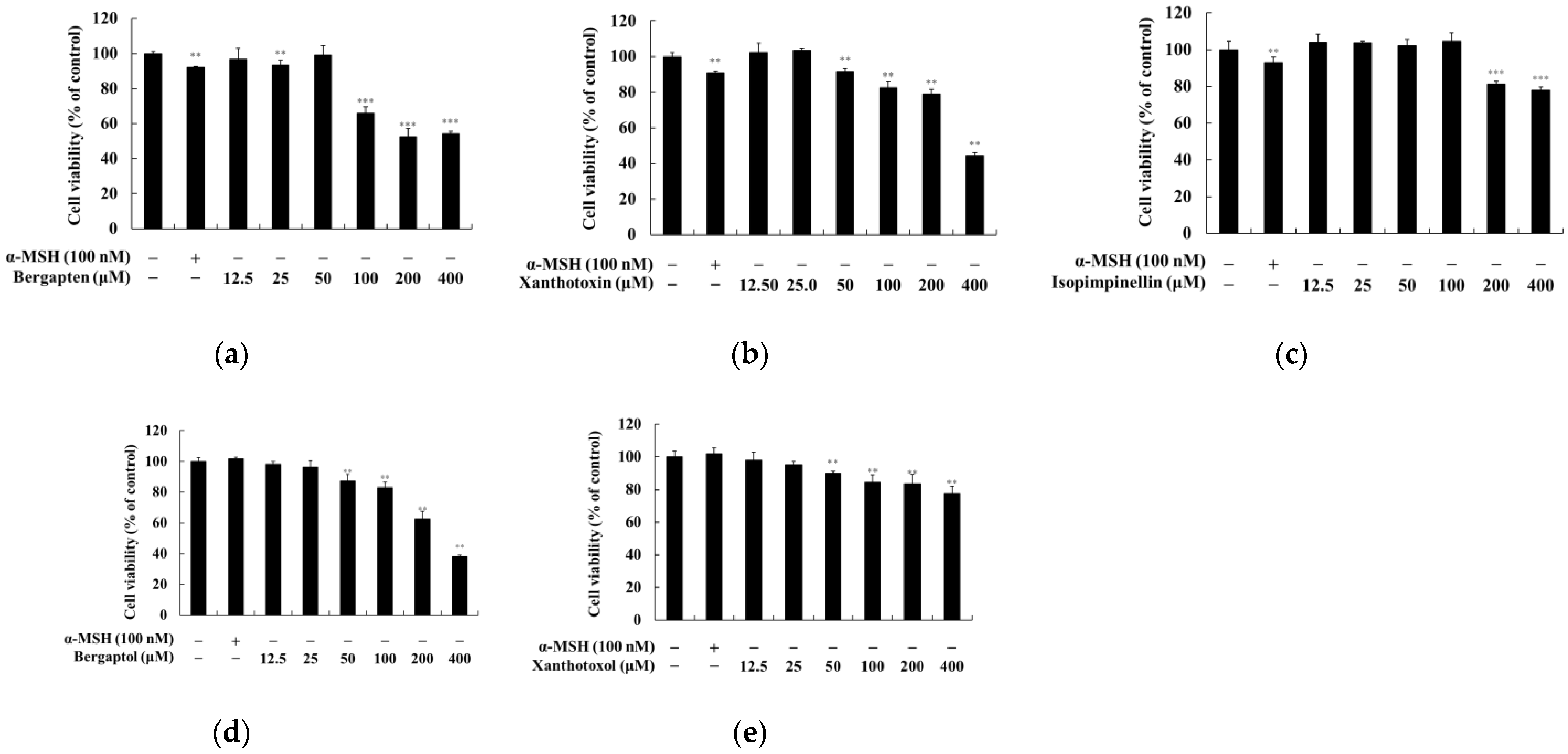
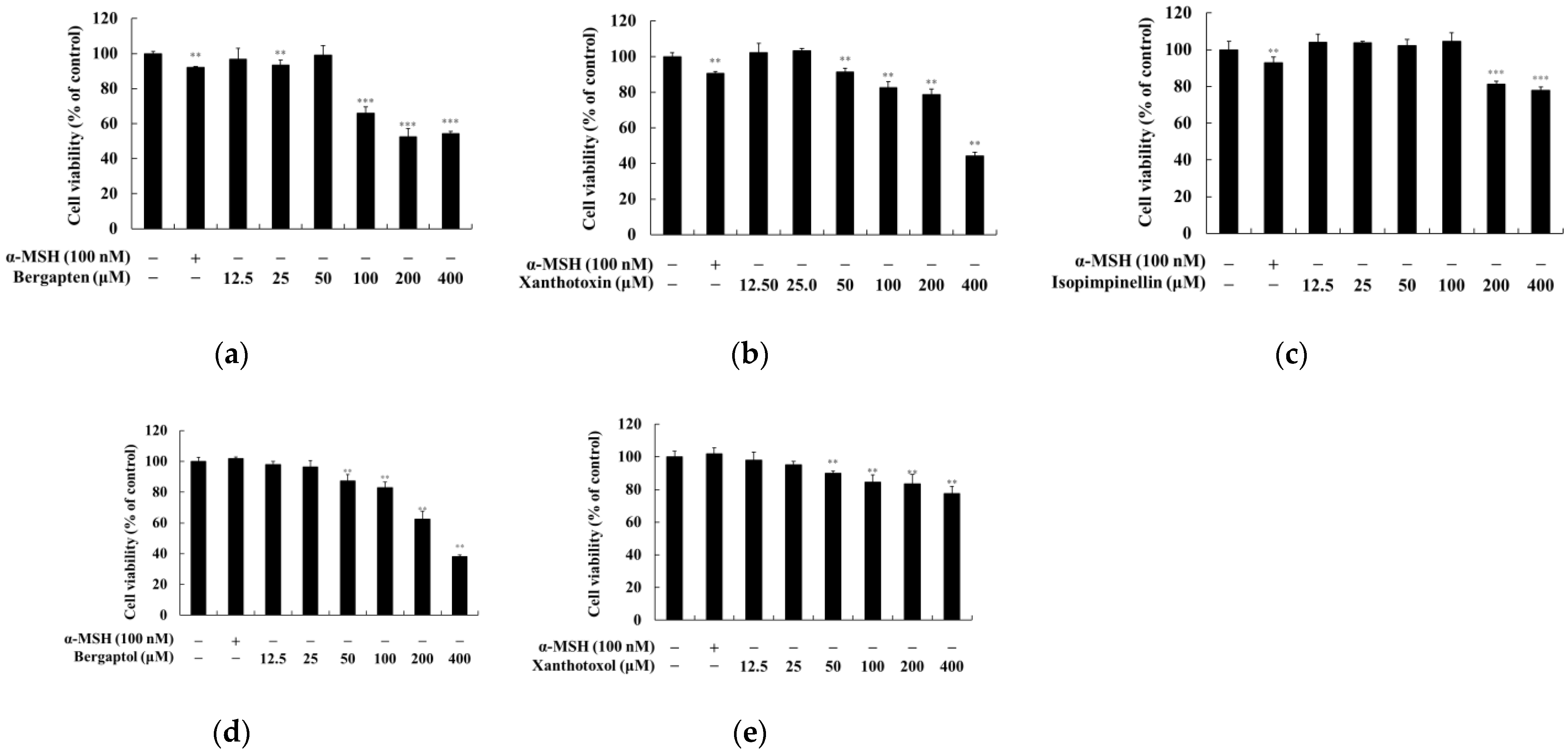
2.1. Effect of Psoralen Derivatives on the Viability of B16F10 Cells
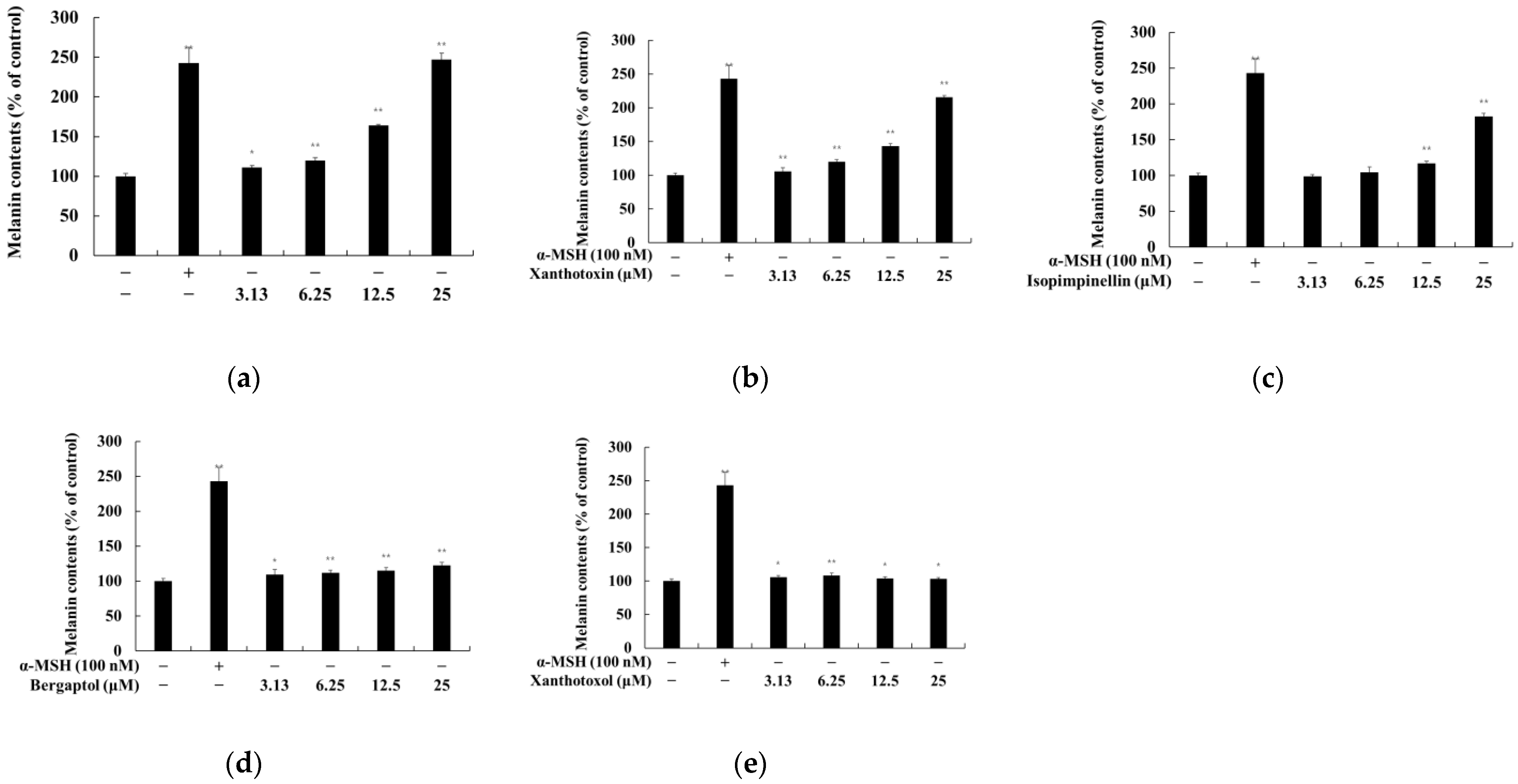
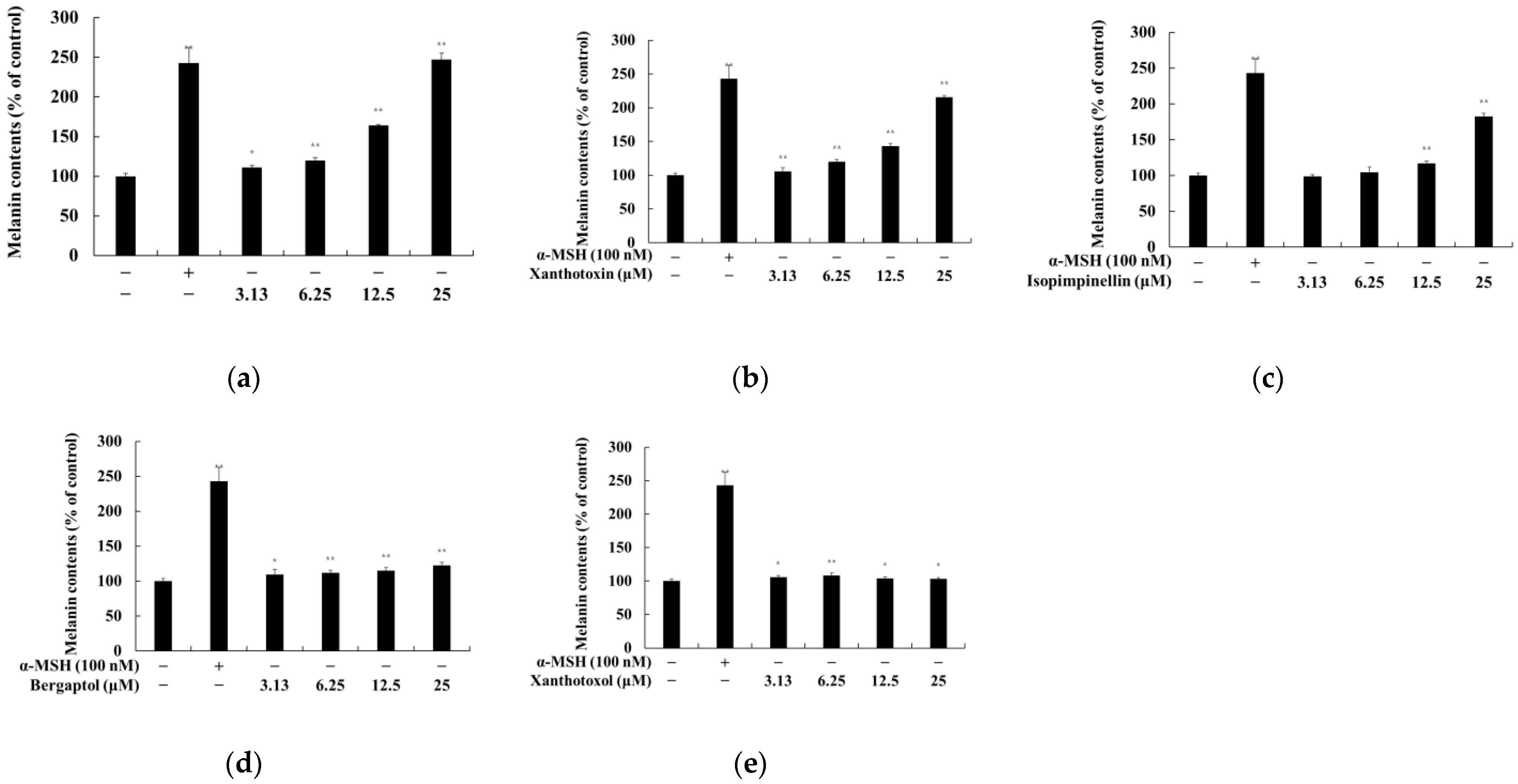
2.2. Effect of Psoralen Derivatives on the Melanin Content of B16F10 Cells
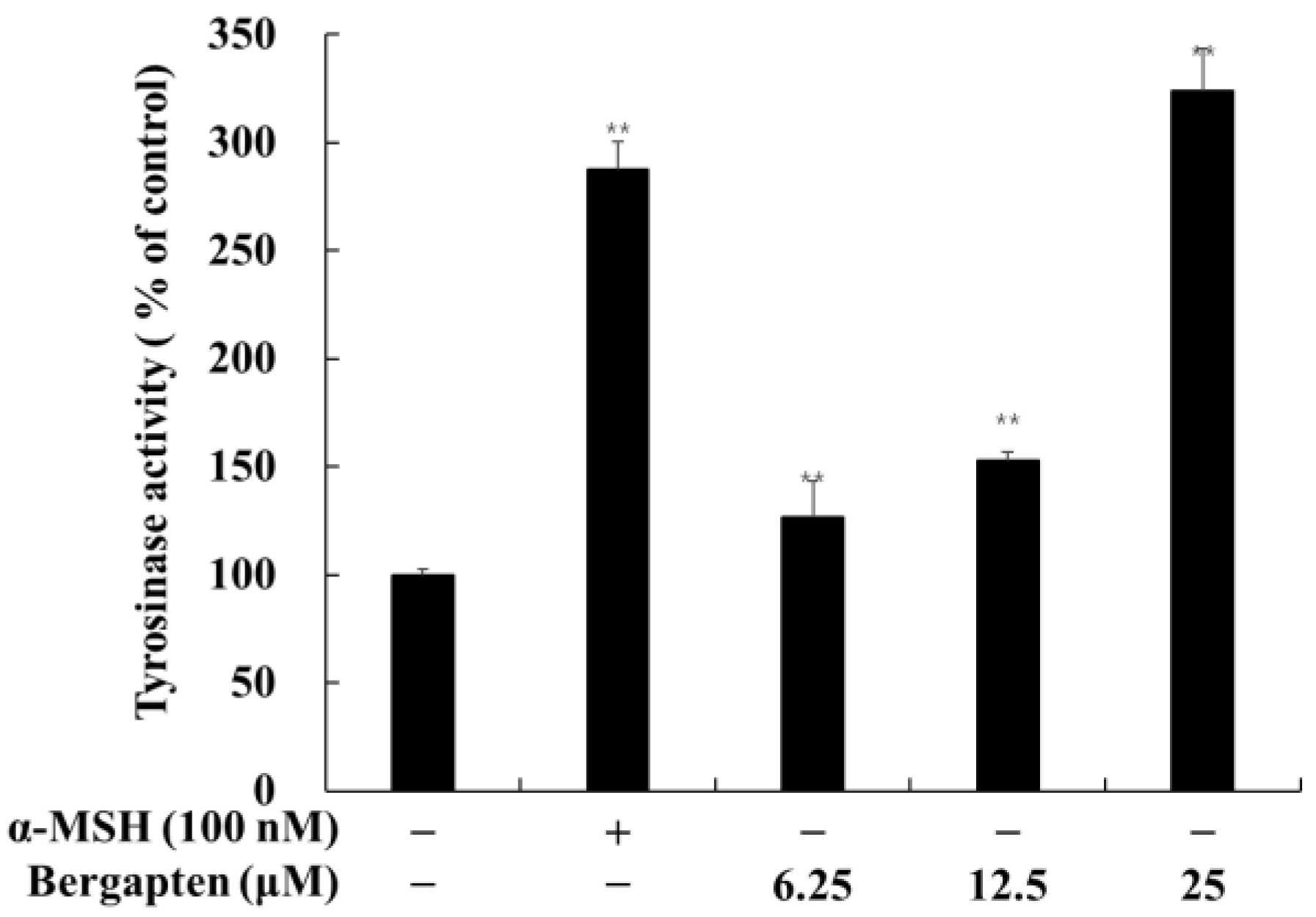
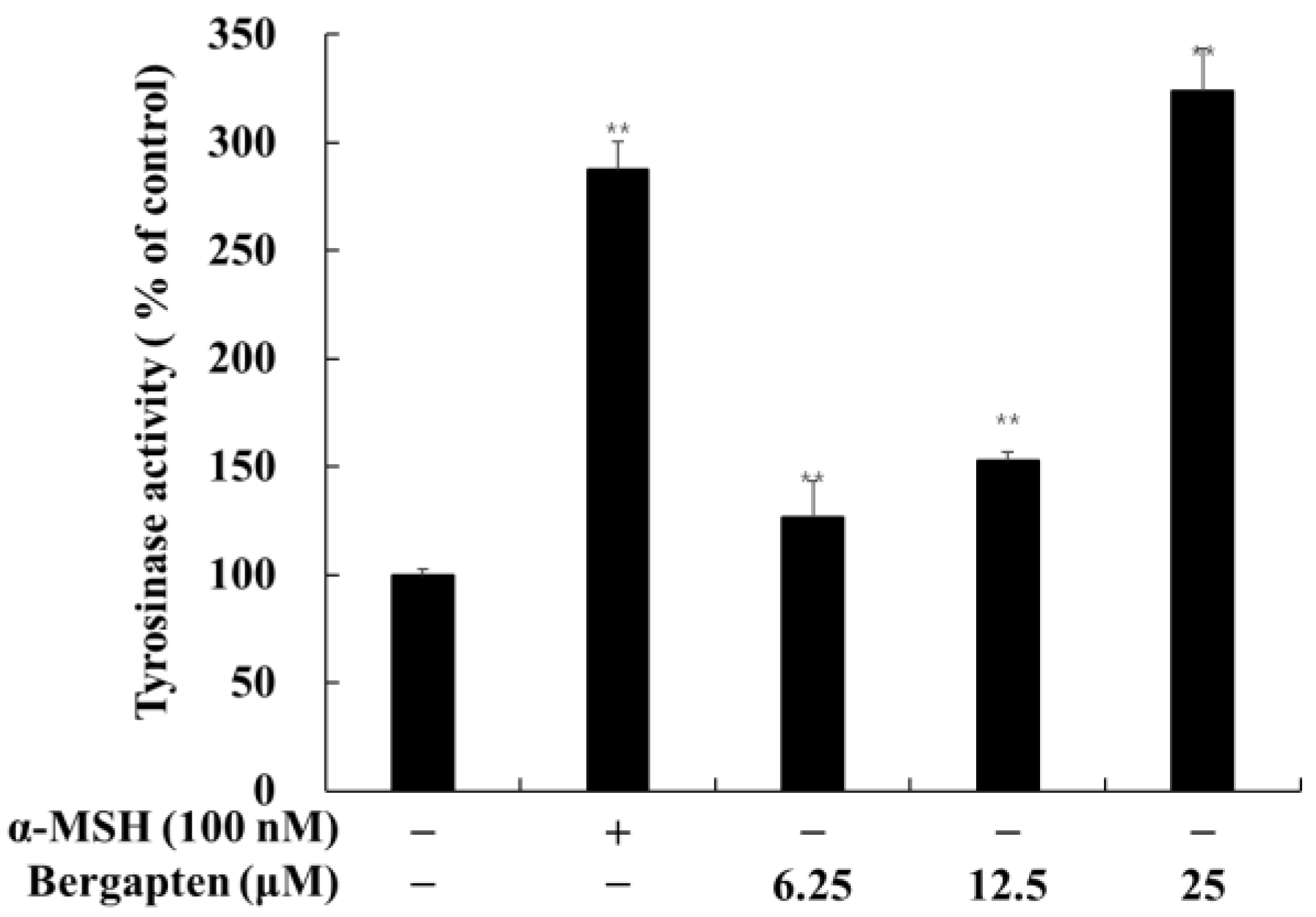
2.3. Effect of Psoralen Derivatives on Tyrosinase Activity in B16F10 Cells
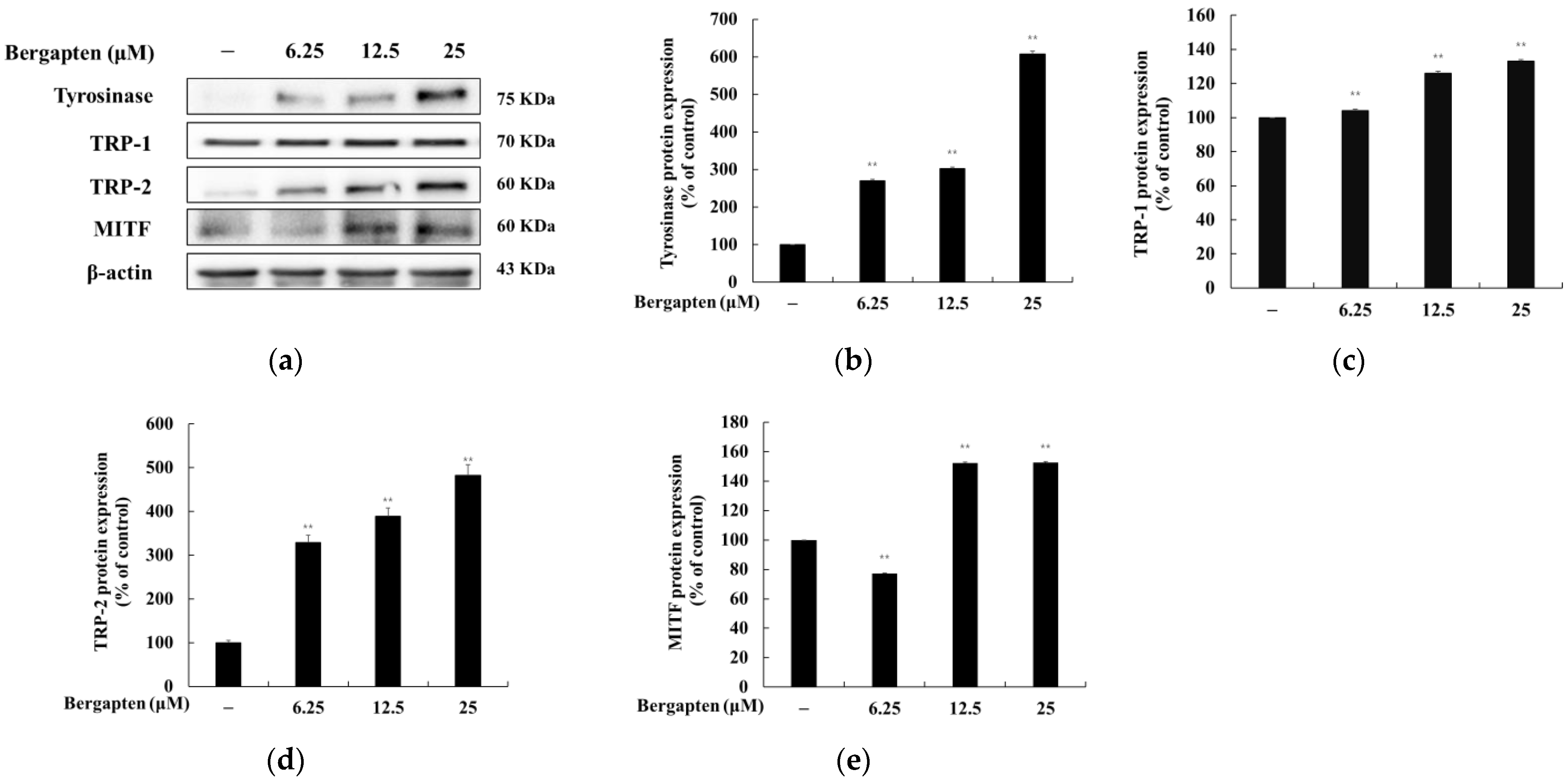
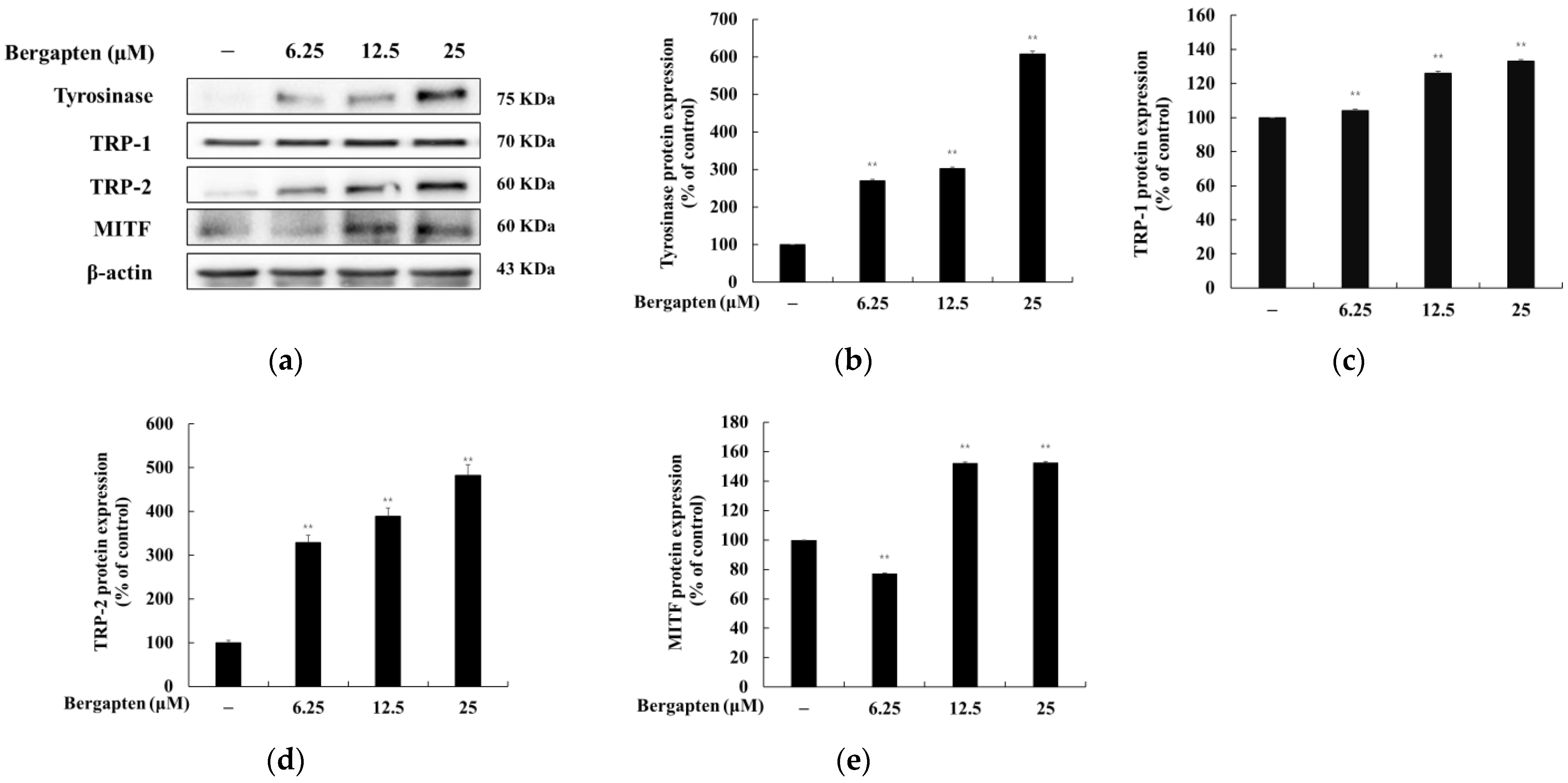
2.4. Effect of Bergapten on the Expression of Melanogenic Enzymes and MITF
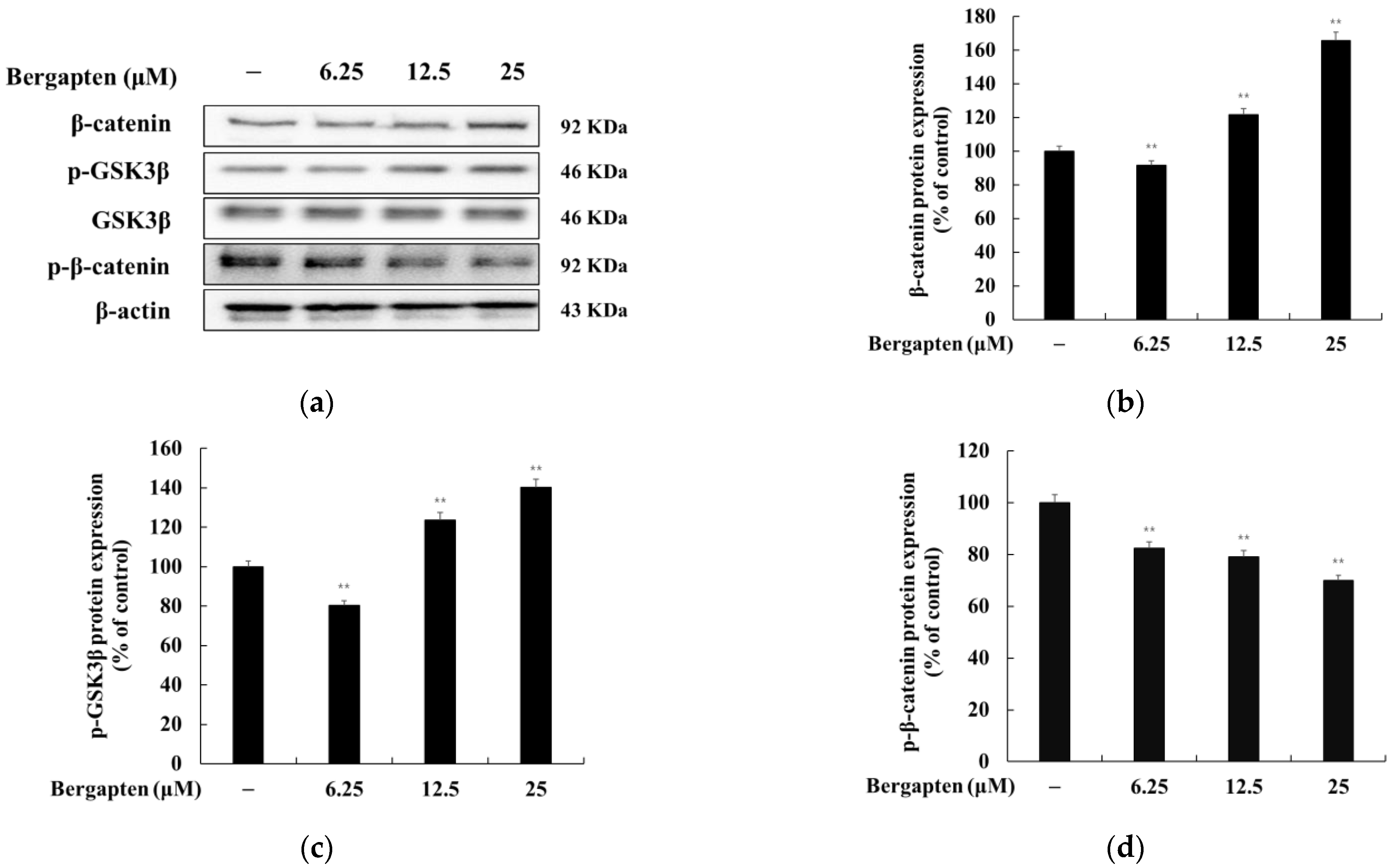
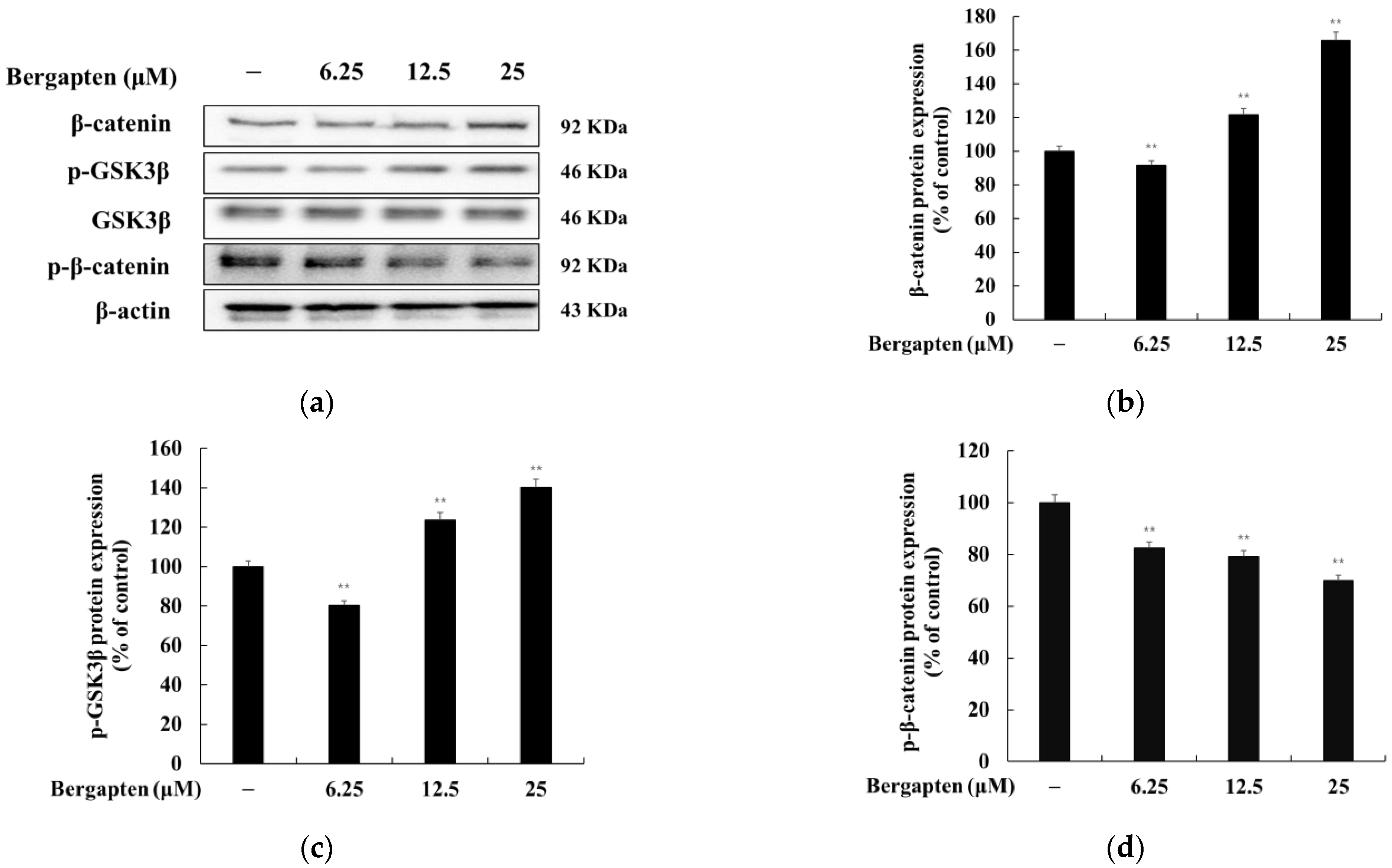
2.5. Effect of Bergapten on the Wnt/β-Catenin Signaling Pathway
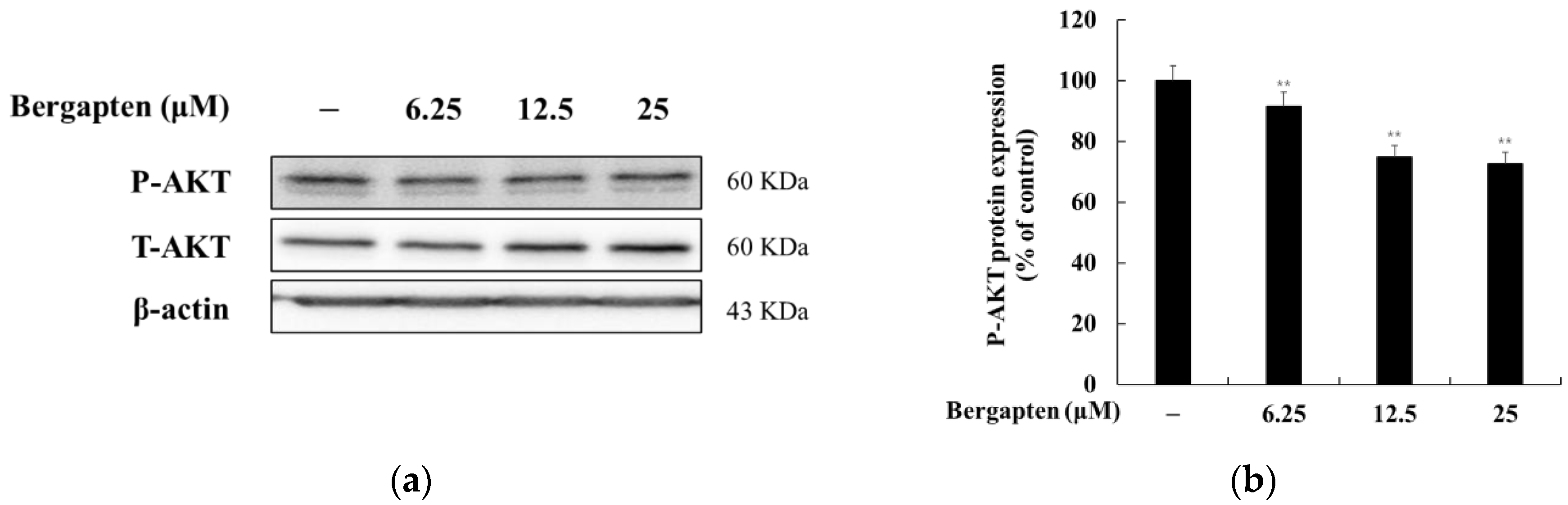
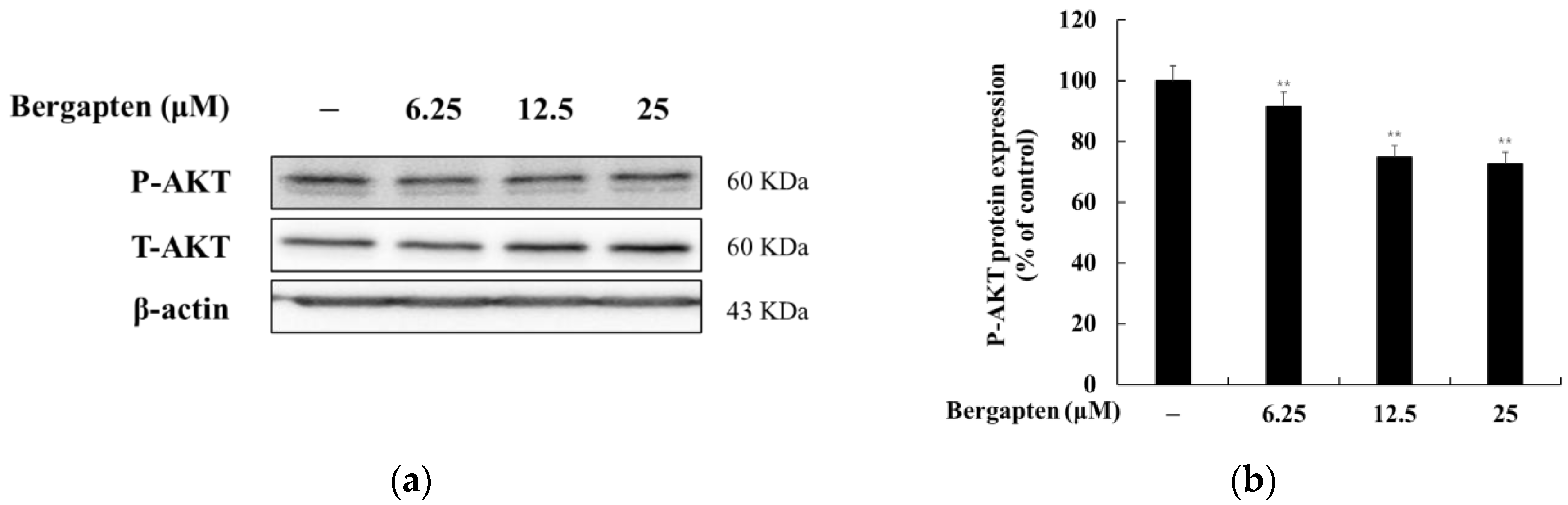
2.6. Effect of Bergapten on the AKT Signaling Pathway
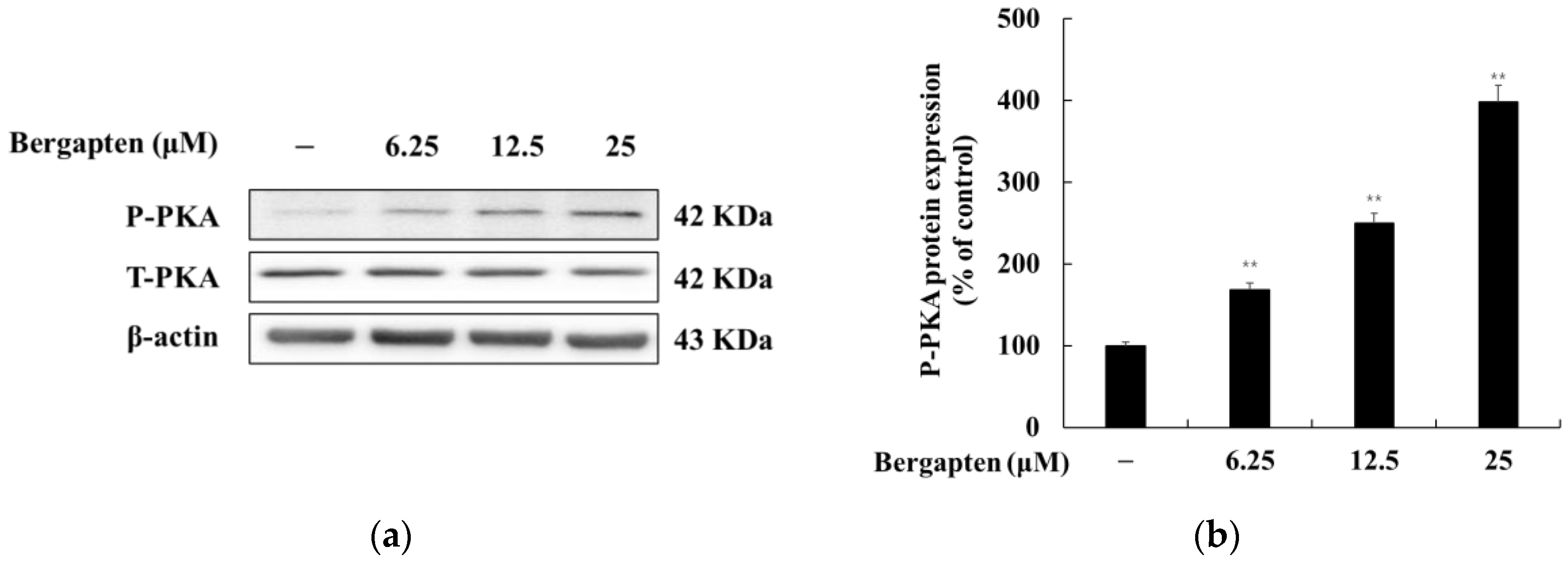
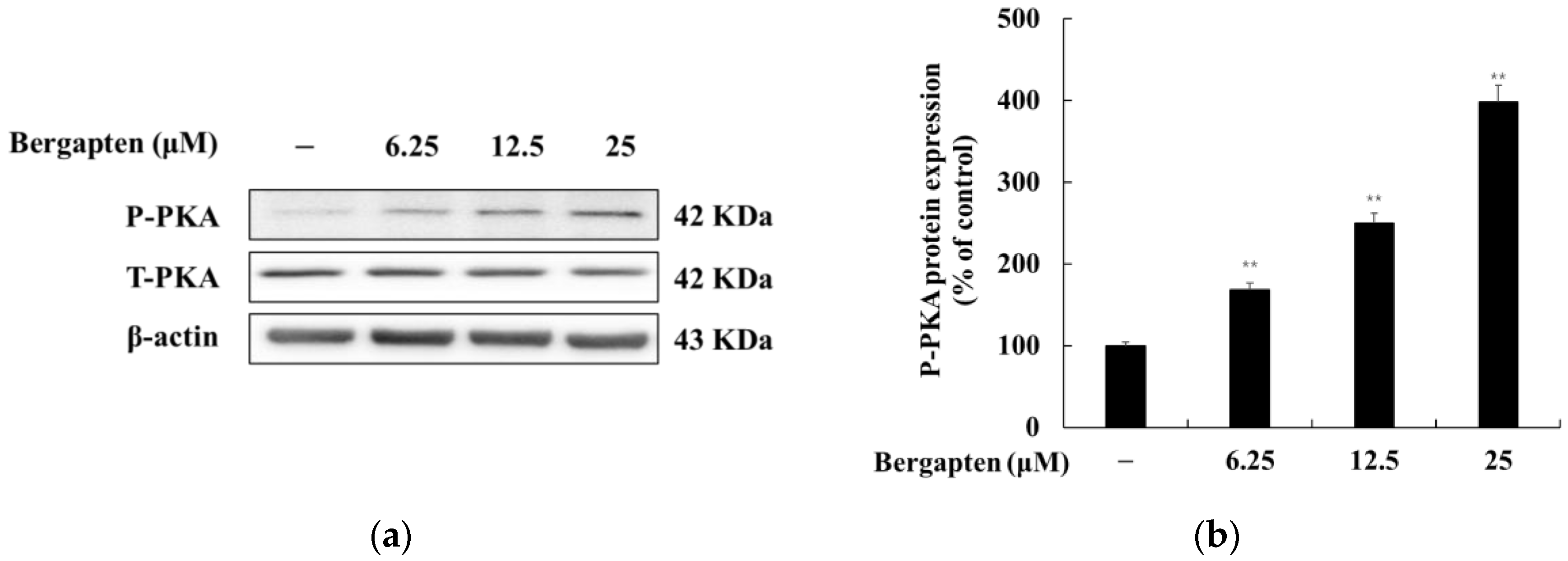
2.7. Effect of Bergapten on the PKA Signaling Pathway
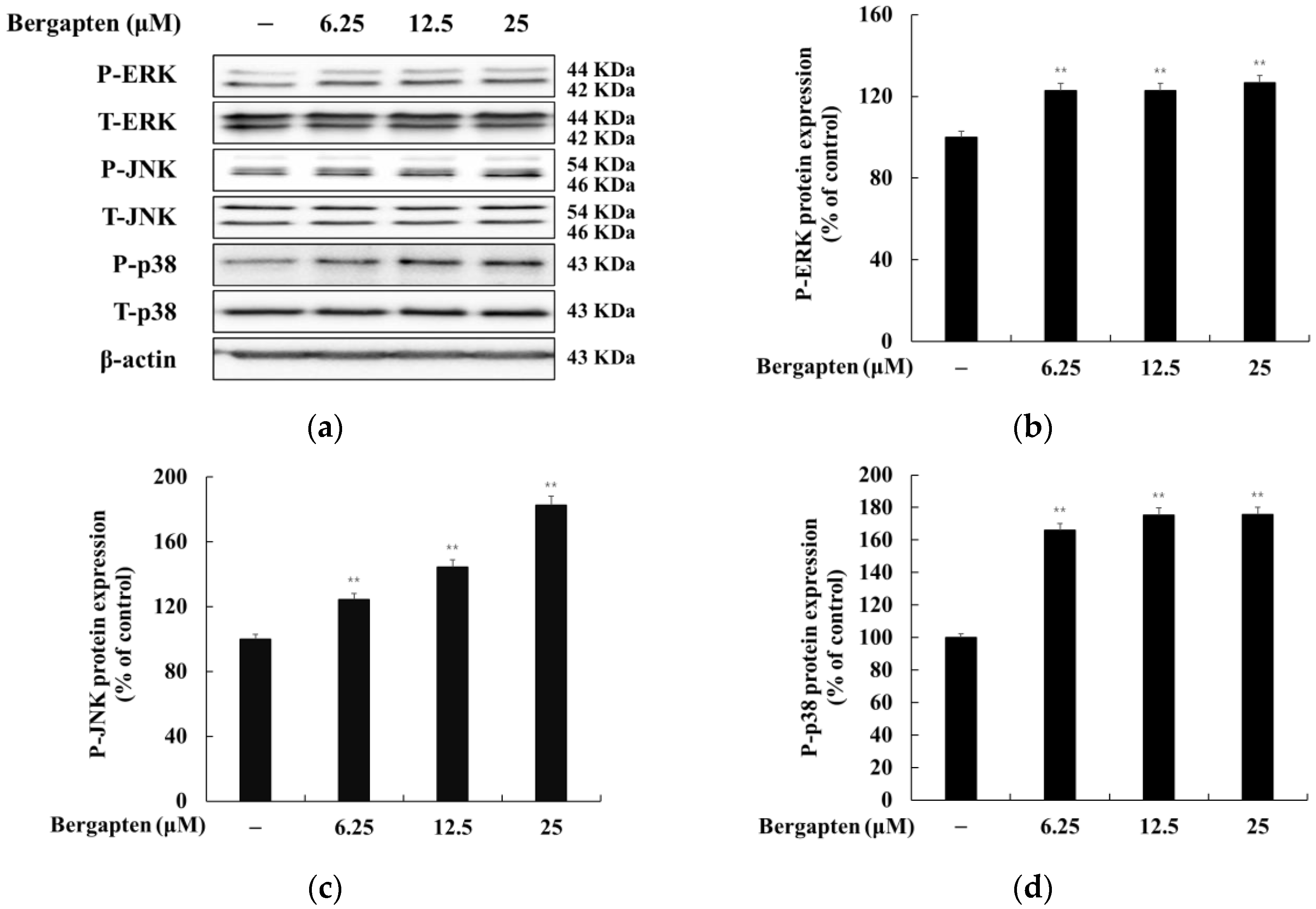
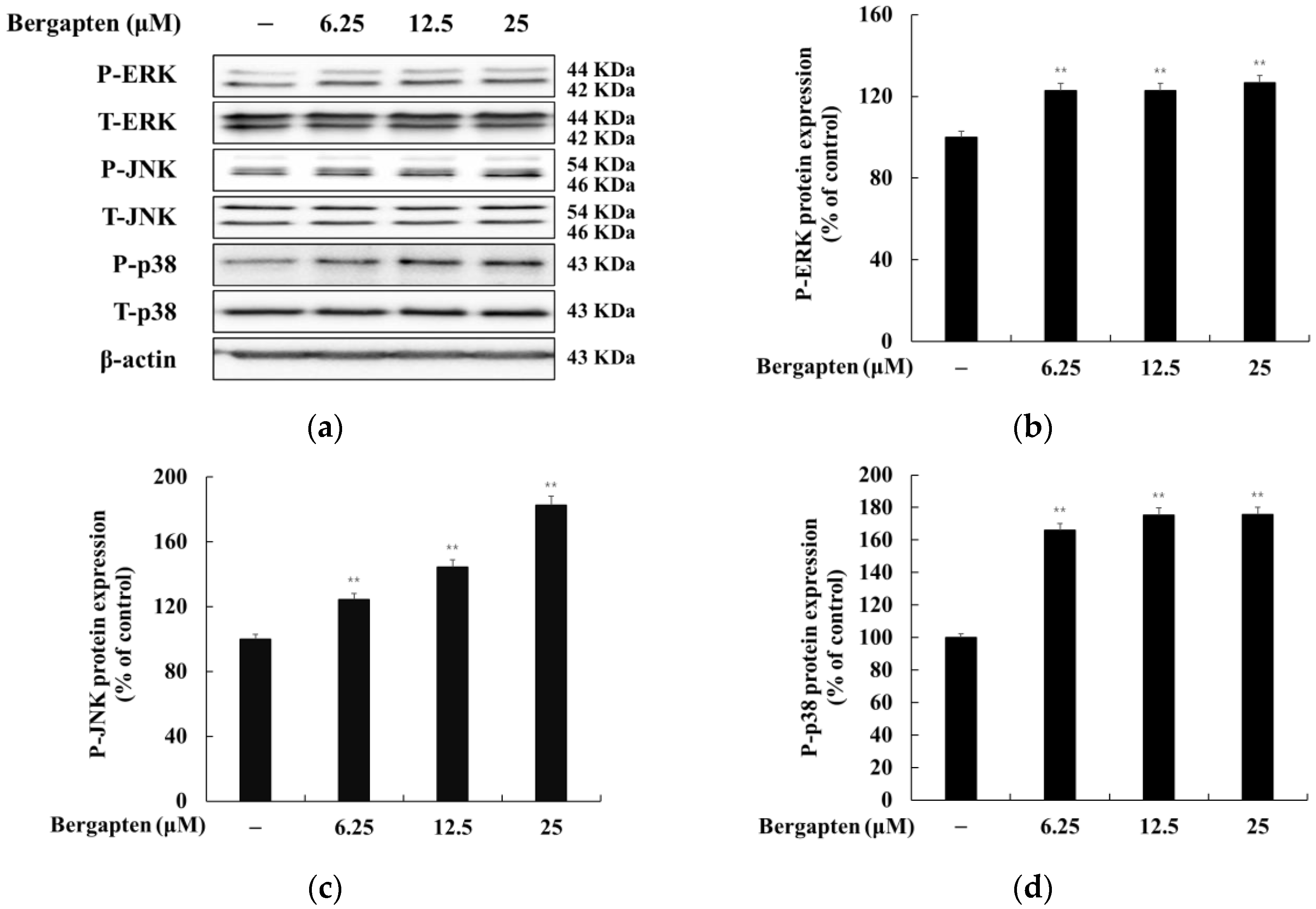
2.8. Effect of Bergapten on the MAPK Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell Viability
4.4. Melanin Content
4.5. Tyrosinase Activity
4.6. Western Blotting
4.7. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Zmijewski, M.A.; Pawelek, J. l-tyrosine and l-dihydroxyphenylalanine as hormone-like regulators of melanocyte functions. Pigment Cell Melanoma Res. 2012, 25, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.G.; Karadeniz, F.; Seo, Y.; Kong, C.S. Anti-Melanogenic Effects of Flavonoid Glycosides from Limonium tetragonum (Thunb.) Bullock via Inhibition of Tyrosinase and Tyrosinase-Related Proteins. Molecules 2017, 22, 1480. [Google Scholar] [CrossRef] [Green Version]
- D’Ischia, M.; Napolitano, A.; Ball, V.; Chen, C.T.; Buehler, M.J. Polydopamine and eumelanin: From structure-property relationships to a unified tailoring strategy. Acc. Chem. Res. 2014, 47, 3541. [Google Scholar] [CrossRef]
- Ren, Y.; Song, X.; Tan, L.; Guo, C.; Wang, M.; Liu, H.; Cao, Z.; Li, Y.; Peng, C. A Review of the Pharmacological Properties of Psoralen. Front. Pharmacol. 2020, 11, 571535. [Google Scholar] [CrossRef]
- Chen, L.; Tang, Z.S.; Song, Z.X.; Liu, Y.R.; Hu, J.H.; Shi, X.B.; Sun, C.; Jiang, D.H.; Li, X.H. Quantitative determination of nine furanocoumarins for quality evaluation of Angelica dahurica from different habitats. Zhongguo Zhong Yao Za Zhi 2019, 44, 3002. [Google Scholar] [PubMed]
- Yu, L.; Zhang, D.M. Studies on chemical constituents from fruits of Paliurus ramosissimus. Zhongguo Zhong Yao Za Zhi 2006, 31, 2049. [Google Scholar]
- Chen, I.S.; Chang, C.T.; Sheen, W.S.; Teng, C.M.; Tsai, I.L.; Duh, C.Y.; Ko, F.N. Coumarins and antiplatelet aggregation constituents from Formosan Peucedanum japonicum. Phytochemistry 1996, 41, 525. [Google Scholar] [CrossRef]
- Ramírez-Pelayo, C.; Martínez-Quiñones, J.; Gil, J.; Durango, D. Coumarins from the peel of citrus grown in Colombia: Composition, elicitation and antifungal activity. Heliyon 2019, 5, e01937. [Google Scholar] [CrossRef] [Green Version]
- Caboni, P.; Saba, M.; Oplos, C.; Aissani, N.; Maxia, A.; Menkissoglu-Spiroudi, U.; Casu, L.; Ntalli, N. Nematicidal activity of furanocoumarins from parsley against Meloidogyne spp. Pest Manag. Sci. 2015, 71, 1099. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Jin, Z.; Zheng, Y.; Xu, Y. Psoralen inhibits the inflammatory response and mucus production in allergic rhinitis by inhibiting the activator protein 1 pathway and the downstream expression of cystatin-SN. Mol. Med. Rep. 2021, 24, 652. [Google Scholar] [CrossRef] [PubMed]
- Jamalis, J.; Yusof, F.S.M.; Chander, S.; Wahab, R.A.; Bhagwat, D.P.; Sankaranarayanan, M.; Almalki, F.; Hadda, T.B. Psoralen Derivatives: Recent Advances of Synthetic Strategy and Pharmacological Properties. Antiinflamm. Antiallergy Agents Med. Chem. 2020, 19, 222. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Hyun, C.G. Inhibitory Effects of Pinostilbene on Adipogenesis in 3T3-L1 Adipocytes: A Study of Possible Mechanisms. Int. J. Mol. Sci. 2021, 22, 13446. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Hyun, C.G. Inhibitory Effects of Pinostilbene Hydrate on Melanogenesis in B16F10 Melanoma Cells via ERK and p38 Signaling Pathways. Int. J. Mol. Sci. 2020, 21, 4732. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Kim, S.; Kim, J.H.; Lee, G.S.; Lee, J.N.; Lee, N.H.; Hyun, C.G. Pratol, an O-Methylated Flavone, Induces Melanogenesis in B16F10 Melanoma Cells via p-p38 and p-JNK Upregulation. Molecules 2017, 22, 1704. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, S.S.; Park, K.J.; An, H.J.; Choi, Y.H.; Lee, N.H.; Hyun, C.G. Methyl jasmonate inhibits lipopolysaccharide-induced inflammatory cytokine production via mitogen-activated protein kinase and nuclear factor-κB pathways in RAW 264.7 cells. Pharmazie 2016, 71, 540. [Google Scholar] [PubMed]
- Yoon, W.J.; Ham, Y.M.; Yoon, H.S.; Lee, W.J.; Lee, N.H.; Hyun, C.G. Acanthoic acid inhibits melanogenesis through tyrosinase downregulation and melanogenic gene expression in B16 melanoma cells. Nat. Prod. Commun. 2013, 8, 1359. [Google Scholar] [PubMed] [Green Version]
- Martins, F.S.; Sy, S.K.B.; Fonseca, M.J.V.; de Freitas, O. Pharmacokinetics, Pharmacodynamics and Dermal Distribution of 5-Methoxypsoralen Based on a Physiologically Based Pharmacokinetic Model to Support Phytotherapy Using Brosimum gaudichaudii. Planta Med. 2020, 86, 276. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Hirata, N.; Kawaguchi, Y.; Yamazaki, M.; Naruto, S.; Shibano, M.; Taniguchi, M.; Baba, K.; Kubo, M. Melanogenesis stimulation in murine b16 melanoma cells by umberiferae plant extracts and their coumarin constituents. Biol. Pharm. Bull. 2005, 28, 1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengeaud, V.; Ortonne, J.P. PUVA (5-methoxypsoralen plus UVA) enhances melanogenesis and modulates expression of melanogenic proteins in cultured melanocytes. J. Investig. Dermatol. 1996, 107, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengeaud, V.; Ortonne, J.P. Regulation of melanogenesis induced by 5-methoxypsoralen without ultraviolet light in murine melanoma cells. Pigment Cell Res. 1994, 7, 245. [Google Scholar] [CrossRef] [PubMed]
- Kinley, J.S.; Moan, J.; Dall’Aqua, F.; Young, A.R. Quantitative assessment of epidermal melanogenesis in C3H/Tif hr/hr mice treated with topical furocoumarins and UVA radiation. J. Investig. Dermatol. 1994, 103, 97. [Google Scholar] [CrossRef] [Green Version]
- Kinley, J.S.; Brunborg, G.; Moan, J.; Young, A.R. Photoprotection by furocoumarin-induced melanogenesis against DNA photodamage in mouse epidermis in vivo. Photochem. Photobiol. 1997, 65, 486. [Google Scholar] [CrossRef]
- Yin, L.; Pang, G.; Niu, C.; Habasi, M.; Dou, J.; Aisa, H.A. A novel psoralen derivative-MPFC enhances melanogenesis via activation of p38 MAPK and PKA signaling pathways in B16 cells. Int. J. Mol. Med. 2018, 41, 3727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, A.; Montalbano, A.; Spanò, V.; Musante, I.; Galietta, L.J.V.; Barraja, P. Furocoumarins as multi-target agents in the treatment of cystic fibrosis. Eur. J. Med. Chem. 2019, 180, 283. [Google Scholar] [CrossRef] [PubMed]
- Dichiara, M.; Prezzavento, O.; Marrazzo, A.; Pittalà, V.; Salerno, L.; Rescifina, A.; Amata, E. Recent advances in drug discovery of phototherapeutic non-porphyrinic anticancer agents. Eur. J. Med. Chem. 2017, 142, 459. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.M.; Raposo, M.M.; Oliveira-Campos, A.M.; Machado, A.E.; Puapairoj, P.; Pedro, M.; Nascimento, M.S.; Portela, C.; Afonso, C.; Pinto, M. Psoralen analogues: Synthesis, inhibitory activity of growth of human tumor cell lines and computational studies. Eur. J. Med. Chem. 2006, 41, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.K.; Chung, Y.C.; Hyun, C.G. Anti-Inflammatory Effects of 6-Methylcoumarin in LPS-Stimulated RAW 264.7 Macrophages via Regulation of MAPK and NF-κB Signaling Pathways. Molecules 2021, 26, 5351. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.K.; Hyun, C.G. 4-Hydroxy-7-Methoxycoumarin Inhibits Inflammation in LPS-activated RAW264.7 Macrophages by Suppressing NF-κB and MAPK Activation. Molecules 2020, 25, 4424. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Chung, Y.C.; Kim, Y.B.; Park, S.M.; Kim, B.S.; Hyun, C.G. 7,8-Dimethoxycoumarin stimulates melanogenesis via MAPKs mediated MITF upregulation. Pharmazie 2020, 75, 107. [Google Scholar]
- Chung, Y.C.; Kim, S.Y.; Hyun, C.G. 8-Methoxycoumarin enhances melanogenesis via the MAPKase signaling pathway. Pharmazie 2019, 74, 529. [Google Scholar]
- Mamat, N.; Dou, J.; Lu, X.; Eblimit, A.; Haji Akber, A. Isochlorogenic acid A promotes melanin synthesis in B16 cell through the β-catenin signal pathway. Acta Biochim. Biophys. Sin. 2017, 49, 800. [Google Scholar] [CrossRef] [Green Version]
- Niu, C.; Yin, L.; Aisa, H.A. Novel Furocoumarin Derivatives Stimulate Melanogenesis in B16 Melanoma Cells by Up-Regulation of MITF and TYR Family via Akt/GSK3β/β-Catenin Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 746. [Google Scholar] [CrossRef] [Green Version]
- Zang, D.; Niu, C.; Aisa, H.A. Amine derivatives of furocoumarin induce melanogenesis by activating Akt/GSK-3β/β-catenin signal pathway. Drug Des. Dev. Ther. 2019, 13, 623. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Kim, S.Y.; Moon, S.J.; Chung, J.H.; Kim, K.H.; Cho, K.H.; Park, K.C. Ceramide inhibits cell proliferation through Akt/PKB inactivation and decreases melanin synthesis in Mel-Ab cells. Pigment Cell Res. 2001, 14, 110. [Google Scholar] [CrossRef]
- Chae, J.K.; Subedi, L.; Jeong, M.; Park, Y.U.; Kim, C.Y.; Kim, H.; Kim, S.Y. Gomisin N Inhibits Melanogenesis through Regulating the PI3K/Akt and MAPK/ERK Signaling Pathways in Melanocytes. Int. J. Mol. Sci. 2017, 18, 471. [Google Scholar] [CrossRef]
- Im, D.S.; Lee, J.M.; Lee, J.; Shin, H.J.; No, K.T.; Park, S.H.; Kim, K. Inhibition of collagenase and melanogenesis by ethanol extracts of Orostachys japonicus A. Berger: Possible involvement of Erk and Akt signaling pathways in melanoma cells. Acta Biochim. Biophys. Sin. 2017, 49, 945. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.H.; Jo, H.G.; Yang, J.H.; Ki, S.H.; Shin, H.J. Antioxidative and Anti-Melanogenic Activities of Bamboo Stems (Phyllostachys nigra variety henosis) via PKA/CREB-Mediated MITF Downregulation in B16F10 Melanoma Cells. Int. J. Mol. Sci. 2018, 19, 409. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.Y.; Hyun, C.G. Chrysoeriol Enhances Melanogenesis in B16F10 Cells Through the Modulation of the MAPK, AKT, PKA, and Wnt/β-Catenin Signaling Pathways. Nat. Prod. Commun. 2022, 17, 1. [Google Scholar] [CrossRef]
- Ko, J.H.; Lee, N.; Kang, H.K.; Kim, B.S.; Lee, J.N.; Hyun, C.G. Anti-melanogenic effects of hot-water extracts from Torreya nucifera via MAPKs and cAMP signaling pathway on B16F10 cells. Pharmazie 2020, 75, 565. [Google Scholar]
- Ullah, S.; Chung, Y.C.; Hyun, C.G. Induction of Melanogenesis by Fosfomycin in B16F10 Cells Through the Upregulation of P-JNK and P-p38 Signaling Pathways. Antibiotics 2020, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.H.; Chung, Y.C.; Hyun, C.G. Tobramycin Promotes Melanogenesis by Upregulating p38 MAPK Protein Phosphorylation in B16F10 Melanoma Cells. Antibiotics 2019, 8, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.C.; Ko, J.H.; Kang, H.K.; Kim, S.; Kang, C.I.; Lee, J.N.; Park, S.M.; Hyun, C.G. Antimelanogenic Effects of Polygonum tinctorium Flower Extract from Traditional Jeju Fermentation via Upregulation of Extracellular Signal-Regulated Kinase and Protein Kinase B Activation. Int. J. Mol. Sci. 2018, 19, 2895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molagoda, I.M.N.; Karunarathne, W.A.H.M.; Park, S.R.; Choi, Y.H.; Park, E.K.; Jin, C.Y.; Yu, H.; Jo, W.S.; Lee, K.T.; Kim, G.Y. GSK-3β-Targeting Fisetin Promotes Melanogenesis in B16F10 Melanoma Cells and Zebrafish Larvae through β-Catenin Activation. Int. J. Mol. Sci. 2020, 21, 312. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Chang, J.L.; Chen, P.R.; Chuang, Y.J.; Tang, S.T.; Pan, S.F.; Lin, T.B.; Chen, K.H.; Chen, M.J. Inhibition of peroxisome proliferator-activated receptor gamma prevents the melanogenesis in murine B16/F10 melanoma cells. Biomed. Res. Int. 2014, 2014, 695797. [Google Scholar] [CrossRef]
- Nam, G.; An, S.K.; Park, I.C.; Bae, S.; Lee, J.H. Daphnetin inhibits α-MSH-induced melanogenesis via PKA and ERK signaling pathways in B16F10 melanoma cells. Biosci. Biotechnol. Biochem. 2022, 86, zbac016. [Google Scholar] [CrossRef]
- Słominski, A.; Moellmann, G.; Kuklinska, E.; Bomirski, A.; Pawelek, J. Positive regulation of melanin pigmentation by two key substrates of the melanogenic pathway, l-tyrosine and l-dopa. J. Cell Sci. 1988, 89 Pt 3, 287. [Google Scholar] [CrossRef]
- Slominski, A.; Moellmann, G.; Kuklinska, E. l-tyrosine, l-dopa, and tyrosinase as positive regulators of the subcellular apparatus of melanogenesis in Bomirski Ab amelanotic melanoma cells. Pigment Cell Res. 1989, 2, 109. [Google Scholar] [CrossRef]
- Diawpanich, P.; Klongpityapong, P.; Veeranondha, S.; Watts, P. DMEM enhances tyrosinase activity in B16 mouse melanoma cells and human melanocytes. Songklanakarin J. Sci. Technol. 2008, 30, 603. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Hyun, C.-G. Mechanistic Insights into the Ameliorating Effect of Melanogenesis of Psoralen Derivatives in B16F10 Melanoma Cells. Molecules 2022, 27, 2613. https://doi.org/10.3390/molecules27092613
Lee Y, Hyun C-G. Mechanistic Insights into the Ameliorating Effect of Melanogenesis of Psoralen Derivatives in B16F10 Melanoma Cells. Molecules. 2022; 27(9):2613. https://doi.org/10.3390/molecules27092613
Chicago/Turabian StyleLee, Yeji, and Chang-Gu Hyun. 2022. "Mechanistic Insights into the Ameliorating Effect of Melanogenesis of Psoralen Derivatives in B16F10 Melanoma Cells" Molecules 27, no. 9: 2613. https://doi.org/10.3390/molecules27092613
APA StyleLee, Y., & Hyun, C.-G. (2022). Mechanistic Insights into the Ameliorating Effect of Melanogenesis of Psoralen Derivatives in B16F10 Melanoma Cells. Molecules, 27(9), 2613. https://doi.org/10.3390/molecules27092613