
The Effect of Beta Adrenoreceptor Blockers on Viability and Cell Colony Formation of Non-Small Cell Lung Cancer Cell Lines A549 and H1299



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
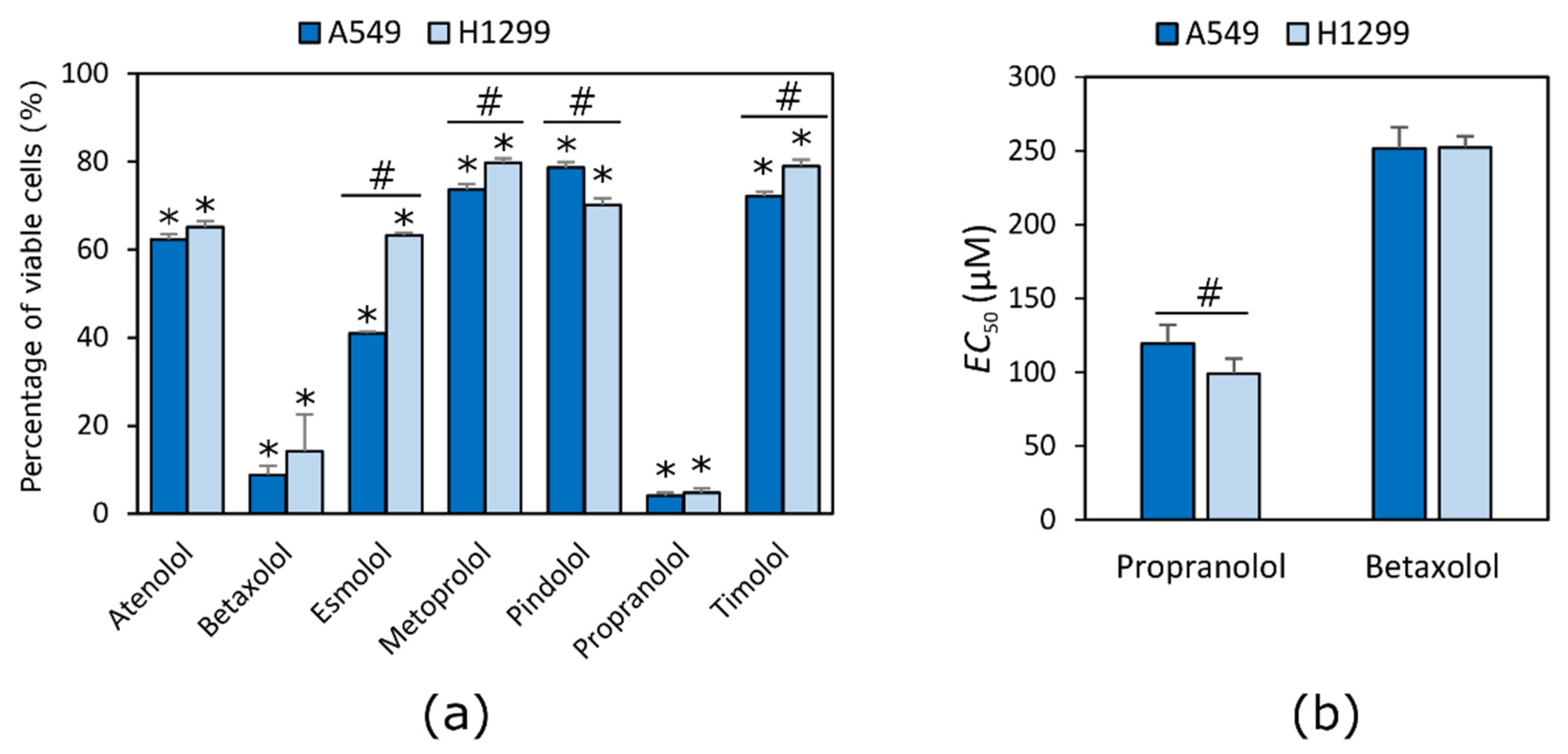
2. Results
2.1. Beta Adrenoblockers Reduce the Viability of NSCLC Cells
2.2. Beta Adrenoblockers Inhibit Growth of Cell Colonies in Concentration-Dependent Way
2.3. Beta Adrenoblockers Mainly Cause Apoptosis
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Cell Colony Formation Assay
4.5. Evaluation of Type of Cell Death
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer Drug Resistance: An Evolving Paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef]
- Siegel, R.; Ma, J.; Zou, Z.; Jemal, A. Cancer Statistics, 2014. CA Cancer J. Clin. 2014, 64, 9–29. [Google Scholar] [CrossRef] [Green Version]
- Hanns, P.; Paczulla, A.M.; Medinger, M.; Konantz, M.; Lengerke, C. Stress and Catecholamines Modulate the Bone Marrow Microenvironment to Promote Tumorigenesis. Cell Stress 2019, 3, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Schuller, H.M.; Cole, B. Regulation of Cell Proliferation by Beta-Adrenergic Receptors in a Human Lung Adenocarcinoma Cell Line. Carcinogenesis 1989, 10, 1753–1755. [Google Scholar] [CrossRef] [PubMed]
- Park, P.G.; Merryman, J.; Orloff, M.; Schuller, H.M. Beta-Adrenergic Mitogenic Signal Transduction in Peripheral Lung Adenocarcinoma: Implications for Individuals with Preexisting Chronic Lung Disease. Cancer Res. 1995, 55, 3504–3508. [Google Scholar]
- Bernabé, D.G.; Tamae, A.C.; Biasoli, É.R.; Oliveira, S.H.P. Stress Hormones Increase Cell Proliferation and Regulates Interleukin-6 Secretion in Human Oral Squamous Cell Carcinoma Cells. Brain Behav. Immun. 2011, 25, 574–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quốc Lương, K.V.; Nguyễn, L.T.H. The Roles of Beta-Adrenergic Receptors in Tumorigenesis and the Possible Use of Beta-Adrenergic Blockers for Cancer Treatment: Possible Genetic and Cell-Signaling Mechanisms. Cancer Manag. Res. 2012, 4, 431–445. [Google Scholar] [CrossRef] [Green Version]
- Shakhar, G.; Ben-Eliyahu, S. In Vivo Beta-Adrenergic Stimulation Suppresses Natural Killer Activity and Compromises Resistance to Tumor Metastasis in Rats. J. Immunol. 1998, 160, 3251–3258. [Google Scholar] [PubMed]
- Cole, S.W.; Sood, A.K. Molecular Pathways: Beta-Adrenergic Signaling in Cancer. Clin. Cancer Res. 2012, 18, 1201–1206. [Google Scholar] [CrossRef] [Green Version]
- Glasner, A.; Avraham, R.; Rosenne, E.; Benish, M.; Zmora, O.; Shemer, S.; Meiboom, H.; Ben-Eliyahu, S. Improving Survival Rates in Two Models of Spontaneous Postoperative Metastasis in Mice by Combined Administration of a Beta-Adrenergic Antagonist and a Cyclooxygenase-2 Inhibitor. J. Immunol. 2010, 184, 2449–2457. [Google Scholar] [CrossRef] [Green Version]
- Wong, H.P.S.; Yu, L.; Lam, E.K.Y.; Tai, E.K.K.; Wu, W.K.K.; Cho, C.-H. Nicotine Promotes Colon Tumor Growth and Angiogenesis through Beta-Adrenergic Activation. Toxicol. Sci. 2007, 97, 279–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, H.P.S.; Yu, L.; Lam, E.K.Y.; Tai, E.K.K.; Wu, W.K.K.; Cho, C.H. Nicotine Promotes Cell Proliferation via Alpha7-Nicotinic Acetylcholine Receptor and Catecholamine-Synthesizing Enzymes-Mediated Pathway in Human Colon Adenocarcinoma HT-29 Cells. Toxicol. Appl. Pharmacol. 2007, 221, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb, Y.; Sorski, L.; Benish, M.; Levi, B.; Melamed, R.; Ben-Eliyahu, S. Improving Postoperative Immune Status and Resistance to Cancer Metastasis: A Combined Perioperative Approach of Immunostimulation and Prevention of Excessive Surgical Stress Responses. Ann. Surg. 2011, 253, 798–810. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.V.; Kim, S.; Donovan, E.L.; Chen, M.; Gross, A.C.; Webster Marketon, J.I.; Barsky, S.H.; Glaser, R. Norepinephrine Upregulates VEGF, IL-8, and IL-6 Expression in Human Melanoma Tumor Cell Lines: Implications for Stress-Related Enhancement of Tumor Progression. Brain Behav. Immun. 2009, 23, 267–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.M.; Liao, Z.X.; Komaki, R.; Welsh, J.W.; O’Reilly, M.S.; Chang, J.Y.; Zhuang, Y.; Levy, L.B.; Lu, C.; Gomez, D.R. Improved Survival Outcomes with the Incidental Use of Beta-Blockers among Patients with Non-Small-Cell Lung Cancer Treated with Definitive Radiation Therapy. Ann. Oncol. 2013, 24, 1312–1319. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-W.; Shahzad, M.M.K.; Lin, Y.G.; Armaiz-Pena, G.; Mangala, L.S.; Han, H.-D.; Kim, H.-S.; Nam, E.J.; Jennings, N.B.; Halder, J.; et al. Surgical Stress Promotes Tumor Growth in Ovarian Carcinoma. Clin. Cancer Res. 2009, 15, 2695–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, X.; Qi, M.; Peng, C.; Zhou, C.; Su, J.; Zeng, W.; Liu, H.; Zhang, J.; Chen, M.; Shen, M.; et al. Propranolol Enhanced the Anti-Tumor Effect of Sunitinib by Inhibiting Proliferation and Inducing G0/G1/S Phase Arrest in Malignant Melanoma. Oncotarget 2018, 9, 802–811. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, K.R.; Yan, S.X.; Heilbroner, S.P.; Sonett, J.R.; Stoopler, M.B.; Shu, C.; Halmos, B.; Wang, T.J.C.; Hei, T.K.; Cheng, S.K. Effects of β-Adrenergic Antagonists on Chemoradiation Therapy for Locally Advanced Non-Small Cell Lung Cancer. J. Clin. Med. 2019, 8, 575. [Google Scholar] [CrossRef] [Green Version]
- Coelho, M.; Imperatori, A.; Chiaravalli, A.M.; Franzi, F.; Castiglioni, M.; Rasini, E.; Luini, A.; Legnaro, M.; Marino, F.; Ribeiro, L.; et al. Beta1- and Beta2-Adrenoceptors Expression Patterns in Human Non-Small Cell Lung Cancer: Relationship with Cancer Histology. J. Neuroimmune Pharmacol. 2019, 14, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.B.; Le, X.; Heymach, J.V. β-Adrenergic Signaling in Lung Cancer: A Potential Role for Beta-Blockers. J. Neuroimmune Pharmacol. 2020, 15, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.A.; Oster, C.G.; Mayer, M.M.; Avery, M.L.; Audus, K.L. Characterization of the A549 Cell Line as a Type II Pulmonary Epithelial Cell Model for Drug Metabolism. Exp. Cell Res. 1998, 243, 359–366. [Google Scholar] [CrossRef]
- Lieber, M.; Smith, B.; Szakal, A.; Nelson-Rees, W.; Todaro, G. A Continuous Tumor-Cell Line from a Human Lung Carcinoma with Properties of Type II Alveolar Epithelial Cells. Int. J. Cancer 1976, 17, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Okabe, T.; Okamoto, I.; Tamura, K.; Terashima, M.; Yoshida, T.; Satoh, T.; Takada, M.; Fukuoka, M.; Nakagawa, K. Differential Constitutive Activation of the Epidermal Growth Factor Receptor in Non-Small Cell Lung Cancer Cells Bearing EGFR Gene Mutation and Amplification. Cancer Res. 2007, 67, 2046–2053. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Pauer, S.H.; Yong, H.M.; An, S.S.; Liggett, S.B. Β2-Adrenergic Receptors Chaperone Trapped Bitter Taste Receptor 14 to the Cell Surface as a Heterodimer and Exert Unidirectional Desensitization of Taste Receptor Function. J. Biol. Chem. 2016, 291, 17616–17628. [Google Scholar] [CrossRef] [Green Version]
- Hu, P.; He, J.; Liu, S.; Wang, M.; Pan, B.; Zhang, W. Β2-Adrenergic Receptor Activation Promotes the Proliferation of A549 Lung Cancer Cells via the ERK1/2/CREB Pathway. Oncol. Rep. 2016, 36, 1757–1763. [Google Scholar] [CrossRef]
- Frishman, W.H. Clinical Significance of Beta 1-Selectivity and Intrinsic Sympathomimetic Activity in a Beta-Adrenergic Blocking Drug. Am. J. Cardiol. 1987, 59, 33F–37F. [Google Scholar] [CrossRef]
- Mason, R.P.; Giles, T.D.; Sowers, J.R. Evolving Mechanisms of Action of Beta Blockers: Focus on Nebivolol. J. Cardiovasc. Pharmacol. 2009, 54, 123–128. [Google Scholar] [CrossRef]
- Kozanoglu, I.; Yandim, M.K.; Cincin, Z.B.; Ozdogu, H.; Cakmakoglu, B.; Baran, Y. New Indication for Therapeutic Potential of an Old Well-Known Drug (Propranolol) for Multiple Myeloma. J. Cancer Res. Clin. Oncol. 2013, 139, 327–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Işeri, O.D.; Sahin, F.I.; Terzi, Y.K.; Yurtcu, E.; Erdem, S.R.; Sarialioglu, F. Beta-Adrenoreceptor Antagonists Reduce Cancer Cell Proliferation, Invasion, and Migration. Pharm. Biol. 2014, 52, 1374–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazawa, T.; Kaira, K.; Shimizu, K.; Shimizu, A.; Mori, K.; Nagashima, T.; Ohtaki, Y.; Oyama, T.; Mogi, A.; Kuwano, H. Prognostic Significance of Β2-Adrenergic Receptor Expression in Non-Small Cell Lung Cancer. Am. J. Transl. Res. 2016, 8, 5059–5070. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ma, Q.; Wang, Z.; Zhang, M.; Guo, K.; Wang, F.; Wu, E. Β2-Adrenoceptor Blockage Induces G1/S Phase Arrest and Apoptosis in Pancreatic Cancer Cells via Ras/Akt/NFκB Pathway. Mol. Cancer 2011, 10, 146. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Xu, Z.; Marignani, P.A. Single-Cell RNA Sequencing for the Identification of Early-Stage Lung Cancer Biomarkers from Circulating Blood. NPJ Genom. Med. 2021, 6, 87. [Google Scholar] [CrossRef] [PubMed]
- Min, H.-Y.; Boo, H.-J.; Lee, H.J.; Jang, H.-J.; Yun, H.J.; Hwang, S.J.; Smith, J.K.; Lee, H.-J.; Lee, H.-Y. Smoking-Associated Lung Cancer Prevention by Blockade of the Beta-Adrenergic Receptor-Mediated Insulin-like Growth Factor Receptor Activation. Oncotarget 2016, 7, 70936–70947. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Che, X.; Zhao, W.; Zhang, D.; Long, H.; Chaudhary, P.; Li, H. Effects of Propranolol in Combination with Radiation on Apoptosis and Survival of Gastric Cancer Cells in Vitro. Radiat. Oncol. 2010, 5, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Ma, Q.; Shen, S.; Hu, H. Inhibition of Pancreatic Cancer Cell Proliferation by Propranolol Occurs through Apoptosis Induction: The Study of Beta-Adrenoceptor Antagonist’s Anticancer Effect in Pancreatic Cancer Cell. Pancreas 2009, 38, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Čeponytė, U.; Paškevičiūtė, M.; Petrikaitė, V. Comparison of NSAIDs Activity in COX-2 Expressing and Non-Expressing 2D and 3D Pancreatic Cancer Cell Cultures. Cancer Manag. Res. 2018, 10, 1543–1551. [Google Scholar] [CrossRef] [Green Version]
- Paškevičiūtė, M.; Petrikaitė, V. Differences of Statin Activity in 2D and 3D Pancreatic Cancer Cell Cultures. Drug Des. Devel. Ther. 2017, 11, 3273–3280. [Google Scholar] [CrossRef] [Green Version]
- Ciancio, G.; Pollack, A.; Taupier, M.A.; Block, N.L.; Irvin, G.L. Measurement of Cell-Cycle Phase-Specific Cell Death Using Hoechst 33342 and Propidium Iodide: Preservation by Ethanol Fixation. J. Histochem. Cytochem. 1988, 36, 1147–1152. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sidorova, M.; Petrikaitė, V. The Effect of Beta Adrenoreceptor Blockers on Viability and Cell Colony Formation of Non-Small Cell Lung Cancer Cell Lines A549 and H1299. Molecules 2022, 27, 1938. https://doi.org/10.3390/molecules27061938
Sidorova M, Petrikaitė V. The Effect of Beta Adrenoreceptor Blockers on Viability and Cell Colony Formation of Non-Small Cell Lung Cancer Cell Lines A549 and H1299. Molecules. 2022; 27(6):1938. https://doi.org/10.3390/molecules27061938
Chicago/Turabian StyleSidorova, Marina, and Vilma Petrikaitė. 2022. "The Effect of Beta Adrenoreceptor Blockers on Viability and Cell Colony Formation of Non-Small Cell Lung Cancer Cell Lines A549 and H1299" Molecules 27, no. 6: 1938. https://doi.org/10.3390/molecules27061938
APA StyleSidorova, M., & Petrikaitė, V. (2022). The Effect of Beta Adrenoreceptor Blockers on Viability and Cell Colony Formation of Non-Small Cell Lung Cancer Cell Lines A549 and H1299. Molecules, 27(6), 1938. https://doi.org/10.3390/molecules27061938