
Automatic Identification of Myeloperoxidase Natural Inhibitors in Plant Extracts

 ,
,  ,
,  and
and 
Abstract
: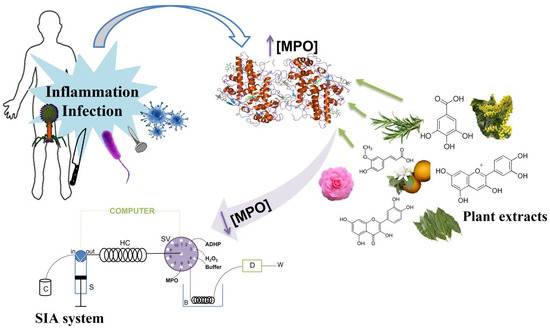
1. Introduction
2. Results and Discussion
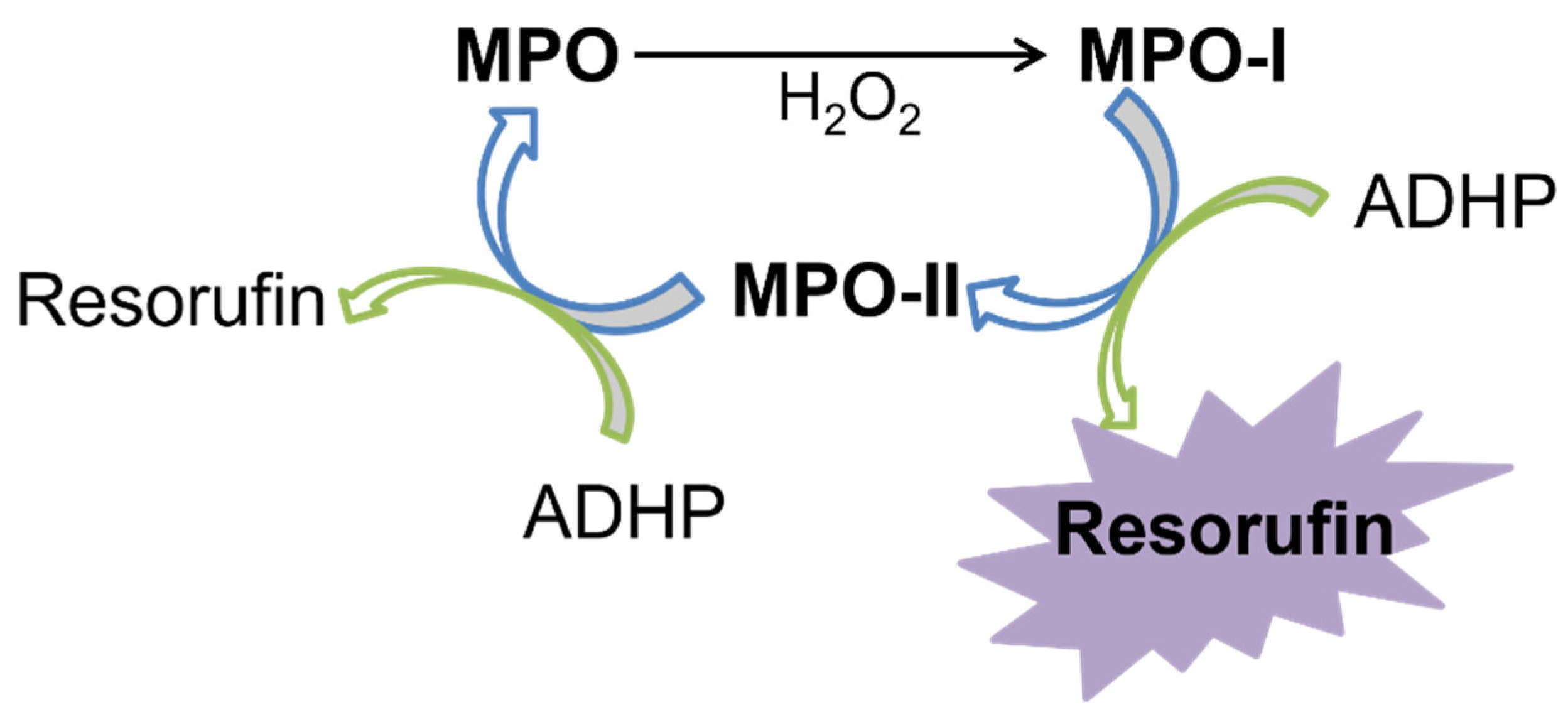
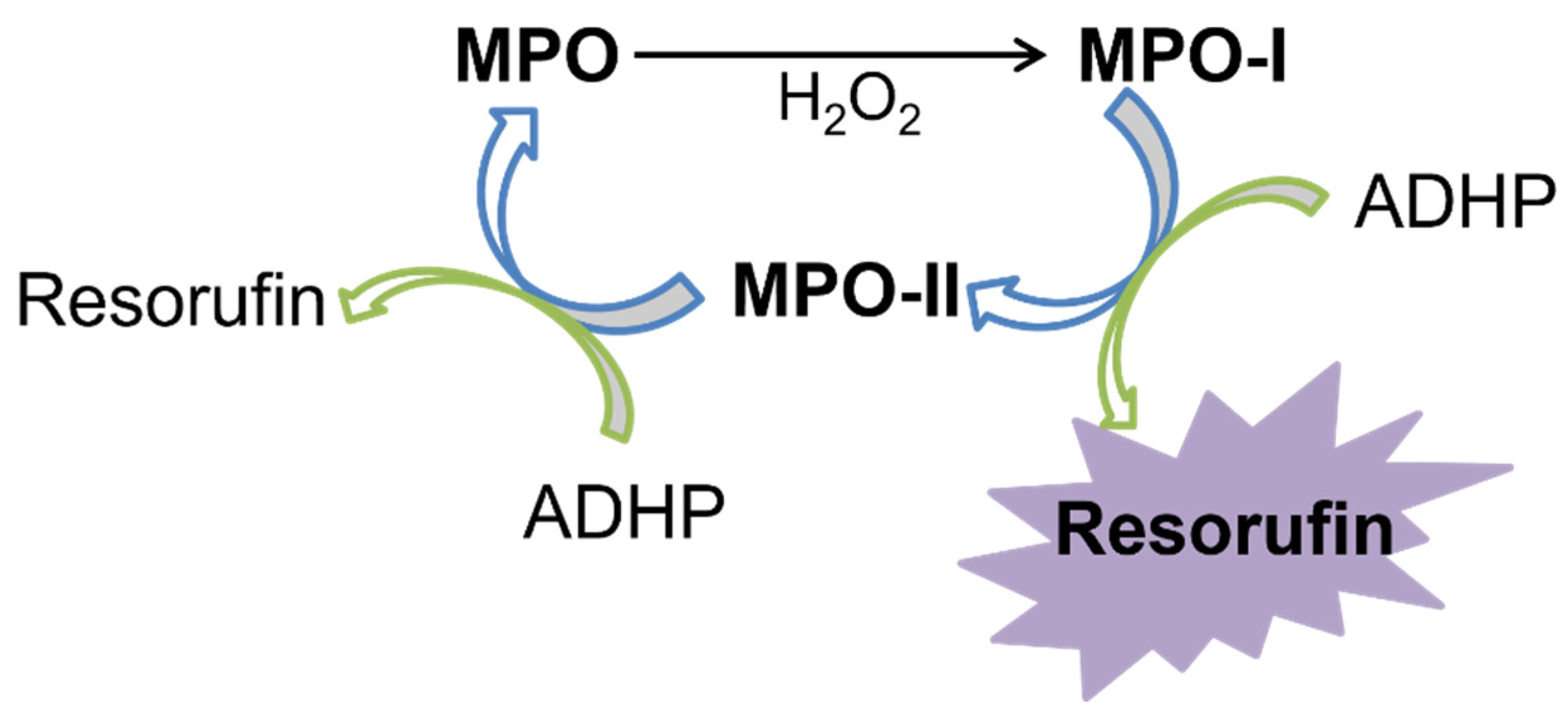
2.1. SIA System Optimization
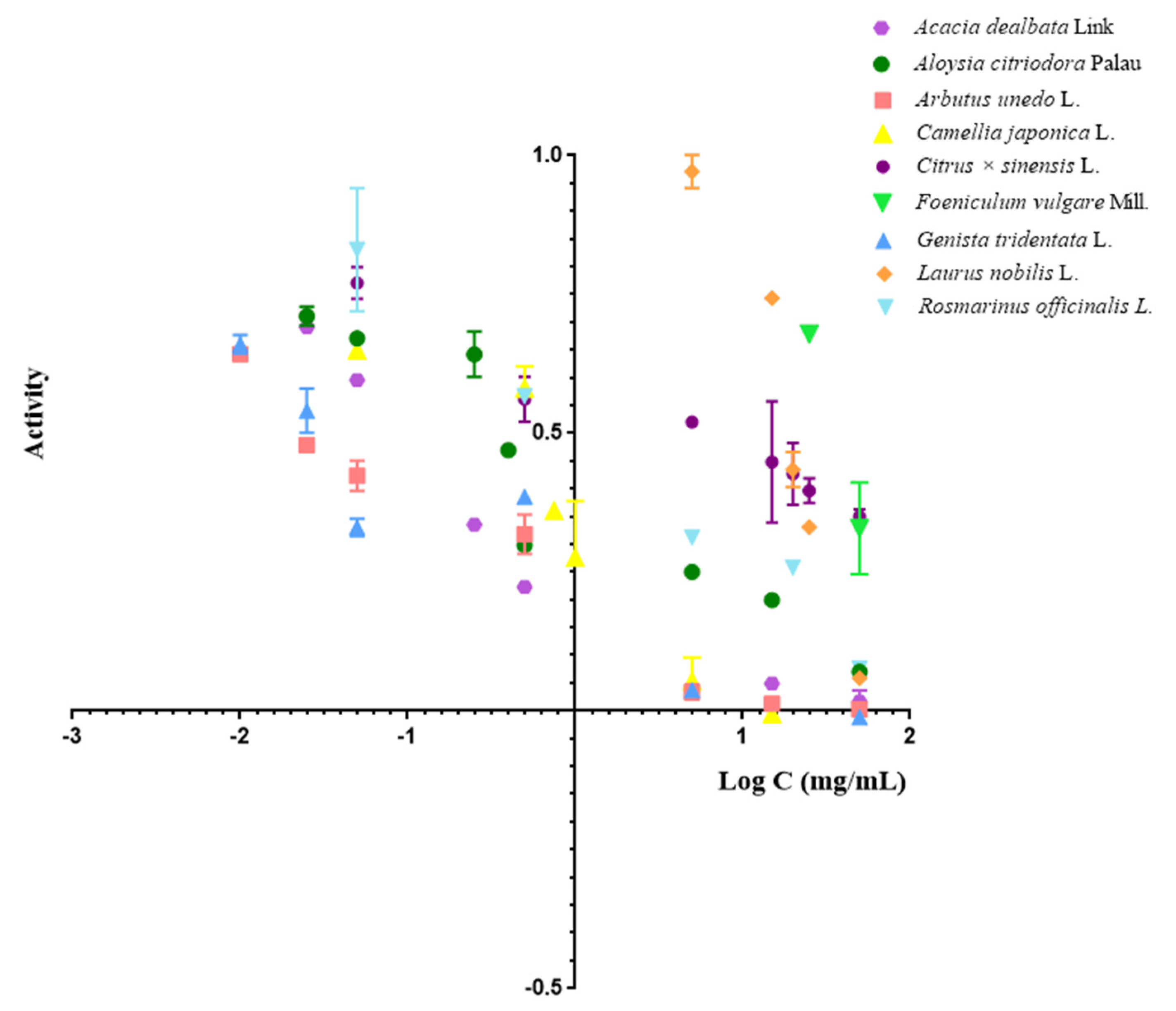
2.2. Evaluation of Plant Extracts Compounds
- Arbutus unedo L.
- Genista tridentata L.
- Aloysia citriodora Palau
- Camellia japónica L.
- Rosmarinus officinalis L.
- Laurus nobilis L.
- 4-Aminobenzohydrazide (positive control)
- Foeniculum vulgare Mill.
3. Materials and Methods
3.1. Reagents
3.2. Sample Treatment
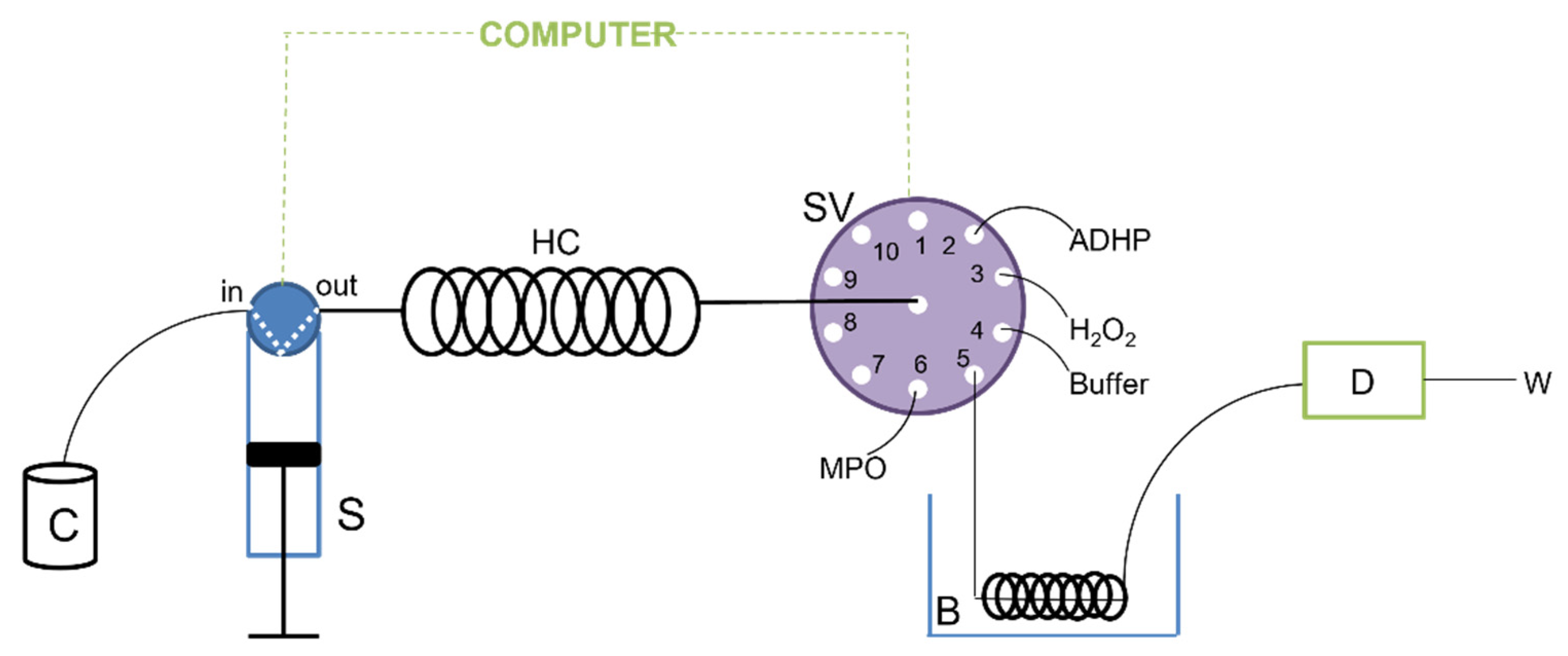
3.3. Apparatus
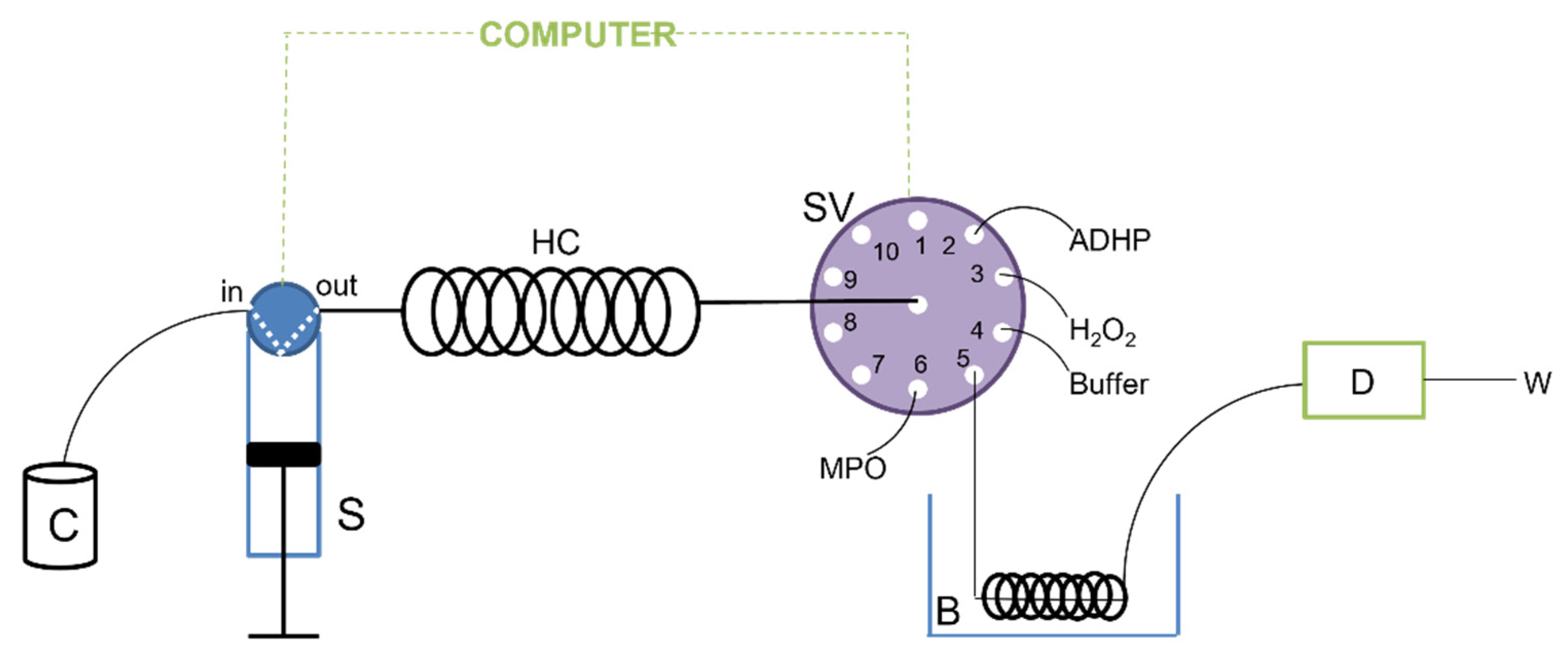
Sequential Injection Analysis Procedure
3.4. Batch Procedure
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ferreira, A.V.; Perelshtein, I.; Perkas, N.; Gedanken, A.; Cunha, J.; Cavaco-Paulo, A. Detection of human neutrophil elastase (HNE) on wound dressings as marker of inflammation. Appl. Microbiol. Biotechnol. 2017, 101, 1443–1454. [Google Scholar] [CrossRef] [PubMed]
- Hasmann, A.; Wehrschuetz-Sigl, E.; Marold, A.; Wiesbauer, H.; Schoeftner, R.; Gewessler, U.; Kandelbauer, A.; Schiffer, D.; Schneider, K.P.; Binder, B.; et al. Analysis of myeloperoxidase activity in wound fluids as a marker of infection. Ann. Clin. Biochem. 2013, 50, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Hasmann, A.; Gewessler, U.; Hulla, E.; Schneider, K.P.; Binder, B.; Francesko, A.; Tzanov, T.; Schintler, M.; Van der Palen, J.; Guebitz, G.M.; et al. Sensor materials for the detection of human neutrophil elastase and cathepsin G activity in wound fluid. Exp. Dermatol. 2011, 20, 508–513. [Google Scholar] [CrossRef]
- Iuonut, A.-M.; Dindelegan, G.C.; Ciuce, C. Proteases as biomarkers in wound healing. Tim. Med. J. 2011, 61, 65–73. [Google Scholar]
- Mota, F.A.R.; Pereira, S.A.P.; Araújo, A.R.T.S.; Passos, M.L.C.; Saraiva, M.L.M.F.S. Biomarkers in the diagnosis of wounds infection: An analytical perspective. TrAC Trends Analyt. Chem. 2021, 143, 116405. [Google Scholar] [CrossRef]
- Tegl, G.; Schiffer, D.; Sigl, E.; Heinzle, A.; Guebitz, G.M. Biomarkers for infection: Enzymes, microbes, and metabolites. Appl. Microbiol. Biotechnol. 2015, 99, 4595–4614. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Milton, A.; Arnold, R.D.; Huang, H.; Smith, F.; Panizzi, J.R.; Panizzi, P. Methods for measuring myeloperoxidase activity toward assessing inhibitor efficacy in living systems. J. Leukoc. Biol. 2016, 99, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Morwinski, I.; Böhme, H.J. Kinetic determination of peroxidase activity of human leukocytes. Z. Med. Lab. Diagn. 1982, 23, 356–360. [Google Scholar]
- de Vargas, F.S.; Almeida, P.D.; de Boleti, A.P.; Pereira, M.M.; de Souza, T.P.; de Vasconcellos, M.C.; Nunez, C.V.; Pohlit, A.M.; Lima, E.S. Antioxidant activity and peroxidase inhibition of Amazonian plants extracts traditionally used as anti-inflammatory. BMC Complem. Altern. Med. 2016, 16, 83. [Google Scholar] [CrossRef] [Green Version]
- Nastasijević, B.; Lazarević-Pašti, T.; Dimitrijević-Branković, S.; Pašti, I.; Vujačić, A.; Joksić, G.; Vasić, V. Inhibition of myeloperoxidase and antioxidative activity of Gentiana lutea extracts. J. Pharm. Biomed. Anal. 2012, 66, 191–196. [Google Scholar] [CrossRef]
- Siedle, B.; Hrenn, A.; Merfort, I. Natural compounds as inhibitors of human neutrophil elastase. Planta Med. 2007, 73, 401–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attiq, A.; Jalil, J.; Husain, K.; Ahmad, W. Raging the War Against Inflammation With Natural Products. Front. Pharm. 2018, 9, 976. [Google Scholar] [CrossRef] [PubMed]
- Jachak, S.M. Cyclooxygenase inhibitory natural products: Current status. Curr. Med. Chem. 2006, 13, 659–678. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Albalat, A.; González, F.V. Natural Products as Cathepsin Inhibitors. Studi Natl. Prod. Chem. 2016, 50, 179–213. [Google Scholar]
- Mehmood, A.; Ishaq, M.; Zhao, L.; Safdar, B.; Rehman, A.U.; Munir, M.; Raza, A.; Nadeem, M.; Iqbal, W.; Wang, C. Natural compounds with xanthine oxidase inhibitory activity: A review. Chem. Biol. Drug Des. 2019, 93, 387–418. [Google Scholar] [CrossRef] [PubMed]
- Kettle, A.J.; Winterbourn, C.C. Mechanism of inhibition of myeloperoxidase by anti-inflammatory drugs. Biochem. Pharm. 1991, 41, 1485–1492. [Google Scholar] [CrossRef]
- Momić, T.; Vujčić, Z.; Vasić, V. Kinetics of inhibition of peroxidase activity of myeloperoxidase by quercetin. Int. J. Chem. Kinet. 2008, 40, 384–394. [Google Scholar] [CrossRef]
- Frumkin, H.; Gerberding, J.L. Toxicological Profile for Phenol; U.S Department of Health and Human Services: Atlanta, GA, USA, 2008.
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Dykes, L.; Rooney, L.W. Phenolic Compounds in Cereal Grains and Their Health Benefits. Cereal Foods World CFW 2007, 52, 105–111. [Google Scholar]
- GutiErrez-Grijalva, E.P.; Ambriz-Pere, D.L.; Leyva-Lopez, N.; Castillo-Lopez, R.I.; Heiedia, J.B. Review: Dietary phenolic compounds, health benefits and bioaccessibility. Arch. Lat. Nutr. 2016, 66, 87–100. [Google Scholar]
- Taamalli, A.; Contreras, M.D.M.; Abu-Reidah, I.M.; Trabelsi, N.; Ben Youssef, N. Quality of Phenolic Compounds: Occurrence, Health Benefits, and Applications in Food Industry. J. Food Qual. 2019, 2019, 9594646. [Google Scholar] [CrossRef]
- Uğur, H.; Çatak, J.; Mızrak, Ö.F.; Çebi, N.; Yaman, M. Determination and evaluation of in vitro bioaccessibility of added vitamin C in commercially available fruit-, vegetable-, and cereal-based baby foods. Food Chem. I 2020, 330, 127166. [Google Scholar] [CrossRef] [PubMed]
- Shiba, Y.; Kinoshita, T.; Chuman, H.; Taketani, Y.; Takeda, E.; Kato, Y.; Naito, M.; Kawabata, K.; Ishisaka, A.; Terao, J.; et al. Flavonoids as substrates and inhibitors of myeloperoxidase: Molecular actions of aglycone and metabolites. Chem. Res. Toxicol. 2008, 21, 1600–1609. [Google Scholar] [CrossRef] [PubMed]
- Tsumbu, C.N.; Deby-Dupont, G.; Tits, M.; Angenot, L.; Frederich, M.; Kohnen, S.; Mouithys-Mickalad, A.; Serteyn, D.; Franck, T. Polyphenol content and modulatory activities of some tropical dietary plant extracts on the oxidant activities of neutrophils and myeloperoxidase. Int. J. Mol. Sci. 2012, 13, 628–650. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Zhang, G.; Liao, Y.; Pan, J.; Gong, D. Dietary Flavonoids as Xanthine Oxidase Inhibitors: Structure-Affinity and Structure-Activity Relationships. J. Agric. Food Chem. 2015, 63, 7784–7794. [Google Scholar] [CrossRef]
- Gonçalves, S.; Moreira, E.; Grosso, C.; Andrade, P.B.; Valentão, P.; Romano, A. Phenolic profile, antioxidant activity and enzyme inhibitory activities of extracts from aromatic plants used in Mediterranean diet. J. Food Sci. Technol. 2017, 54, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Chen, H.; Du, Q.; Shen, J. Targeting Myeloperoxidase (MPO) Mediated Oxidative Stress and Inflammation for Reducing Brain Ischemia Injury: Potential Application of Natural Compounds. Front. Physiol. 2020, 11, 433. [Google Scholar] [CrossRef]
- Lazarević-Pasti, T.; Leskovac, A.; Vasić, V. Myeloperoxidase Inhibitors as Potential Drugs. Curr. Drug Metab. 2015, 16, 168–190. [Google Scholar] [CrossRef]
- Tunali, S.; Boztas, F.Y.; Yanardag, R. The inhibitory effects of plant extracts, vitamins and amino acids on myeloperoxidase activity. J. Fac. Pharm. Istanb. Univ. 2020, 50, 125. [Google Scholar] [CrossRef]
- Khalil, N.M.; Pepato, M.T.; Brunetti, I.L. Free Radical Scavenging Profile and Myeloperoxidase Inhibition of Extracts from Antidiabetic Plants: Bauhinia forficata and Cissus sicyoides. Biol. Res. 2008, 41, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.A.; Mallick, N.; Feroz, Z. Anti-inflammatory effects of Citrus sinensis L.; Citrus paradisi L. and their combinations. Pak. J. Pharm. Sci. 2016, 29, 843–852. [Google Scholar]
- Teke, M.; Sezgintürk, M.K.; Dinçkaya, E. A biosensor based on Bay leaf (Laurus nobilis L.) tissue homogenate: Improvement of the stability characteristics by a simple bio-imprinted technique. Artif. Cells Blood Substit. Immobil. Biotechnol. 2008, 36, 445–456. [Google Scholar] [CrossRef]
- Sahin Basak, S.; Candan, F. Effect of Laurus nobilis L. Essential Oil and its Main Components on α-glucosidase and Reactive Oxygen Species Scavenging Activity. Iran. J. Pharm. Res. 2013, 12, 367–379. [Google Scholar]
- Oboh, G.; Olasehinde, T.; Ademosun, A. Inhibition of Enzymes linked to Type-2 Diabetes and Hypertension by Essential oils from Peels of orange and lemon. Int. J. Food Prop. 2017, 20, S586–S594. [Google Scholar] [CrossRef] [Green Version]
- Subehan; Zaidi, S.F.; Kadota, S.; Tezuka, Y. Inhibition on human liver cytochrome P450 3A4 by constituents of fennel (Foeniculum vulgare): Identification and characterization of a mechanism-based inactivator. J. Agric. Food Chem. 2007, 55, 10162–10167. [Google Scholar] [CrossRef]
- El Haouari, M.; López, J.J.; Mekhfi, H.; Rosado, J.A.; Salido, G.M. Antiaggregant effects of Arbutus unedo extracts in human platelets. J. Ethnopharmacol. 2007, 113, 325–331. [Google Scholar] [CrossRef]
- Ruzicka, J.; Marshall, G.D. Sequential injection: A new concept for chemical sensors, process analysis and laboratory assays. Anal. Chim. Acta 1990, 237, 329–343. [Google Scholar] [CrossRef]
- ABCAM. Myeloperoxidase Inhibitor Screening Assay Kit; Abcamwebsite: Waltham, MA, USA, 2019. [Google Scholar]
- Eisenthdal, R.; Danson, M.J. Enzyme Assays—A pratical Approach; Oxford University Press: Oxford, UK, 2002; pp. 19–20. [Google Scholar]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) Models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef]
- Kyselova, Z. Toxicological aspects of the use of phenolic compounds in disease prevention. Interdiscip. Toxicol. 2011, 4, 173–183. [Google Scholar] [CrossRef]
- Malle, E.; Furtmüller, P.G.; Sattler, W.; Obinger, C. Myeloperoxidase: A target for new drug development? Br. J. Pharm. 2007, 152, 838–854. [Google Scholar] [CrossRef] [Green Version]
- Anvisa, M.D.S.E. Monografia da Espécie Foeniculum Vulgare Mill; Saúde, M.D., Ed.; Ed. Brasília: Brasília, Brazil, 2015. [Google Scholar]
- Badgujar, S.B.; Patel, V.V.; Bandivdekar, A.H. Foeniculum vulgare Mill: A review of its botany, phytochemistry, pharmacology, contemporary application, and toxicology. BioMed Res. Int. 2014, 2014, 842674. [Google Scholar] [CrossRef] [Green Version]
- Salami, M.; Rahimmalek, M.; Ehtemam, M.H. Inhibitory effect of different fennel (Foeniculum vulgare) samples and their phenolic compounds on formation of advanced glycation products and comparison of antimicrobial and antioxidant activities. Food Chem. 2016, 213, 196–205. [Google Scholar] [CrossRef]
- Miguel, M.; Faleiro, M.L.; Guerreiro, A.; Antunes, M. Arbutus unedo L.: Chemical and Biological Properties. Molecules 2014, 19, 15799–15823. [Google Scholar] [CrossRef] [Green Version]
- Macchioni, V.; Santarelli, V.; Carbone, K. Phytochemical Profile, Antiradical Capacity and α-Glucosidase Inhibitory Potential of Wild Arbutus unedo L. Fruits from Central Italy: A Chemometric Approach. Plants 2020, 9, 12. [Google Scholar] [CrossRef]
- Sharma, H.A.; Barber, J.T.; Ensley, H.E.; Polito, M.A. A comparison of the toxicity and metabolism of phenol and chlorinated phenols by Lemna gibba, with special reference to 2,4,5-trichlorophenol. Environ. Toxicol. Chem. 1997, 16, 346–350. [Google Scholar] [CrossRef]
- Flambó, D. Biological Activities of Flavonoids: Antimicrobial Activity; Fernando Pessoa University: Porto, Portugal, 2013. [Google Scholar]
- Pawlowska, A.M.; De Leo, M.; Braca, A. Phenolics of Arbutus unedo L. (Ericaceae) fruits: Identification of anthocyanins and gallic acid derivatives. J. Agric. Food Chem. 2006, 54, 10234–10238. [Google Scholar] [CrossRef]
- Fortalezas, S.; Tavares, L.; Pimpão, R.; Tyagi, M.; Pontes, V.; Alves, P.M.; McDougall, G.; Stewart, D.; Ferreira, R.B.; Santos, C.N. Antioxidant Properties and Neuroprotective Capacity of Strawberry Tree Fruit (Arbutus unedo). Nutrients 2010, 2, 214–229. [Google Scholar] [CrossRef]
- Dua, A.; Garg, G.; Mahajan, R. Polyphenols, flavonoids and antimicrobial properties of methanolic extract of fennel (Foeniculum vulgare Miller). Eur. J. Exp. Biol. 2013, 3, 203–208. [Google Scholar]
- Konovalov, D.; Alieva, N. Phenolic compounds of laurus nobilis (review). Pharm. Pharm. 2019, 7, 244–259. [Google Scholar] [CrossRef]
- Stefanova, G.; Girova, T.; Gochev, V.; Stoyanova, M.; Petkova, Z.; Stoyanova, A.; Zheljazkov, V.D. Comparative study on the chemical composition of laurel (Laurus nobilis L.) leaves from Greece and Georgia and the antibacterial activity of their essential oil. Heliyon 2020, 6, e05491. [Google Scholar] [CrossRef]
- Gao, J.; Hu, J.; Hu, D.; Yang, X. A Role of Gallic Acid in Oxidative Damage Diseases: A Comprehensive Review. Nat. Prod. Commun. 2019, 14, 1934578X19874174. [Google Scholar] [CrossRef] [Green Version]
- Roriz, C.L.; Barros, L.; Carvalho, A.M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Pterospartum tridentatum, Gomphrena globosa and Cymbopogon citratus: A phytochemical study focused on antioxidant compounds. Food Res. Int. 2014, 62, 684–693. [Google Scholar] [CrossRef] [Green Version]
- Aires, A.; Marrinhas, E.; Carvalho, R.; Dias, C.; Saavedra, M.J. Phytochemical Composition and Antibacterial Activity of Hydroalcoholic Extracts of Pterospartum tridentatum and Mentha pulegium against Staphylococcus aureus Isolates. BioMed Res. Int. 2016, 2016, 5201879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitor, R.F.; Mota-Filipe, H.; Teixeira, G.; Borges, C.; Rodrigues, A.I.; Teixeira, A.; Paulo, A. Flavonoids of an extract of Pterospartum tridentatum showing endothelial protection against oxidative injury. J. Ethnopharmacol. 2004, 93, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.S.A. Plantas Medicinais: Um Estudo Etnobotânico em Alguns Concelhos do Distrito de Vila Real; Universidade da Beira Interior: Vila Real, Spain, 2017. [Google Scholar]
- Kadhom, I.M.; Ibrahim, N.S.; Al-zubaidy, N.A. Estimation of Phenolic Compounds And Evaluation Of Their Antioxidant Activity Of Some Parts Of The Orange Plant (Citrus sinensis, L.). Eur. J. Mol. Clin. Med. 2020, 7, 4811–4822. [Google Scholar]
- Kheiria, H.; Sotomayor, J.; Jordán, M.; Bouzid, S. Identification and Quantification of Phenolic Compounds of Tunisian Rosmarinus officinalis L. Asian J. Chem. 2013, 25, 9299–9301. [Google Scholar]
- Bentahar, A.; Bouaziz, A.; Djidel, S.; Khennouf, S. Phenolic content and antioxidant activity of ethanolic extracts from Citrus sinensis L. and Citrus reticulata L. fruits. J. Drug Deliv. Ther. 2020, 10, 308–313. [Google Scholar] [CrossRef]
- Adomako-Bonsu, A.; Chan, S.; Pratten, M.; Fry, J. Antioxidant activity of rosmarinic acid and its principal metabolites in chemical and cellular systems: Importance of physico-chemical characteristics. Toxicol. Vitr. 2017, 40, 248–255. [Google Scholar] [CrossRef]
- Christensen, R.D.; Rothstein, G. Neutrophil myeloperoxidase concentration: Changes with development and during bacterial infection. Pediatr. Res. 1985, 19, 1278–1282. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name of Plant Extract | Common Name of Plant Extract | IC50 (mg/mL) ± SD |
|---|---|---|
| Acacia dealbata Link | Mimosa; Acacia | 0.086 ± 0.001 |
| Aloysia citriodora Palau | Lemon verbena | 0.282 ± 0.040 |
| Arbutus unedo L. | Strawberry tree | 0.029 ± 0.002 |
| Camellia Japonica L. | Japanese camellia | 0.303 ± 0.066 |
| Citrus × sinensis | Orange | 5.4 ± 3.1 |
| Foeniculum vulgare Mill. | Fennel | 35.4 ± 3.5 |
| Genista tridentata L. | Prickly broom | 0.032 ± 0.004 |
| Laurus nobilis L. | Laurel | 19.62 ± 0.28 |
| Rosmarinus officinalis L. | Rosemary | 1.02 ± 0.28 |
| Plant Extract | IC50 (mg/mL) Obtain in SIA ± SD | IC50 (mg/mL) Obtain in Batch ± SD |
|---|---|---|
| Aloysia citriodora Palau | 0.282 ± 0.040 | 0.258 ± 0.029 |
| Arbutus unedo L. | 0.029 ± 0.002 | 0.004 ± 0.001 |
| Camellia Japonica L. | 0.303 ± 0.066 | 0.290 ± 0.083 |
| Foeniculum vulgare Mill. | 35.4 ± 3.5 | 50.6 ± 9.7 |
| Genista tridentata L. | 0.032 ± 0.004 | 0.044 ± 0.013 |
| Laurus nobilis L. | 19.62 ± 0.28 | 13.7 ± 1.4 |
| Rosmarinus officinalis L. | 1.02 ± 0.28 | 1.67 ± 0.29 |
| 4-Aminobenzohydrazide (positive control) | 24.8 ± 4.9 | 17.4 ± 4.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mota, F.A.R.; Pereira, S.A.P.; Araújo, A.R.T.S.; Gullón, B.; Passos, M.L.C.; Saraiva, M.L.M.F.S. Automatic Identification of Myeloperoxidase Natural Inhibitors in Plant Extracts. Molecules 2022, 27, 1825. https://doi.org/10.3390/molecules27061825
Mota FAR, Pereira SAP, Araújo ARTS, Gullón B, Passos MLC, Saraiva MLMFS. Automatic Identification of Myeloperoxidase Natural Inhibitors in Plant Extracts. Molecules. 2022; 27(6):1825. https://doi.org/10.3390/molecules27061825
Chicago/Turabian StyleMota, Fátima A. R., Sarah A. P. Pereira, André R. T. S. Araújo, Beatriz Gullón, Marieta L. C. Passos, and Maria Lúcia M. F. S. Saraiva. 2022. "Automatic Identification of Myeloperoxidase Natural Inhibitors in Plant Extracts" Molecules 27, no. 6: 1825. https://doi.org/10.3390/molecules27061825
APA StyleMota, F. A. R., Pereira, S. A. P., Araújo, A. R. T. S., Gullón, B., Passos, M. L. C., & Saraiva, M. L. M. F. S. (2022). Automatic Identification of Myeloperoxidase Natural Inhibitors in Plant Extracts. Molecules, 27(6), 1825. https://doi.org/10.3390/molecules27061825