
Phenolics as GABAA Receptor Ligands: An Updated Review


Abstract
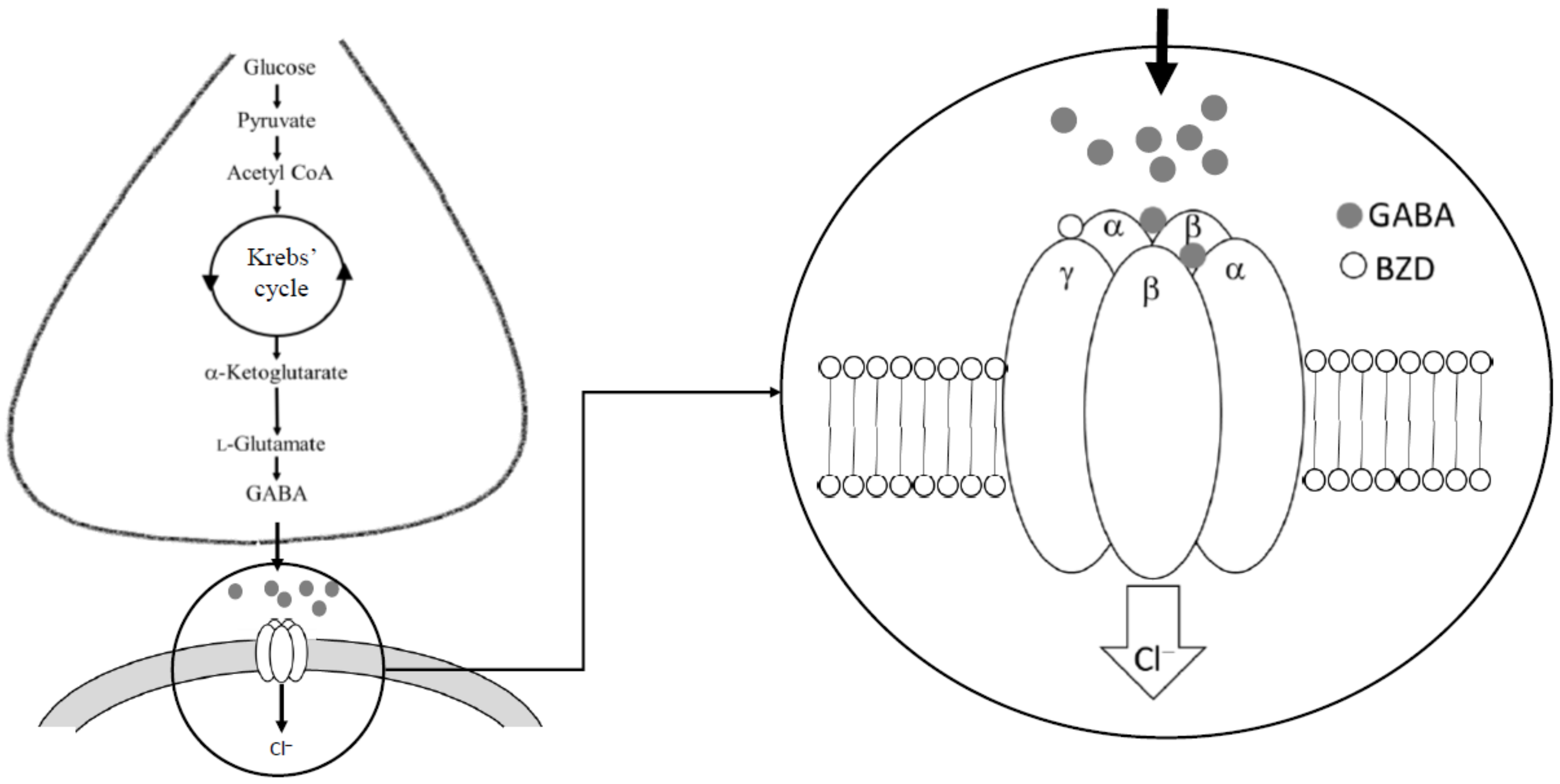
1. Introduction
2. Natural Products as Potential GABA Modulators
2.1. Phenolics as Potential GABAergic Compounds
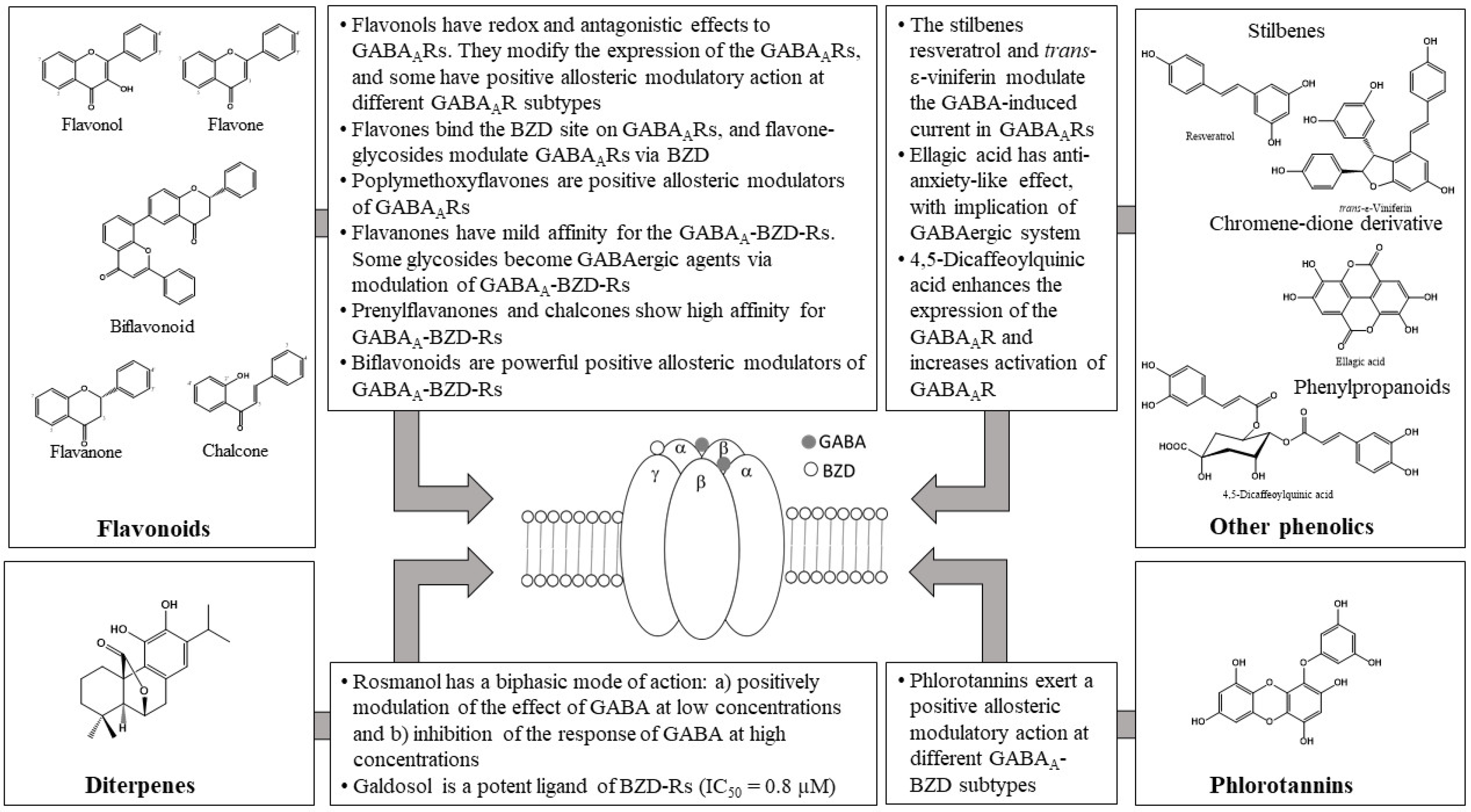
2.2. Flavonoids and GABAA Receptors
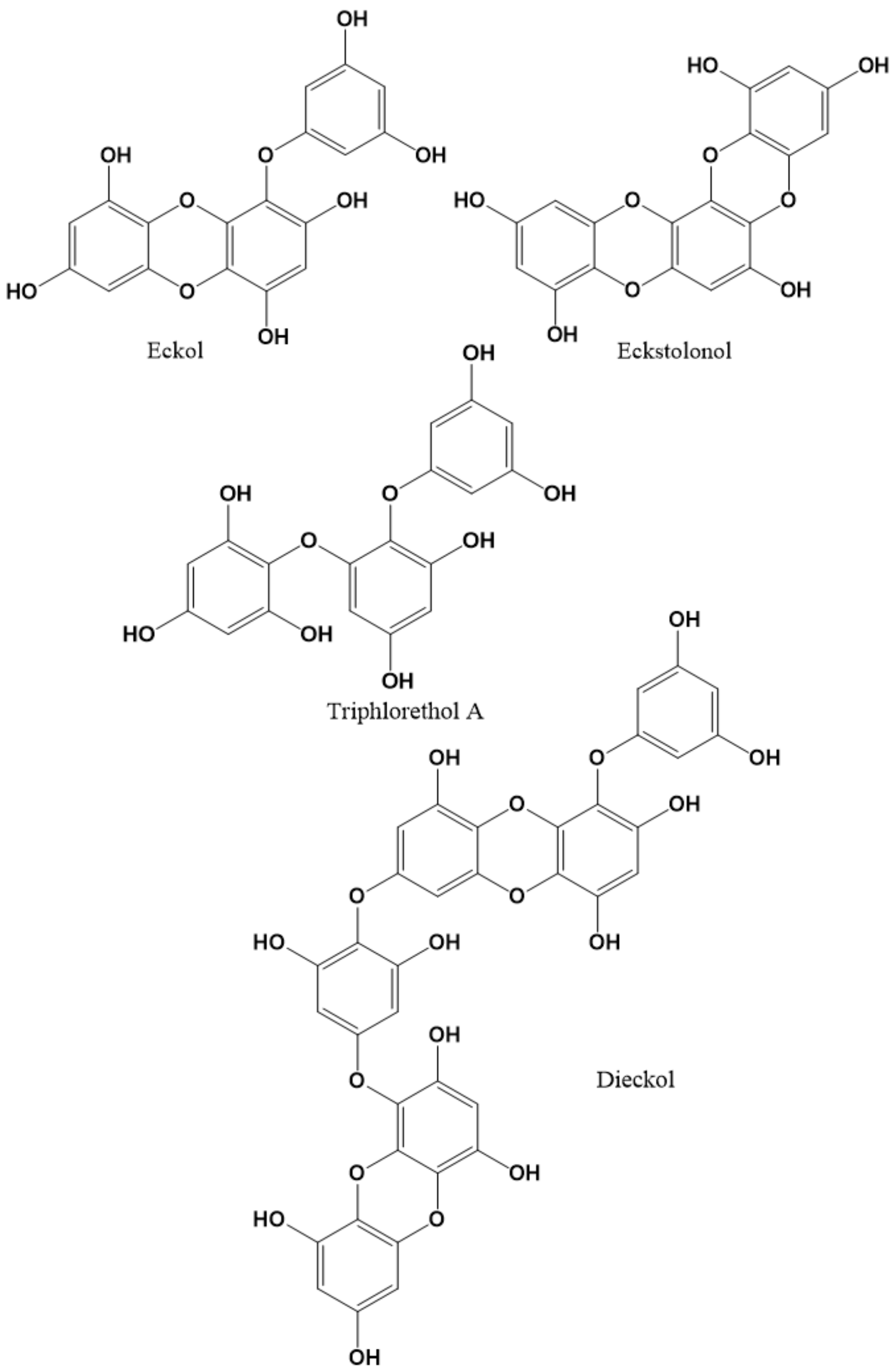
2.3. Phlorotannins as Modulators of GABAARs
2.4. Other Types of Compounds Active on GABAARs
2.5. Discussion and Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Awapara, J.; Landua, A.J.; Fuerst, R.; Seale, B. Free γ-aminobutyric acid in brain. J. Biol. Chem. 1950, 187, 35–39. [Google Scholar] [CrossRef]
- Roberts, E.; Frankel, S. γ-Aminobutyric acid in brain: Its formation from glutamic acid. J. Biol. Chem. 1950, 187, 55–63. [Google Scholar] [CrossRef]
- Johnston, G.A.R. GABAA receptor channel pharmacology. Curr. Pharm. Des. 2005, 11, 1867–1885. [Google Scholar] [CrossRef] [PubMed]
- Lydiard, R.B. The role of GABA in anxiety disorders. J. Clin. Psychiatry 2003, 64 (Suppl. 3), 21–27. [Google Scholar]
- Prévot, T.; Sibille, E. Altered GABA-mediated information processing and cognitive dysfunctions in depression and other brain disorders. Mol. Psychiatry 2021, 26, 151–167. [Google Scholar] [CrossRef]
- Richards, A.; Kanady, J.C.; Neylan, T. Sleep disturbance in PTSD and other anxiety-related disorders: An updated review of clinical features, physiological characteristics, and psychological and neurobiological mechanisms. Neuropsychopharmacology 2020, 45, 55–73. [Google Scholar] [CrossRef]
- Treiman, D.M. GABAergic mechanisms in epilepsy. Epilepsia 2001, 42 (Suppl. 3), 8–12. [Google Scholar] [CrossRef]
- Lener, M.S.; Niciu, M.J.; Ballard, E.D.; Park, M.; Park, L.T.; Nugent, A.C.; Zarate, C.A. Glutamate and gamma-aminobutyric acid systems in the pathophysiology of major depression and antidepressant response to ketamine. Biol. Psychiatry 2017, 81, 886–897. [Google Scholar] [CrossRef]
- Adell, A. Brain NMDA Receptors in schizophrenia and depression. Biomolecules 2020, 10, E947. [Google Scholar] [CrossRef]
- Solas, M.; Puerta, E.; Ramirez, M.J. Treatment options in Alzheimer´s disease: The GABA story. Curr. Pharm. Des. 2015, 21, 4960–4971. [Google Scholar] [CrossRef]
- Muñoz, M.D.; de la Fuente, N.; Sánchez-Capelo, A. TGF-β/Smad3 signalling modulates GABA neurotransmission: Implications in Parkinson’s disease. Int. J. Mol. Sci. 2020, 21, E590. [Google Scholar] [CrossRef] [PubMed]
- Mousavi Majd, A.; Ebrahim Tabar, F.; Afghani, A.; Ashrafpour, S.; Dehghan, S.; Gol, M.; Ashrafpour, M.; Pourabdolhossein, F. Inhibition of GABA A receptor improved spatial memory impairment in the local model of demyelination in rat hippocampus. Behav. Brain Res. 2018, 336, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Escartín Pérez, R.E.; Mancilla Díaz, J.M.; Cortés Salazar, F.; López Alonso, V.E.; Florán Garduño, B. CB1/5-HT/GABA interactions and food intake regulation. Prog. Brain Res. 2021, 259, 177–196. [Google Scholar] [PubMed]
- Brohan, J.; Goudra, B.G. The role of GABA receptor agonists in anesthesia and sedation. CNS Drugs 2017, 31, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xiong, B.R.; Zhang, L.Q.; Huang, X.; Yuan, X.; Tian, Y.K.; Tian, X.B. The role of the GABAergic system in diseases of the central nervous system. Neuroscience 2021, 470, 88–99. [Google Scholar] [CrossRef]
- Watanabe, M.; Maemura, K.; Kanbara, K.; Tamayama, T.; Hayasaki, H. GABA and GABA receptors in the central nervous system and other organs. Int. Rev. Cytol. 2002, 213, 1–47. [Google Scholar]
- Serrano-Regal, M.P.; Bayón-Cordero, L.; Ordaz, R.P.; Garay, E.; Limon, A.; Arellano, R.O.; Matute, C.; Sánchez-Gómez, M.V. Expression and function of GABA Receptors in myelinating cells. Front Cell. Neurosci. 2020, 14, 256. [Google Scholar] [CrossRef]
- Owens, D.F.; Kriegstein, A.R. Is there more to GABA than synaptic inhibition? Nat. Rev. Neurosci. 2002, 3, 715–727. [Google Scholar] [CrossRef]
- Lee, S.E.; Lee, Y.; Lee, G.H. The regulation of glutamic acid decarboxylases in GABA neurotransmission in the brain. Arch. Pharm. Res. 2019, 42, 1031–1039. [Google Scholar] [CrossRef]
- Olsen, R.W.; Betz, H. GABA and Glycine. In Basic Neurochemistry: Molecular, Cellular and Medical Aspects, 7th ed.; Siegel, G.J., Albers, R.W., Brady, S.T., Price, D.L., Eds.; Elsevier Academic Press: Burlington, VT, USA, 2006; pp. 291–301. [Google Scholar]
- Jung, H.Y.; Kwon, H.J.; Kim, W.; Nam, S.M.; Kim, J.W.; Hahn, K.R.; Yoo, D.Y.; Yoon, Y.S.; Choi, S.Y.; Kim, D.W.; et al. Role of pyridoxine in GABA synthesis and degradation in the hippocampus. Tissue Cell. 2019, 61, 72–78. [Google Scholar] [CrossRef]
- Ghit, A.; Assal, D.; Al-Shami, A.S.; Hussein, D.E.E. GABAA receptors: Structure, function, pharmacology, and related disorders. J. Genet. Eng. Biotechnol. 2021, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Łątka, K.; Jończyk, J.; Bajda, M. γ-Aminobutyric acid transporters as relevant biological target: Their function, structure, inhibitors and role in the therapy of different diseases. Int. J. Biol. Macromol. 2020, 158, 750–772. [Google Scholar] [CrossRef] [PubMed]
- Doyon, N.; Vinay, L.; Prescott, S.A.; De Koninck, Y. Chloride regulation: A dynamic equilibrium crucial for synaptic inhibition. Neuron 2016, 89, 1157–1172. [Google Scholar] [CrossRef]
- Sieghart, W.; Savić, M.M. International Union of Basic and Clinical Pharmacology. CVI: GABAA receptor subtype- and function-selective ligands: Key issues in translation to humans. Pharmacol. Rev 2018, 70, 836–878. [Google Scholar] [CrossRef] [PubMed]
- Wasowski, C.; Marder, M. Flavonoids as GABAA receptor ligands: The whole story? J. Exp. Pharmacol. 2012, 4, 9–24. [Google Scholar] [PubMed]
- Waldvogel, H.J.; Faull, R.L.M. The diversity of GABAA receptor subunit distribution in the normal and Huntington’s disease human brain. Adv. Pharmacol. 2015, 73, 223–264. [Google Scholar] [PubMed]
- Vogt, K. Diversity in GABAergic signaling. Adv. Pharmacol. 2015, 73, 203–222. [Google Scholar]
- Chen, X.; Keramidas, A.; Lynch, J.W. Physiological and pharmacological properties of inhibitory postsynaptic currents mediated by α5β1γ2, α5β2γ2 and α5β3γ2 GABAA receptors. Neuropharmacology 2017, 125, 243–253. [Google Scholar] [CrossRef]
- Nilsson, J.; Sterner, O. Modulation of GABAA receptors by natural products and the development of novel synthetic ligands for the benzodiazepine binding site. Curr. Drug Targets. 2011, 12, 1674–1688. [Google Scholar] [CrossRef]
- Hanrahan, J.R.; Chebib, M.; Johnston, G.A. Flavonoid modulation of GABAA receptors. Br. J. Pharmacol. 2011, 163, 234–245. [Google Scholar] [CrossRef]
- Wang, F.; Shing, M.; Huen, Y.; Tsang, S.Y.; Xue, H. Neuroactive flavonoids interacting with GABAA receptor complex. Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Dias, R.; Sheppard, W.F.A.; Fradley, R.L.; Garrett, E.M.; Stanley, J.L.; Tye, S.J.; Goodacre, S.; Lincoln, R.J.; Cook, S.M.; Conley, R.; et al. Evidence for a significant role of α3-containing GABAA receptors in mediating the anxiolytic effects of benzodiazepines. J. Neurosci. 2005, 25, 10682–10688. [Google Scholar] [CrossRef] [PubMed]
- Niturad, C.E.; Lev, D.; Kalscheuer, V.M.; Charzewska, A.; Schubert, J.; Lerman-Sagie, T.; Kroes, H.Y.; Oegema, R.; Traverso, M.; Specchio, N.; et al. Rare GABRA3 variants are associated with epileptic seizures, encephalopathy and dysmorphic features. Brain 2017, 140, 2879–2894. [Google Scholar] [PubMed]
- Syed, P.; Durisic, N.; Harvey, R.J.; Sah, P.; Lynch, J.W. Effects of GABAA receptor α3 subunit epilepsy mutations on inhibitory synaptic signaling. Front. Mol. Neurosci. 2020, 13, 602559. [Google Scholar] [CrossRef]
- Collinson, N.; Kuenzi, F.M.; Jarolimek, W.; Maubach, K.A.; Cothliff, R.; Sur, C.; Smith, A.; Otu, F.M.; Howell, O.; Atack, J.R.; et al. Enhanced learning and memory and altered GABAergic synaptic transmission in mice lacking the alpha 5 subunit of the GABAA receptor. J. Neurosci. 2002, 22, 5572–5580. [Google Scholar] [CrossRef]
- Rudolph, U.; Möhler, H. GABA-based therapeutic approaches: GABAA receptor subtype functions. Curr. Opin. Pharmacol. 2006, 6, 18–23. [Google Scholar] [CrossRef]
- Yamatsu, A.; Yamashita, Y.; Maru, I.; Yang, J.; Tatsuzaki, J.; Kim, M. The improvement of sleep by oral intake of GABA and Apocynum venetum leaf extract. J. Nutr. Sci. Vitaminol. 2015, 61, 182–187. [Google Scholar] [CrossRef]
- Mineo, L.; Concerto, C.; Patel, D.; Mayorga, T.; Paula, M.; Chusid, E.; Aguglia, E.; Battaglia, F. Valeriana officinalis root extract modulates cortical excitatory circuits in humans. Neuropsychobiology 2017, 75, 46–51. [Google Scholar] [CrossRef]
- Sarris, J.; Byrne, G.J.; Bousman, C.A.; Cribb, L.; Savage, K.M.; Holmes, O.; Murphy, J.; Macdonald, P.; Short, A.; Nazareth, S.; et al. Kava for generalised anxiety disorder: A 16-week double-blind, randomised, placebo-controlled study. Aust. N.Z.J. Psychiatry 2020, 54, 288–297. [Google Scholar] [CrossRef]
- Kennedy, D.; Okello, E.; Chazot, P.; Howes, M.J.; Ohiomokhare, S.; Jackson, P.; Haskell-Ramsay, C.; Khan, J.; Forster, J.; Weightman, E. Volatile terpenes and brain function: Investigation of the cognitive and mood effects of Mentha × piperita L. essential oil with In vitro properties relevant to central nervous system function. Nutrients 2018, 10, 1029. [Google Scholar] [CrossRef]
- Andrade, J.C.; Monteiro, Á.B.; Andrade, H.H.N.; Gonzaga, T.K.S.N.; Silva, P.R.; Alves, D.N.; Castro, R.D.; Maia, M.S.; Scotti, M.T.; Sousa, D.P.; et al. Involvement of GABAA receptors in the anxiolytic-like effect of hydroxycitronellal. Biomed. Res. Int. 2021, 2021, 9929805. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Che, Y.; Wang, H. Bergamot essential oil attenuate aluminum-induced anxiety-like behavior through antioxidation, anti-inflammatory and GABA regulation in rats. Food Chem. Toxicol. 2020, 145, 111766. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Lee, Y.; Lee, H.J.; Kwon, Y.S.; Chun, W. In silico screening of GABA aminotransferase inhibitors from the constituents of Valeriana officinalis by molecular docking and molecular dynamics simulation study. J. Mol. Model. 2020, 26, 228. [Google Scholar] [CrossRef] [PubMed]
- Niu, W.H.; Wu, F.; Cao, W.Y.; Wu, Z.G.; Chao, Y.C.; Liang, C. Network pharmacology for the identification of phytochemicals in traditional Chinese medicine for COVID-19 that may regulate interleukin-6. Biosci. Rep. 2021, 41, 20202583. [Google Scholar] [CrossRef]
- Jahani, R.; Mojab, F.; Mahboubi, A.; Nasiri, A.; Tahamtani, A.; Faizi, M. An in-vivo study on anticonvulsant, anxiolytic, and sedative-hypnotic effects of the polyphenol-rich Thymus kotschyanus extract; evidence for the involvement of GABAA receptors. Iran J. Pharm. Res. 2019, 18, 1456–1465. [Google Scholar]
- Jäger, A.K.; Saaby, L. Flavonoids and the CNS. Molecules 2011, 16, 1471–1485. [Google Scholar] [CrossRef]
- Cho, S.; Yang, H.; Jeon, Y.J.; Lee, C.J.; Jin, Y.H.; Baek, N.I.; Kim, D.; Kang, S.M.; Yoon, M.; Yong, H.; et al. Phlorotannins of the edible brown seaweed Ecklonia cava Kjellman induce sleep via positive allosteric modulation of gamma-aminobutyric acid type A-benzodiazepine receptor: A novel neurological activity of seaweed polyphenols. Food Chem. 2012, 132, 1133–1142. [Google Scholar] [CrossRef]
- Cho, S.; Han, D.; Kim, S.B.; Yoon, M.; Yang, H.; Jin, Y.H.; Jo, J.; Yong, H.; Lee, S.H.; Jeon, Y.J.; et al. Depressive effects on the central nervous system and underlying mechanism of the enzymatic extract and its phlorotannin-rich fraction from Ecklonia cava edible brown seaweed. Biosci. Biotechnol. Biochem. 2012, 76, 163–168. [Google Scholar] [CrossRef]
- Cho, S.; Yoon, M.; Pae, A.N.; Jin, Y.H.; Cho, N.C.; Takata, Y.; Urade, Y.; Kim, S.; Kim, J.S.; Yang, H.; et al. Marine polyphenol phlorotannins promote non-rapid eye movement sleep in mice via the benzodiazepine site of the GABAA receptor. Psychopharmacology 2014, 231, 2825–2837. [Google Scholar] [CrossRef]
- Kwon, S.; Yoon, M.; Lee, J.; Moon, K.D.; Kim, D.; Kim, S.B.; Cho, S. A standardized phlorotannin supplement attenuates caffeine-induced sleep disruption in mice. Nutrients 2019, 11, 556. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Phenolics: From chemistry to biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Free radicals and antioxidants—quo vadis? Trends Pharmacol. Sci. 2011, 32, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Poljsak, B.; Šuput, D.; Milisav, I. Achieving the balance between ROS and antioxidants: When to use the synthetic antioxidants. Oxid. Med. Cell Longev. 2013, 2013, 956792. [Google Scholar] [CrossRef] [PubMed]
- Azzi, A. Antioxidants: Wonder drugs or quackery? Biofactors 2017, 43, 785–788. [Google Scholar] [CrossRef]
- Jucá, M.M.; Filho, F.M.S.C.; de Almeida, J.C.; Mesquita, D.D.S.; Barriga, J.R.M.; Dias, K.C.F.; Barbosa, T.M.; Vasconcelos, L.C.; Leal, L.K.A.M.; Ribeiro, J.E.; et al. Flavonoids: Biological activities and therapeutic potential. Nat. Prod. Res. 2020, 34, 692–705. [Google Scholar] [CrossRef]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial activities of flavonoids: Structure-activity relationship and mechanism. Curr. Med. Chem. 2015, 22, 132–149. [Google Scholar] [CrossRef]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure-activity relationship: An update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]
- Serafini, M.; Peluso, I.; Raguzzini, A. Flavonoids as anti-inflammatory agents. Proc. Nutr. Soc. 2010, 69, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, C.R.; Carvalho, T.T.; Manchope, M.F.; Artero, N.A.; Rasquel-Oliveira, F.S.; Fattori, V.; Casagrande, R.; Verri, W.A., Jr. Therapeutic potential of flavonoids in pain and inflammation: Mechanisms of action, pre-clinical and clinical data, and pharmaceutical development. Molecules 2020, 25, 762. [Google Scholar] [CrossRef] [PubMed]
- Calis, Z.; Mogulkoc, R.; Baltaci, A.K. The roles of flavonols/flavonoids in neurodegeneration and neuroinflammation. Mini Rev. Med. Chem. 2020, 20, 1475–1488. [Google Scholar] [CrossRef]
- Zakaryan, H.; Arabyan, E.; Oo, A.; Zandi, K. Flavonoids: Promising natural compounds against viral infections. Arch. Virol. 2017, 162, 2539–2551. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as anticancer agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef]
- Hazafa, A.; Rehman, K.U.; Jahan, N.; Jabeen, Z. The role of polyphenol (flavonoids) compounds in the treatment of cancer cells. Nutr. Cancer 2020, 72, 386–397. [Google Scholar] [CrossRef]
- Fan, H.R.; Du, W.F.; Zhu, T.; Wu, Y.J.; Liu, Y.M.; Wang, Q.; Wang, Q.; Gu, X.; Shan, X.; Denge, S. Quercetin reduces cortical GABAergic transmission and alleviates MK-801-induced hyperactivity. EBioMedicine 2018, 34, 201–213. [Google Scholar] [CrossRef]
- Calero, C.I.; González, A.N.B.; Gasulla, J.; Álvarez, S.; Evelson, P.; Calvo, D.J. Quercetin antagonism of GABAAρ₁ receptors is prevented by ascorbic acid through a redox-independent mechanism. Eur. J. Pharmacol. 2013, 714, 274–280. [Google Scholar] [CrossRef]
- Moghbelinejad, S.; Alizadeh, S.; Mohammadi, G.; Khodabandehloo, F.; Rashvand, Z.; Najafipour, R.; Nassiri-Asl, M. The effects of quercetin on the gene expression of the GABAA receptor α5 subunit gene in a mouse model of kainic acid-induced seizure. J. Physiol. Sci. 2017, 67, 339–343. [Google Scholar] [CrossRef]
- Aguirre-Hernández, E.; González-Trujano, M.E.; Terrazas, T.; Herrera-Santoyo, J.; Guevara-Fefer, P. Anxiolytic and sedative-like effects of flavonoids from Tilia americana var. mexicana: GABAergic and serotonergic participation. Salud Ment. 2016, 39, 37–46. [Google Scholar]
- Zhao, X.; Li, X.L.; Liu, X.; Wang, C.; Zhou, D.S.; Ma, Q.; Zhou, W.H.; Hu, Z.Y. Antinociceptive effects of fisetin against diabetic neuropathic pain in mice: Engagement of antioxidant mechanisms and spinal GABAA receptors. Pharmacol. Res. 2015, 102, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Ma, Z.G.; Rowlands, D.K.; Gou, Y.L.; Fok, K.L.; Wong, H.Y.; Yu, M.K.; Tsang, L.L.; Mu, L.; Chen, L.; et al. Flavonoid myricetin modulates GABAA receptor activity through activation of Ca2+ channels and CaMK-II pathway. Evid. Based Complement. Alternat. Med. 2012, 2012, 758097. [Google Scholar] [CrossRef] [PubMed]
- Karim, N.; Irshad, S.; Khan, I.; Mohammad, A.; Anis, I.; Shah, M.R.; Khan, I.; Chebib, M. GABAA receptor modulation and neuropharmacological activities of viscosine isolated from Dodonaea viscosa (Linn). Pharmacol. Biochem. Behav. 2015, 136, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Hernández-León, A.; González-Trujano, M.E.; Fernández-Guasti, A. The anxiolytic-like effect of rutin in rats involves GABAA receptors in the basolateral amygdala. Behav. Pharmacol. 2017, 28, 303–312. [Google Scholar] [CrossRef]
- Kim, J.W.; Kim, C.S.; Hu, Z.; Han, J.Y.; Kim, S.K.; Yoo, S.K.; Yeo, Y.M.; Chong, M.S.; Lee, K.; Hong, J.T.; et al. Enhancement of pentobarbital-induced sleep by apigenin through chloride ion channel activation. Arch. Pharm. Res. 2012, 35, 367–373. [Google Scholar] [CrossRef]
- Chang, C.Y.; Lin, T.Y.; Lu, C.W.; Wang, C.C.; Wang, Y.C.; Chou, S.S.; Wang, S.J. Apigenin, a natural flavonoid, inhibits glutamate release in the rat hippocampus. Eur. J. Pharmacol. 2015, 762, 72–81. [Google Scholar] [CrossRef]
- Hara, K.; Haranishi, Y.; Terada, T.; Takahashi, Y.; Nakamura, M.; Sata, T. Effects of intrathecal and intracerebroventricular administration of luteolin in a rat neuropathic pain model. Pharmacol. Biochem. Behav. 2014, 125, 78–84. [Google Scholar] [CrossRef]
- Shen, M.L.; Wang, C.H.; Lin, C.H.; Zhou, N.; Kao, S.T.; Wu, D.C. Luteolin attenuates airway mucus overproduction via inhibition of the GABAergic system. Sci. Rep. 2016, 6, 32756. [Google Scholar] [CrossRef]
- Shen, M.L.; Wang, C.H.; Chen, R.Y.; Zhou, N.; Kao, S.T.; Wu, D.C. Luteolin inhibits GABAA receptors in HEK cells and brain slices. Sci. Rep. 2016, 6, 27695. [Google Scholar] [CrossRef]
- De la Peña, J.B.I.; Kim, C.A.; Lee, H.L.; Yoon, S.Y.; Kim, H.J.; Hong, E.Y.; Kim, G.H.; Ryu, J.H.; Lee, Y.S.; Kim, K.M.; et al. Luteolin mediates the antidepressant-like effects of Cirsium japonicum in mice, possibly through modulation of the GABAA receptor. Arch. Pharm. Res. 2014, 37, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Landa, J.F.; Guillén-Ruiz, G.; Hernández-López, F.; Cueto-Escobedo, J.; Rivadeneyra-Domínguez, E.; Bernal-Morales, B.; Herrera-Huerta, E.V. Chrysin reduces anxiety-like behavior through actions on GABAA receptors during metestrus-diestrus in the rat. Behav. Brain Res. 2021, 397, 112952. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Landa, J.F.; Hernández-López, F.; Cueto-Escobedo, J.; Herrera-Huerta, E.V.; Rivadeneyra-Domínguez, E.; Bernal-Morales, B.; Romero-Avendaño, E. Chrysin (5,7-dihydroxyflavone) exerts anxiolytic-like effects through GABAA receptors in a surgical menopause model in rats. Biomed. Pharmacother. 2019, 109, 2387–2395. [Google Scholar] [CrossRef] [PubMed]
- German-Ponciano, L.J.; Costa, B.P.D.; Feitosa, L.M.; Campos, K.D.S.; Chaves, S.N.d.S.; Cueto-Escobedo, J.; Lima-Maximino, M.; Rodríguez-Landa, J.F.; Maximino, C. Chrysin, but not flavone backbone, decreases anxiety-like behavior in animal screens. Neurochem. Int. 2020, 140, 104850. [Google Scholar] [CrossRef]
- Cueto-Escobedo, J.; Andrade-Soto, J.; Lima-Maximino, M.; Maximino, C.; Hernández-López, F.; Rodríguez-Landa, J.F. Involvement of GABAergic system in the antidepressant-like effects of chrysin (5,7-dihydroxyflavone) in ovariectomized rats in the forced swim test: Comparison with neurosteroids. Behav. Brain Res. 2020, 386, 112590. [Google Scholar] [CrossRef]
- Yoon, S.Y.; de la Peña, I.C.; Shin, C.Y.; Son, K.H.; Lee, Y.S.; Ryu, J.H.; Cheong, J.H.; Ko, K.H. Convulsion-related activities of Scutellaria flavones are related to the 5,7-dihydroxyl structures. Eur. J. Pharmacol. 2011, 659, 155–160. [Google Scholar] [CrossRef]
- Zhang, S.Q.; Obregon, D.; Ehrhart, J.; Deng, J.; Tian, J.; Hou, H.; Giunta, B.; Sawmiller, D.; Tan, J. Baicalein reduces β-amyloid and promotes nonamyloidogenic amyloid precursor protein processing in an Alzheimer’s disease transgenic mouse model. J. Neurosci. Res. 2013, 91, 1239–1246. [Google Scholar] [CrossRef]
- Chang, H.H.; Yi, P.L.; Cheng, C.H.; Lu, C.Y.; Hsiao, Y.T.; Tsai, Y.F.; Li, C.L.; Chang, F.C. Biphasic effects of baicalin, an active constituent of Scutellaria baicalensis Georgi, in the spontaneous sleep-wake regulation. J. Ethnopharmacol. 2011, 135, 359–368. [Google Scholar] [CrossRef]
- Hui, K.M.; Wang, X.H.; Xue, H. Interaction of flavones from the roots of Scutellaria baicalensis with the benzodiazepine site. Planta Med. 2000, 66, 91–93. [Google Scholar] [CrossRef]
- Fong, S.Y.K.; Li, C.; Ho, Y.C.; Li, R.; Wang, Q.; Wong, Y.C.; Xue, H.; Zuo, Z. Brain uptake of bioactive flavones in Scutellariae radix and its relationship to anxiolytic effect in mice. Mol. Pharm. 2017, 14, 2908–2916. [Google Scholar] [CrossRef]
- Cho, S.; Park, J.H.; Pae, A.N.; Han, D.; Kim, D.; Cho, N.C.; No, K.T.; Yang, H.; Yoon, M.; Lee, C.; et al. Hypnotic effects and GABAergic mechanism of licorice (Glycyrrhiza glabra) ethanol extract and its major flavonoid constituent glabrol. Bioorg. Med. Chem. 2012, 20, 3493–3501. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, J.; Subramanian, V.; Nadipelly, J.S.; Kathirvelu, P.; Sayeli, V.; Cheriyan, B.V. Sedative-hypnotic like effect of 5-methoxyflavone in mice and investigation on possible mechanisms by in vivo and In silico methods. Biomed. Pharmacother. 2018, 108, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, J.; Subramanian, V.; Nadipelly, J.; Kathirvelu, P.; Sayeli, V.; Cheriyan, B.V. Anxiolytic-like activity of 5-methoxyflavone in mice with involvement of GABAergic and serotonergic systems—in vivo and In silico evidences. Eur. Neuropsychopharmacol. 2020, 36, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, D.; Imbrosci, B.; Mittmann, T. Modulatory effects of the novel TrkB receptor agonist 7,8-dihydroxyflavone on synaptic transmission and intrinsic neuronal excitability in mouse visual cortex In vitro. Eur. J. Pharmacol. 2013, 709, 64–71. [Google Scholar] [CrossRef]
- Karim, N.; Gavande, N.; Wellendorph, P.; Johnston, G.A.; Hanrahan, J.R.; Chebib, M. 3-Hydroxy-2’-methoxy-6-methylflavone: A potent anxiolytic with a unique selectivity profile at GABAA receptor subtypes. Biochem. Pharmacol. 2011, 82, 1971–1983. [Google Scholar] [CrossRef]
- Karim, N.; Curmi, J.; Gavande, N.; Johnston, G.A.; Hanrahan, J.R.; Tierney, M.L.; Chebib, M. 2’-Methoxy-6-methylflavone: A novel anxiolytic and sedative with subtype selective activating and modulating actions at GABAA receptors. Br. J. Pharmacol. 2012, 165, 880–896. [Google Scholar] [CrossRef]
- Shajib, M.S.; Rashid, R.B.; Ming, L.C.; Islam, S.; Sarker, M.M.R.; Nahar, L.; Sarker, S.D.; Datta, B.K.; Rashid, M.A. Polymethoxyflavones from Nicotiana plumbaginifolia (Solanaceae) exert antinociceptive and neuropharmacological effects in mice. Front. Pharmacol. 2018, 9, 85. [Google Scholar] [CrossRef]
- Liu, Z.; Lindemeyer, A.K.; Liang, J.; Wallner, M.; Shao, X.M.; Shao, Y.; Tao, Y.; Olsen, R.W. Flavonoids isolated from Tibetan medicines, binding to GABAA receptor and the anticonvulsant activity. Phytomedicine 2018, 50, 1–7. [Google Scholar] [CrossRef]
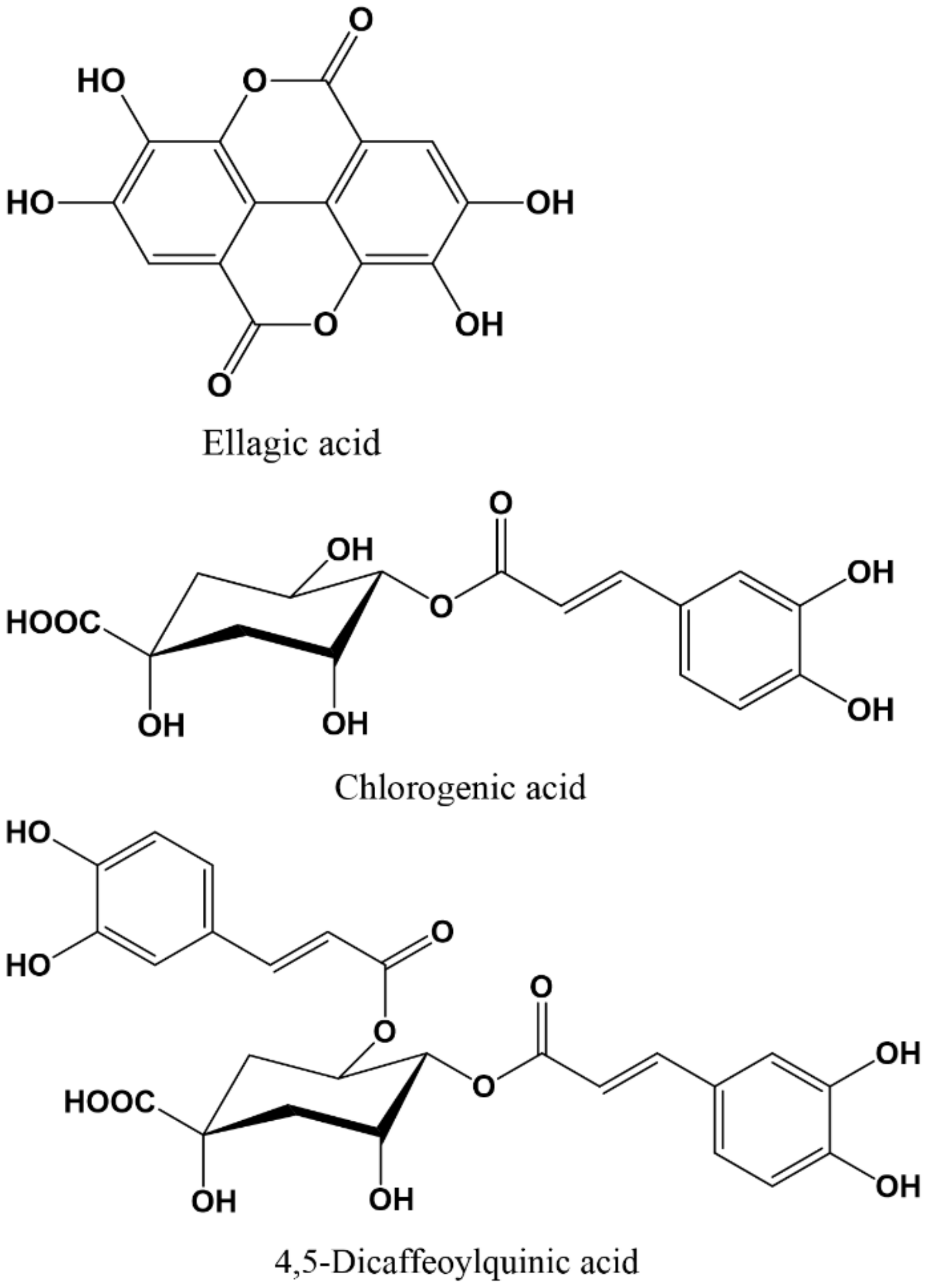
- Sheng, N.; Zheng, H.; Li, M.; Li, M.; Wang, Z.; Peng, Y.; Yu, H.; Zhang, J. 4,5-Caffeoylquinic acid and scutellarin, identified by integrated metabolomics and proteomics approach as the active ingredients of Dengzhan Shengmai, act against chronic cerebral hypoperfusion by regulating glutamatergic and GABAergic synapses. Pharmacol. Res. 2020, 152, 104636. [Google Scholar] [CrossRef]
- Dai, J.; Chen, L.; Qiu, Y.M.; Li, S.Q.; Xiong, W.H.; Yin, Y.H.; Jia, F.; Jiang, J.Y. Activations of GABAergic signaling, HSP70 and MAPK cascades are involved in baicalin's neuroprotection against gerbil global ischemia/reperfusion injury. Brain Res. Bull. 2013, 90, 1–9. [Google Scholar] [CrossRef]
- Yin, H.; Bhattarai, J.P.; Oh, S.M.; Park, S.J.; Ahn, D.K.; Han, S.K. Baicalin activates glycine and γ-aminobutyric acid receptors on substantia gelatinosa neurons of the trigeminal subsnucleus caudalis in juvenile mice. Am. J. Chin. Med. 2016, 44, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, E.; Nassiri-Asl, M.; Shafeei, M.; Sheikhi, M. Neuroprotective effects of vitexin, a flavonoid, on pentylenetetrazole-induced seizure in rats. Chem. Biol. Drug Des. 2012, 80, 274–278. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, D.R.; Todo, A.H.; Rêgo, G.M.; Cerutti, J.M.; Cavalheiro, A.J.; Rando, D.G.G.; Cerutti, S.M. Flavones-bound in benzodiazepine site on GABAA receptor: Concomitant anxiolytic-like and cognitive-enhancing effects produced by Isovitexin and 6-C-glycoside-Diosmetin. Eur. J. Pharmacol. 2018, 831, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhai, W.M.; Yang, Y.X.; Shi, J.L.; Liu, Q.T.; Liu, G.L.; Fang, N.; Li, J.; Guo, J.Y. GABA and 5-HT systems are implicated in the anxiolytic-like effect of spinosin in mice. Pharmacol. Biochem. Behav. 2015, 128, 41–49. [Google Scholar] [CrossRef]
- Anderson, W.; Barrows, M.; López, F.; Rogers, S.; Ortiz-Coffie, A.; Norman, D.; Hodges, J.; McDonald, K.; Barnes, D.; McCall, S.; et al. Investigation of the anxiolytic effects of naringenin, a component of Mentha aquatica, in the male Sprague-Dawley rat. Holist. Nurs. Pract. 2012, 26, 52–57. [Google Scholar] [CrossRef]
- Benkherouf, A.Y.; Logrén, N.; Somborac, T.; Kortesniemi, M.; Soini, S.L.; Yang, B.; Salo-Ahen, O.M.H.; Laaksonen, O.; Uusi-Oukarie, M. Hops compounds modulatory effects and 6-prenylnaringenin dual mode of action on GABAA receptors. Eur. J. Pharmacol. 2020, 873, 172962. [Google Scholar] [CrossRef]
- Benkherouf, A.Y.; Soini, S.L.; Stompor, M.; Uusi-Oukari, M. Positive allosteric modulation of native and recombinant GABAA receptors by hops prenylflavonoids. Eur. J. Pharmacol. 2019, 852, 34–41. [Google Scholar] [CrossRef]
- Shen, Y.; Lindemeyer, A.K.; González, C.; Shao, X.M.; Spigelman, I.; Olsen, R.W.; Liang, J. Dihydromyricetin as a novel anti-alcohol intoxication medication. J. Neurosci. 2012, 32, 390–401. [Google Scholar] [CrossRef]
- Liang, J.; Shen, Y.; Shao, X.M.; Scott, M.B.; Ly, E.; Wong, S.; Nguyen, A.; Tan, K.; Kwon, B.; Olsen, R.W.; et al. Dihydromyricetin prevents fetal alcohol exposure-induced behavioral and physiological deficits: The roles of GABAA receptors in adolescence. Neurochem. Res. 2014, 39, 1147–1161. [Google Scholar] [CrossRef]
- Kumar, A.; Lalitha, S.; Mishra, J. Hesperidin potentiates the neuroprotective effects of diazepam and gabapentin against pentylenetetrazole-induced convulsions in mice: Possible behavioral, biochemical and mitochondrial alterations. Indian J. Pharmacol. 2014, 46, 309–315. [Google Scholar] [CrossRef]
- Carballo-Villalobos, A.I.; González-Trujano, M.E.; Pellicer, F.; López-Muñoz, F.J. Antihyperalgesic effect of hesperidin improves with diosmin in experimental neuropathic pain. Biomed. Res. Int. 2016, 2016, 8263463. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Baburin, I.; Plitzko, I.; Hering, S.; Hamburger, M. HPLC-based activity profiling for GABAA receptor modulators from the traditional Chinese herbal drug Kushen (Sophora flavescens root). Mol. Divers. 2011, 15, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, S. Antinociceptive effect of the citrus flavonoid eriocitrin on postoperative pain conditions. J. Pain Res. 2020, 13, 805–815. [Google Scholar] [CrossRef]
- Hoffmann, K.M.; Beltrán, L.; Ziemba, P.M.; Hatt, H.; Gisselmann, G. Potentiating effect of glabridin from Glycyrrhiza glabra on GABAA receptors. Biochem. Biophys. Rep. 2016, 6, 197–202. [Google Scholar] [PubMed]
- Jin, Z.; Kim, S.; Cho, S.; Kim, I.H.; Han, D.; Jin, Y.H. Potentiating effect of glabridin on GABAA receptor-mediated responses in dorsal raphe neurons. Planta Med. 2013, 79, 1408–1412. [Google Scholar] [CrossRef]
- Cho, S.; Kim, S.; Jin, Z.; Yang, H.; Han, D.; Baek, N.I.; Jo, J.; Cho, C.W.; Park, J.H.; Shimizu, M.; et al. Isoliquiritigenin, a chalcone compound, is a positive allosteric modulator of GABAA receptors and shows hypnotic effects. Biochem Biophys Res. Commun. 2011, 413, 637–642. [Google Scholar] [CrossRef]
- Alatorre, A.; Oviedo-Chávez, A.; Villalobos, N.; Ríos, A.; Barrientos, R.; Querejeta, E. The local application of a flavonoid, (−)-epicatechin, increases the spiking of globus pallidus neurons in a dose-dependent manner and diminishes the catalepsy induced by haloperidol. Behav. Pharmacol. 2015, 26, 117–124. [Google Scholar] [CrossRef]
- Park, K.S.; Han, J.Y.; Moon, D.C.; Hong, J.T.; Oh, K.W. (−)-Epigallocatechin-3-O-gallate augments pentobarbital-induced sleeping behaviors through Cl− channel activation. J. Med. Food 2011, 14, 1456–1462. [Google Scholar] [CrossRef]
- Shrestha, S.; Park, J.H.; Lee, D.Y.; Cho, J.G.; Cho, S.; Yang, H.J.; Yong, H.I.; Yoon, M.S.; Han, D.S.; Baek, N.I.; et al. Rhus parviflora and its biflavonoid constituent, rhusflavone, induce sleep through the positive allosteric modulation of GABAA-benzodiazepine receptors. J. Ethnopharmacol. 2012, 142, 213–220. [Google Scholar] [CrossRef]
- Hansen, R.S.; Paulsen, I.; Davies, M. Determinants of amentoflavone interaction at the GABAA receptor. Eur. J. Pharmacol. 2005, 519, 199–207. [Google Scholar] [CrossRef]
- Campbell, E.L.; Chebib, M.; Johnston, G.A. The dietary flavonoids apigenin and (−)-epigallocatechin gallate enhance the positive modulation by diazepam of the activation by GABA of recombinant GABAA receptors. Biochem. Pharmacol. 2004, 68, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Deeb, O.; Shaik, B.; Agrawal, V.K. Exploring QSARs of the interaction of flavonoids with GABAA receptor using MLR, ANN and SVM techniques. J. Enzyme Inhib. Med. Chem. 2014, 29, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.S.; Mamun, A.A.; Rahman, M.A.; Kabir, M.T.; Alkahtani, S.; Alanazi, I.S.; Perveen, A.; Ashraf, G.M.; Bin-Jumah, M.N.; Abdel-Daim, M.M. Exploring the promise of flavonoids to combat neuropathic pain: From molecular mechanisms to therapeutic implications. Front. Neurosci. 2020, 14, 478. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Silva, J.; Shao, A.S.; Liang, J.; Wallner, M.; Shao, X.M.; Li, M.; Olsen, R.W. Flavonoid compounds isolated from Tibetan herbs, binding to GABAA receptor with anxiolytic property. J. Ethnopharmacol. 2021, 267, 113630. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Goutman, J.D.; Calvo, D.J. Studies on the mechanisms of action of picrotoxin, quercetin and pregnanolone at the GABA rho 1 receptor. Br. J. Pharmacol. 2004, 141, 717–727. [Google Scholar] [CrossRef]
- Calero, C.I.; Calvo, D.J. Redox modulation of homomeric rho1 GABA receptors. J. Neurochem. 2008, 105, 2367–2374. [Google Scholar] [CrossRef]
- Sun, Z.Q.; Meng, F.H.; Tu, L.X.; Sun, L. Myricetin attenuates the severity of seizures and neuroapoptosis in pentylenetetrazole kindled mice by regulating the of BDNF-TrkB signaling pathway and modulating matrix metalloproteinase-9 and GABAA. Exp. Ther. Med. 2019, 17, 3083–3091. [Google Scholar] [CrossRef]
- Carry, E.; Kshatriya, D.; Silva, J.; Davies, D.L.; Yuan, B.; Wu, Q.; Patel, H.; Park, E.R.; Gilleran, J.; Hao, L.; et al. Identification of dihydromyricetin and metabolites in serum and brain associated with acute anti-ethanol intoxicating effects in mice. Int. J. Mol. Sci. 2021, 22, 7460. [Google Scholar] [CrossRef]
- Hall, B.J.; Karim, N.; Chebib, M.; Johnston, G.A.R.; Hanrahan, J.R. Modulation of ionotropic GABA receptors by 6-methoxyflavanone and 6-methoxyflavone. Neurochem. Res. 2014, 39, 1068–1078. [Google Scholar] [CrossRef]
- Copmans, D.; Orellana-Paucar, A.M.; Steurs, G.; Zhang, Y.; Ny, A.; Foubert, K.; Exarchou, V.; Siekierska, A.; Kim, Y.; De Borggraeve, W.; et al. Methylated flavonoids as anti-seizure agents: Naringenin 4',7-dimethyl ether attenuates epileptic seizures in zebrafish and mouse models. Neurochem. Int. 2018, 112, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.K.A.; da Silva, A.W.; Moura, A.L.d.S.; Sales, K.V.B.; Marinho, E.M.; Cardoso, J.d.N.M.; Marinho, M.M.; Bandeira, P.N.; Magalhães, F.E.A.; Marinho, E.S.; et al. Chalcones reverse the anxiety and convulsive behavior of adult zebrafish. Epilepsy Behav. 2021, 117, 107881. [Google Scholar] [CrossRef] [PubMed]
- Isaza, J.H.; Torres, H.G. Preparation and chromatographic analysis of phlorotannins. J. Chromatogr. Sci. 2013, 51, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, J.; Keekan, K.K.; Anil, S.; Bhatnagar, I.; Kim, S.K. Phlorotannins. Encyclopedia Food Chem. 2019, 3, 515–527. [Google Scholar]
- Shrestha, S.; Zhang, W.; Smid, S.D. Phlorotannins: A review on biosynthesis, chemistry and bioactivity. Food Biosci. 2021, 39, 100832. [Google Scholar] [CrossRef]
- Lee, J.; Jun, M. Dual BACE1 and cholinesterase inhibitory effects of phlorotannins from Ecklonia cava—an In vitro and In silico study. Mar. Drugs 2019, 17, 91. [Google Scholar] [CrossRef]
- Kwon, S.; Jung, J.H.; Cho, S.; Moon, K.D.; Lee, J. Dieckol is a natural positive allosteric modulator of GABAA-benzodiazepine receptors and enhances inhibitory synaptic activity in cultured neurons. Nutr. Neurosci. 2019, 24, 835–842. [Google Scholar] [CrossRef]
- Yoon, M.; Kim, J.S.; Seo, S.; Lee, K.; Um, M.Y.; Lee, J.; Jung, J.; Cho, S. Dieckol, a major marine polyphenol, enhances non-rapid eye movement sleep in mice via the GABAA-benzodiazepine receptor. Front. Pharmacol. 2020, 11, 494. [Google Scholar] [CrossRef]
- Yoon, M.; Cho, S. Triphlorethol A, a dietary polyphenol from seaweed, decreases sleep latency and increases non-rapid eye movement sleep in mice. Mar. Drugs. 2018, 16, 139. [Google Scholar] [CrossRef]
- Um, M.Y.; Kim, J.Y.; Han, J.K.; Kim, J.; Yang, H.; Yoon, M.; Kim, J.; Kang, S.W.; Cho, S. Phlorotannin supplement decreases wake after sleep onset in adults with self-reported sleep disturbance: A randomized, controlled, double-blind clinical and polysomnographic study. Phytother. Res. 2018, 32, 698–704. [Google Scholar] [CrossRef]
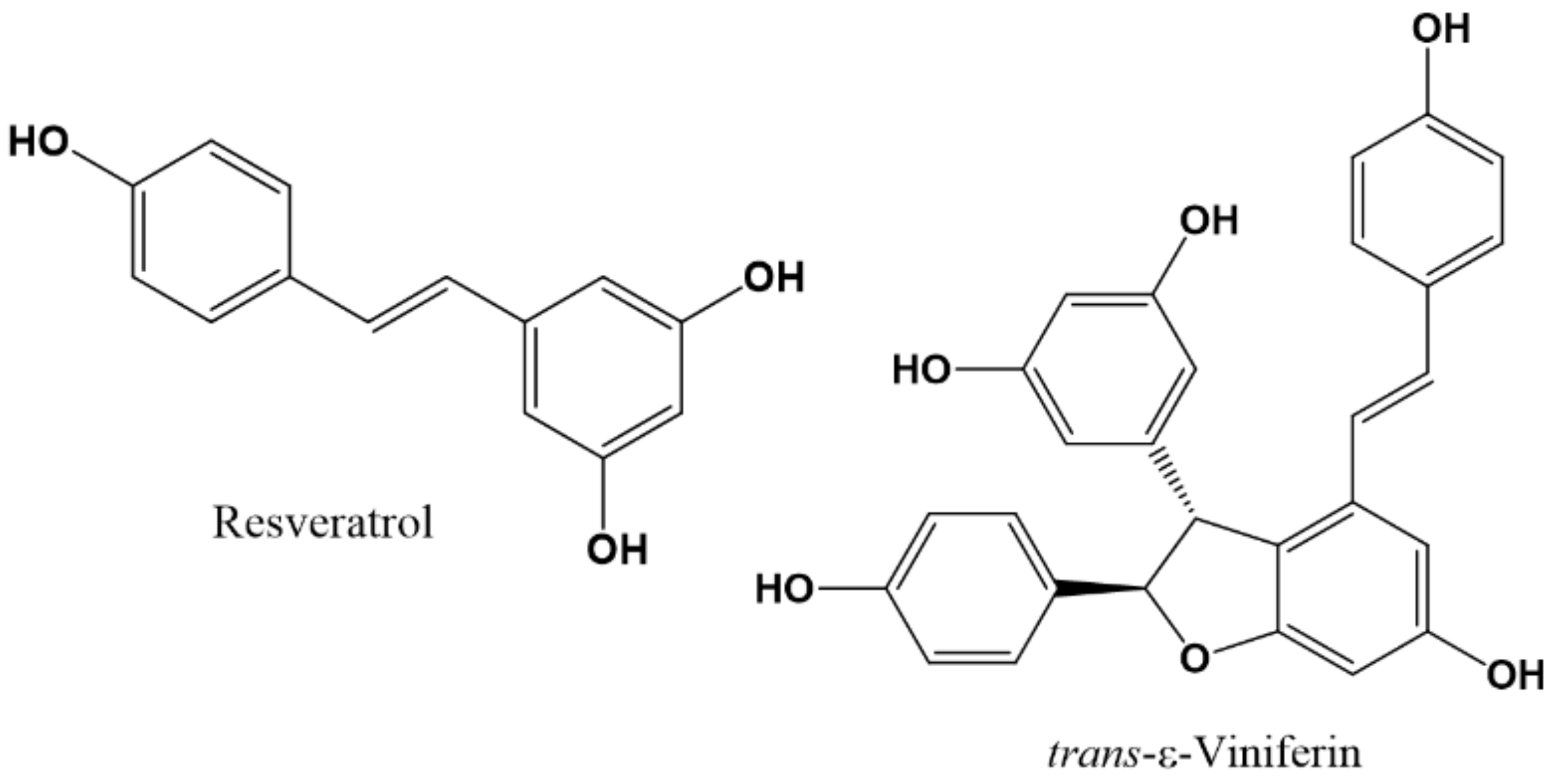
- Hamid, K.; Ng, I.; Tallapragada, V.J.; Hibbs, D.E.; Hanrahan, J.; Groundwater, P.W. The differential effects of resveratrol and trans-ε-viniferin on the GABA-induced current in GABAA receptor subtypes expressed in Xenopus laevis oocytes. J. Pharm. Pharm. Sci. 2015, 18, 328–338. [Google Scholar]
- Soh, M.S.; Lynch, J.W. Selective modulators of α5-containing GABAA receptors and their therapeutic significance. Curr. Drug Targets 2015, 16, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; You, Z.; Li, M.; Pang, L.; Cheng, J.; Wang, L. protective effect of resveratrol on the brain in a rat model of epilepsy. Neurosci. Bull. 2017, 33, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Sloviter, R.S.; Zappone, C.A.; Harvey, B.D.; Bumanglag, A.V.; Bender, R.A.; Frotscher, M. ‘‘Dormant basket cell’’ hypothesis revisited: Relative vulnerabilities of dentate gyrus mossy cells and inhibitory interneurons after hippocampal status epilepticus in the rat. J. Comp. Neurol. 2003, 459, 44–76. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Xu, Q.; Zhang, L.; Kong, D.; Ma, R.; Wang, L. Protective effect of resveratrol against kainate-induced temporal lobe epilepsy in rats. Neurochem. Res. 2009, 34, 1393–1400. [Google Scholar] [CrossRef]
- Yaşar, S.N.; Can, O.D.; Oztürk, N.; Sagratini, G.; Ricciutelli, M.; Vittori, S.; Maggi, F. Central nervous system activities of Hypericum origanifolium extract via GABAergic and opioidergic mechanisms. Phytother. Res. 2013, 27, 877–884. [Google Scholar] [CrossRef]
- Ríos, J.L.; Giner, R.M.; Marín, M.; Recio, M.C. A pharmacological update of ellagic acid. Planta Med. 2018, 84, 1068–1093. [Google Scholar] [CrossRef]
- Girish, C.; Raj, V.; Arya, J.; Balakrishnan, S. Involvement of the GABAergic system in the anxiolytic-like effect of the flavonoid ellagic acid in mice. Eur. J. Pharmacol. 2013, 710, 49–58. [Google Scholar] [CrossRef]
- Abdelhalim, A.; Chebib, M.; Aburjai, T.; Johnston, G.A.; Hanrahan, J. GABAA receptor modulation by compounds isolated from Salvia triloba L. Adv. Biol. Chem. 2014, 4, 148–159. [Google Scholar] [CrossRef]
- Abdelhalim, A.; Karim, N.; Chebib, M.; Aburjai, T.; Khan, I.; Johnston, G.A.; Hanrahan, J. Antidepressant, anxiolytic and antinociceptive activities of constituents from Rosmarinus officinalis. J. Pharm. Pharm. Sci. 2015, 18, 448–459. [Google Scholar] [CrossRef]
- Kavvadias, D.; Monschein, V.; Sand, P.; Riederer, P.; Schreier, P. Constituents of sage (Salvia officinalis) with In vitro affinity to human brain benzodiazepine receptor. Planta Med. 2003, 69, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jeon, Y.J. Efficacy and safety of a dieckol-rich extract (AG-dieckol) of brown algae, Ecklonia cava, in pre-diabetic individuals: A double-blind, randomized, placebo-controlled clinical trial. Food Funct. 2015, 6, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.; Dordevic, A.L.; Ryan, L.; Bonham, M.P. The impact of a single dose of a polyphenol-rich seaweed extract on postprandial glycaemic control in healthy adults: A randomised cross-over trial. Nutrients 2018, 10, 270. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jung, Y.; Lee, E.; Jang, S.; Ryu, D.H.; Kwon, O.; Hwang, G.S. Urinary metabolomic profiling analysis and evaluation of the effect of Ecklonia cava extract intake. Nutrients 2020, 12, 1407. [Google Scholar] [CrossRef]
- Baldrick, F.R.; McFadden, K.; Ibars, M.; Sung, C.; Moffatt, T.; Megarry, K.; Thomas, K.; Mitchell, P.; Wallace, J.M.W.; Pourshahidi, L.K.; et al. Impact of a (poly)phenol-rich extract from the brown algae Ascophyllum nodosum on DNA damage and antioxidant activity in an overweight or obese population: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 688–700. [Google Scholar] [CrossRef]
- Haskell-Ramsay, C.F.; Jackson, P.A.; Dodd, F.L.; Forster, J.S.; Bérubé, J.; Levinton, C.; Kennedy, D.O. Acute post-prandial cognitive effects of brown seaweed extract in humans. Nutrients 2018, 10, 85. [Google Scholar] [CrossRef]
- Kyselova, Z. Toxicological aspects of the use of phenolic compounds in disease prevention. Interdiscip. Toxicol. 2011, 4, 173–183. [Google Scholar] [CrossRef]
- Perveen, S.; Al-Taweel, A.M. Phenolic compounds from the natural sources and their cytotoxicity. In Phenolic Compounds; Soto-Hernández, M., Palma-Tenango, M., Garcia-Mateos, M.R., Eds.; IntechOpen Book Series: London, UK, 2017. [Google Scholar]
- Anku, W.W.; Mamo, M.A.; Govender, P.P. Phenolic compounds in water: Sources, reactivity, toxicity and treatment methods. In Phenolic Compounds; Soto-Hernández, M., Palma-Tenango, M., García-Mateos, M.R., Eds.; IntechOpen Book Series: London, UK, 2017. [Google Scholar]
- Thavasi, V.; Leong, L.P.; Bettens, R.P. Investigation of the influence of hydroxy groups on the radical scavenging ability of polyphenols. J. Phys. Chem. A 2006, 110, 4918–4923. [Google Scholar] [CrossRef]
- Kim, A.R.; Shin, T.S.; Lee, M.S.; Park, J.Y.; Park, K.E.; Yoon, N.Y.; Kim, J.S.; Choi, J.S.; Jang, B.C.; Byun, D.S.; et al. Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and anti-inflammatory properties. J. Agric. Food. Chem. 2009, 57, 3483–3489. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 Flavonol | ||
|---|---|---|
| Compound | Properties | Reference |
| Quercetin (5,7,3′,4′-tetrahydroxy) | Quercetin acts as a negative allosteric GABAAR modulator with antipsychotic activity. These results justify further therapeutic development of the excitatory-inhibitory imbalance disorders. | [69] |
| Quercetin antagonistic actions on GABAAρ₁Rs are mediated through a redox-independent allosteric mechanism. | [70] | |
| GABAAα5R could be a mechanism for reducing seizure severity (at anticonvulsive doses) or even be used a marker of seizure severity. | [71] | |
| Quercetin and its glycosides (rutin and isoquercitrin) are partially responsible for the anxiolytic and sedative-like effect of Tilia americana var. mexicana through the GABA/BZD and serotoninergic 5-HT1A receptors. | [72] | |
| Fisetin (7,3′,4′-trihydroxy) | Treatment with fisentin can delay or correct neuropathic hyperalgesia and allodynia in mice with type 1 diabetes. The analgesia caused by fisetin may be linked with its antioxidant activity. Spinal GABAARs are likely rendered as downstream targets. | [73] |
| Myricetin (3,5,7,3′,4′,5′-pentahydroxy) | Myricetin enhances GABAAR activity via the calcium channel/Ca2+/calmodulin-dependent protein kinase II dependent mechanism, which is distinctively different from that of most existing BZD-binding site agonists of GABAAR. | [74] |
| Viscosine (5,7,4′-trihydroxy-3,6-dimethoxy) | The anxiolytic and anticonvulsant actions of viscosine are likely mediated via its positive allosteric modulatory action at different GABAAR subtypes. | [75] |
| Glycosides | ||
| Rutin (quercetin 3-O-rutinoside) | The anxiolytic-like effect involves GABAergic neurotransmission without implication of BZD receptors. | [76] |
| Rutin (quercetin 3-O-rutinoside) Isoquercitrin (quercetin-3-O-glucoside) | Leaves of Tilia americana var. mexicana have anxiolytic and sedative-like effects and its flavonoids, quercetin, rutin and isoquercitrin, are partially responsible due to the involvement of GABA/BZD and serotoninergic 5-HT1A receptors. | [72] |
 Flavone | ||
|---|---|---|
| Compound | Properties | Reference |
| Apigenin (5,7,4’-trihydroxy) | Apigenin enhances pentobarbital-induced sleep behaviours through chloride ion channel activation. | [77] |
| Apigenin reduces the frequency of spontaneous excitatory postsynaptic currents without affecting their amplitude, suggesting a presynaptic mechanism. | [78] | |
| Luteolin (5,7,3’,4’-trihydroxy) | Luteolin ameliorates mechanical and cold hyperalgesia at least in part by activation of GABAARs in a flumazenil-insensitive manner and μ-opioid receptors in the spinal cord. | [79] |
| Luteolin attenuates mucus overproduction and goblet cell hyperplasia in an animal asthma model at least partially by inhibition of GABAAR activities. | [80] | |
| Luteolin has negative modulatory effects on both recombinant and endogenous GABAARs and inhibits phasic rather than tonic inhibition in the hippocampus. | [81] | |
| The antidepressant-like effect of Cirsium japonicum could be mediated by luteolin through the potentiation of the GABAA receptor-Cl− ion channel complex. | [82] | |
| Chrysin (5,7-dihydroxy) | Chrysin prevents anxiety-like behaviour during metestrus-diestrus in two unconditioned models. These effects were mediated by actions on GABAARs. | [83] |
| Chrysin produces anxiolytic-like effects through actions on GABAARs in a model of surgical menopause in rats. | [84] | |
| Chrysin has more than one mechanism of action in addition to its action at the GABAA-BZD receptor complex, and also could be involved in its free radical scavenging abilities. | [85] | |
| The acute antidepressant-like effects of chrysin, similar to neurosteroids, are mediated by the GABA-binding site at GABAARs in ovariectomized rats. | [86] | |
| Baicalein (5,6,7-trihydroxy) | The anticonvulsive effect of baicalein was mediated by the BZD binding site of GABAAR. The 5,7-dihydroxyl group is present in the structure of the three flavones, playing a key role in inducing convulsion-related activities. | [87] |
| Baicalein promotes non-amyloidogenic processing of APP, thereby reducing Aβ production and improving cognitive performance by activation of GABAARs. | [88] | |
| It exhibits biphasic effects on sleep–wake regulation; decreases the SW sleep during the light period and increases SW sleep and REM sleep during the dark period. | [89] | |
| Wogonin (5,7-dihydroxy-8-methoxy) | Wogonin is a 2.8-fold stronger ligand to the BZD binding site (Ki = 2.03 μM) compared to baicalein (Ki = 5.69 μM). | [90] |
| Oroxylin A (5,7-dihydroxy-6-methoxy) | In vitro studies reveal that oroxylin A blocked muscimol-induced intracellular Cl− influx. | [87] |
| Oroxylin A has the highest brain uptake and the highest affinity to brain tissues (In vitro tissue binding assay) compared to other flavones. This flavone, a GABAA antagonist, can suppress the anxiolytic effects of other flavones present in the extract. | [91] | |
| Glabrol (7,4’-dihydroxy-8,3’-di-isoprenyl) | Glabrol inhibits [3H]-flumazenil binding site to the GABAA-BZD receptors in the rat cerebral cortex membrane with a binding affinity (Ki) of 1.63 μM. The isoprenyl groups may play a key role in binding to GABAA-BZD receptors. Glabrol increases sleep duration and decreases sleep latency via a positive allosteric modulation of GABAA-BZD receptors. | [92] |
| 5-Methoxyflavone | In silico studies indicate that 5-methoxyflavone exhibits good binding affinity towards GABAA, adenosine, glycine and NMDA receptors by H-bond interactions, justifying its hypnotic effect. | [93] |
| In silico studies demonstrate a good binding affinity of 5-methoxyflavone towards GABAA (α2 subunit-containing) and serotoninergic 5-HT1A receptors by H-bond interactions. | [94] | |
| 7,8-Dihydroxyflavone | 7,8-Dihydroxyflavone causes a selective reduction in the strength of GABAergic inhibition after incubation with acute cortical slices. | [95] |
| 3-Hydroxy-2’methoxy-6-methylflavone | 3-Hydroxy-2’methoxy-6-methylflavone has an anxiolytic effect without sedation or myorelaxation through positive allosteric modulation of the α2β2/3γδ2L and direct activation of α4β2/3δ GABAAR subtypes. | [96] |
| 2’-Methoxy-6-methylflavone | 2’-Methoxy-6-methylflavone could be used as a tool to study the complex nature of the activation and modulation of GABAAR subtypes. | [97] |
| 3,5,6,7,8,3’-hexamethoxy-4’,5’-methylenedioxyflavone | This methylenedioxyflavone shows anxiolytic-like activity in the EPM but locomotor responses remain unchanged. | [98] |
| 6,7,4’,5’-dimethylenedioxy-3,5,3’-trimethoxyflavone | This methoxyflavone has anxiolytic-like activity in the EPM test involving GABAAR reversed by flumazenil. | [98] |
| 3,3’,4’,5,5’,8-hexamethoxy-6,7-methylenedioxyflavone | This methylenedioxyflavone shows anxiolytic-like activity in the EPM test, with the implication of GABAAR, but locomotor responses remain unchanged. | [98] |
| 6-Methoxyflavone/8-Methoxyflavone 5,7,2’,4’-Tetrahydroxy-6,5’-dimethoxyflavone | The structure-activity relationships analysis of 28 flavonoids indicate that 6-and/or 8-methoxy flavones had the most potent binding affinity to GABAARs. Of them, compound 5,7,2’,4’-tetrahydroxy-6,5’-dimethoxyflavone (IC50 0.10 μM) had the higher anticonvulsant activity against chemically-induced and electrogenic seizures without myorelaxation and sedation. | [99] |
| Glycosides | ||
| Scutellarin (scutellarein-7-O-glucuronide) | Scutellarin is identified by integrated metabolomics and proteomics approach as the active ingredient of Dengzhan Shengmai acting against chronic cerebral hypoperfusion due to the regulation of glutamatergic and GABAergic synapses. | [100] |
| Baicalin (baicalein 7-O-glucuronide) | Baicalin does not change intracellular Cl− concentration, whereas its aglycone does. Glycosylation has a negative influence on the affinity for the BZD-binding site of the GABAAR. | [87] |
| Baicalin activates GABAergic signalling, HSP70 and MAPKs cascades in global ischemia, which may be a mechanism underlying the baicalin’s neuroprotection. | [101] | |
| Baicalin inhibits SG neurons activating the BZD-sensitive GABAAR and/or glycine receptors, becoming a potential target for orofacial pain modulation. | [102] | |
| Vitexin (apigenin 8-C-glucoside) | Vitexin has anticonvulsant effects possibly through interaction at the BZD-binding site of the GABAAR complex. | [103] |
| Isovitexin (apigenin 6-C-glucoside) | Isovitexin could exert its memory-enhancing and anxiolytic-like effects via GABAA R modulation through its BZD-binding site. | [104] |
| Spinosin (apigenin 7-O-methyl-6-O-diglucoside) | Spinosin exerts anxiolytic-like effects with a mechanism of action modulated by GABAA and serotoninergic 5-HT1A receptors. | [105] |
 Flavanone | ||
|---|---|---|
| Compound | Properties | Reference |
| Naringenin (5,7,4’-O-trihydroxy) | Naringenin does not produce anxiolysis by modulation of the GABAARs but it does decrease motor movements. | [106] |
| 6-Prenylnaringenin | 6-Prenylnaringenin acts as a positive allosteric modulator at α+ β− binding interface as well as a null modulator at the flumazenil-sensitive α+ γ2− binding interface on GABAARs. | [107] |
| 8-Prenylnaringenin | 8-Prenylnaringenin potentiated GABA-induced displacement of [3H] ethynylbicycloorthobenzoate radioligand binding assay in native and recombinant α1β3γ2, α2β3γ2 and α6β3δ receptors (HEK293 cells, IC50 of 7.3 µM). | [108] |
| Dihydromyricetin = ampelopsin (5,7,3’,4’,5’-O-tetrahydroxy) | Dihydromyricetin competitively inhibits the BZD-site [3H]-flunitrazepam binding site (IC50 4.36 μM), suggesting the interaction with the BZD sites on GABAAR. | [109] |
| Dihydromyricetin prevents foetal alcohol exposure consequences in pregnant rats, avoiding the alterations in physiology, behaviour, and hippocampal GABAAR function. | [110] | |
| Hesperidin (hesperetin 7-O-rutinoside) | Hesperidin possesses anticonvulsant activity which might be mediated through the modulation of GABA-BZD receptor action. | [111] |
| The antihyperalgesic effect of hesperidin combined with diosmin involves central activity partially modulated by D2, GABAA, and opioid receptors, but not serotoninergic 5-HT1A receptors. | [112] | |
| Kushenol I (3β,7,2’,4’-tetrahydroxy-5-methoxy-8-lavandulyl) (–)-Kurarinone (7,2’,4’-trihydroxy-5-methoxy-8-lavandulyl) Sophoraflavanone G (5,7,2’,4’-tetrahydroxy-8-lavandulyl) | These flavanones act as GABAAR modulators. They induced the IGABA enhancement in Xenopus oocytes transiently expressing GABAAR with subunit composition: α1β2γ2S. EC50 are 5.0, 8.1 and 15.0 μM, respectively. | [113] |
| Isoxanthohumol (7,4’-dihidroxy-5-methoxy-8-isoprenyl) | Isoxanthohumol potentiates GABA-induced displacement of [3H]-ethynylbicycloorthobenzoate radioligand binding assay in native and recombinant α1β3γ2, α2β3γ2 and α6β3δ receptors (HEK293 cells) in a concentration-dependent manner with an IC50 of 11.6 µM. | [108] |
| Glycosides | ||
| Eriocitrin (eriodictyol 7-O-rutinoside) (eriodictyol is 5,7,3’,4’-tetrahydroxy flavanone) | The antinociceptive effect of eriocitrin is blocked by naltrexone (opioid receptor antagonist) and bicuculline (GABAAR antagonist). Therefore, the antinociception in postoperative pain conditions could be mediated through opioid and GABAARs. | [114] |
 |  |  |
|---|---|---|
| Isoflavan | Chalcone | (−)-Epicatechin |
| Compound | Properties | Reference |
| Isoflavanes | ||
| Glabridin: (3R)-isoflavan-2′,4′-diol-7,8-dimethyl-6”-pyrano | Glabridin acts on GABAAR β subunit by a mechanism involving the M286 residue, a key amino acid at the binding site for general anaesthetics. | [115] |
| Glabridin may exhibit sedative and hypnotic effects by GABAAR actions which potentiates GABAergic inhibition in dorsal raphe neurons. Flumazenil did not inhibit glabridin-induced potentiation. | [116] | |
| Chalcones | ||
| Isoliquiritigenin (4,2′,4′-trihihidroxy) | Isoliquiritigenin has higher affinity for GABAA-BZD receptors than diazepam. Its effect on GABA currents was blocked by flumazenil and ZK-93426). Therefore, isoliquiritigenin produces hypnotic effects by positive allosteric modulation of GABAA-BZD receptors. | [117] |
| Kuraridine (2,4,4′,6′-tetrahydroxy-2′-methoxy-5′-lavandulyl) | Kuraridine acts as a GABAAR modulator. It induces IGABA enhancement in Xenopus oocytes transiently expressing GABAARs with subunit composition: α1β2γ2S (EC50 4.0 μM). | [113] |
| Xanthohumol (4,2′,4′-trihidroxy-6′-methoxy-3′-isoprenyl) | Xanthohumol potentiates GABA-induced displacement of [3H] the ethynylbicycloorthobenzoate radioligand binding assay in native and recombinant α1β3γ2, α2β3γ2 and α6β3δ receptors (HEK293 cells, IC50 of 29.7 µM) | [108] |
| Catechins | ||
| (−)-Epicatechin (5,7,3′,4′-tetrahydroxy) | Epicatechin increases the basal firing rate of neurons in the globus pallidus and antagonizes the inhibitory effect of GABA. A bilateral infusion into the globus pallidus diminishes the catalepsy induced by haloperidol. | [118] |
| (−)-Epigallocatechin-3-O-gallate | Epigallocatechin gallate increases Cl− influx in primary cultured cerebellar cells and decreases GABAARs α-subunit expression, whereas it has no effect on the expression of β- and γ-subunits. | [119] |
| Biflavonoids | ||
|---|---|---|
 Mesuaferrone B |  Rhusflavone |  Agathisflavone |
| Mesuaferrone B, rhusflavone, and agathisflavone competitively inhibit the ³H-Ro 15-1788 flumazenil binding site with a Ki of 0.280, 0.045, and 0.091 μM, respectively. In addition, rhusflavone has sedative-hypnotic effects and is the most potent at inducing sleep. The decrease in sleep latency and increase in sleep duration seems to be due to the presence of a conjugated ketone and C6-C8” linkage in rhusflavone. It induces sleep via the positive allosteric modulation of GABAA-BZD receptors. | [120] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ríos, J.-L.; Schinella, G.R.; Moragrega, I. Phenolics as GABAA Receptor Ligands: An Updated Review. Molecules 2022, 27, 1770. https://doi.org/10.3390/molecules27061770
Ríos J-L, Schinella GR, Moragrega I. Phenolics as GABAA Receptor Ligands: An Updated Review. Molecules. 2022; 27(6):1770. https://doi.org/10.3390/molecules27061770
Chicago/Turabian StyleRíos, José-Luis, Guillermo R. Schinella, and Inés Moragrega. 2022. "Phenolics as GABAA Receptor Ligands: An Updated Review" Molecules 27, no. 6: 1770. https://doi.org/10.3390/molecules27061770
APA StyleRíos, J.-L., Schinella, G. R., & Moragrega, I. (2022). Phenolics as GABAA Receptor Ligands: An Updated Review. Molecules, 27(6), 1770. https://doi.org/10.3390/molecules27061770