
Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone from Syzygium nervosum Seeds on Antiproliferative, DNA Damage, Cell Cycle Arrest, and Apoptosis in Human Cervical Cancer Cell Lines

 , , ,
, , ,  , and
, and
Abstract
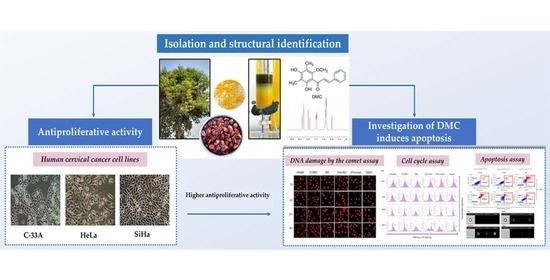
:
1. Introduction
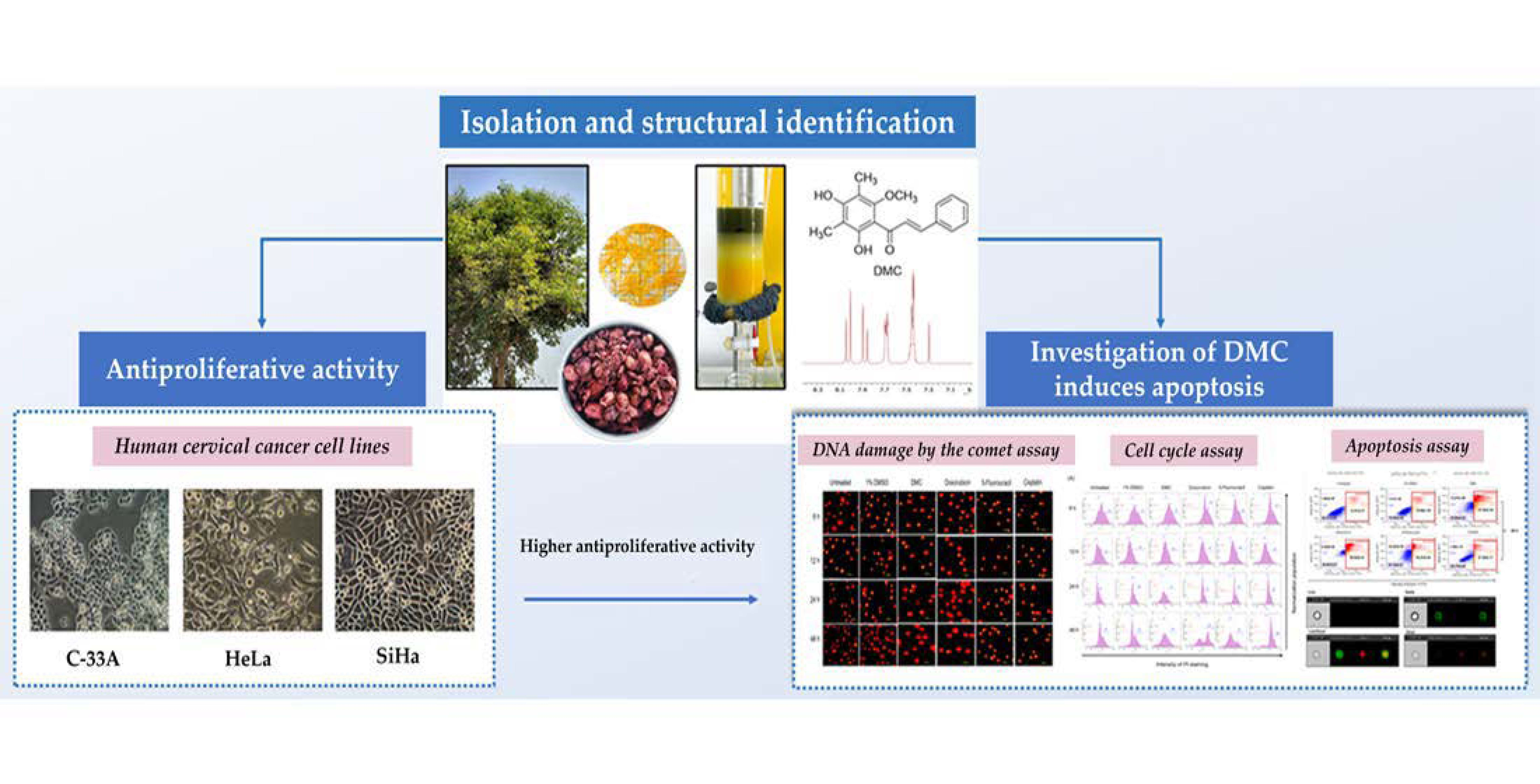
2. Results
2.1. Isolation and Structural Identification of DMC
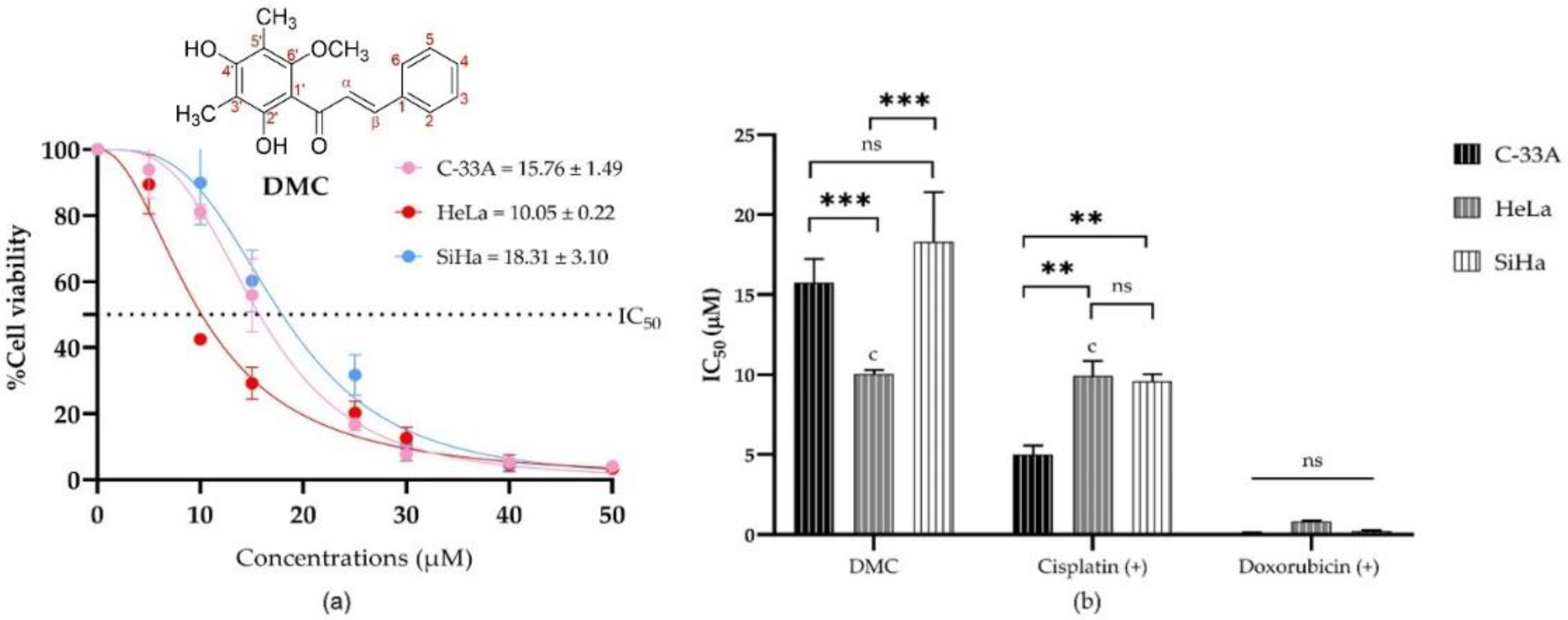
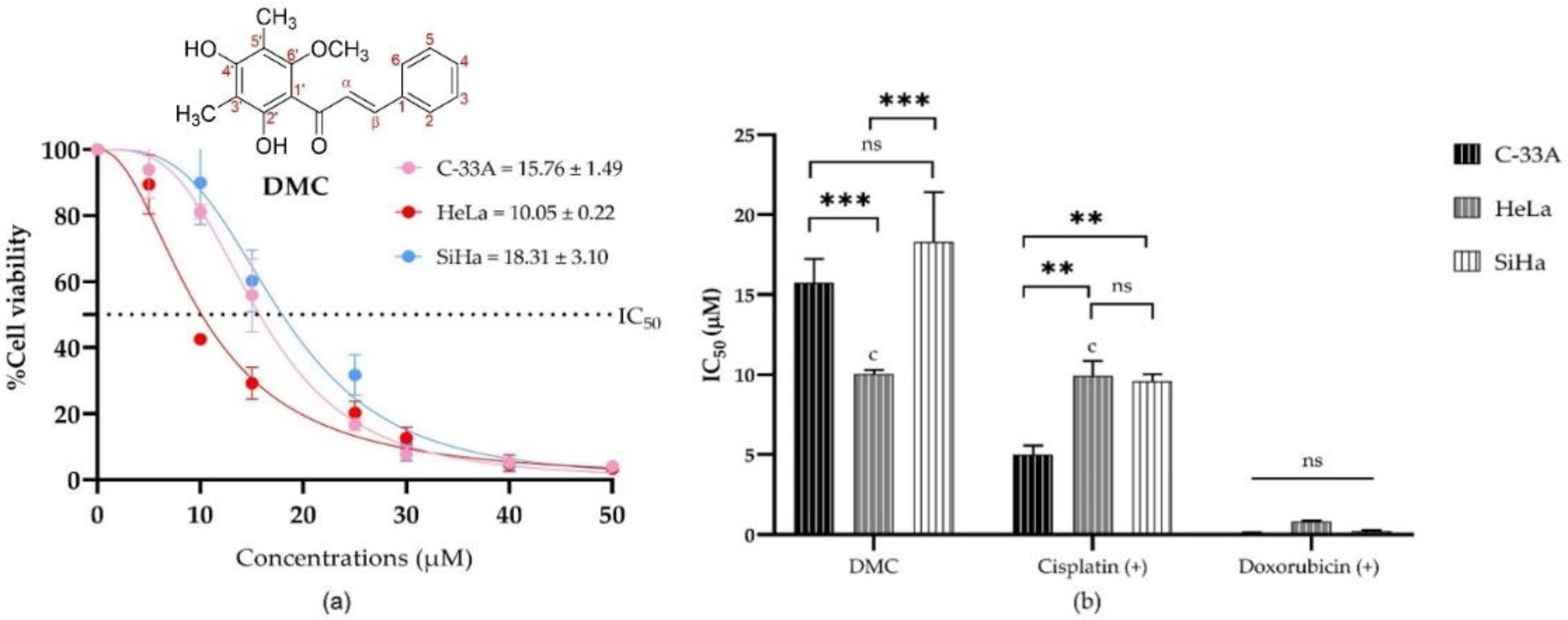
2.2. Antiproliferative Activity of DMC on Cervical Cancer Cell Lines
2.3. Investigation of Compound-Induced Apoptosis
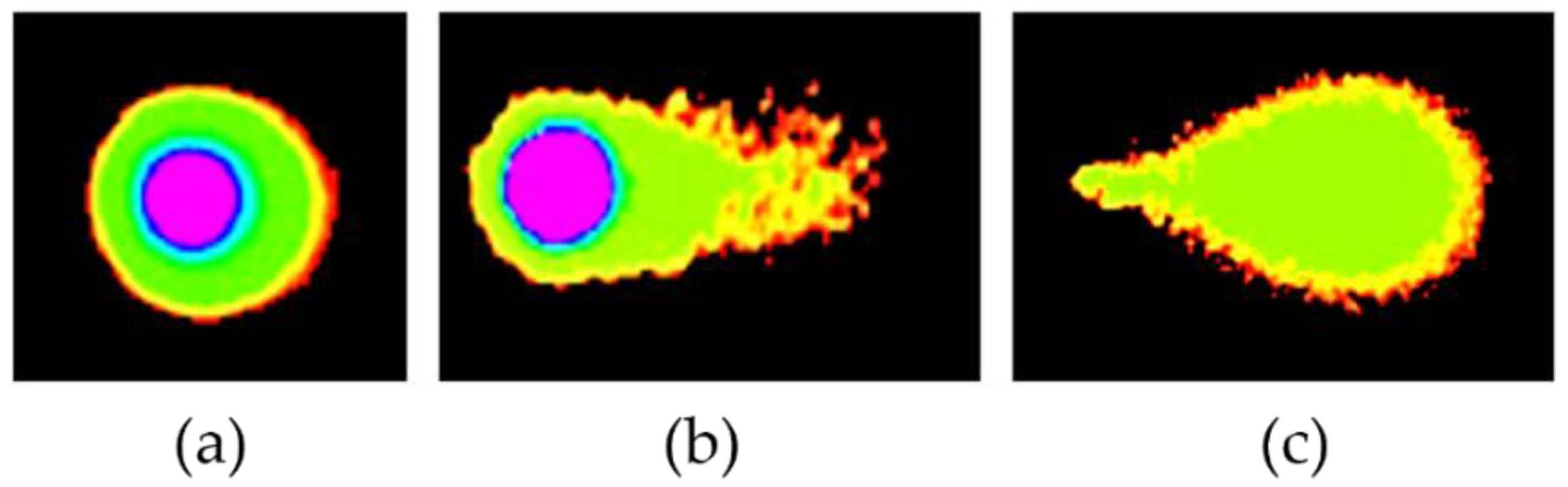
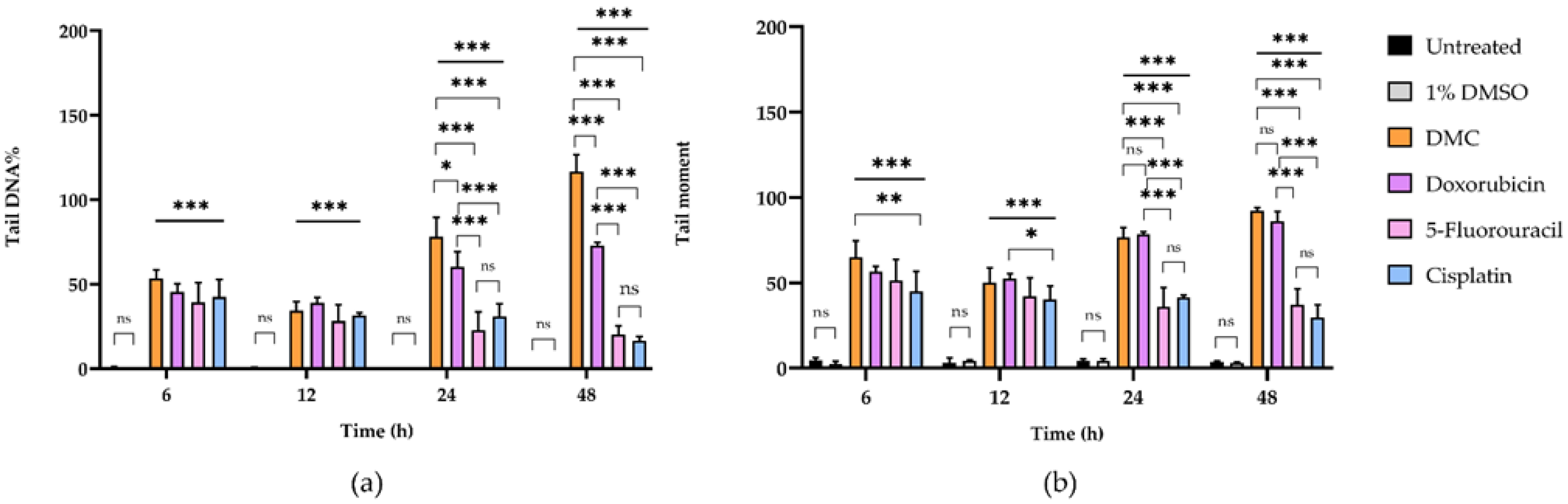
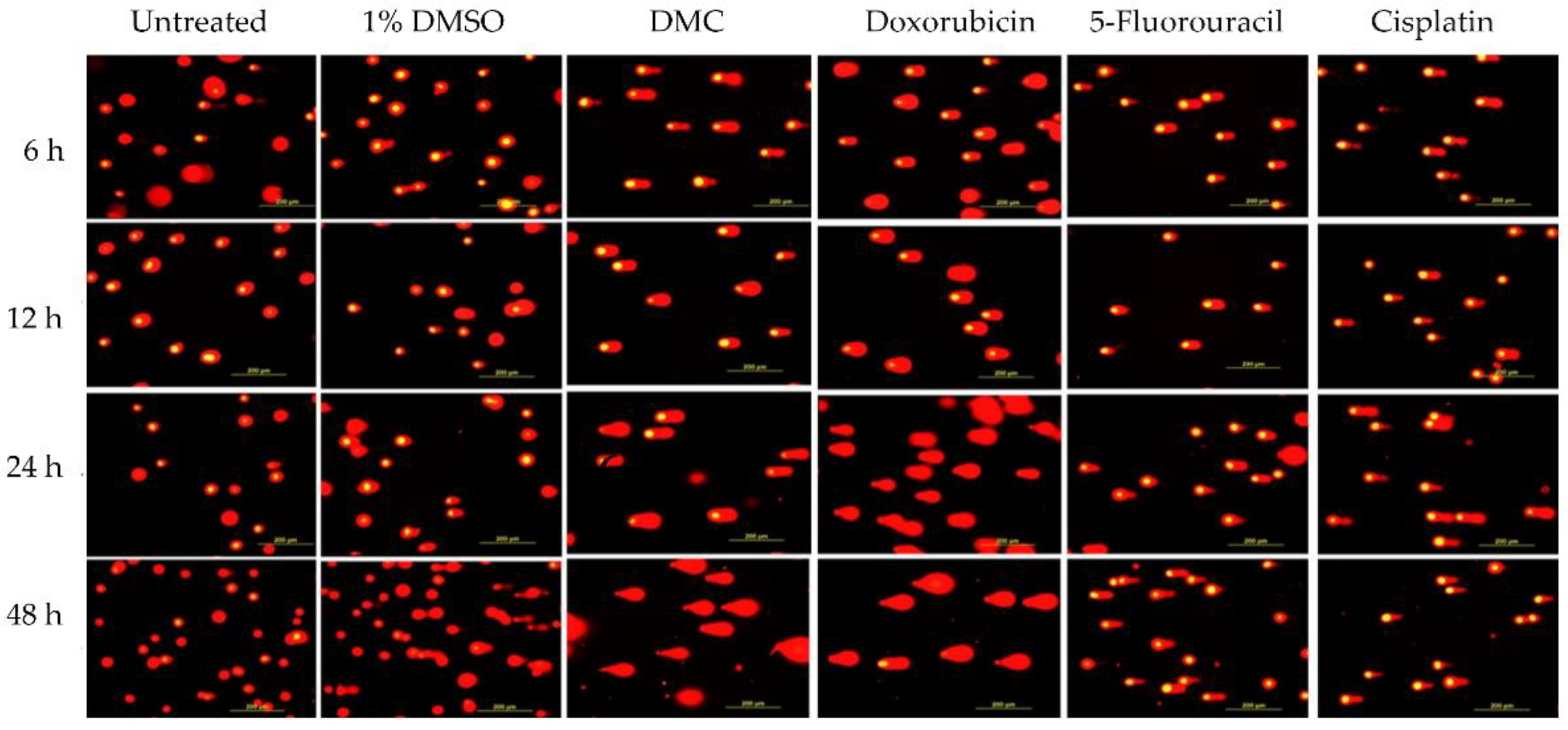
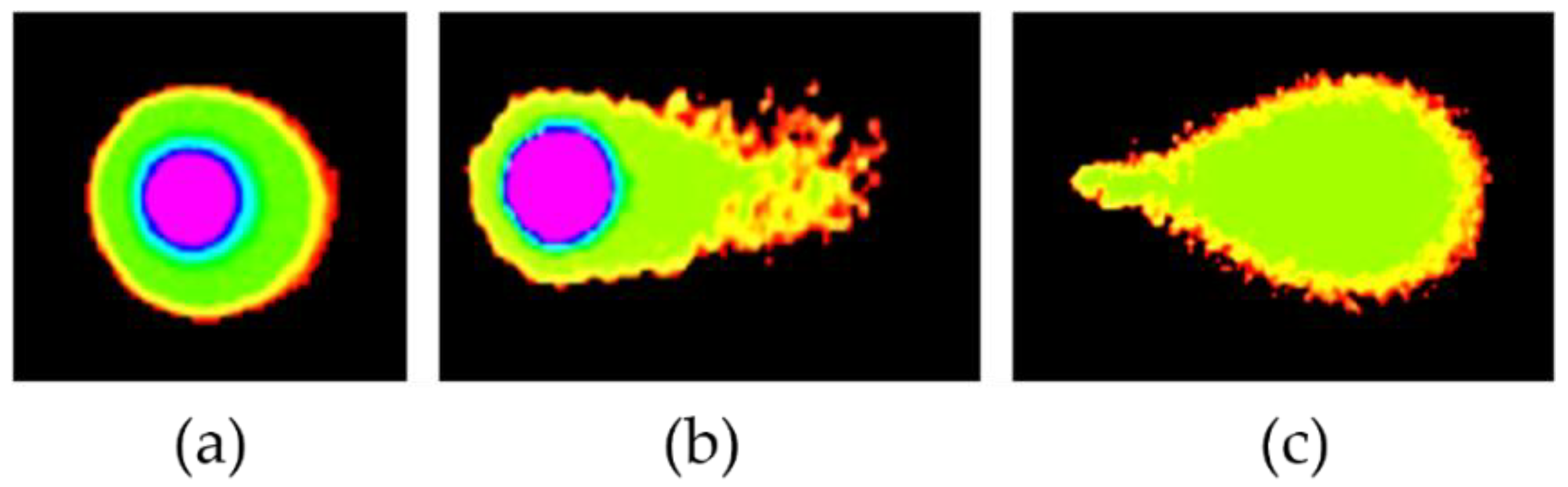
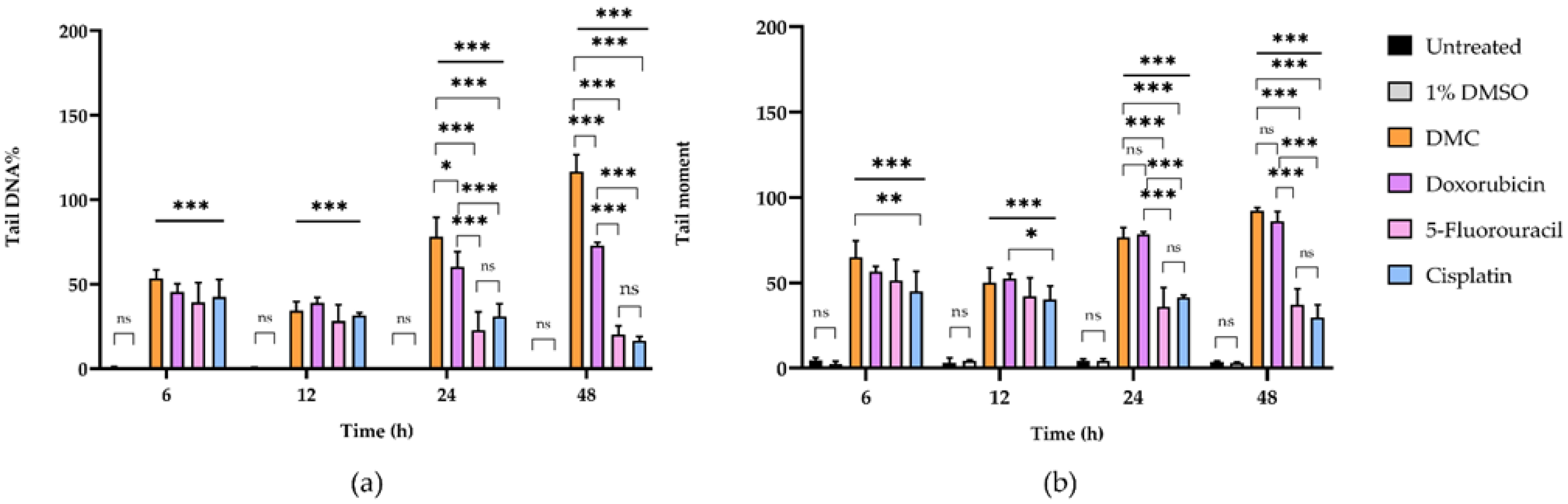
2.3.1. DMC-Induced DNA Damage Pathways
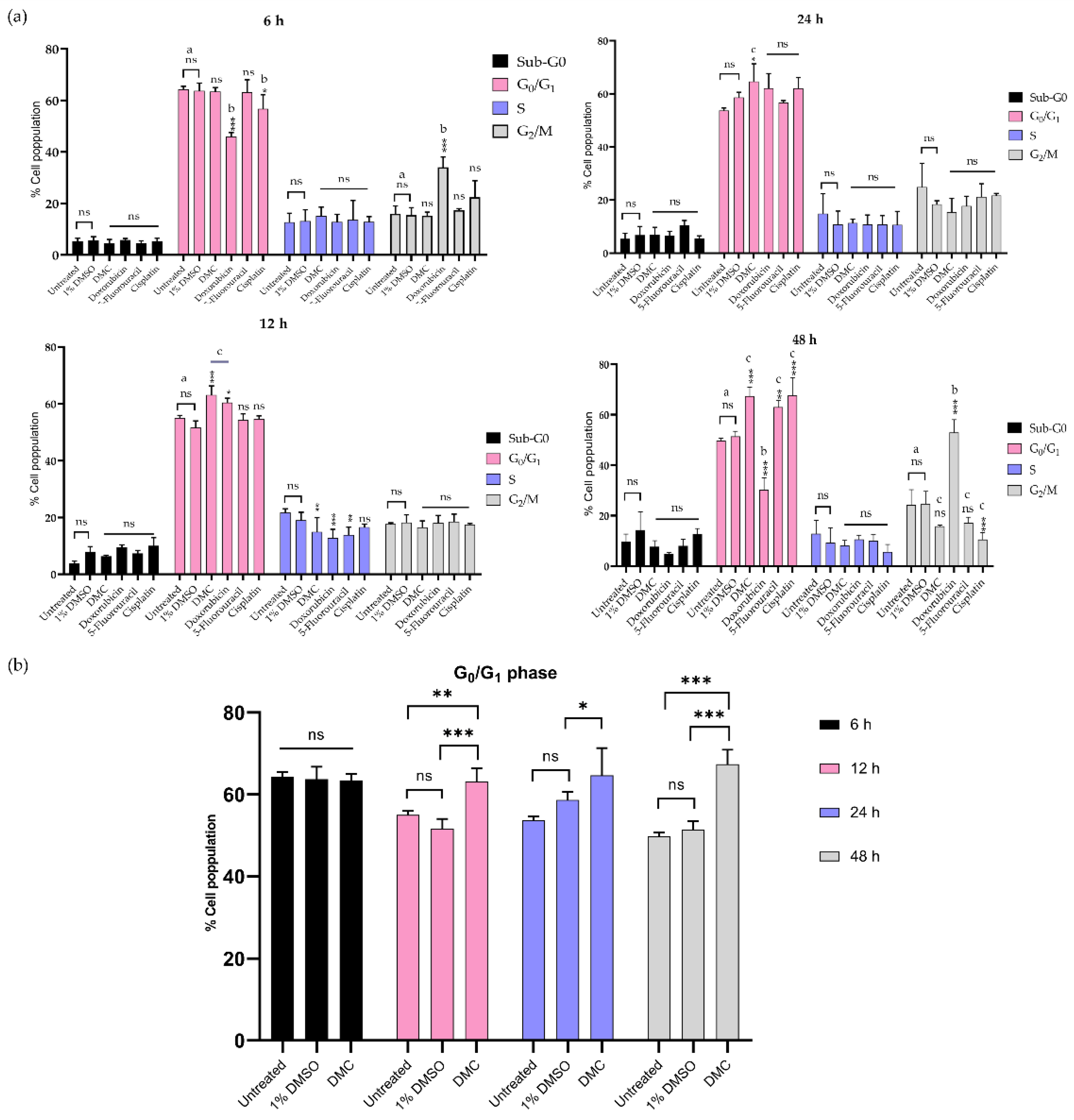
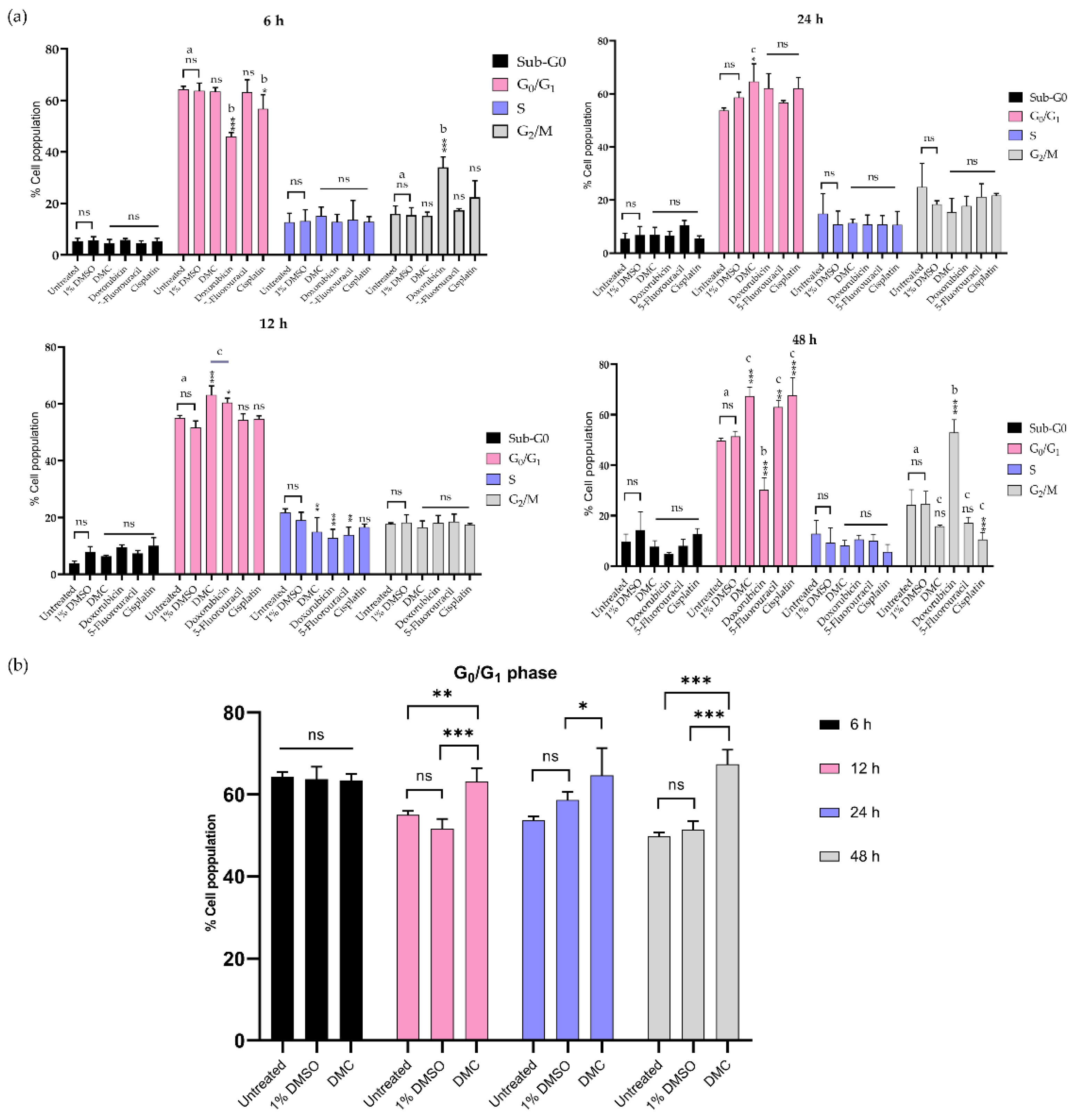
2.3.2. Inhibition of the G0/G1 and G2/M Phases
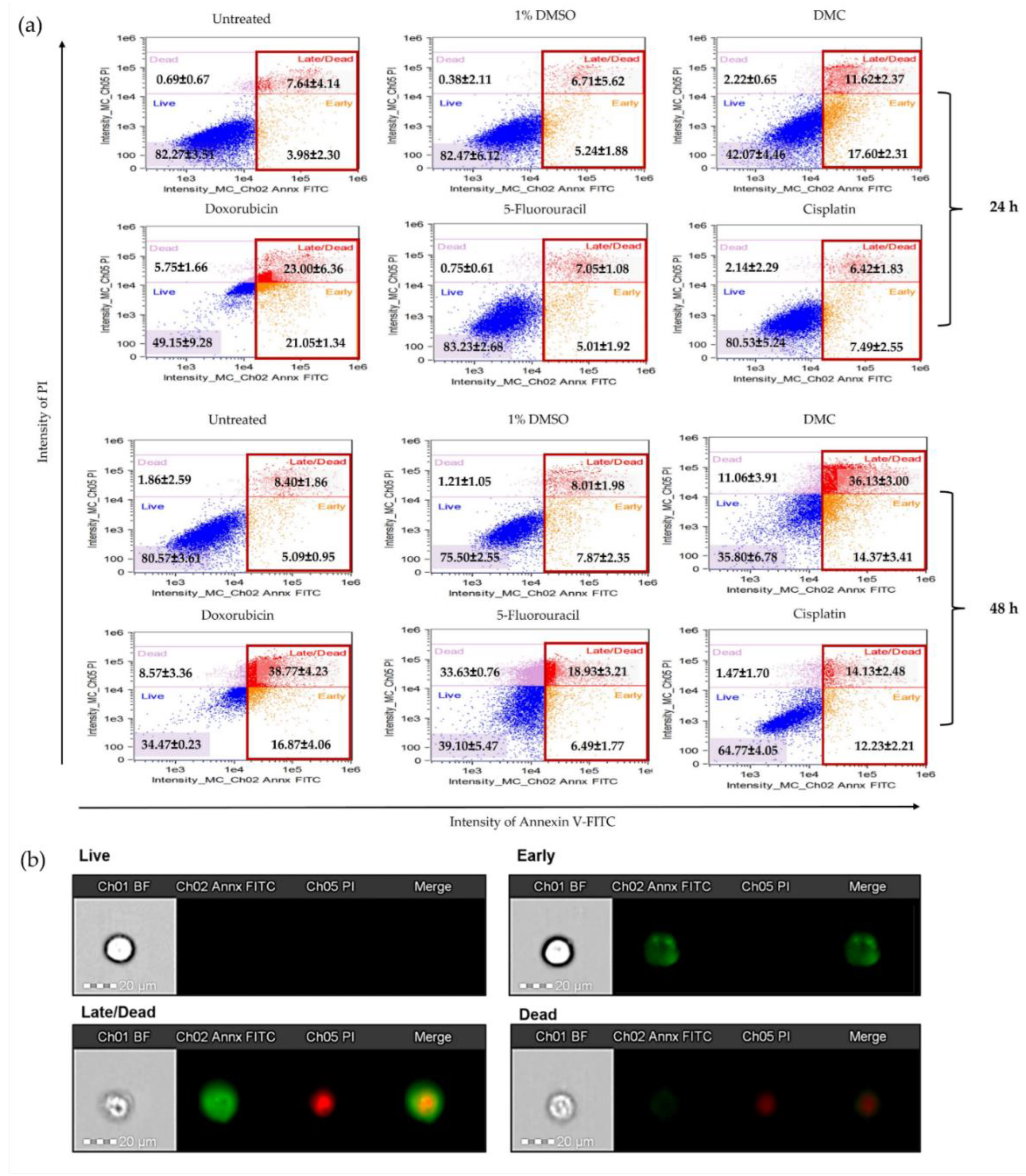
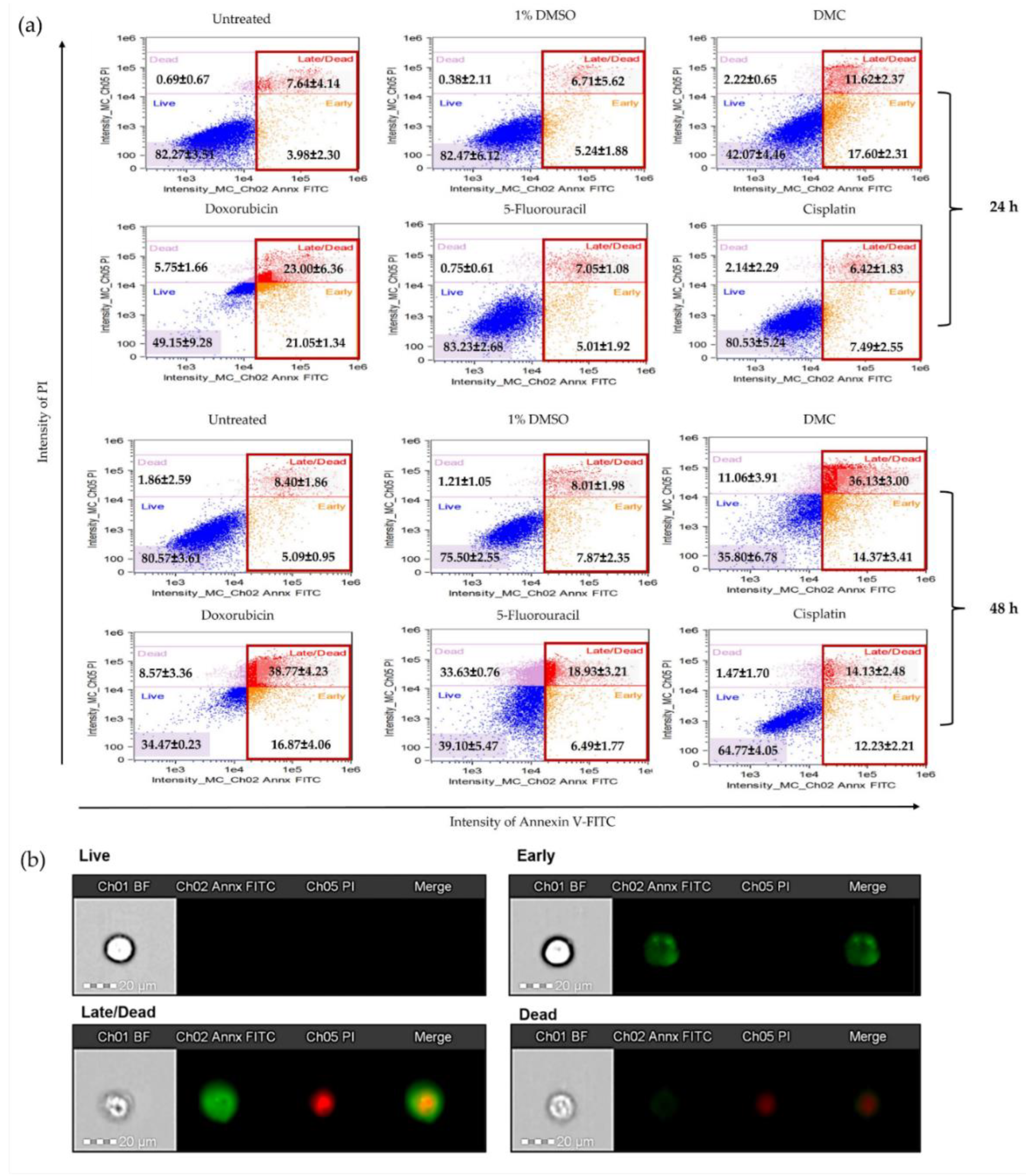
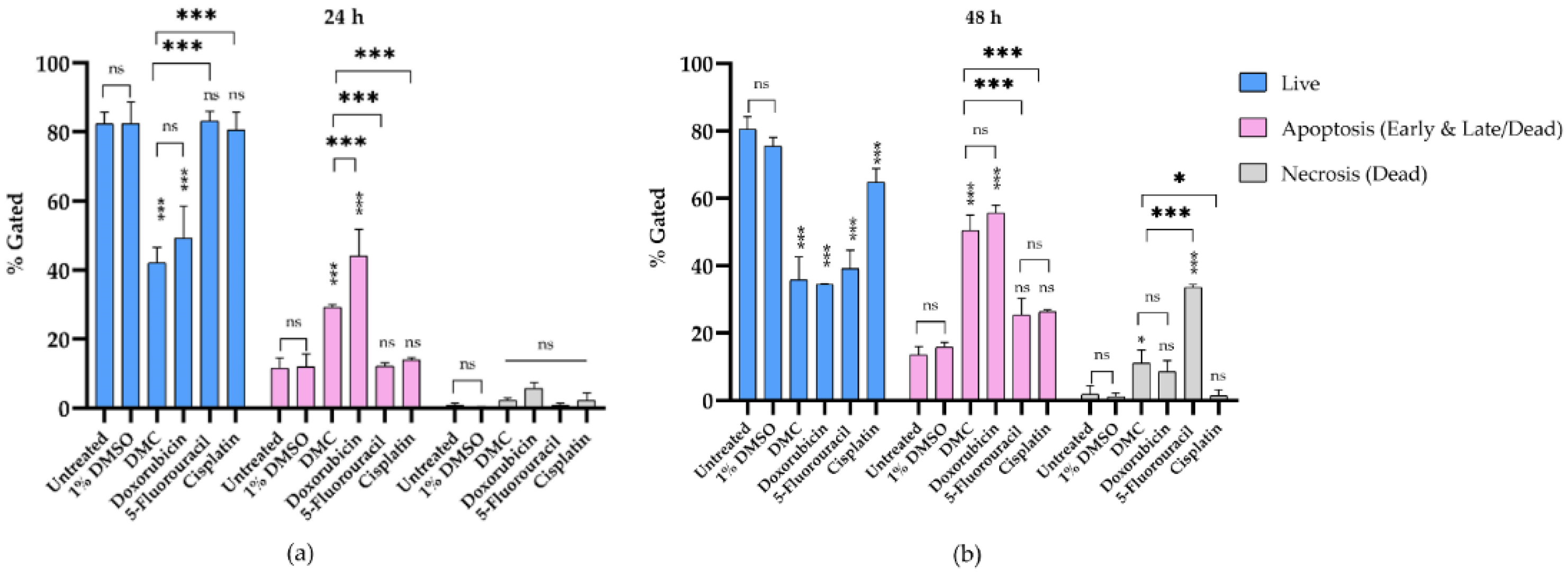
2.3.3. DMC-Induced Apoptosis
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures for Structural Characterisation
4.2. Standard Anti-Cancer Drugs
4.3. Cell Culture
4.4. Isolation and Structural Identification of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone
4.5. Determination of Antiproliferative Activity
4.6. DNA Damage Assessed by the Comet Assay
4.7. Cell Cycle Assay by Flow Cytometry
4.8. Annexin V-FITC and Propidium Iodide Staining to Evaluate Apoptosis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Wilailak, S.; Kengsakul, M.; Kehoe, S. Worldwide initiatives to eliminate cervical cancer. Int. J. Gynecol. Obstet. 2021, 155, 102–106. [Google Scholar] [CrossRef]
- Bhatla, N.; Aoki, D.; Sharma, D.N.; Sankaranarayanan, R. Cancer of the cervix uteri. Int. J. Gynecol. Obstet. 2018, 143, 22–36. [Google Scholar] [CrossRef]
- Marth, C.; Landoni, F.; Mahner, S.; McCormack, M.; Gonzalez-Martin, A.; Colombo, N.; ESMO Guidelines Committee. Cervical cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2017, 28, iv72–iv83. [Google Scholar] [CrossRef]
- Han, R.; Yang, Y.M.; Dietrich, J.; Luebke, A.; Mayer-Pröschel, M.; Noble, M. Systemic 5-fluorouracil treatment causes a syn-drome of delayed myelin destruction in the central nervous system. J. Biol. 2008, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, L.; Ghiselli, S.; Guaran, V.; Chicca, M.; Simoni, E.; Olivetto, E.; Lelli, G.; Martini, A. Correlation of adverse effects of cisplatin administration in patients affected by solid tumours: A retrospective evaluation. Oncol. Rep. 2013, 29, 1285–1292. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Woodle, M.C.; Mixson, A.J. Advances in Delivery Systems for Doxorubicin. J. Nanomed. Nanotechnol. 2018, 9, 519. [Google Scholar] [CrossRef]
- Pham, G.N.; Nguyen, T.T.T.; Nguyen-Ngoc, H. Ethnopharmacology, phytochemistry, and pharmacology of Syzygium ner-vosum. Evidence-Based Complement. Altern. Med. 2020, 2020, 8263670. [Google Scholar]
- Inboot, W.; Taya, S.; Chailungka, A.; Meepowpan, P.; Wongpoomchai, R. Genotoxicity and antigenotoxicity of the methanol extract of Cleistocalyx nervosum var. paniala seed using a Salmonella mutation assay and rat liver micronucleus tests. Mol. Cell. Toxicol. 2012, 8, 19–24. [Google Scholar] [CrossRef]
- Yu, W.-G.; He, H.; Qian, J.; Lu, Y.-H. Dual Role of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone in inhibiting high-mobility group box 1 secretion and blocking its pro-inflammatory activity in hepatic inflammation. J. Agric. Food Chem. 2014, 62, 11949–11956. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-H.; Wang, X.-H.; Yi, Y.-H.; Lee, K.-H. Anti-AIDS agents 54. A potent anti-HIV chalcone and flavonoids from genus Desmos. Bioorg. Med. Chem. Lett. 2003, 13, 1813–1815. [Google Scholar] [CrossRef]
- Luo, Y.; Lu, Y. 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone inhibits apoptosis of MIN6 cells via improving mitochon-drial function. Pharmazie 2012, 67, 798–803. [Google Scholar]
- Chailungka, A.; Junpirom, T.; Pompimon, W.; Nuntasaen, N.; Meepowpan, P. Two flavonoids first isolated from the seed of Syzygium nervosum and preliminary study of their anticancer and anti-HIV-1 reverse transcriptase activities. Maejo Int. J. Sci. Technol. 2017, 11, 58–67. [Google Scholar]
- Khamto, N.; Chaichuang, L.; Rithchumpon, P.; Phupong, W.; Bhoopong, P.; Tateing, S.; Pompimon, W.; Semakul, N.; Chomsri, N.; Meepowpan, P. Synthesis, cytotoxicity evaluation and molecular docking studies on 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone derivatives. RSC Adv. 2021, 11, 31433–31447. [Google Scholar] [CrossRef]
- Santana-Méridas, O.; González-Coloma, A.; Sánchez-Vioque, R. Agricultural residues as a source of bioactive natural products. Phytochem. Rev. 2012, 11, 447–466. [Google Scholar] [CrossRef]
- Srivastava, R.; Shaw, A.K.; Kulshreshtha, D.K. Triterpenoids and chalcone from Syzygium samarangense. Phytochemistry 1995, 38, 687–689. [Google Scholar] [CrossRef]
- Amor, E.C.; Villaseñor, I.M.; Yasin, A.; Choudhary, M.I. Prolyl endopeptidase inhibitors from Syzygium samarangense (Blume) Merr. & L. M. Perry. Z. Naturforsch. C 2004, 59, 86–92. [Google Scholar]
- Hadisaputri, Y.E.; Cahyana, N.; Muchtaridi, M.; Lesmana, R.; Rusdiana, T.; Chaerunisa, A.Y.; Sufiawati, I.; Rostinawati, T.; Subarnas, A. Apoptosis-mediated antiproliferation of A549 lung cancer cells mediated by Eugenia aquea leaf compound 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone and its molecular interaction with caspase receptor in molecular docking simulation. Oncol. Lett. 2020, 19, 3551–3557. [Google Scholar] [CrossRef]
- Ye, C.-L.; Qian, F.; Wei, D.-Z.; Lu, Y.-H.; Liu, J.-W. Induction of apoptosis in K562 human leukemia cells by 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone. Leuk. Res. 2005, 29, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.; Kim, Y.-J.; Amor, E.C.; Lee, J.W.; Kim, H.-C.; Kim, H.J.; Yang, H.O. Induction of autophagy by dimethyl cardamonin is associated with proliferative arrest in human colorectal carcinoma HCT116 and LOVO cells. J. Cell. Biochem. 2011, 112, 2471–2479. [Google Scholar] [CrossRef] [PubMed]
- Tuan, H.N.; Minh, B.H.; Tran, P.T.; Lee, J.H.; Van Oanh, H.; Ngo, Q.M.T.; Nguyen, Y.N.; Lien, P.T.K.; Tran, M.H. The Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone from Cleistocalyx operculatus buds on human pancreatic cancer cell lines. Molecules 2019, 24, 2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, C.-L.; Liu, J.-W.; Wei, D.-Z.; Lu, Y.-H.; Qian, F. In vivo antitumor activity by 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone in a solid human carcinoma xenograft model. Cancer Chemother. Pharmacol. 2005, 56, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.-L.; Lai, Y.-F. 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone, from buds of Cleistocalyx operculatus, induces apop-tosis in human hepatoma SMMC-7721 cells through a reactive oxygen species-dependent mechanism. Cytotechnology 2016, 68, 331–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rammohan, A.; Reddy, J.S.; Sravya, G.; Rao, C.N.; Zyryanov, G.V. Chalcone synthesis, properties and medicinal applications: A review. Environ. Chem. Lett. 2020, 18, 433–458. [Google Scholar] [CrossRef]
- Subarnas, A.; Diantini, A.; Abdulah, R.; Zuhrotun, A.; Hadisaputri, E.; Puspitasari, M.Y.; Yamazaki, C.I.; Kuwano, H.; Koyama, H. Apoptosis induced in MCF-7 human breast cancer cells by 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone isolated from Eugenia aquea Burm f. leaves. Oncol. Lett. 2015, 9, 2303–2306. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.R. The Comet Assay for DNA Damage and Repair: Principles, Applications, and Limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Olive, P.L.; Banáth, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef]
- Speit, G.; Hartmann, A. The Comet Assay: A Sensitive Genotoxicity Test for the Detection of DNA Damage. Methods Mol. Biol. 2005, 291, 085–096. [Google Scholar] [CrossRef]
- Jagetia, A.; Jagetia, G.C.; Jha, S. Naringin, a grapefruit flavanone, protects V79 cells against the bleomycin-induced genotoxicity and decline in survival. J. Appl. Toxicol. 2006, 27, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.-T.; Tung, B.-T.; Nguyen, P.-H.; Thuong, P.-T.; Yoo, S.-S.; Kim, E.-H.; Kim, S.-K.; Oh, W.-K. C-Methylated Flavonoids from Cleistocalyx operculatus and Their Inhibitory Effects on Novel Influenza A (H1N1) Neuraminidase. J. Nat. Prod. 2010, 73, 1636–1642. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, G.; Zhang, P.; Fan, J. Programmed cell death and its role in inflammation. Mil. Med. Res. 2015, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wu, P.; Shen, X.-L.; Wei, X.-Y.; Jiang, Z.-H. Synthesis, cytotoxic activity and drug combination study of tertiary amine derivatives of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone. RSC Adv. 2017, 7, 48031–48038. [Google Scholar] [CrossRef] [Green Version]
- Deans, A.J.; West, S.C. DNA interstrand crosslink repair and cancer. Nat. Rev. Cancer 2011, 11, 467–480. [Google Scholar] [CrossRef] [Green Version]
- He, G.; Siddik, Z.H.; Huang, Z.; Wang, R.; Koomen, J.; Kobayashi, R.; Khokhar, A.R.; Kuang, J. Induction of p21 by p53 fol-lowing DNA damage inhibits both Cdk4 and Cdk2 activities. Oncogene 2005, 24, 2929–2943. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yang, X.; Li, X.; Li, C.; Zhao, L.; Zhou, Y.; Hou, H. Butein sensitizes HeLa cells to cisplatin through the AKT and ERK/p38 MAPK pathways by targeting FoxO3a. Int. J. Mol. Med. 2015, 36, 957–966. [Google Scholar] [CrossRef] [Green Version]
- Kciuk, M.; Bukowski, K.; Marciniak, B.; Kontek, R. Advances in DNA Repair—Emerging Players in the Arena of Eukaryotic DNA Repair. Int. J. Mol. Sci. 2020, 21, 3934. [Google Scholar] [CrossRef]
- Jain, A.; Jahagirdar, D.; Nilendu, P.; Sharma, N.K. Molecular approaches to potentiate cisplatin responsiveness in carcinoma therapeutics. Expert Rev. Anticancer Ther. 2017, 17, 815–825. [Google Scholar] [CrossRef]
- Mann, J.R.; Backlund, M.G.; Dubois, R.N. Mechanisms of Disease: Inflammatory mediators and cancer prevention. Nat. Clin. Pract. Oncol. 2005, 2, 202–210. [Google Scholar] [CrossRef]
- Norbury, C.J.; Zhivotovsky, B. DNA damage-induced apoptosis. Oncogene 2004, 23, 2797–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erasimus, H.; Gobin, M.; Niclou, S.; Van Dyck, E. DNA repair mechanisms and their clinical impact in glioblastoma. Mutat. Res. Mutat. Res. 2016, 769, 19–35. [Google Scholar] [CrossRef]
- Ngabire, D.; Seong, Y.-A.; Patil, M.P.; Niyonizigiye, I.; Seo, Y.B.; Kim, G.-D. Induction of apoptosis and G1 phase cell cycle arrest by Aster incisus in AGS gastric adenocarcinoma cells. Int. J. Oncol. 2018, 53, 2300–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darzynkiewicz, Z.; Juan, G.; Bedner, E. Determining Cell Cycle Stages by Flow Cytometry. Curr. Protoc. Cell Biol. 1999, 1, 8.4.1–8.4.18. [Google Scholar] [CrossRef] [PubMed]
- Rajamanikyam, M.; Vadlapudi, V.; Parvathaneni, S.P.; Koude, D.; Sripadi, P.; Misra, S.; Amanchy, R.; Upadhyayula, S.M. Isolation and characterization of phthalates from Brevibacterium mcbrellneri that cause cytotoxicity and cell cycle arrest. EXCLI J. 2017, 16, 375–387. [Google Scholar] [CrossRef]
- Kantapan, J.; Paksee, S.; Chawapun, P.; Sangthong, P.; Dechsupa, N. Pentagalloyl Glucose- and Ethyl Gallate-Rich Extract from Maprang Seeds Induce Apoptosis in MCF-7 Breast Cancer Cells through Mitochondria-Mediated Pathway. Evid.-Based Complement. Altern. Med. 2020, 2020, 1–19. [Google Scholar] [CrossRef]
- Bian, S.; Zhao, Y.; Li, F.; Lu, S.; Wang, S.; Bai, X.; Liu, M.; Zhao, D.; Wang, J.; Guo, D. 20(S)-Ginsenoside Rg3 Promotes HeLa Cell Apoptosis by Regulating Autophagy. Molecules 2019, 24, 3655. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Half Maximal Inhibitory Concentration a (IC50, µM) | ||
|---|---|---|---|
| C-33A | HeLa | SiHa | |
| DMC | 15.76 ± 1.49 | 10.05 ± 0.22 c | 18.31 ± 3.10 |
| Doxorubicin (+) b | 0.08 ± 0.06 | 0.82 ± 0.07 | 0.23 ± 0.05 |
| 5-Fluorouracil (+) b | 88.43 ± 2.07 | ≥1000 | 205.20 ± 10.61 |
| Cisplatin (+) b | 5.01 ± 0.56 | 9.93 ± 0.16 c | 9.59 ± 0.45 |
| Incubation Time (h) | Sample | DNA Damage Parameters a | |||||
|---|---|---|---|---|---|---|---|
| Tail DNA% | Comparison (Fold Change) | Tail Moment | Comparison (Fold Change) | ||||
| Untreated | 1% DMSO | Untreated | 1% DMSO | ||||
| 6 | Untreated | 4.61 ± 1.62 | - | 2 | 0.52 ± 0.63 | - | 4 |
| 1% DMSO | 2.38 ± 1.63 | 1 | - | 0.14 ± 0.16 | 0 | - | |
| DMC | 65.07 ± 9.57 | 14 | 27 | 53.29 ± 5.21 | 102 | 381 | |
| Doxorubicin | 56.61 ± 3.09 | 12 | 24 | 45.49 ± 4.65 | 87 | 325 | |
| 5-Fluorouracil | 51.33 ± 12.29 | 11 | 22 | 39.33 ± 11.53 | 76 | 281 | |
| Cisplatin | 45.00 ± 11.71 | 10 | 19 | 42.50 ± 10.38 | 82 | 304 | |
| 12 | Untreated | 3.33 ± 2.66 | - | 1 | 0.32 ± 0.43 | - | 1 |
| 1% DMSO | 4.43 ± 0.63 | 1 | - | 0.24 ± 0.10 | 1 | - | |
| DMC | 49.98 ± 8.78 | 15 | 11 | 34.37 ± 5.31 | 107 | 143 | |
| Doxorubicin | 52.37 ± 3.03 | 16 | 12 | 38.84 ± 3.25 | 121 | 162 | |
| 5-Fluorouracil | 44.07 ± 8.13 | 13 | 10 | 28.20 ± 9.70 | 88 | 118 | |
| Cisplatin | 40.40 ± 7.74 | 12 | 9 | 31.45 ± 1.67 | 98 | 131 | |
| 24 | Untreated | 4.44 ± 0.86 | - | 1 | 0.25 ± 0.12 | - | 1 |
| 1% DMSO | 4.16 ± 1.42 | 1 | - | 0.22 ± 0.20 | 1 | - | |
| DMC | 76.55 ± 6.00 | 17 | 18 | 78.13 ± 11.41 | 313 | 355 | |
| Doxorubicin | 78.39 ± 1.64 | 18 | 19 | 60.12 ± 9.20 | 240 | 273 | |
| 5-Fluorouracil | 36.08 ± 11.23 | 8 | 9 | 22.69 ± 10.96 | 91 | 103 | |
| Cisplatin | 41.40 ± 1.49 | 9 | 10 | 30.96 ± 7.40 | 124 | 141 | |
| 48 h | Untreated | 3.67 ±0.67 | - | 1 | 0.14 ± 0.03 | - | 2 |
| 1% DMSO | 3.06 ± 0.48 | 1 | - | 0.09 ± 0.04 | 1 | - | |
| DMC | 92.17 ± 1.76 | 25 | 30 | 116.65 ± 9.93 | 833 | 1296 | |
| Doxorubicin | 86.13 ± 5.81 | 23 | 28 | 72.92 ± 1.91 | 521 | 810 | |
| 5-Fluorouracil | 37.32 ± 9.20 | 10 | 12 | 20.09 ± 5.30 | 144 | 223 | |
| Cisplatin | 29.71 ± 7.51 | 8 | 10 | 16.56 ± 2.57 | 118 | 184 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Utama, K.; Khamto, N.; Meepowpan, P.; Aobchey, P.; Kantapan, J.; Sringarm, K.; Roytrakul, S.; Sangthong, P. Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone from Syzygium nervosum Seeds on Antiproliferative, DNA Damage, Cell Cycle Arrest, and Apoptosis in Human Cervical Cancer Cell Lines. Molecules 2022, 27, 1154. https://doi.org/10.3390/molecules27041154
Utama K, Khamto N, Meepowpan P, Aobchey P, Kantapan J, Sringarm K, Roytrakul S, Sangthong P. Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone from Syzygium nervosum Seeds on Antiproliferative, DNA Damage, Cell Cycle Arrest, and Apoptosis in Human Cervical Cancer Cell Lines. Molecules. 2022; 27(4):1154. https://doi.org/10.3390/molecules27041154
Chicago/Turabian StyleUtama, Kraikrit, Nopawit Khamto, Puttinan Meepowpan, Paitoon Aobchey, Jiraporn Kantapan, Korawan Sringarm, Sittiruk Roytrakul, and Padchanee Sangthong. 2022. "Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone from Syzygium nervosum Seeds on Antiproliferative, DNA Damage, Cell Cycle Arrest, and Apoptosis in Human Cervical Cancer Cell Lines" Molecules 27, no. 4: 1154. https://doi.org/10.3390/molecules27041154
APA StyleUtama, K., Khamto, N., Meepowpan, P., Aobchey, P., Kantapan, J., Sringarm, K., Roytrakul, S., & Sangthong, P. (2022). Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone from Syzygium nervosum Seeds on Antiproliferative, DNA Damage, Cell Cycle Arrest, and Apoptosis in Human Cervical Cancer Cell Lines. Molecules, 27(4), 1154. https://doi.org/10.3390/molecules27041154