
Prevention of Enzymatic Browning by Natural Extracts and Genome-Editing: A Review on Recent Progress


 ,
,  ,
,
Abstract
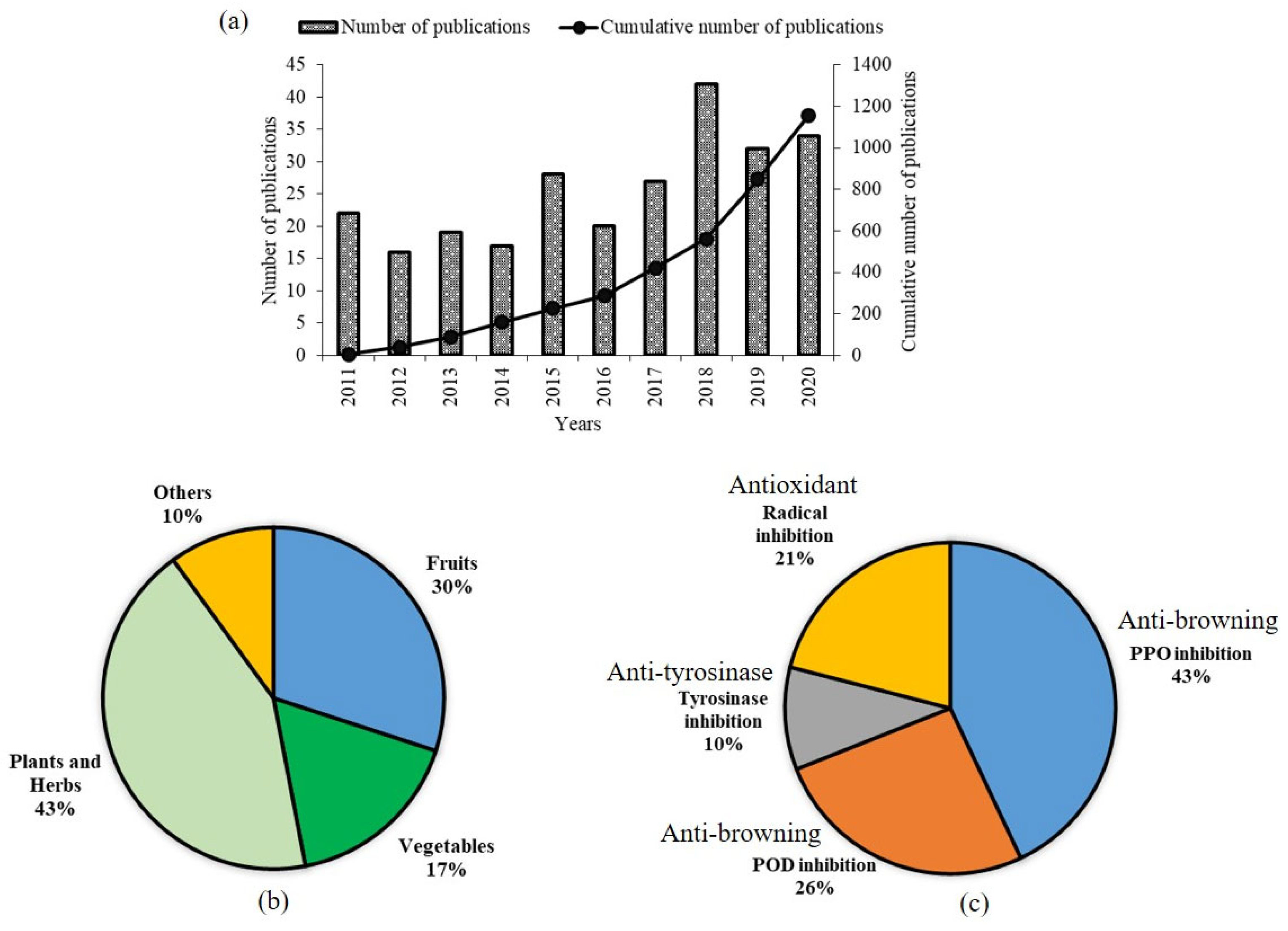
1. Introduction
1.1. Key Motivations of Natural-Based Anti-Browning Research
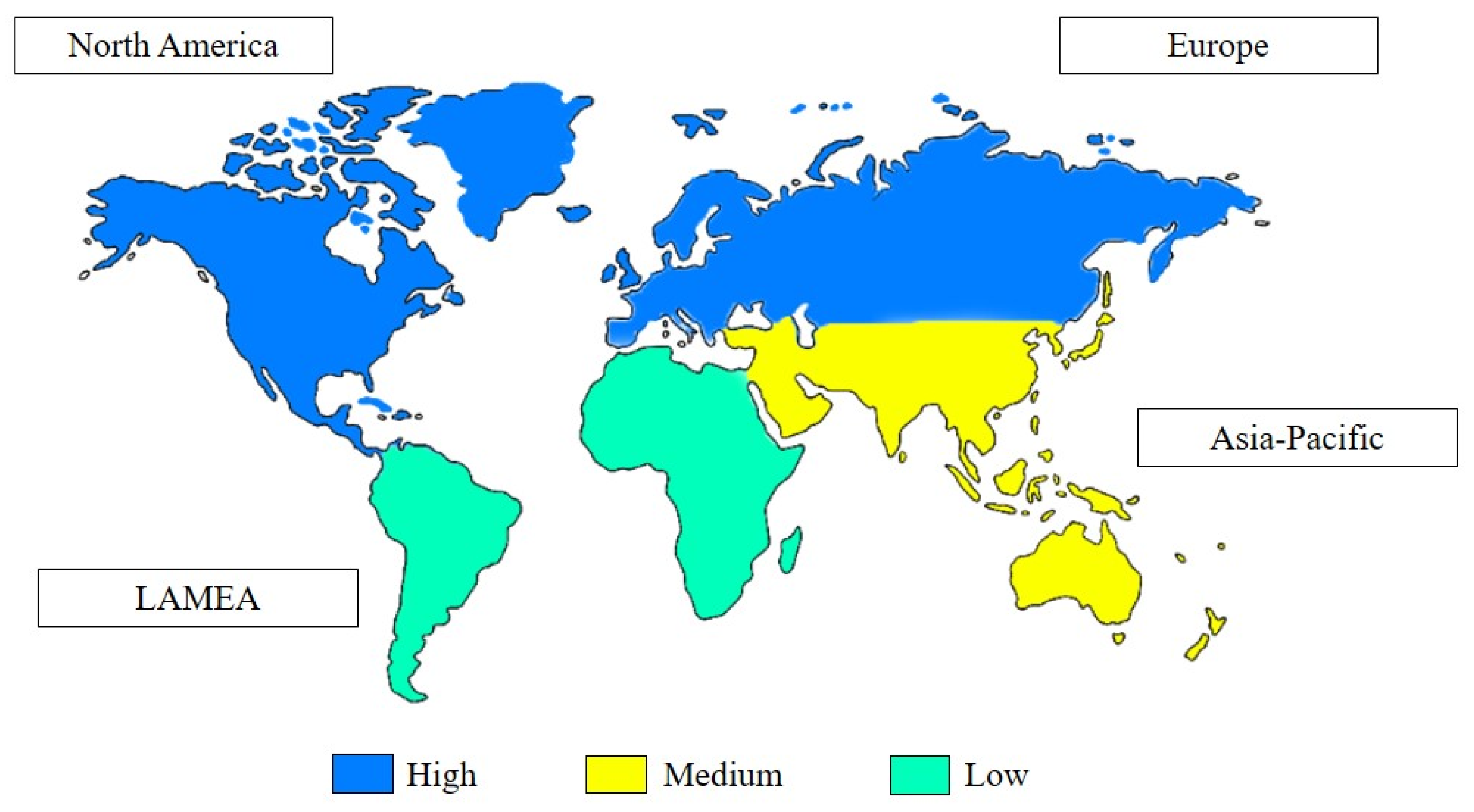
1.2. Market Potential
1.3. Synthetic Versus Natural Sources
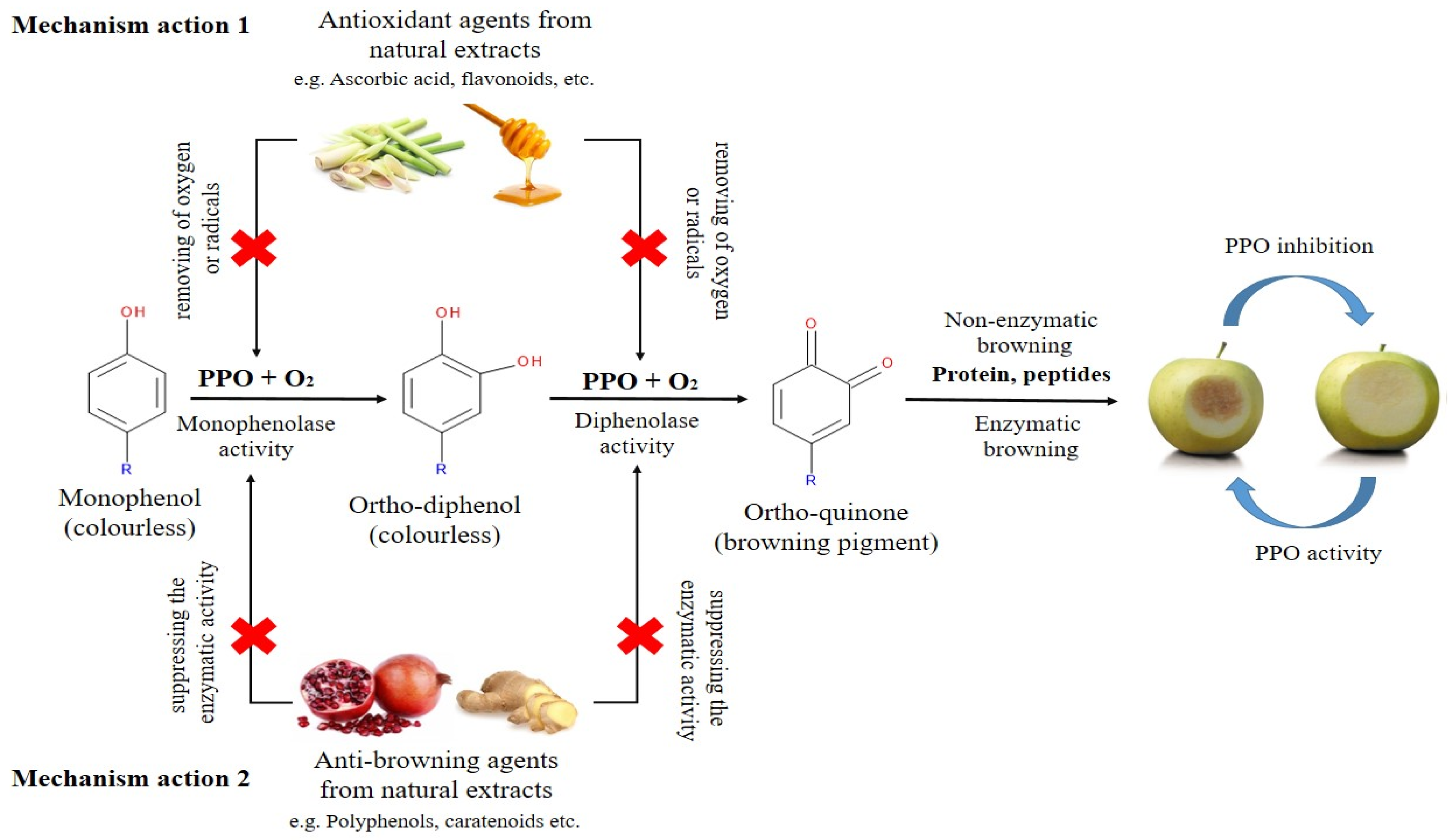
2. Browning Control Mechanisms
3. Compounds Exhibiting Anti-Browning Properties
3.1. Polyphenols
3.2. Carotenoids
3.3. Terpenoids
3.4. Organic Acids
3.5. Bioactive Peptides
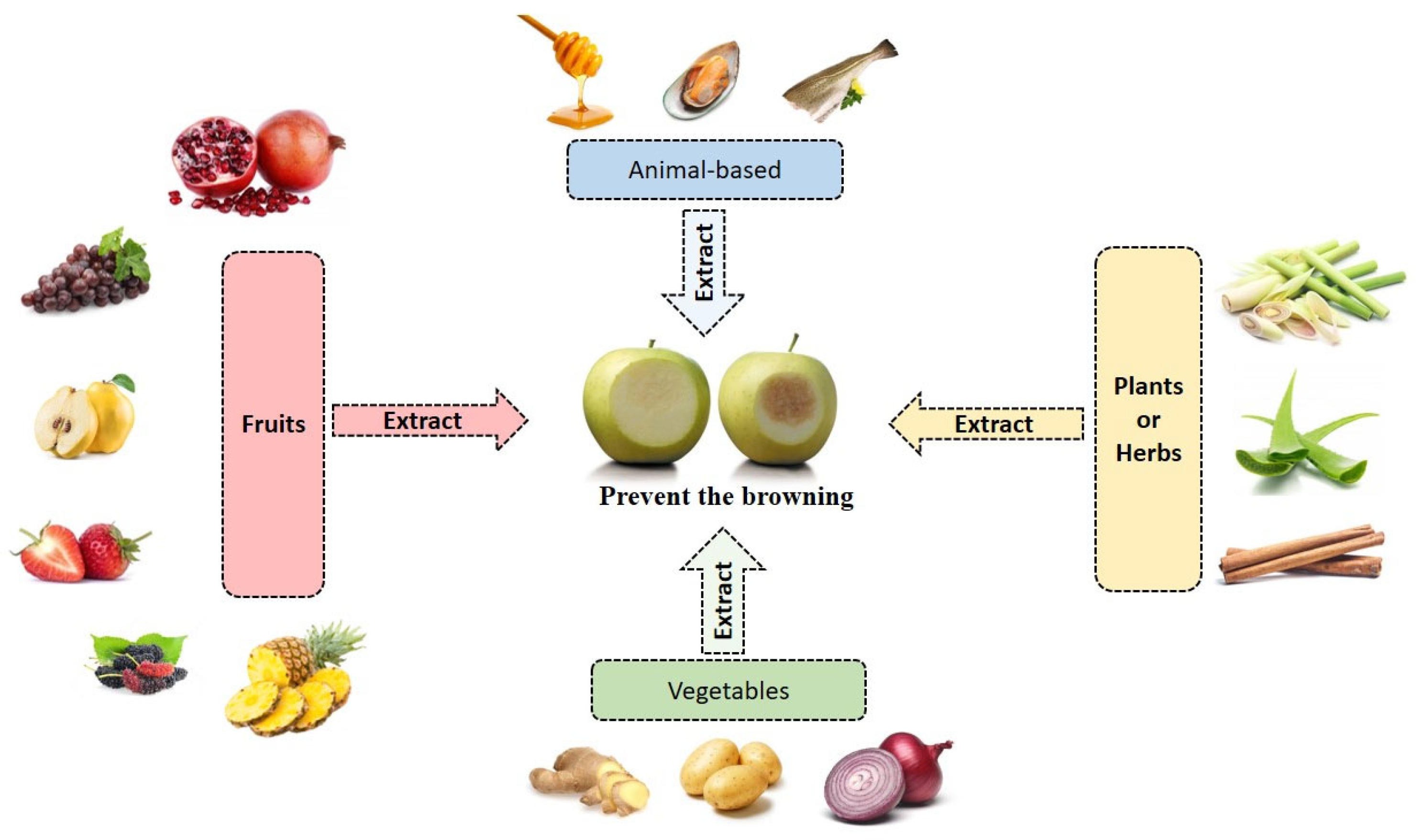
4. Extracts Derived from Fruit Sources

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Sources | Bioactive Compounds/Extracts | Biological Activity | Mechanism of Action | Experimental Results | References |
|---|---|---|---|---|---|
| Fruits sources | |||||
| Unripe grape | Polyphenols (Caffeic acid, catechin, chlorogenic acid, gallic acid, epicatechin, epigallocatechin gallate), organic acids (citric acid, fumaric acid, malic acid, oxalic acid, succinic acid, tartaric acid) | Anti-tyrosinase, antioxidant | -Tyrosinase inhibition -DPPH and FRAP inhibition | The tyrosinase inhibitory activity varied between 67.8% and 68.2%. The grape juice showed the 1 mg/mL DPPH inhibition and 2 mg/mL FRAP inhibition. The anti-tyrosinase and antioxidant properties of unripe grape juices were attributed to the synergistic effects of polyphenols. | Tinello and Lante [71] |
| Unripe grape (Vitis vinifera) | Polyphenol (catechin chlorogenic acid, epicatechin, procyanidin B1 and procyanidin B2), organic acids (Ascorbic acid, citric acid, malic acid) | Antioxidant, anti-browning | -DPPH, ABTS and FRAP inhibition -Inhibit PPO enzymes | The unripe grape juice significantly inhibits the DPPH (3.62 mg/g), ABTS (6.43 mg/g) and FRAP (7.64 mg/g) activity. High antioxidant capacity induced high PPO inhibition activity (30%). | Tinello et al. [61] |
| Quince (Cydonia oblonga) seed | Quince seed extract | Antioxidant | -DPPH inhibition | Quince seed extract significantly reduced the rate of softening, maintained the color values, and reduced the weight loss (3.95%) of tested samples stored at 4 °C for 10 days. Quince seed extract exhibited a strong inhibition of DPPH activity (40.48%). | Kozlu and Elmaci [72] |
| Strawberry leaves and branches | Phenolic compounds (gallic acid, procyanidin B type, catechin, epicatechin, and flavonols) | Antioxidant, anti-browning | -ABTS inhibition -Inhibit PPO and POD enzymes | Strawberry leaves extract showed the inhibiting activity in ABTS (IC50: 0.65 mg/mL), PPO (IC50: 53.92 mg/mL) and POD (IC50: 0.77 mg/mL). Strawberry branches extract showed the inhibiting activity in ABTS (IC50: 0.75 mg/mL), PPO (IC50: 5.97 mg/mL) and POD (IC50: 2.25 mg/mL). | Dias et al. [2] |
| Coconut liquid endosperm | Coconut water | Anti-browning | -Inhibit PPO enzymes | Inhibition of PPO at least 20% during the cold storage (4 °C for 9 days). | Supapvanich et al. [74] |
| Coconut liquid endosperm | Coconut water | Anti-browning | -Inhibit PPO enzymes | The result revealed that visual appearance of the fresh-cut apple was maintained by 50% of coconut water immersion, which lowered the browning index, browning score, and maintained the whiteness index during the cold storage (4 °C) for 7 days. | Supapvanich et al. [75] |
| Mulberry (Morus alba Linn) root bark | 2-arylbenzofuran | Antioxidant, anti-tyrosinase | -DPPH inhibition -Tyrosinase inhibition | The 2-arylbenzofuran derived from M. alba root bark manifested the good inhibitory of DPPH activity (IC50 ranging from 11.58 to 55.73 µM). The 2-arylbenzofuran exhibited good tyrosinase inhibition with IC50 value of 4.45 µM. | Paudel et al. [9] |
| Mulberry (M. alba) twigs | Polyphenol (flavone morin, rutin, resveratrol, maclurin) | Anti-tyrosinase | -Tyrosinase inhibition | The flavone compounds in M. alba twigs were found to inhibit tyrosinase activity with IC50 values between 0.07–8.0 μM. | Zhang et al. [79] |
| Pomegranate (Punica granatum L.) peel | Phenolic compound (Gallic acid) | Antioxidant, anti-tyrosinase | -DPPH inhibition -Tyrosinase inhibition | The IC50 for DPPH inhibitory activity varied between 307 to 42 mg/g. The tyrosinase inhibitory activity was approximately 60%. | Turrini et al. [83] |
| Tomato (Solanum lycopersicum L.) skin | Carotenoid (Lycopene) | Antioxidant, antimicrobial | -FRAP inhibition -Microbial growth inhibition | Lycopene of tomato skin showed FRAP inhibition activity with 824.0 mg/kg to 860.9 mg/kg. Lycopene treatment reduced the sample browning and inhibited the microbial growth after 9 days stored at 5 °C. | Martínez-Hernández et al. [55] |
| Mango (Mangifera indica) peel | Phenolic compound (Mangiferin, protocatechuic and gallic acid) | Anti-browning | -Inhibit PPO enzymes | The phenolic compounds of mango peels showed inhibitory effect on PPO with an IC50 of 0.3 mg/mL. | Jirasuteeruk and Theerakulkait [88] |
| Pineapple (Ananas comosus) | Phenolic compound | Anti-browning | N.M | High phenolic contents and reduced the browning development with low browning index (less than 1). | Supapvanich et al. [90] |
| Vegetables sources | |||||
| Ginger (Zingiber officinale) | Ginger extract | Anti-browning | -Inhibit PPO and POD enzymes | The ginger extract showed the 60.90% inhibitory activity of PPO and 48.10% inhibitory activity of POD. | Weerawardana et al. [92] |
| Potato (Solanum tuberosum) peel | Phenolic compound (caffeic acid and chlorogenic acid) | Antioxidant | -ABTS and FRAP inhibition | The potato peel extract showed ABTS and FRAP inhibitory effect with the total antioxidant capacity values of 0.21 μmol/mL and 0.28 μmol/mL, respectively. Reduced browning and slowed down the fruits softening can be observed during the storage for 3 days at 4 °C. | Venturi et al. [93] |
| Onion (Allium cepa) | Onion extract | Anti-browning | -Inhibit PPO enzymes | The onion extract manifested the good inhibition percentage on PPO activity (15.89–33.11%). | Lim and Wong [7] |
| Onion (Allium cepa) | Onion extract | Antioxidant | -ABTS inhibition | The onion extract displayed a good antiradical activity in ABTS (62.10%) after 30 days of storage. | Bustos et al. [94] |
| Onion (Allium cepa) | Polyphenols and flavonoids | Antioxidant, anti-browning | -DPPH and ABTS inhibition | The onion extracts exhibited higher DPPH inhibition activity (0.16 mmol/g) and ABTS inhibition activity (1 mmol/g). The onion extracts not only inhibited the browning development but also improved the nutritional quality of apple juice. | Lee et al. [95] |
| Chili pepper (Capsicum sp.) | Ascorbic acid | Anti-browning | -Inhibit PPO enzymes | The PPO activity inhibition by extracts of chili pepper was 70%. | Mercimek et al. [96] |
| Chili pepper (Capsicum sp.) | Chili pepper extract | Anti-browning | -Inhibit PPO enzymes | The chili pepper extracts showed the great potential of inhibition (45.97%) on PPO enzyme. | Lim et al. [97] |
| Plants and Herbs sources | |||||
| Mangrove plant leaves (Bruguiera gymnorhiza) | Polyphenols (Tannins) | Anti-tyrosinase, anti-browning, antioxidant | -Tyrosinase inhibition -DPPH, ABTS and FRAP inhibition -Inhibit PPO and POD enzymes | The tannins extracted from the B. gymnorhiza leaves exhibited strong anti-tyrosinase activity (IC50: 123.90 µg/mL). The IC50 value for antioxidant in DPPH, ABTS and FRAP were 88.81 µg/mL, 105.03 µg/mL, 1052.27 mg/g, respectively. Effective inhibited PPO and POD activity in fresh-cut lotus root stored at 4 °C for 17 days. | Liu et al. [98] |
| Mangrove plant leaves (Bruguiera gymnorhiza) | Flavonoids and phenolic compounds | Anti-browning | -Inhibit PPO enzymes | B. gymnorhiza leaves exhibited the greatest inhibition on sweet potato PPO (at least 50%). | Lim et al. [13] |
| Oregano Herb (Origanum vulgare) aerial parts | O. vulgare extract | Anti-browning | -Inhibit PPO enzymes | Prevention of enzymatic browning by PPO inhibition (64.50%). | Tanhaş et al. [99] |
| Citronella (Cymbopogon nardus) hydrosols | Terpenoids | Anti-browning | -Inhibit PPO and POD enzymes | Effectively suppressed the browning development by lowering the PPO and POD activities. | Xiao et al. [59] |
| Cinnamon | Cinnamon essential oil | Anti-browning | -Inhibit PPO enzymes | Decreased browning by inhibiting the PPO activity (80–90%). | Xu et al. [100] |
| Purslane (Portulaca oleracea L.) | Polyphenols and alkaloids | Anti-browning | -Inhibit PPO and POD enzymes | The 0.05% (w/w) purslane extract inhibited the PPO and POD activity in entire 8 days storage at 4 °C. | Liu et al. [68] |
| Purslane (Portulaca oleracea L.) | Purslane extract | Anti-browning | -Inhibit PPO and POD enzymes | Coupling the ultrasound treatment with purslane extract (0.02%) greatly promote the anti-browning effects on the fresh-cut potato across entire 8 days storage at 4 °C. | Zhu et al. [101] |
| Green tea leaves | Green tea extract | Anti-browning | -Inhibit PPO enzymes | Green tea extract (3 mg/mL) inhibit the PPO activity at least 86%. | Klimczak and Gliszczyńska-Świgło [102] |
| Green tea leaves (Camellia sinensis) | Flavonoids (catechins) | Anti-browning | -Inhibit PPO enzymes | Green tea extract showed high PPO inhibitory activity at least 80%. | Chang and Kim [103] |
| Aloe (Aloe vera) | A. vera extract | Anti-browning | -Inhibit PPO and POD enzymes | The fresh-cut fruits treated with A. vera extract showed the lowest browning score (1.94%), reduced the PPO and POD activity across entire 6 days storage at 4 °C. | Supapvanich et al. [104] |
| Stevia plant (Stevia rebaudiana) leaves | Stevia leaves extract | Anti-browning | -Inhibit PPO and POD enzymes | Reducing the PPO and POD activity with the incubation time when the Stevia leaves extract were added. | Criado et al. [105] |
| Others | |||||
| Manuka Honey | Honey extract | Anti-browning | -Inhibit PPO enzymes | The inhibitory effect of honey extract on PPO activity varied from a range of 41.39% to 48.0%. | Lim et al. [97] |
| Egg white | Peptides | Antioxidant, Anti-tyrosinase | -DPPH, and ABTS inhibition -Tyrosinase inhibition | The bioactive peptide derived from egg white exhibit the good antioxidant activity in DPPH (20%), ABTS (0.3 mg/mL) and tyrosinase inhibition (IC50: 2.90 mg/mL). | Thaha et al. [67] |
| Cod fish skin | Peptides | Anti-browning | -Inhibit PPO and POD enzymes | The cod fish derived bioactive peptides significantly inhibit the PPO and POD activity during the entire 8 days storage at 4 °C. | Liu et al. [106] |
| Buffalo whey | Peptides | Antioxidant, Anti-browning | -ABTS inhibition -Inhibit PPO enzymes | The buffalo whey peptides exhibited significant increase in antioxidant activity and PPO-inhibitory activity (50% inhibition). | da Silva et al. [69] |
| Blue mussel (Mytilus edulis) | Organic compound (Hypotaurine and sulfinic acids) | Anti-browning | -Inhibit PPO enzymes | The hypotaurine and sulfinic acids compound found in blue mussel extract exhibited great inhibition of PPO activity (ranging from 89% to 100%). | Schulbach et al. [107] |
5. Extracts Derived from Vegetables Sources
6. Extracts Derived from Plants or Herbs Sources
7. Extracts Derived from Other Sources
8. Potential Application of Genome-Editing in Preventing Browning Activity and Improving Postharvest Quality
8.1. Current Application of CRISPR/Cas Technology in Preventing Browning
8.2. Challenge in the Application of CRISPR/Cas9 System in Crop Plants
8.3. Other Application of Genome-Editing Tools in Preventing Browning Effect
9. Method of Incorporations with Natural Extracts in Preventing Browning
10. Patents
11. Conclusions and Future Research Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tinello, F.; Lante, A. Recent Advances in Controlling Polyphenol Oxidase Activity of Fruit And Vegetable Products. Innov. Food Sci. Emerg.Technol. 2018, 50, 73–83. [Google Scholar] [CrossRef]
- Dias, C.; Fonseca, A.M.A.; Amaro, A.L.; Vilas-Boas, A.A.; Oliveira, A.; Santos, S.A.O.; Silvestre, A.J.D.; Rocha, S.M.; Isidoro, N.; Pintado, M. Natural-Based Antioxidant Extracts as Potential Mitigators of Fruit Browning. Antioxidant 2020, 9, 715. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.M.; Kwon, E.B.; Lee, B.; Kim, C.Y. Recent Trends in Controlling the Enzymatic Browning of Fruit and Vegetable Products. Molecules 2020, 25, 2754. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, S.; Simpson, B.K. Enzymes in Food Bioprocessing—Novel Food Enzymes, Applications, and Related Techniques. Curr. Opin. Food Sci. 2018, 19, 30–35. [Google Scholar] [CrossRef]
- Paravisini, L.; Peterson, D.G. Mechanisms Non-Enzymatic Browning in Orange Juice During Storage. Food Chem. 2019, 289, 320–327. [Google Scholar] [CrossRef]
- Shrestha, L.; Kulig, B.; Moscetti, R.; Massantini, R.; Pawelzik, E.; Hensel, O.; Sturm, B. Optimisation of Physical and Chemical Treatments to Control Browning Development and Enzymatic Activity on Fresh-cut Apple Slices. Foods 2020, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.Y.; Wong, C.W. Inhibitory Effect of Chemical and Natural Anti-Browning Agents on Polyphenol Oxidase from Ginger (Zingiber officinale Roscoe). J. Food Sci. Technol. 2018, 55, 3001–3007. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Suri, K.; Shevkani, K.; Kaur, A.; Kaur, A.; Singh, N. Enzymatic Browning of Fruit And Vegetables: A Review. In Enzymes in Food Technology; Springer: New York, NY, USA, 2018; pp. 63–78. [Google Scholar]
- Paudel, P.; Seong, S.H.; Wagle, A.; Min, B.S.; Jung, H.A.; Choi, J.S. Antioxidant and Anti-Browning Property of 2-Arylbenzofuran Derivatives From Morus Alba Linn Root Bark. Food Chem. 2020, 309, 125739. [Google Scholar] [CrossRef]
- De Corato, U. Improving the Shelf-Life and Quality of Fresh and Minimally-Processed Fruits and Vegetables for A Modern Food Industry: A Comprehensive Critical Review from The Traditional Technologies into The Most Promising Advancements. Crit. Rev. Food Sci. Nutr. 2020, 60, 940–975. [Google Scholar] [CrossRef]
- Bobo-García, G.; Arroqui, C.; Merino, G.; Vírseda, P. Antibrowning Compounds for Minimally Processed Potatoes: A Review. Food Rev. Int. 2020, 36, 529–546. [Google Scholar] [CrossRef]
- Favre, L.C.; Dos Santos, C.; Lopez-Fernandez, M.P.; Mazzobre, M.F.; Buera, M.D.P. Optimization of Beta-Cyclodextrin-Based Extraction of Antioxidant and Anti-Browning Activities from Thyme Leaves by Response Surface Methodology. Food Chem. 2018, 265, 86–95. [Google Scholar] [CrossRef]
- Lim, W.Y.; Cheng, Y.W.; Lian, L.B.; Chan, E.W.C.; Wong, C.W. Inhibitory Effect of Malaysian Coastal Plants on Banana (Musa acuminata colla “Lakatan”), Ginger (Zingiber officinale Roscoe) and Sweet Potato (Ipomoea batatas) Polyphenol Oxidase. J. Food Sci. Technol. 2020, 58, 4178–4184. [Google Scholar] [CrossRef] [PubMed]
- Salminen, W.F.; Russotti, G. Synergistic Interaction of Ascorbic Acid and Green Tea Extract in Preventing The Browning of Fresh Cut Apple Slices. J. Food Process. Preserv. 2017, 41, e13192. [Google Scholar] [CrossRef]
- Yang, Z.K.; Zou, X.B.; Li, Z.H.; Huang, X.W.; Zhai, X.D.; Zhang, W.; Shi, J.Y.; Tahir, H.E. Improved Postharvest Quality of Cold Stored Blueberry by Edible Coating Based on Composite Gum Arabic/Roselle Extract. Food Bioproc. Technol. 2019, 12, 1537–1547. [Google Scholar] [CrossRef]
- Sarkar, S.; Ahmed, M.; Mozumder, N.; Saeid, A. Isolation and Characterization of Bromelain Enzyme from Pineapple and Its Utilization As Anti-Browning Agent. Process Eng. J. 2017, 1, 52–58. [Google Scholar]
- Fernandes, S.S.; Coelho, M.S.; de las Mercedes Salas-Mellado, M. Bioactive Compounds as Ingredients of Functional Foods: Polyphenols, Carotenoids, Peptides from Animal and Plant Sources New. In Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2019; pp. 129–142. [Google Scholar]
- Peanparkdee, M.; Iwamoto, S. Bioactive Compounds from By-Products of Rice Cultivation and Rice Processing: Extraction and Application in The Food and Pharmaceutical Industries. Trends Food Sci. Technol. 2019, 86, 109–117. [Google Scholar] [CrossRef]
- Wang, Y.X.; Zhou, F.H.; Zuo, J.H.; Zheng, Q.L.; Gao, L.P.; Wang, Q.; Jiang, A.L. Pre-storage Treatment of Mechanically-Injured Green Pepper (Capsicum annuum L.) Fruit with Putrescine Reduces Adverse Physiological Responses. Postharvest Biol. Technol. 2018, 145, 239–246. [Google Scholar] [CrossRef]
- Ortiz, C.M.; Vicente, A.R.; Fields, R.P.; Grillo, F.; Labavitch, J.M.; Donis-Gonzalez, I.; Crisosto, C.H. Walnut (Juglans regia L.) Kernel Postharvest Deterioration As Affected by Pellicle Integrity, Cultivar and Oxygen Concentration. Postharvest Biol. Technol. 2019, 156, 110948. [Google Scholar] [CrossRef]
- FAO. The Future of Food and Agriculture–Trends and Challenges; FAO: Rome, Italy, 2017; ISBN 9789251095515. [Google Scholar]
- Erdinc, B.; Demir, K.; Torcuk, A.I. Postharvest Putrescine and Ultrasound Treatments to Improve Quality and Postharvest Life of Table Grapes (Vitis vinifera L.) cv. Michele Palieri. J. Cent. Eur. Agric. 2017, 18, 598–615. [Google Scholar] [CrossRef]
- Putnik, P.; Bursać Kovačević, D.; Herceg, K.; Levaj, B. Influence of Cultivar, Anti-Browning Solutions, Packaging Gasses, and Advanced Technology on Browning in Fresh-Cut Apples During Storage. J. Food Process Eng. 2017, 40, e12400. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, W.; Liu, S.; Liu, C.; Zheng, L. Effects of Melatonin Treatment on The Enzymatic Browning and Nutritional Quality of Fresh-Cut Pear Fruit. Food Chem. 2019, 299, 125116. [Google Scholar] [CrossRef]
- Sarpong, F.; Yu, X.J.; Zhou, C.S.; Oteng-Darko, P.; Amenorfe, L.P.; Wu, B.G.; Bai, J.W.; Ma, H.L. Drying Characteristic, Enzyme Inactivation and Browning Pigmentation Kinetics of Controlled Humidity-Convective Drying of Banana Slices. Heat Mass. Transf. 2018, 54, 3117–3130. [Google Scholar] [CrossRef]
- Mirshekari, A.; Madani, B.; Golding, J.B. Aloe Vera Gel Treatment Delays Postharvest Browning of White Button Mushroom (Agaricus bisporus). J. Food Meas.Charact. 2019, 13, 1250–1256. [Google Scholar] [CrossRef]
- Huang, S.J.; Lin, S.Y.; Wang, T.T.; Hsu, F.C. Combining Acetic Acid and Ethanol as An Anti-Browning Treatment for Lettuce Butt Discoloration Through Repression of The Activity and Expression of Phenylalanine Ammonia Lyase. Postharvest Biol.Technol. 2020, 164, 111151. [Google Scholar] [CrossRef]
- Bayrak, S.; Ozturk, C.; Demir, Y.; Alim, Z.; Kufrevioglu, O.I. Purification of Polyphenol Oxidase from Potato and Investigation of the Inhibitory Effects of Phenolic Acids on Enzyme Activity. Protein Pept. Lett. 2020, 27, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ren, L.; Li, M.; Qian, J.; Fan, J.; Du, B. Effects of Clove Essential Oil and Eugenol on Quality and Browning Control of Fresh-Cut Lettuce. Food Chem. 2017, 214, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Kerdudo, A.; Burger, P.; Merck, F.; Dingas, A.; Rolland, Y.; Michel, T.; Fernandez, X. Development of A Natural Ingredient–Natural Preservative: A Case Study. C. R. Chim. 2016, 19, 1077–1089. [Google Scholar] [CrossRef]
- Mei, J.; Ma, X.; Xie, J. Review on Natural Preservatives for Extending Fish Shelf Life. Foods 2019, 8, 490. [Google Scholar] [CrossRef] [PubMed]
- Misra, N.N.; Koubaa, M.; Roohinejad, S.; Juliano, P.; Alpas, H.; Inacio, R.S.; Saraiva, J.A.; Barba, F.J. Landmarks in The Historical Development of Twenty First Century Food Processing Technologies. Food Res. Int. 2017, 97, 318–339. [Google Scholar] [CrossRef]
- Hintz, T.; Matthews, K.K.; Di, R. The Use of Plant Antimicrobial Compounds for Food Preservation. Biomed. Res. Int. 2015, 2015, 12. [Google Scholar] [CrossRef]
- Baqutayan, S.M.; Mohamad, R.; Azman, R.R.; Hassan, N.A. The Implementation of Contract Farming of Fresh Fruits and Vegetables (FFV) for Smallholders in Malaysia: Government Roles and Initiatives. J. Sci. Technol. Policy Manag. 2017, 3, 1–8. [Google Scholar] [CrossRef]
- Sillani, S.; Nassivera, F. Consumer Behavior in Choice of Minimally Processed Vegetables and Implications for Marketing Strategies. Trends Food Sci. Technol. 2015, 46, 339–345. [Google Scholar] [CrossRef][Green Version]
- Research, G.V. Food Preservatives Market Size, Share and Trends Analysis Report by Type (Natural, Synthetic), by Function (Anti-microbial, Anti-oxidant), by Application (Meat and Poultry, Beverages), and Segment Forecasts, 2020–2027. Available online: https://www.grandviewresearch.com/industry-analysis/food-preservatives-market (accessed on 24 November 2021).
- Intelligence, M. Food Preservatives Market-Growth, Trends, Covid-19 Impact, and Forecasts (2021–2026). Available online: https://www.mordorintelligence.com/industry-reports/food-preservatives-market (accessed on 24 November 2021).
- Singla, G.; Chaturvedi, K.; Sandhu, P.P. Status and Recent Trends in Fresh-Cut Fruits and Vegetables. In Fresh-Cut Fruits and Vegetables; Elsevier: Amsterdam, The Netherlands, 2020; pp. 17–49. [Google Scholar]
- Ozsoy, O.; Aras, S.; Ozkan, A.; Parlak, H.; Gemici, B.; Uysal, N.; Aslan, M.; Yargicoglu, P.; Agar, A. The Effect of Ingested Sulfite on Active Avoidance in Normal and Sulfite Oxidase-Deficient Aged Rats. Toxicol. Mech. Methods. 2017, 27, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, N.; Nematollahi, Z.; Hoseini, E. Effect of Sodium Metabisulfite on Rat Ovary and Lipid Peroxidation. Iran. J. Toxicol. 2016, 10, 23–28. [Google Scholar] [CrossRef]
- Shekarforoush, S.; Ebrahimi, Z.; Hoseini, M. Sodium Metabisulfite-Induced Changes on Testes, Spermatogenesis and Epididymal Morphometric Values in Adult Rats. Int. J. Reprod. Biomed. 2015, 13, 765–770. [Google Scholar] [CrossRef]
- Vally, H.; Misso, N.L. Adverse Reactions to The Sulphite Additives. Gastroenterol. Hepatol. Bed. Bench. 2012, 5, 16–23. [Google Scholar]
- Shekarforoush, S.; Ebrahimi, P.; Fathabad, A.A.; Farzanfar, E. Effect of Sodium Metabisulfite on Oxidative Stress and Lipid Peroxidation Biomarkers. Curr. Nutr. Food. Sci. 2020, 16, 114–117. [Google Scholar] [CrossRef]
- Ozturk, O.H.; Oktar, S.; Aydin, M.; Kucukatay, V. Effect of Sulfite on Antioxidant Enzymes and Lipid Peroxidation in Normal and Sulfite Oxidase-Deficient Rat Erythrocytes. J. Physiol. Biochem. 2010, 66, 205–212. [Google Scholar] [CrossRef]
- Franco, R.; Martinez-Pinilla, E. Chemical Rules on The Assessment of Antioxidant Potential in Food and Food Additives Aimed at Reducing Oxidative Stress and Neurodegeneration. Food Chem. 2017, 235, 318–323. [Google Scholar] [CrossRef]
- Research, A.M. Natural Food Preservatives Market Outlook-2026. Available online: https://cutt.ly/bOQWEtQ (accessed on 24 November 2021).
- Sae-leaw, T.; Benjakul, S. Prevention of Melanosis in Crustaceans by Plant Polyphenols: A Review. Trends Food Sci. Technol. 2019, 85, 1–9. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Tundis, R.; Menichini, F. Natural and Synthetic Tyrosinase Inhibitors as Antibrowning Agents: An Update. Compr. Rev. Food Sci. Food Saf. 2012, 11, 378–398. [Google Scholar] [CrossRef]
- Nokthai, P.; Lee, V.S.; Shank, L. Molecular Modeling of Peroxidase and Polyphenol Oxidase: Substrate Specificity and Active Site Comparison. Int. J. Mol. Sci. 2010, 11, 3266–3276. [Google Scholar] [CrossRef]
- Cirkovic Velickovic, T.D.; Stanic-Vucinic, D.J. The Role of Dietary Phenolic Compounds in Protein Digestion and Processing Technologies to Improve Their Antinutritive Properties. Compr. Rev. Food Sci. Food Saf. 2018, 17, 82–103. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total Phenolic Content, Flavonoid Content and Antioxidant Potential of Wild Vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of Antioxidant Potential of Plants and Its Relevance to Therapeutic Applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef]
- Mezzomo, N.; Ferreira, S.R.S. Carotenoids Functionality, Sources, and Processing by Supercritical Technology: A Review. J. Chem. 2016, 2016, 3164312. [Google Scholar] [CrossRef]
- Strati, I.; Oreopoulou, V. Recovery of Carotenoids from Tomato Processing By-Products–A Review. Food Res. Int. 2014, 65, 311–321. [Google Scholar] [CrossRef]
- Martínez-Hernández, G.B.; Castillejo, N.; Artés-Hernández, F. Effect of Fresh–Cut Apples Fortification with Lycopene Microspheres, Revalorized from Tomato By-Products, During Shelf Life. Postharvest Biol. Technol. 2019, 156, 110925. [Google Scholar] [CrossRef]
- Spagna, G.; Barbagallo, R.N.; Chisari, M.; Branca, F. Characterization of a Tomato Polyphenol Oxidase and Its Role in Browning and Lycopene Content. J. Agric. Food Chem. 2005, 53, 2032–2038. [Google Scholar] [CrossRef] [PubMed]
- Proshkina, E.; Plyusnin, S.; Babak, T.; Lashmanova, E.; Maganova, F.; Koval, L.; Platonova, E.; Shaposhnikov, M.; Moskalev, A. Terpenoids as Potential Geroprotectors. Antioxidant 2020, 9, 529. [Google Scholar] [CrossRef] [PubMed]
- Reyes, B.A.; Dufourt, E.C.; Ross, J.; Warner, M.J.; Tanquilut, N.C.; Leung, A.B. Selected Phyto and Marine Bioactive Compounds: Alternatives for the Treatment of Type 2 Diabetes. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; Volume 55, pp. 111–143. [Google Scholar]
- Xiao, Y.; He, J.; Zeng, J.; Yuan, X.; Zhang, Z.; Wang, B. Application of Citronella and Rose Hydrosols Reduced Enzymatic Browning of Fresh-Cut Taro. J Food Biochem. 2020, 44, e13283. [Google Scholar] [CrossRef]
- Nakatsu, T.; Lupo, A.T., Jr.; Chinn, J.W., Jr.; Kang, R.K. Biological Activity of Essential Oils and Their Constituents. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2000; Volume 21, pp. 571–631. [Google Scholar]
- Tinello, F.; Mihaylova, D.; Lante, A. Effect of Dipping Pre-Treatment with Unripe Grape Juice on Dried “Golden Delicious” Apple Slices. Food Bioproc. Tech. 2018, 11, 2275–2285. [Google Scholar] [CrossRef]
- Hajfathalian, M.; Ghelichi, S.; García-Moreno, P.J.; Moltke Sørensen, A.-D.; Jacobsen, C. Peptides: Production, Bioactivity, Functionality, and Applications. Crit. Rev. Food Sci. Nutr. 2018, 58, 3097–3129. [Google Scholar] [CrossRef] [PubMed]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current Trends and Perspectives of Bioactive Peptides. Crit. Rev. Food Sci. Nutr. 2018, 58, 2273–2284. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Sillero, J.; Gharsallaoui, A.; Prentice, C. Peptides from Fish By-product Protein Hydrolysates and Its Functional Properties: An Overview. Mar. Biotechnol. 2018, 20, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Tonolo, F.; Moretto, L.; Grinzato, A.; Fiorese, F.; Folda, A.; Scalcon, V.; Ferro, S.; Arrigoni, G.; Bellamio, M.; Feller, E. Fermented Soy-Derived Bioactive Peptides Selected by a Molecular Docking Approach Show Antioxidant Properties Involving the Keap1/Nrf2 Pathway. Antioxidants 2020, 9, 1306. [Google Scholar] [CrossRef] [PubMed]
- Puangphet, A.; Tiyaboonchai, W.; Thongsook, T. Inhibitory Effect of Sericin Hydrolysate on Polyphenol Oxidase and Browning of Fresh-Cut Products. Int. Food Res. J. 2015, 22, 1623–1630. [Google Scholar]
- Thaha, A.; Wang, B.S.; Chang, Y.W.; Hsia, S.M.; Huang, T.C.; Shiau, C.Y.; Hwang, D.F.; Chen, T.Y. Food-Derived Bioactive Peptides with Antioxidative Capacity, Xanthine Oxidase and Tyrosinase Inhibitory Activity. Processes 2021, 9, 747. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Q.; Lu, Y.; Li, Y.; Li, T.; Zhou, B.; Qiao, L. Effect of Purslane (Portulaca oleracea L.) Extract on Anti-Browning of Fresh-Cut Potato Slices During Storage. Food Chem. 2019, 283, 445–453. [Google Scholar] [CrossRef]
- da Silva, J.D.F.; Correa, A.P.F.; Kechinski, C.P.; Brandelli, A. Buffalo Cheese Whey Hydrolyzed with Alcalase As An Antibrowning Agent in Minimally Processed Apple. J. Food Sci. Technol. 2018, 55, 3731–3738. [Google Scholar] [CrossRef]
- Dávila, I.; Robles, E.; Egüés, I.; Labidi, J.; Gullón, P. The Biorefinery Concept for The Industrial Valorization of Grape Processing By-Products. In Handbook of Grape Processing By-Products; Elsevier: Amsterdam, The Netherlands, 2017; pp. 29–53. [Google Scholar]
- Tinello, F.; Lante, A. Evaluation of Antibrowning and Antioxidant Activities in Unripe Grapes Recovered During Bunch Thinning. Aust. J. Grape Wine. Res. 2017, 23, 33–41. [Google Scholar] [CrossRef]
- Kozlu, A.; Elmaci, Y. Quince Seed Mucilage as Edible Coating for Mandarin Fruit; Determination of The Quality Characteristics During Storage. J. Food Process. Preserv. 2020, 44, e14854. [Google Scholar] [CrossRef]
- Moualek, I.; Aiche, G.I.; Guechaoui, N.M.; Lahcene, S.; Houali, K. Antioxidant and Anti-Inflammatory Activities of Arbutus Unedo Aqueous Extract. Asian. Pac. J. Trop. Biomed. 2016, 6, 937–944. [Google Scholar] [CrossRef]
- Supapvanich, S.; Yimpong, A.; Srisuwanwichan, J. Browning Inhibition on Fresh-Cut Apple by The Immersion of Liquid Endosperm from Mature Coconuts. J. Food Sci. Technol. 2020, 57, 4424–4431. [Google Scholar] [CrossRef]
- Supapvanich, S.; Anartnet, D.; Krungpree, C. Efficiency of Coconut Water Immersion Inhibiting Browning Incidence on Cut-Surface of Fresh-Cut ‘Gala’apples During Storage. In Proceedings of the 4th International Conference on Engineering, Applied Sciences and Technology (ICEAST 2018), Phuket, Thailand, 4–7 July 2018. [Google Scholar]
- Schulz, M.; Chim, J.F. Nutritional and Bioactive Value of Rubus Berries. Food. Biosci. 2019, 31, 100438. [Google Scholar] [CrossRef]
- Wen, P.; Hu, T.G.; Linhardt, R.J.; Liao, S.T.; Wu, H.; Zou, Y.X. Mulberry: A review of Bioactive Compounds and Advanced Processing Technology. Trends Food Sci. Technol. 2019, 83, 138–158. [Google Scholar] [CrossRef]
- Chang, L.W.; Juang, L.J.; Wang, B.S.; Wang, M.Y.; Tai, H.M.; Hung, W.J.; Chen, Y.J.; Huang, M.H. Antioxidant and Antityrosinase Activity of Mulberry (Morus alba L.) Twigs and Root Bark. Food Chem. Toxicol. 2011, 49, 785–790. [Google Scholar] [CrossRef]
- Zhang, L.; Tao, G.; Chen, J.; Zheng, Z.P. Characterization of A New Flavone and Tyrosinase Inhibition Constituents from The Twigs of Morus alba L. Molecules 2016, 21, 1130. [Google Scholar] [CrossRef]
- Kandylis, P.; Kokkinomagoulos, E. Food Applications and Potential Health Benefits of Pomegranate and its Derivatives. Foods 2020, 9, 122. [Google Scholar] [CrossRef]
- El Barnossi, A.; Moussaid, F.; Iraqi Housseini, A. Tangerine, Banana and Pomegranate Peels Valorisation for Sustainable Environment: A Review. Biotechnol. Rep. 2021, 29, e00574. [Google Scholar] [CrossRef]
- Derakhshan, Z.; Ferrante, M.; Tadi, M.; Ansari, F.; Heydari, A.; Hosseini, M.S.; Conti, G.O.; Sadrabad, E.K. Antioxidant Activity and Total Phenolic Content of Ethanolic Extract of Pomegranate Peels, Juice and Seeds. Food Chem. Toxicol. 2018, 114, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Turrini, F.; Malaspina, P.; Giordani, P.; Catena, S.; Zunin, P.; Boggia, R. Traditional Decoction and PUAE Aqueous Extracts of Pomegranate Peels as Potential Low-Cost Anti-Tyrosinase Ingredients. Appl. Sci. 2020, 10, 2795. [Google Scholar] [CrossRef]
- PA Silva, Y.; Borba, B.C.; Pereira, V.A.; Reis, M.G.; Caliari, M.; Brooks, M.S.-L.; Ferreira, T.A. Characterization of Tomato Processing By-Product for Use As A Potential Functional Food Ingredient: Nutritional Composition, Antioxidant Activity and Bioactive Compounds. Int. J. Food Sci. Nutr. 2019, 70, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Ruales, J.; Baenas, N.; Moreno, D.A.; Stinco, C.M.; Meléndez-Martínez, A.J.; García-Ruiz, A. Biological Active Ecuadorian Mango ‘Tommy Atkins’ Ingredients—An Opportunity to Reduce Agrowaste. Nutrients 2018, 10, 1138. [Google Scholar] [CrossRef] [PubMed]
- Jirasuteeruk, C.; Theerakulkait, C. Inhibitory Effect of Different Varieties of Mango Peel Extract on Enzymatic Browning in Potato Puree. Agric. Nat. Resour. 2020, 54, 217–222. [Google Scholar] [CrossRef]
- Rojas-Bravo, M.; Rojas-Zenteno, E.G.; Hernandez-Carranza, P.; Avila-Sosa, R.; Aguilar-Sanchez, R.; Ruiz-Lopez, I.I.; Ochoa-Velasco, C.E. A Potential Application of Mango (Mangifera indica L. cv Manila) Peel Powder to Increase the Total Phenolic Compounds and Antioxidant Capacity of Edible Films and Coatings. Food Bioproc. Tech. 2019, 12, 1584–1592. [Google Scholar] [CrossRef]
- Jirasuteeruk, C.; Theerakulkait, C. Ultrasound-Assisted Extraction of Phenolic Compounds from Mango (Mangifera indica cv. Chok Anan) Peel and Its Inhibitory Effect on Enzymatic Browning of Potato Puree. Food Technol. Biotechnol. 2019, 57, 350–357. [Google Scholar] [CrossRef]
- Chaisakdanugull, C.; Theerakulkait, C.; Wrolstad, R.E. Pineapple Juice and Its Fractions in Enzymatic Browning Inhibition of Banana [Musa (AAA Group) Gros Michel]. J. Agric. Food Chem. 2007, 55, 4252–4257. [Google Scholar] [CrossRef]
- Supapvanich, S.; Mitrsang, P.; Srinorkham, P. Effects of “Queen” and “Smooth Cayenne” Pineapple Fruit Core Extracts on Browning Inhibition of Fresh-Cut Wax Apple Fruit During Storage. Int. Food Res. J. 2017, 24, 559. [Google Scholar]
- Boonyaritthongchai, P.; Supapvanich, S.; Wongaree, C.; Uthairatanakij, A.; Jitareerat, P.; Pongprasert, N.; Kaewmanee, N. Application of Natural Extracts from Pineapple Juice on Inhibiting Browning Symptom of Fresh-Cut “Nam Dok Mai” Mango. In Proceedings of the IV Asia Symposium on Quality Management in Postharvest Systems, Jeonju, Korea, 12–14 September 2017; pp. 235–240. [Google Scholar]
- Weerawardana, M.B.S.; Thiripuranathar, G.; Paranagama, P.A. Natural Antibrowning Agents against Polyphenol Oxidase Activity in Annona muricata and Musa acuminata. J. Chem. 2020, 2020, 1904798. [Google Scholar] [CrossRef]
- Venturi, F.; Bartolini, S.; Sanmartin, C.; Orlando, M.; Taglieri, I.; Macaluso, M.; Lucchesini, M.; Trivellini, A.; Zinnai, A.; Mensuali, A. Potato Peels as a Source of Novel Green Extracts Suitable as Antioxidant Additives for Fresh-Cut Fruits. Appl. Sci. 2019, 9, 2431. [Google Scholar] [CrossRef]
- Bustos, M.C.; Mazzobre, M.F.; Buera, M.P. Stabilization of Refrigerated Avocado Pulp: Chemometrics-Assessed Antibrowning Allium and Brassica Extracts as Effective Lipid Oxidation Retardants. Food Bioproc. Tech. 2017, 10, 1142–1153. [Google Scholar] [CrossRef]
- Lee, B.; Seo, J.D.; Rhee, J.K.; Kim, C.Y. Heated Apple Juice Supplemented with Onion Has Greatly Improved Nutritional Quality and Browning Index. Food Chem. 2016, 201, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Mercimek, H.A.; Guzeldag, G.; Ucan, F.; Guler, K.C.; Karaman, M.; Karayilan, R. Inhibition of Polyphenol Oxidase Purified from Potato (Solanum tuberosum). Rom. Biotechnol. Lett. 2015, 20, 10961–10968. [Google Scholar]
- Lim, W.Y.; Cheun, C.F.; Wong, C.W. Inhibition of Enzymatic Browning in Sweet Potato (Ipomoea batatas (L.)) with Chemical and Natural Anti-Browning Agents. J. Food Process. Preserv. 2019, 43, e14195. [Google Scholar] [CrossRef]
- Liu, X.L.; Chen, T.; Wang, Q.; Liu, J.A.; Lu, Y.H.; Shi, Y. Structure Analysis and Study of Biological Activities of Condensed Tannins From Bruguiera Gymnorhiza (L.) Lam and Their Effect on Fresh-Cut Lotus Roots. Molecules 2021, 26, 1369. [Google Scholar] [CrossRef] [PubMed]
- Tanhaş, E.; Martin, E.; Korucu, E.N.; Dirmenci, T. Effect of Aqueous Extract, Hydrosol, and Essential Oil Forms of Some Endemic Origanum L.(Lamiaceae) Taxa on Polyphenol Oxidase Activity in Fresh-Cut Mushroom Samples. J. Food Process. Preserv. 2020, 44, e14726. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, L.; Miao, J.Y.; Yu, W.Z.; Zou, L.Q.; Zhou, W.; Liu, W. Effect of Cinnamon Essential Oil Nanoemulsion Combined with Ascorbic Acid on Enzymatic Browning of Cloudy Apple Juice. Food Bioprocess. Technol. 2020, 13, 860–870. [Google Scholar] [CrossRef]
- Zhu, Y.; Du, X.; Zheng, J.; Wang, T.; You, X.; Liu, H.; Liu, X. The Effect of Ultrasonic on Reducing Anti-Browning Minimum Effective Concentration of Purslane Extract on Fresh-Cut Potato Slices During Storage. Food Chem. 2021, 343, 128401. [Google Scholar] [CrossRef]
- Klimczak, I.; Gliszczyńska-Świgło, A. Green Tea Extract As An Anti-Browning Agent for Cloudy Apple Juice. J. Sci. Food Agric. 2017, 97, 1420–1426. [Google Scholar] [CrossRef]
- Chang, M.S.; Kim, G.H. Effects of a Combined Treatment of Hot Water with Green Tea Extract and NaCl on the Postharvest Quality of Fresh-cut Burdocks. Korean. J. Hortic. Sci. Technol. 2015, 33, 364–374. [Google Scholar] [CrossRef]
- Supapvanich, S.; Mitrsang, P.; Srinorkham, P.; Boonyaritthongchai, P.; Wongs-Aree, C. Effects of Fresh Aloe Vera Gel Coating on Browning Alleviation of Fresh Cut Wax Apple (Syzygium samarangenese) Fruit cv. Taaptimjaan. J. Food Sci. Technol. 2016, 53, 2844–2850. [Google Scholar] [CrossRef]
- Criado, M.N.; Civera, M.; Martiez, A.; Rodrigo, D. Use of Weibull Distribution to Quantify The Antioxidant Effect of Stevia Rebaudiana on Oxidative Enzymes. LWT Food Sci. Technol. 2015, 60, 985–989. [Google Scholar] [CrossRef]
- Liu, X.; Lu, Y.Z.; Yang, Q.; Yang, H.Y.; Li, Y.; Zhou, B.Y.; Li, T.T.; Gao, Y.; Qiao, L.P. Cod Peptides Inhibit Browning in Fresh-Cut Potato Slices: A Potential Anti-Browning Agent of Random Peptides for Regulating Food Properties. Postharvest Biol. Technol. 2018, 146, 36–42. [Google Scholar] [CrossRef]
- Schulbach, K.F.; Johnson, J.V.; Simonne, A.H.; Kim, J.M.; Jeong, Y.; Yagiz, Y.; Marshall, M.R. Polyphenol Oxidase Inhibitor from Blue Mussel (Mytilus edulis) Extract. J. Food Sci. 2013, 78, C425–C431. [Google Scholar] [CrossRef] [PubMed]
- Ezeonu, C.S. Impact of Garlic, White Onion and Ginger Extracts on Enzymatic Browning in Sweet Potato. JPAS 2021, 21, 20–27. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mandal, N.; Dey, A.; Mondal, B. An Approach Towards Optimization of The Extraction of Polyphenolic Antioxidants from Ginger (Zingiber officinale). J. Food Sci. Technol. 2014, 51, 3301–3308. [Google Scholar] [CrossRef]
- Sampaio, S.L.; Petropoulos, S.A.; Alexopoulos, A.; Heleno, S.A.; Santos-Buelga, C.; Barros, L.; Ferreira, I.C.F.R. Potato Peels as Sources of Functional Compounds for The Food Industry: A Review. Trends Food Sci. Technol. 2020, 103, 118–129. [Google Scholar] [CrossRef]
- Choi, S.H.; Kozukue, N.; Kim, H.J.; Friedman, M. Analysis of Protein Amino Acids, Non-Protein Amino Acids and Metabolites, Dietary Protein, Glucose, Fructose, Sucrose, Phenolic, and Flavonoid Content and Antioxidative Properties of Potato Tubers, Peels, and Cortexes (Pulps). J. Food. Compost. Anal. 2016, 50, 77–87. [Google Scholar] [CrossRef]
- Tinello, F.; Mihaylova, D.; Lante, A. Valorization of Onion Extracts as Anti-Browning Agents. Food Sci. Appl. Biotechnol. 2020, 3, 16–21. [Google Scholar] [CrossRef]
- Golder, M.; Sadhu, S.K.; Biswas, B.; Islam, T. Comparative Pharmacologic Profiles of Leaves and Hypocotyls of A Mangrove Plant: Bruguiera gymnorrhiza. Adv. Tradit.Med. 2020, 20, 395–403. [Google Scholar] [CrossRef]
- Romani, V.P.; Prentice-Hernandez, C.; Martins, V.G. Active and Sustainable Materials from Rice Starch, Fish Protein and Oregano Essential Oil for Food Packaging. Ind. Crops Prod. 2017, 97, 268–274. [Google Scholar] [CrossRef]
- Kosakowska, O.; Węglarz, Z.; Bączek, K. Yield and Quality of “Greek oregano” (Origanum vulgare L. subsp. hirtum) Herb from Organic Production System in Temperate Climate. Ind. Crops Prod. 2019, 141, 111782. [Google Scholar] [CrossRef]
- Kumoro, A.; Wardhani, D.; Retnowati, D.; Haryani, K. A Brief Review on The Characteristics, Extraction and Potential Industrial Applications of Citronella Grass (Cymbopogon nardus) And Lemongrass (Cymbopogon citratus) Essential Oils. In Proceedings of the International Conference on Chemical and Material Engineering (ICCME 2020), Kyoto, Japan, 9–12 October 2020; p. 012118. [Google Scholar]
- Lante, A.; Tinello, F. Citrus Hydrosols As Useful By-Products for Tyrosinase Inhibition. Innov. Food. Sci. Emerg. Technol. 2015, 27, 154–159. [Google Scholar] [CrossRef]
- Rashid, Z.; Khan, M.R.; Mubeen, R.; Hassan, A.; Saeed, F.; Afzaal, M. Exploring the Effect of Cinnamon Essential Oil to Enhance The Stability and Safety of Fresh Apples. J. Food Process. Preserv. 2020, 44, e14926. [Google Scholar] [CrossRef]
- Davila-Rodriguez, M.; Lopez-Malo, A.; Palou, E.; Ramirez-Corona, N.; Jimenez-Munguia, M.T. Antimicrobial Activity of Nanoemulsions of Cinnamon, Rosemary, and Oregano Essential Oils on Fresh Celery. LWT- Food Sci. Technol. 2019, 112, 108247. [Google Scholar] [CrossRef]
- Wan, J.; Zhong, S.; Schwarz, P.; Chen, B.; Rao, J. Physical Properties, Antifungal and Mycotoxin Inhibitory Activities of Five Essential Oil Nanoemulsions: Impact of Oil Compositions and Processing Parameters. Food Chem. 2019, 291, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Chugh, V.; Mishra, V.; Dwivedi, S.; Sharma, K. Purslane (Portulaca oleracea L.): An Underutilized Wonder Plant with Potential Pharmacological Value. Pharm. Innov. J. 2019, 8, 236–246. [Google Scholar]
- Xing, L.; Zhang, H.; Qi, R.; Tsao, R.; Mine, Y. Recent Advances in the Understanding of the Health Benefits and Molecular Mechanisms Associated with Green Tea Polyphenols. J. Agric. Food Chem. 2019, 67, 1029–1043. [Google Scholar] [CrossRef] [PubMed]
- Maan, A.A.; Nazir, A.; Khan, M.K.I.; Ahmad, T.; Zia, R.; Murid, M.; Abrar, M. The Therapeutic Properties and Applications of Aloe Vera: A Review. J. Herb. Med. 2018, 12, 1–10. [Google Scholar] [CrossRef]
- Ahmad, J.; Khan, I.; Blundell, R.; Azzopardi, J.; Mahomoodally, M.F. Stevia rebaudiana Bertoni.: An Updated Review of Its Health Benefits, Industrial Applications and Safety. Trends Food Sci. Technol. 2020, 100, 177–189. [Google Scholar] [CrossRef]
- Tedeschi, T.; Anzani, C.; Ferri, M.; Marzocchi, S.; Caboni, M.F.; Monari, S.; Tassoni, A. Enzymatic Digestion of Calf Fleshing Meat By-Products: Antioxidant and Anti-Tyrosinase Activity of Protein Hydrolysates, and Identification of Fatty Acids. Foods 2021, 10, 755. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.M.; El-Gizawy, A.M.; El-Bassiouny, R.E.I.; Saleh, M.A. The Role of Various Amino Acids in Enzymatic Browning Process in Potato Tubers, and Identifying The Browning Products. Food Chem. 2016, 192, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Metje-Sprink, J.; Menz, J.; Modrzejewski, D.; Sprink, T. DNA-free Genome Editing: Past, Present and Future. Front. Plant Sci. 2019, 9, 1957. [Google Scholar] [CrossRef] [PubMed]
- González, M.N.; Massa, G.A.; Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.S.; Storani, L.; Décima Oneto, C.A.; Hofvander, P.; Feingold, S.E. Reduced Enzymatic Browning in Potato Tubers by Specific Editing of A Polyphenol Oxidase Gene Via Ribonucleoprotein Complexes Delivery of the Crispr/Cas9 System. Front. Plant Sci. 2020, 10, 1649. [Google Scholar] [CrossRef] [PubMed]
- Maioli, A.; Gianoglio, S.; Moglia, A.; Acquadro, A.; Valentino, D.; Milani, A.M.; Prohens, J.; Orzaez, D.; Granell, A.; Lanteri, S.; et al. Simultaneous CRISPR/Cas9 Editing of Three PPO Genes Reduces Fruit Flesh Browning in Solanum melongena L. Front. Plant Sci. 2020, 11, 1883. [Google Scholar] [CrossRef]
- Wan, L.; Wang, Z.; Tang, M.; Hong, D.; Sun, Y.; Ren, J.; Zhang, N.; Zeng, H. CRISPR-Cas9 Gene Editing for Fruit and Vegetable Crops: Strategies and Prospects. Horticulturae 2021, 7, 193. [Google Scholar] [CrossRef]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef]
- Gupta, D.; Bhattacharjee, O.; Mandal, D.; Sen, M.K.; Dey, D.; Dasgupta, A.; Kazi, T.A.; Gupta, R.; Sinharoy, S.; Acharya, K.; et al. CRISPR-Cas9 System: A New-fangled Dawn in Gene Editing. Life Sci. 2019, 232, 116636. [Google Scholar] [CrossRef]
- Feng, Z.; Zhang, B.; Ding, W.; Liu, X.; Yang, D.L.; Wei, P.; Cao, F.; Zhu, S.; Zhang, F.; Mao, Y.; et al. Efficient Genome Editing in Plants Using a CRISPR/Cas System. Cell Res. 2013, 23, 1229–1232. [Google Scholar] [CrossRef]
- Li, J.F.; Norville, J.E.; Aach, J.; McCormack, M.; Zhang, D.; Bush, J.; Church, G.M.; Sheen, J. Multiplex and Homologous Recombination–mediated Genome Editing in Arabidopsis and Nicotiana benthamiana Using Guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.D.; Kamoun, S. Targeted Mutagenesis in the Model Plant Nicotiana Benthamiana Using Cas9 RNA-guided Endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef]
- Kumlehn, J.; Pietralla, J.; Hensel, G.; Pacher, M.; Puchta, H. The CRISPR/Cas Revolution Continues: From Efficient Gene Editing for Crop Breeding to Plant Synthetic Biology. J. Integr. Plant Biol. 2018, 60, 1127–1153. [Google Scholar] [CrossRef] [PubMed]
- Demirer, G.S.; Silva, T.N.; Jackson, C.T.; Thomas, J.B.; Ehrhardt, D.W.; Rhee, S.Y.; Mortimer, J.C.; Landry, M.P. Nanotechnology to Advance CRISPR–Cas Genetic Engineering of Plants. Nat. Nanotechnol. 2021, 16, 243–250. [Google Scholar] [CrossRef]
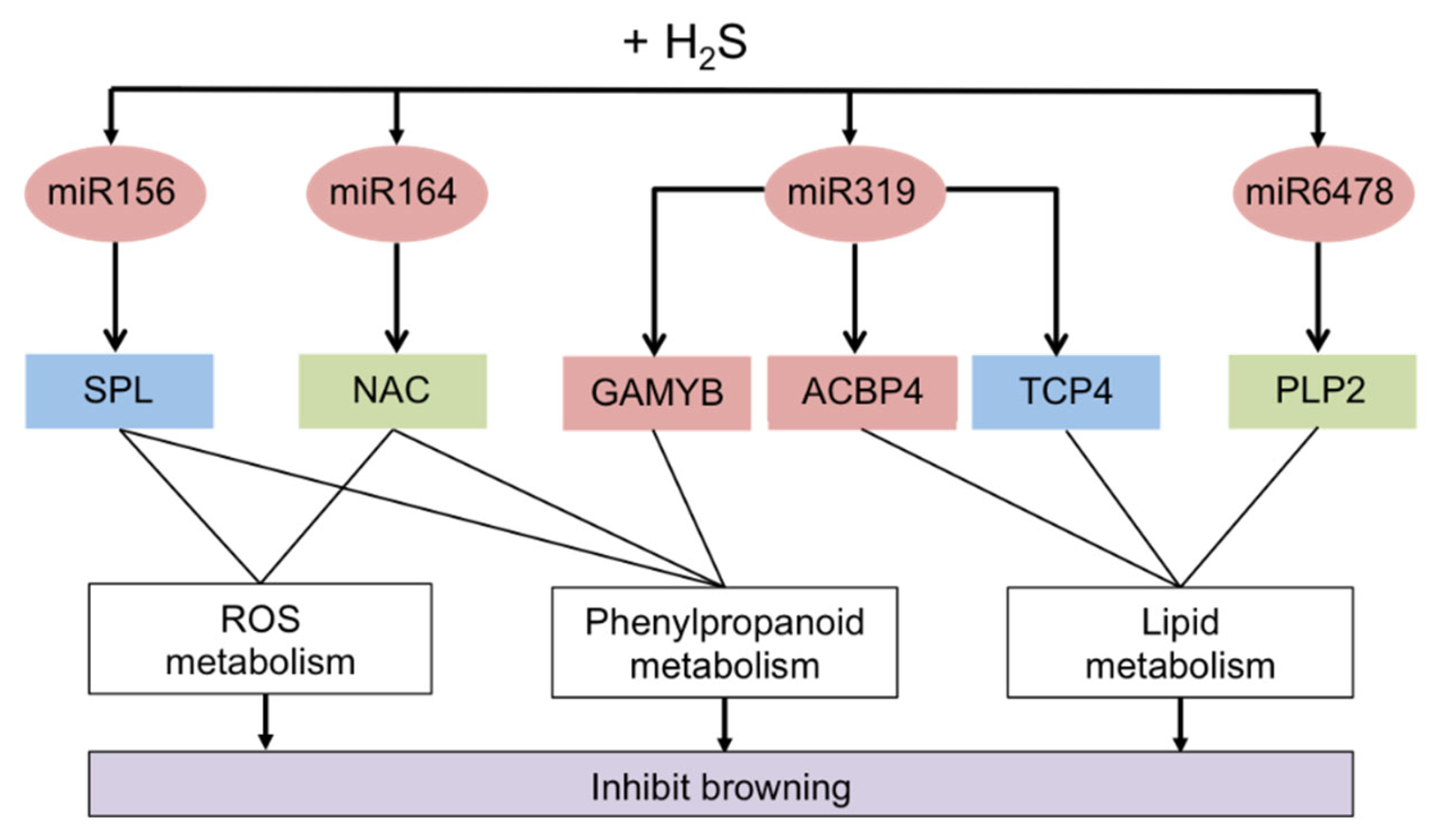
- Chen, C.; Liu, C.; Jiang, A.; Zhao, Q.; Zhang, Y.; Hu, W. miRNA and Degradome Sequencing Identify miRNAs and Their Target Genes Involved in the Browning Inhibition of Fresh-Cut Apples by Hydrogen Sulfide. J. Agric. Food Chem. 2020, 68, 8462–8470. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, Y.; Tang, R.; Qu, H.; Duan, X.; Jiang, Y. Banana sRNAome and Degradome Identify Micrornas Functioning in Differential Responses to Temperature Stress. BMC Genom. 2019, 20, 33. [Google Scholar] [CrossRef]
- Chi, M.; Liu, C.; Su, Y.; Tong, Y.; Liu, H. Bioinformatic Prediction of Upstream microRNAs of PPO and Novel microRNAs in Potato. Can. J. Plant Sci. 2015, 95, 871–877. [Google Scholar] [CrossRef]
- Wang, C.; Ye, J.; Tang, W.; Liu, Z.; Zhu, C.; Wang, M.; Wan, J. Loop Nucleotide Polymorphism in A Putative miRNA Precursor Associated with Seed Length in Rice (Oryza sativa L.). Int. J. Biol. Sci. 2013, 9, 578. [Google Scholar] [CrossRef] [PubMed]
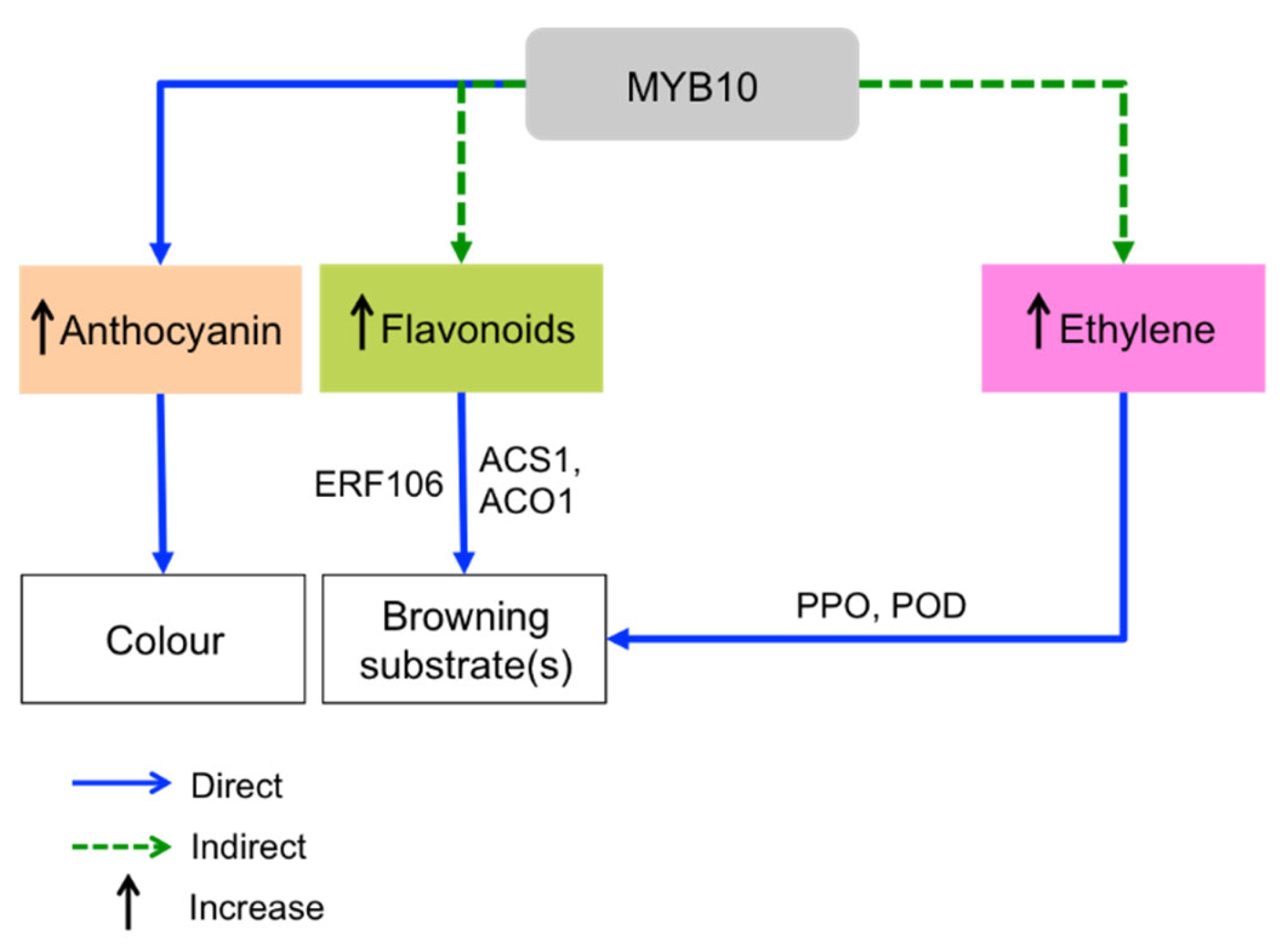
- Espley, R.V.; Leif, D.; Plunkett, B.; McGhie, T.; Henry-Kirk, R.; Hall, M.; Johnston, J.W.; Punter, M.P.; Boldingh, H.; Nardozza, S.; et al. Red to Brown: An Elevated Anthocyanic Response in Apple Drives Ethylene to Advance Maturity and Fruit Flesh Browning. Front. Plant Sci. 2019, 10, 1248. [Google Scholar] [CrossRef]
- Li, R.; Fu, D.; Zhu, B.; Luo, Y.; Zhu, H. CRISPR/Cas9-mediated Mutagenesis of lncRNA1459 Alters Tomato Fruit Ripening. Plant J. 2019, 94, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Vrebalov, J.; Ruezinsky, D.; Padmanabhan, V.; White, R.; Medrano, D.; Drake, R.; Schuch, W.; Giovannoni, J. A MADS-box Gene Necessary for Fruit Ripening at the Tomato Ripening-inhibitor (Rin) Locus. Science 2002, 296, 343–346. [Google Scholar] [CrossRef]
- Giovannoni, J.J. Fruit Ripening Mutants Yield Insights into Ripening Control. Curr. Opin. Plant Biol. 2007, 10, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Osorio, S.; Alba, R.; Damasceno, C.M.; Lopez-Casado, G.; Lohse, M.; Zanor, M.I.; Tohge, T.; Usadel, B.; Rose, J.K.; Fei, Z.; et al. Systems Biology of Tomato Fruit Development: Combined Transcript, Protein, and Metabolite Analysis of Tomato Transcription Factor (Nor, Rin) and Ethylene Receptor (Nr) Mutants Reveals Novel Regulatory Interactions. Plant Physiol. 2011, 157, 405–425. [Google Scholar] [CrossRef]
- Vrebalov, J.; Pan, I.L.; Arroyo, A.J.M.; McQuinn, R.; Chung, M.; Poole, M.; Rose, J.; Seymour, G.; Grandillo, S.; Giovannoni, J.; et al. Fleshy Fruit Expansion and Ripening Are Regulated by the Tomato SHATTERPROOF Gene TAGL1. Plant Cell 2009, 21, 3041–3062. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.; Zhao, F.; Li, R.; Xu, C.; Chen, K.; Xiao, H. The Zinc Finger Transcription Factor SlZFP2 Negatively Regulates Abscisic Acid Biosynthesis and Fruit Ripening in Tomato. Plant Physiol. 2015, 167, 931–949. [Google Scholar] [CrossRef]
- Kim, H.J.; Sujiwo, J.; Kim, H.J.; Jang, A. Effects of Dipping Chicken Breast Meat Inoculated with Listeria monocytogenes in Lyophilized Scallion, Garlic, and Kiwi Extracts on Its Physicochemical Quality. Food Sci. Anim. Resour. 2019, 39, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Khalafalla, F.A.; Ali, F.H.; Hassan, A.-R.H. Quality Improvement and Shelf-Life Extension of Refrigerated Nile Tilapia (Oreochromis niloticus) Fillets Using Natural Herbs. Beni-Seuf Univ. J. Basic Appl. Sci. 2015, 4, 33–40. [Google Scholar] [CrossRef]
- Nicosia, M.G.L.; Pangallo, S.; Raphael, G.; Romeo, F.V.; Strano, M.C.; Rapisarda, P.; Droby, S.; Schena, L. Control of Postharvest Fungal Rots on Citrus Fruit and Sweet Cherries Using A Pomegranate Peel Extract. Postharvest Biol. Technol. 2016, 114, 54–61. [Google Scholar] [CrossRef]
- Ncama, K.; Magwaza, L.S.; Mditshwa, A.; Tesfay, S.Z. Plant-Based Edible Coatings for Managing Postharvest Quality of Fresh Horticultural Produce: A Review. Food. Packag. Shelf Life. 2018, 16, 157–167. [Google Scholar] [CrossRef]
- Yousuf, B.; Srivastava, A.K.; Ahmad, S. Application of Natural Fruit Extract and Hydrocolloid-Based Coating to Retain Quality of Fresh-Cut Melon. J. Food Sci. Technol. 2020, 57, 3647–3658. [Google Scholar] [CrossRef]
- Choulitoudi, E.; Velliopoulou, A.; Tsimogiannis, D.; Oreopoulou, V. Effect of Active Packaging with Satureja Thymbra Extracts on The Oxidative Stability of Fried Potato Chips. Food. Packag. Shelf Life. 2020, 23, 100455. [Google Scholar] [CrossRef]
- Umaraw, P.; Munekata, P.E.S.; Verma, A.K.; Barba, F.J.; Singh, V.P.; Kumar, P.; Lorenzo, J.M. Edible Films/Coating with Tailored Properties for Active Packaging of Meat, Fish and Derived Products. Trends Food Sci. Technol. 2020, 98, 10–24. [Google Scholar] [CrossRef]
- Qiao, L.; Gao, M.; Zheng, J.; Zhang, J.; Lu, L.; Liu, X. Novel Browning Alleviation Technology for Fresh-Cut Products: Preservation Effect of The Combination of Sonchus Oleraceus L. Extract and Ultrasound in Fresh-Cut Potatoes. Food Chem. 2021, 348, 129132. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Zhang, J.; Zhang, H.; Dzah, C.S.; Zandile, M.; Duan, Y.; Ma, H.; Luo, X. Advances in Ultrasound Assisted Extraction of Bioactive Compounds from Cash Crops–A Review. Ultrason. Sonochem. 2018, 48, 538–549. [Google Scholar] [CrossRef]
- Lee, K.; Jeong, Y.; Yoon, B. Developing An Research and Development (R&D) Process Improvement System to Simulate The Performance of R&D Activities. Comput. Ind. 2017, 92, 178–193. [Google Scholar] [CrossRef]







| Patent Number | Approval Year | Country | Natural Sources | Patent Title | Summary of Inventions |
|---|---|---|---|---|---|
| CN111066878A | 2020 | China | Fruit and vegetable | Natural fruit and vegetable preservative and preparation process thereof | This patent related the natural fruit and vegetable extract as a potential fresh-keeping agent. |
| US6224926B1 | 2019 | United States | Pineapple | Natural anti-browning and antioxidant compositions and methods for making the same | This patent describes active ingredients containing pineapple juice as a browning and oxidizing inhibitor. |
| AU2017260170A1 | 2017 | Australia | Acerola cherry | Juice products and methods for reduced enzymatic browning | This patent is related to the anti-browning potentials of acerola cherry fruit to reduce the oxidation process produced by PPO. |
| US20160338368A1 | 2016 | United States | Green tea | Fruit and vegetable preservative | This invention describes the preservation of fruits and vegetables using green tea extract. |
| US20140127369A1 | 2014 | United States | Chrysanthemum indicum | Composition containing Chrysanthemum indicum L. extract for preventing discoloration | This patent is related to anti-browning agents containing the extracts of Chrysanthemum indicum for preventing browning of fruits and vegetables. |
| KR20120135778A | 2011 | South Korea | Citrus peel | Anti-browning agents containing the extracts of pericarp of Citrus linn | This invention relates to the utilization of citrus-peel extract for browning prevention. |
| KR101300603B1 | 2011 | South Korea | Chrysanthemum indicum | Anti-browning agents containing the extracts of Chrysanthemum indicum | This invention relates to browning prevention by using the active ingredient extract from Chrysanthemum indicum. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdan, N.; Lee, C.H.; Wong, S.L.; Fauzi, C.E.N.C.A.; Zamri, N.M.A.; Lee, T.H. Prevention of Enzymatic Browning by Natural Extracts and Genome-Editing: A Review on Recent Progress. Molecules 2022, 27, 1101. https://doi.org/10.3390/molecules27031101
Hamdan N, Lee CH, Wong SL, Fauzi CENCA, Zamri NMA, Lee TH. Prevention of Enzymatic Browning by Natural Extracts and Genome-Editing: A Review on Recent Progress. Molecules. 2022; 27(3):1101. https://doi.org/10.3390/molecules27031101
Chicago/Turabian StyleHamdan, Norfadilah, Chia Hau Lee, Syie Luing Wong, Che Ellysa Nurshafika Che Ahmad Fauzi, Nur Mirza Aqilah Zamri, and Ting Hun Lee. 2022. "Prevention of Enzymatic Browning by Natural Extracts and Genome-Editing: A Review on Recent Progress" Molecules 27, no. 3: 1101. https://doi.org/10.3390/molecules27031101
APA StyleHamdan, N., Lee, C. H., Wong, S. L., Fauzi, C. E. N. C. A., Zamri, N. M. A., & Lee, T. H. (2022). Prevention of Enzymatic Browning by Natural Extracts and Genome-Editing: A Review on Recent Progress. Molecules, 27(3), 1101. https://doi.org/10.3390/molecules27031101