
Towards Better Sinomenine-Type Drugs to Treat Rheumatoid Arthritis: Molecular Mechanisms and Structural Modification
 ,
,
Abstract
1. Introduction
2. Direct Targets of Sinomenine for Treating RA
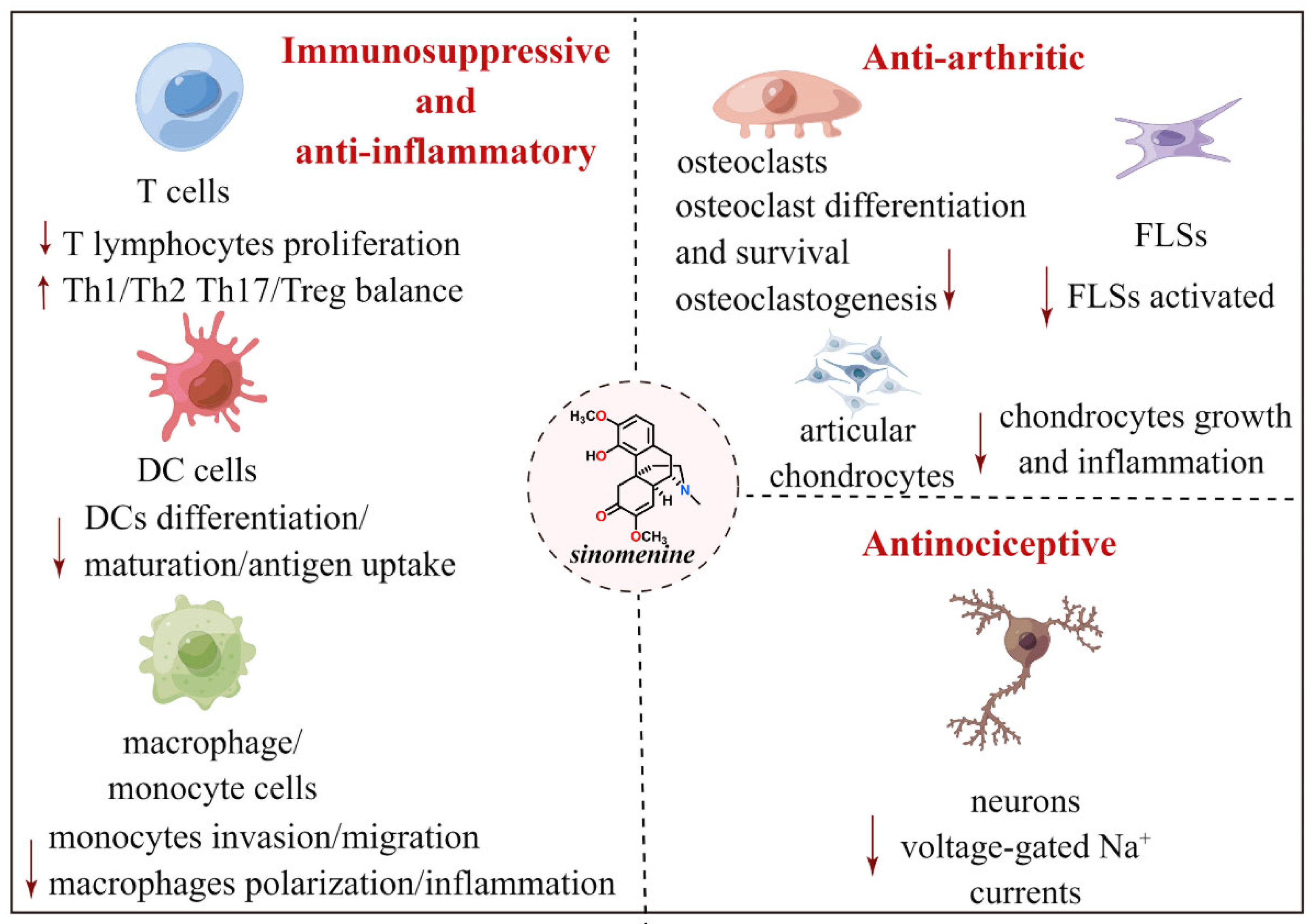
3. Immunosuppressive and Anti-Inflammatory Effects of Sinomenine
4. Antiarthritic Effect of Sinomenine
5. Antinociceptive Effect of Sinomenine
6. Novel Sinomenine-Derived Anti-RA Agents
6.1. Sinomenine Derivatives with Enhanced Anti-RA Effects
6.2. SAR Analysis of Sinomenine Derivatives for RA Treatment
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sparks, J.A. Rheumatoid arthritis. Ann. Intern. Med. 2019, 170, ITC1–ITC16. [Google Scholar] [CrossRef] [PubMed]
- Aletaha, D.; Smolen, J.S. Diagnosis and management of rheumatoid arthritis: A review. JAMA 2018, 320, 1360–1372. [Google Scholar] [CrossRef] [PubMed]
- Weyand, C.M.; Goronzy, J.J. The immunology of rheumatoid arthritis. Nat. Immunol. 2021, 22, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, A.; Swain, N.; Gupta, B. Understanding the role and uses of alternative therapies for the management of rheumatoid arthritis. Curr. Rheumatol. Rev. 2022, 18, 89–100. [Google Scholar] [CrossRef]
- Zhao, X.X.; Peng, C.; Zhang, H.; Qin, L.P. Sinomenium acutum: A review of chemistry, pharmacology, pharmacokinetics, and clinical use. Pharm. Biol. 2012, 50, 1053–1061. [Google Scholar] [CrossRef]
- Ishiwari, N. An alkaloid of Sinomenium diversifolius. Chugai Iji Shimpo 1920, 959, 277–296. [Google Scholar]
- Zhang, M.W.; Wang, X.H.; Shi, J.; Yu, J.G. Sinomenine in cardio-cerebrovascular diseases: Potential therapeutic effects and pharmacological evidences. Front. Cardiovasc. Med. 2021, 8, 749113. [Google Scholar] [CrossRef]
- Jiang, W.; Fan, W.; Gao, T.; Li, T.; Yin, Z.; Guo, H.; Wang, L.; Han, Y.; Jiang, J.D. Analgesic mechanism of sinomenine against chronic pain. Pain Res. Manag. 2020, 2020, 1876862. [Google Scholar] [CrossRef]
- Huang, R.Y.; Pan, H.D.; Wu, J.Q.; Zhou, H.; Li, Z.G.; Qiu, P.; Zhou, Y.Y.; Chen, X.M.; Xie, Z.X.; Xiao, Y.; et al. Comparison of combination therapy with methotrexate and sinomenine or leflunomide for active rheumatoid arthritis: A randomized controlled clinical trial. Phytomedicine 2019, 57, 403–410. [Google Scholar] [CrossRef]
- Wang, Q.; Li, X.K. Immunosuppressive and anti-inflammatory activities of sinomenine. Int. Immunopharmacol. 2011, 11, 373–376. [Google Scholar] [CrossRef]
- Tang, J.; Raza, A.; Chen, J.; Xu, H. A systematic review on the sinomenine derivatives. Mini-Rev. Med. Chem. 2018, 18, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ji, J.; Feng, Z.; Hou, X.; Luo, Y.; Mei, Z. A network pharmacology approach to explore the potential targets underlying the effect of sinomenine on rheumatoid arthritis. Int. Immunopharmacol. 2020, 80, 106201. [Google Scholar] [CrossRef] [PubMed]
- Kurko, J.; Besenyei, T.; Laki, J.; Glant, T.T.; Mikecz, K.; Szekanecz, Z. Genetics of rheumatoid arthritis-A comprehensive review. Clin. Rev. Allergy Immunol. 2013, 45, 170–179. [Google Scholar] [CrossRef]
- Marquez, A.; Martin, J.; Carmona, F.D. Emerging aspects of molecular biomarkers for diagnosis, prognosis and treatment response in rheumatoid arthritis. Expert Rev. Mol. Diagn. 2016, 16, 663–675. [Google Scholar] [CrossRef]
- Hoppe, B.; Dorner, T. Coagulation and the fibrin network in rheumatic disease: A role beyond haemostasis. Nat. Rev. Rheumatol. 2012, 8, 738–746. [Google Scholar] [CrossRef]
- Li, G.Q.; Fang, Y.X.; Liu, Y.; Meng, F.R.; Wu, X.; Zhang, C.W.; Zhang, Y.; Liu, D.; Gao, B. MALAT1-driven inhibition of Wnt signal impedes proliferation and inflammation in fibroblast-Like synoviocytes through CTNNB1 promoter methylation in rheumatoid arthritis. Hum. Gene Ther. 2019, 30, 1008–1022. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, H.J.; Ji, T.F.; Zhang, C.J. Chemoproteomics-based target profiling of sinomenine reveals multiple protein regulators of inflammation. Chem. Commun. 2021, 57, 5981–5984. [Google Scholar] [CrossRef]
- Nathan, C. Points of control in inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef]
- Liu, L.; Resch, K.; Kaever, V. Inhibition of lymphocyte proliferation by the anti-arthritic drug sinomenine. Int. J. Immunopharmacol. 1994, 16, 685–691. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, J.; Hou, W.; Wang, D.; Li, F.; Zhang, Y.; Yuan, F. Immunoregulatory effects of sinomenine on the T-bet/GATA-3 ratio and Th1/Th2 cytokine balance in the treatment of mesangial proliferative nephritis. Int. Immunopharmacol. 2009, 9, 894–899. [Google Scholar] [CrossRef]
- Shu, L.; Yin, W.; Zhang, J.; Tang, B.; Kang, Y.X.; Ding, F.; Hua, Z.C. Sinomenine inhibits primary CD4+ T-cell proliferation via apoptosis. Cell Biol. Int. 2007, 31, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, C.; Jin, N.; Xie, Z.; Fei, L.; Jia, Z.; Wu, Y. Sinomenine promotes differentiation but impedes maturation and co-stimulatory molecule expression of human monocyte-derived dendritic cells. Int. Immunopharmacol. 2007, 7, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, J.; Yu, K.; Liu, Y.; Chen, X. Sinomenine inhibits maturation of monocyte-derived dendritic cells through blocking activation of NF-kappa B. Int. Immunopharmacol. 2007, 7, 637–645. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, J.; Guo, Z.; Liu, Q.; Chen, T.; Wang, X.; Cao, X. Requirement for ERK activation in sinomenine-induced apoptosis of macrophages. Immunol. Lett. 2005, 98, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.Q.; Chen, L.H.; Li, X.J.; Lin, Z.B.; Li, W.D. Sinomenine influences capacity for invasion and migration in activated human monocytic THP-1 cells by inhibiting the expression of MMP-2, MMP-9, and CD147. Acta Pharmacol. Sin. 2009, 30, 435–441. [Google Scholar] [CrossRef]
- Tong, B.; Yu, J.; Wang, T.; Dou, Y.; Wu, X.; Kong, L.; Dai, Y.; Xia, Y. Sinomenine suppresses collagen-induced arthritis by reciprocal modulation of regulatory T cells and Th17 cells in gut-associated lymphoid tissues. Mol. Immunol. 2015, 65, 94–103. [Google Scholar] [CrossRef]
- Tong, B.; Yuan, X.; Dou, Y.; Wu, X.; Wang, Y.; Xia, Y.; Dai, Y. Sinomenine induces the generation of intestinal Treg cells and attenuates arthritis via activation of aryl hydrocarbon receptor. Lab. Investig. 2016, 96, 1076–1086. [Google Scholar] [CrossRef]
- Jegerschold, C.; Pawelzik, S.C.; Purhonen, P.; Bhakat, P.; Gheorghe, K.R.; Gyobu, N.; Mitsuoka, K.; Morgenstern, R.; Jakobsson, P.J.; Hebert, H. Structural basis for induced formation of the inflammatory mediator prostaglandin E2. Proc. Natl. Acad. Sci. USA. 2008, 105, 11110–11115. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, J.X.; Luo, J.F.; Cheng, C.S.; Leung, E.L.; Li, Y.; Su, X.H.; Liu, Z.Q.; Chen, T.B.; Duan, F.G.; et al. Suppressing mPGES-1 expression by sinomenine ameliorates inflammation and arthritis. Biochem. Pharmacol. 2017, 142, 133–144. [Google Scholar] [CrossRef]
- Luo, J.F.; Yao, Y.D.; Cheng, C.S.; Lio, C.K.; Liu, J.X.; Huang, Y.F.; He, F.; Xie, Y.; Liu, L.; Liu, Z.Q.; et al. Sinomenine increases the methylation level at specific GCG site in mPGES-1 promoter to facilitate its specific inhibitory effect on mPGES-1. Biochim. Biophys. Acta Gene Regul. Mech. 2022, 1865, 194813. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Zhu, W.; Ma, C.; Ruan, J.; Long, H.; Wang, Y. Sinomenine inhibits the progression of rheumatoid arthritis by regulating the secretion of inflammatory cytokines and monocyte/macrophage subsets. Front. Immunol. 2018, 9, 2228. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Zhang, X.; Dou, Y.; Wei, Z.; Tao, Y.; Xia, Y.; Dai, Y. Gut-sourced vasoactive intestinal polypeptide induced by the activation of alpha7 nicotinic acetylcholine receptor substantially contributes to the anti-inflammatory effect of sinomenine in collagen-induced arthritis. Front. Pharmacol. 2018, 9, 675. [Google Scholar] [CrossRef] [PubMed]
- Zhi, Y.K.; Li, J.; Yi, L.; Zhu, R.L.; Luo, J.F.; Shi, Q.P.; Bai, S.S.; Li, Y.W.; Du, Q.; Cai, J.Z.; et al. Sinomenine inhibits macrophage M1 polarization by downregulating alpha7nAChR via a feedback pathway of alpha7nAChR/ERK/Egr-1. Phytomedicine 2022, 100, 154050. [Google Scholar] [CrossRef] [PubMed]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.J.; Liu, J.X.; Xie, Y.; Luo, P.; Liu, Z.Q.; Liu, L.; Zhou, H. Suppression of macrophage migration by down-regulating Src/FAK/P130Cas activation contributed to the anti-inflammatory activity of sinomenine. Pharmacol. Res. 2021, 167, 105513. [Google Scholar] [CrossRef]
- de Punder, Y.M.; van Riel, P.L. Rheumatoid arthritis: Understanding joint damage and physical disability in RA. Nat. Rev. Rheumatol. 2011, 7, 260–261. [Google Scholar] [CrossRef]
- Okamoto, K.; Takayanagi, H. Osteoclasts in arthritis and Th17 cell development. Int. Immunopharmacol. 2011, 11, 543–548. [Google Scholar] [CrossRef]
- He, L.G.; Li, X.L.; Zeng, X.Z.; Duan, H.; Wang, S.; Lei, L.S.; Li, X.J.; Liu, S.W. Sinomenine induces apoptosis in RAW 264.7 cell-derived osteoclasts in vitro via caspase-3 activation. Acta Pharmacol. Sin. 2014, 35, 203–210. [Google Scholar] [CrossRef]
- He, L.; Duan, H.; Li, X.; Wang, S.; Zhang, Y.; Lei, L.; Xu, J.; Liu, S.; Li, X. Sinomenine down-regulates TLR4/TRAF6 expression and attenuates lipopolysaccharide-induced osteoclastogenesis and osteolysis. Eur. J. Pharmacol. 2016, 779, 66–79. [Google Scholar] [CrossRef]
- Zhou, B.; Lu, X.; Tang, Z.; Liu, D.; Zhou, Y.; Zeng, P.; Xiong, H. Influence of sinomenine upon mesenchymal stem cells in osteoclastogenesis. Biomed. Pharmacother. 2017, 90, 835–841. [Google Scholar] [CrossRef]
- Wu, Z.; Ma, D.; Yang, H.; Gao, J.; Zhang, G.; Xu, K.; Zhang, L. Fibroblast-like synoviocytes in rheumatoid arthritis: Surface markers and phenotypes. Int. Immunopharmacol. 2021, 93, 107392. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fang, Y.; Huang, W.; Zhou, X.; Wang, M.; Zhong, B.; Peng, D. Effect of sinomenine on cytokine expression of macrophages and synoviocytes in adjuvant arthritis rats. J. Ethnopharmacol. 2005, 98, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.; Su, X.; Lei, K.; Liu, Z.; Lu, L.; Wu, Q.; Pan, H.; Huang, Q.; Zhao, Y.; Wang, M.; et al. Sinomenine protects bone from destruction to ameliorate arthritis via activating p62(Thr269/Ser272)-Keap1-Nrf2 feedback loop. Biomed. Pharmacother. 2021, 135, 111195. [Google Scholar] [CrossRef]
- Yi, L.; Lyn, Y.J.; Peng, C.; Zhu, R.L.; Bai, S.S.; Liu, L.; Wang, P.X.; Zhou, H.; Dong, Y. Sinomenine inhibits fibroblast-like synoviocyte proliferation by regulating alpha7nAChR expression via ERK/Egr-1 pathway. Int. Immunopharmacol. 2018, 56, 65–70. [Google Scholar] [CrossRef]
- Yi, L.; Ke, J.; Liu, J.; Lai, H.; Lv, Y.; Peng, C.; Zhi, Y.; Du, Q.; Liu, L.; Wang, P.; et al. Sinomenine increases adenosine A2A receptor and inhibits NF-kappaB to inhibit arthritis in adjuvant-induced-arthritis rats and fibroblast-like synoviocytes through alpha7nAChR. J. Leukoc. Biol. 2021, 110, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Pap, T.; Korb-Pap, A. Cartilage damage in osteoarthritis and rheumatoid arthritis--two unequal siblings. Nat. Rev. Rheumatol. 2015, 11, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, C.; Zhang, H. Lipopolysaccharides-mediated injury to chondrogenic ATDC5 cells can be relieved by Sinomenine via downregulating microRNA-192. Phytother. Res. 2019, 33, 1827–1836. [Google Scholar] [CrossRef]
- Wu, Y.; Lin, Z.; Yan, Z.; Wang, Z.; Fu, X.; Yu, K. Sinomenine contributes to the inhibition of the inflammatory response and the improvement of osteoarthritis in mouse-cartilage cells by acting on the Nrf2/HO-1 and NF-kappaB signaling pathways. Int. Immunopharmacol. 2019, 75, 105715. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, L.; Chen, P.; Liang, G. Myeloid differentiation primary response protein 88 (MyD88): The central hub of TLR/IL-1R signaling. J. Med. Chem. 2020, 63, 13316–13329. [Google Scholar] [CrossRef]
- Ramirez-Perez, S.; Oregon-Romero, E.; Reyes-Perez, I.V.; Bhattaram, P. Targeting MyD88 downregulates inflammatory mediators and pathogenic processes in PBMC from DMARDs-naive rheumatoid arthritis patients. Front. Pharmacol. 2021, 12, 800220. [Google Scholar] [CrossRef]
- Mu, H.; Yao, R.B.; Zhao, L.J.; Shen, S.Y.; Zhao, Z.M.; Cai, H. Sinomenine decreases MyD88 expression and improves inflammation-induced joint damage progression and symptoms in rat adjuvant-induced arthritis. Inflammation 2013, 36, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Hao, J.; Wiesenfeld-Hallin, Z.; Wang, D.Q.; Xu, X.J. Analgesic effect of sinomenine in rodents after inflammation and nerve injury. Eur. J. Pharmacol. 2013, 721, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Sun, Y.; Mao, L.; Liu, C.; Jiang, B.; Zhang, W.; Li, J.X. Antinociceptive effects of sinomenine in a rat model of postoperative pain. Br. J. Pharmacol. 2016, 173, 1693–1702. [Google Scholar] [CrossRef]
- Lee, J.Y.; Yoon, S.Y.; Won, J.; Kim, H.B.; Kang, Y.; Oh, S.B. Sinomenine produces peripheral analgesic effects via inhibition of voltage-gated sodium currents. Neuroscience 2017, 358, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.; Coghi, P.; Law, B.; Liu, L.; Wong, V. The present and future synthetic strategies of structural modifications of sinomenine. Org. Chem. Front. 2020, 7, 4089–4107. [Google Scholar] [CrossRef]
- Wu, F.C.; Feng, X.; Wu, K.M.; Cheng, G.F.; Huang, Y.M.; Ye, X.R.; Qiu, P.; Zheng, X.M. A Series of Sinomenine Derivatives and Their Preparation Methods. China Patent CN 101092397, 19 January 2011. [Google Scholar]
- Zhao, Z.; Xiao, J.; Wang, J.; Dong, W.; Peng, Z.; An, D. Anti-inflammatory effects of novel sinomenine derivatives. Int. Immunopharmacol. 2015, 29, 354–360. [Google Scholar] [CrossRef]
- Zhao, Z.J.; Zhao, C.; Xiao, J.; Wang, J.C. Transdermal permeation and anti-inflammation activities of novel sinomenine derivatives. Molecules 2016, 21, 1520. [Google Scholar] [CrossRef]
- Jin, J.; Teng, P.; Liu, H.L.; Wu, J.; Liu, Y.M.; Xu, Q.; Li, J.X. Microfluidics assisted synthesis and bioevaluation of sinomenine derivatives as antiinflammatory agents. Eur. J. Med. Chem. 2013, 62, 280–288. [Google Scholar] [CrossRef]
- Zhou, Z.; Qiu, N.; Ou, Y.; Wei, Q.; Tang, W.; Zheng, M.; Xing, Y.; Li, J.J.; Ling, Y.; Li, J.; et al. N-Demethylsinomenine, an active metabolite of sinomenine, attenuates chronic neuropathic and inflammatory pain in mice. Sci. Rep. 2021, 11, 9300. [Google Scholar] [CrossRef]
- Teng, P.; Liu, H.L.; Zhang, L.; Feng, L.L.; Huai, Y.; Deng, Z.S.; Sun, Y.; Xu, Q.; Li, J.X. Synthesis and biological evaluation of novel sinomenine derivatives as anti-inflammatory agents. Eur. J. Med. Chem. 2012, 50, 63–74. [Google Scholar] [CrossRef]
- Zhou, Y.R.; Zhao, Y.; Bao, B.H.; Li, J.X. SND-117, a sinomenine bivalent alleviates type II collagen-induced arthritis in mice. Int. Immunopharmacol. 2015, 26, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.C.; Bi, E.G.; Lou, Y.T.; Wu, X.D.; Liu, Z.D.; Zou, J.; Wang, Y.; Ma, Z.; Lin, G.M.; Sun, S.H.; et al. Novel sinomenine derivative 1032 improves immune suppression in experimental autoimmune encephalomyelitis. Biochem. Biophys. Res. Commun. 2010, 391, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.T.; Hou, J.; Wang, M.; Ma, L.Y.; Wu, L.L.; Wang, S.Z.; Sun, B.; Yao, Z. Regio-controlled synthesis of unsymmetrical pyrazine-fused sinomenine derivatives and discriminate substitution effects on TNF-α inhibitory activity. Tetrahedron 2014, 70, 5475–5482. [Google Scholar] [CrossRef]
- Ni, P.; Liu, Y.; Man, J.; Li, W.; Xue, S.; Lu, T.; Su, Z.; Zhou, C. C16, a novel sinomenine derivatives, promoted macrophage reprogramming toward M2-like phenotype and protected mice from endotoxemia. Int. J. Immunopath. Ph. 2021, 35, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Dai, Z.; Zhang, T.; Gu, Y.; Cai, D.; Lu, M.; Zhang, Z.; Zeng, Q.; Shang, B.; Xu, B.; et al. Synthesis and biological evaluation of novel sinomenine derivatives as anti-inflammatory and analgesic agent. RSC Adv. 2022, 12, 30001–30007. [Google Scholar] [CrossRef]
- Zhang, T.; Ouyang, X.; Gou, S.; Zhang, Y.; Yan, N.; Chang, L.; Li, B.; Zhang, F.; Liu, H.; Ni, J. Novel synovial targeting peptide-sinomenine conjugates as a potential strategy for the treatment of rheumatoid arthritis. Int. J. Pharm. 2022, 617, 121628. [Google Scholar] [CrossRef]
- Huang, L.; Dong, Y.; Wu, J.; Wang, P.; Zhou, H.; Li, T.; Liu, L. Sinomenine-induced histamine release-like anaphylactoid reactions are blocked by tranilast via inhibiting NF-kappaB signaling. Pharmacol. Res. 2017, 125, 150–160. [Google Scholar] [CrossRef]
- Gao, L.N.; Zhong, B.; Wang, Y. Mechanism underlying antitumor effects of sinomenine. Chin. J. Integr. Med. 2019, 25, 873–878. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
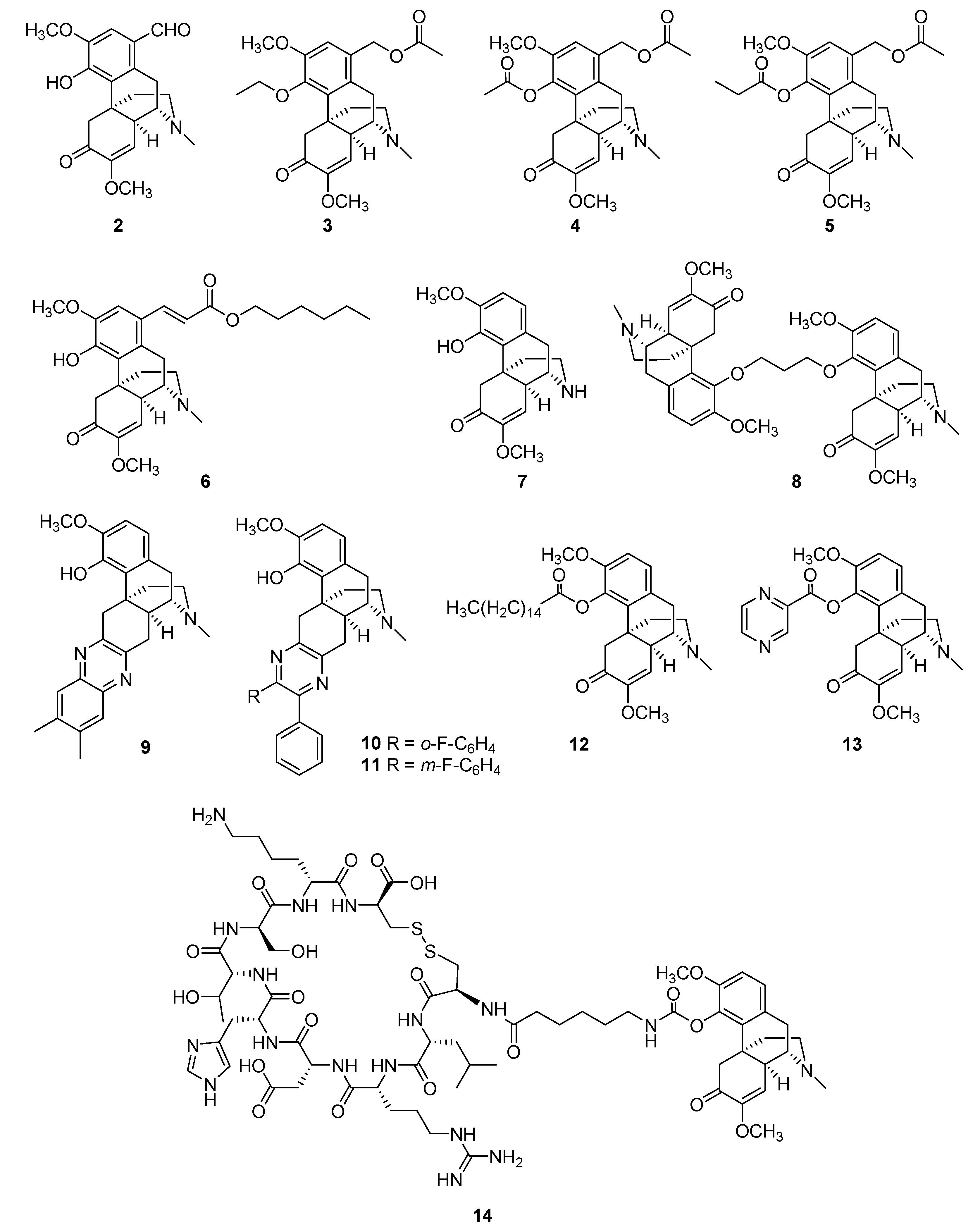
| Compound | Cell Model and Efficacy | Animal Model | Potential Targets or Pathways | Cytotoxicity or Side Effects | Ref. |
|---|---|---|---|---|---|
| 2 | rat splenocytes | croton oil-induced ear edema | IL-2 | — 1 | [56] |
| 3 | LPS-induced RAW264.7 cells | dimethylbenzene-induced ear edema, carrageenan-induced paw edema | IL-1β, TNF-α | — | [57] |
| 4, 5 | LPS-induced RAW264.7 cells | dimethylbenzene-induced ear edema, carrageenan-induced paw edema | IL-1β, TNF-α | no obvious cytotoxicity in peritoneal macrophages at concentrations below 0.02 mg/mL | [58] |
| 6 | TNF-α-induced mouse embryonic fibroblasts | carrageenan-induced paw edema | NF-κB | cytotoxicity in mouse embryonic fibroblasts, IC50 = 38.9 μM | [59] |
| 7 | — | chronic constriction injury model, complete Freund’s adjuvant-induced mechanical allodynia | GABAA receptor | no sedation or allergic reactions at a dose of 80 mg/kg | [60] |
| 8 | LPS-induced RAW264.7 cells, NO production inhibition, IC50 = 15.2 μM (sinomenine concentration >200 μM); fibroblast-like synovial cells (FLSCs) | LPS-induced septic shock model, collagen-induced arthritis | TNF-α, IL-6, NF-κB, IκBα; not the MAPK pathway | cytotoxicity in RAW264.7 cells, IC50 = 41.1 μM (sinomenine, IC50 >200 μM); no inhibition of FLSC growth at 30 μM | [61,62] |
| 9 | Bone-marrow-derived dendritic cells | EAE model | TNF-α, IL-6 | [63] | |
| 10, 11 | LPS-induced peritoneal macrophages, 99% inhibition of TNF-α activity at 10 μM (sinomenine, 11% inhibitory rate) | — | TNF-α | — | [64] |
| 12 | LPS-induced ANA-1/peritoneal macrophages | LPS-induced endotoxemia model | P38/AKT or STAT1 pathways | — | [65] |
| 13 | LPS-induced RAW264.7 cells, NO production inhibition, IC50 = 30.28 ± 1.70 μM (sinomenine, IC50 = 70.86 ± 1.00 μM) | carrageenan-induced paw edema, acetic-acid-induced writhing test | NF-κB, iNOS | no obvious cytotoxicity in RAW264.7 cells | [66] |
| 14 | — | carrageenan-induced paw edema, AIA model | — | no hemolytic activity | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Zhang, S.; Liao, J.; Gong, Z.; Chai, X.; Lyu, H. Towards Better Sinomenine-Type Drugs to Treat Rheumatoid Arthritis: Molecular Mechanisms and Structural Modification. Molecules 2022, 27, 8645. https://doi.org/10.3390/molecules27248645
Zhang C, Zhang S, Liao J, Gong Z, Chai X, Lyu H. Towards Better Sinomenine-Type Drugs to Treat Rheumatoid Arthritis: Molecular Mechanisms and Structural Modification. Molecules. 2022; 27(24):8645. https://doi.org/10.3390/molecules27248645
Chicago/Turabian StyleZhang, Cuili, Shujie Zhang, Jingjing Liao, Zipeng Gong, Xin Chai, and Haining Lyu. 2022. "Towards Better Sinomenine-Type Drugs to Treat Rheumatoid Arthritis: Molecular Mechanisms and Structural Modification" Molecules 27, no. 24: 8645. https://doi.org/10.3390/molecules27248645
APA StyleZhang, C., Zhang, S., Liao, J., Gong, Z., Chai, X., & Lyu, H. (2022). Towards Better Sinomenine-Type Drugs to Treat Rheumatoid Arthritis: Molecular Mechanisms and Structural Modification. Molecules, 27(24), 8645. https://doi.org/10.3390/molecules27248645