
Compensatory Transcriptional Response of Fischerella thermalis to Thermal Damage of the Photosynthetic Electron Transfer Chain

 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Prevalence of the D1 Isoform in the Fischerella thermalis PSII Reaction Center
2.2. Effect of Temperature on the Transcription of Photosynthesis-Related Genes in Fischerella thermalis
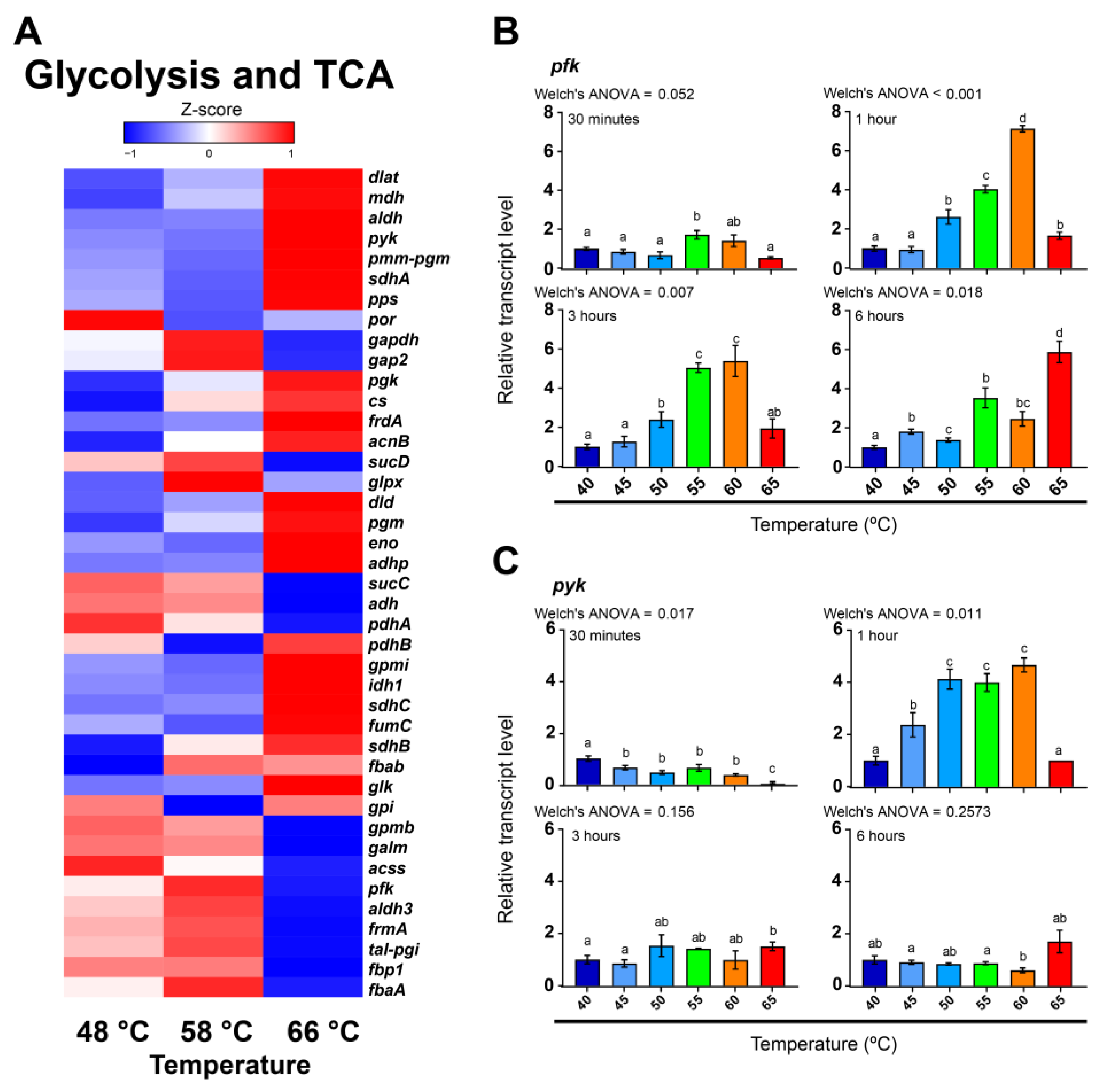
2.3. Alternative Energy Pathways and Co-Expression of Energy Metabolism Genes in Fischerella thermalis
2.4. Effect of Temperature on the Photosynthetic Efficiency of Fischerella thermalis
3. Discussion
3.1. Possible Loss of D1 Gene Copies and Retention of the Photoresistant D1INT Isoform in Fischerella thermalis
3.2. Photosynthesis-Related Genes Are Differentially Regulated by Temperature in Fischerella thermalis
3.3. Emerging Evidence of Mixotrophic Metabolism by Fischerella thermalis in Relation to Its Thermotolerance
4. Materials and Methods
4.1. Phylogenetic Reconstruction and Analysis of the D1 Isoforms of Fischerella thermalis
4.2. Fischerella thermalis PCC 7521 Culture Conditions and Temperature Treatments
4.3. RNA Extraction
4.4. cDNA Synthesis
4.5. qPCR Analysis
4.6. Metatranscriptomic Analysis of Porcelana Hot Spring and El Tatio Environmental Samples
4.7. Co-Expression Network Analysis and Pathway Overrepresentation
4.8. Metabolic Network of Fischerella thermalis Reconstruction
4.9. Chlorophyll Fluorescence Measurement of Temperature-Treated Fischerella thermalis PCC 7521 Cultures under Continuous Light Conditions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- McGowan, H.A.; Lowry, A.L.; Gray, M.A. Identification of Optimum Temperatures for Photosynthetic Production in Subtropical Coastal Ecosystems: Implications for CO2 Sequestration in a Warming World. J. Geophys. Res. Biogeosci. 2020, 125, e2020JG005678. [Google Scholar] [CrossRef]
- Partensky, F.; Hess, W.R.; Vaulot, D. Prochlorococcus, a Marine Photosynthetic Prokaryote of Global Significance. Microbiol. Mol. Biol. Rev. 1999, 63, 106–127. [Google Scholar] [CrossRef] [PubMed]
- Laws, E.A.; Falkowski, P.G.; Smith, W.O., Jr.; Ducklow, H.; McCarthy, J.J. Temperature Effects on Export Production in the Open Ocean. Glob. Biogeochem. Cycles 2000, 14, 1231–1246. [Google Scholar] [CrossRef]
- Carr, M.-E.; Friedrichs, M.A.M.; Schmeltz, M.; Aita, M.N.; Antoine, D.; Arrigo, K.R.; Asanuma, I.; Aumont, O.; Barber, R.; Behrenfeld, M.; et al. A Comparison of Global Estimates of Marine Primary Production from Ocean Color. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2006, 53, 741–770. [Google Scholar] [CrossRef]
- Alcorta, J.; Alarcón-Schumacher, T.; Salgado, O.; Díez, B. Taxonomic Novelty and Distinctive Genomic Features of Hot Spring Cyanobacteria. Front. Genet. 2020, 11, 568223. [Google Scholar] [CrossRef]
- Miller, S.R.; Castenholz, R.W. Evolution of Thermotolerance in Hot Spring Cyanobacteria of the Genus Synechococcus. Appl. Environ. Microbiol. 2000, 66, 4222–4229. [Google Scholar] [CrossRef]
- Steunou, A.-S.; Bhaya, D.; Bateson, M.M.; Melendrez, M.C.; Ward, D.M.; Brecht, E.; Peters, J.W.; Kühl, M.; Grossman, A.R. In Situ Analysis of Nitrogen Fixation and Metabolic Switching in Unicellular Thermophilic Cyanobacteria Inhabiting Hot Spring Microbial Mats. Proc. Natl. Acad. Sci. USA 2006, 103, 2398–2403. [Google Scholar] [CrossRef]
- Alcamán-Arias, M.E.; Pedrós-Alió, C.; Tamames, J.; Fernández, C.; Pérez-Pantoja, D.; Vásquez, M.; Díez, B. Diurnal Changes in Active Carbon and Nitrogen Pathways Along the Temperature Gradient in Porcelana Hot Spring Microbial Mat. Front. Microbiol. 2018, 9, 2353. [Google Scholar] [CrossRef]
- Prieto-Barajas, C.M.; Alfaro-Cuevas, R.; Valencia-Cantero, E.; Santoyo, G. Effect of Seasonality and Physicochemical Parameters on Bacterial Communities in Two Hot Spring Microbial Mats from Araró, Mexico. Rev. Mex. Biodivers. 2017, 88, 616–624. [Google Scholar] [CrossRef]
- Cox, A.; Shock, E.L.; Havig, J.R. The Transition to Microbial Photosynthesis in Hot Spring Ecosystems. Chem. Geol. 2011, 280, 344–351. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat Stress: An Overview of Molecular Responses in Photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under Stressful Environments: An Overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Hüve, K.; Bichele, I.; Rasulov, B.; Niinemets, Ü. When It Is Too Hot for Photosynthesis: Heat-Induced Instability of Photosynthesis in Relation to Respiratory Burst, Cell Permeability Changes and H2O2 Formation. Plant Cell Environ. 2011, 34, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Rowland, J.G.; Pang, X.; Suzuki, I.; Murata, N.; Simon, W.J.; Slabas, A.R. Identification of Components Associated with Thermal Acclimation of Photosystem II in Synechocystis Sp. PCC6803. PLoS ONE 2010, 5, e10511. [Google Scholar] [CrossRef]
- Finsinger, K.; Scholz, I.; Serrano, A.; Morales, S.; Uribe-lorio, L.; Mora, M.; Sittenfeld, A.; Weckesser, J.; Hess, W.R. Characterization of True-Branching Cyanobacteria from Geothermal Sites and Hot Springs of Costa Rica. Environ. Microbiol. 2008, 10, 460–473. [Google Scholar] [CrossRef]
- Schwabe, H. Über Die Algen Der Karlsbader Warmen Quellen. Linnaea 1837, 11, 109–127. [Google Scholar]
- Koch, R.; Kupczok, A.; Stucken, K.; Ilhan, J.; Hammerschmidt, K.; Dagan, T. Plasticity First: Molecular Signatures of a Complex Morphological Trait in Filamentous Cyanobacteria. BMC Evol. Biol. 2017, 17, 209. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. J. Gen. Microbiol. 1979, 110, 1–61. [Google Scholar] [CrossRef]
- Alcamán, M.E.; Fernandez, C.; Delgado, A.; Bergman, B.; Díez, B. The Cyanobacterium Mastigocladus Fulfills the Nitrogen Demand of a Terrestrial Hot Spring Microbial Mat. ISME J. 2015, 9, 2290–2303. [Google Scholar] [CrossRef]
- Alcamán, M.E.; Alcorta, J.; Bergman, B.; Vásquez, M.; Polz, M.; Díez, B. Physiological and Gene Expression Responses to Nitrogen Regimes and Temperatures in Mastigocladus Sp. Strain CHP1, a Predominant Thermotolerant Cyanobacterium of Hot Springs. Syst. Appl. Microbiol. 2017, 40, 102–113. [Google Scholar] [CrossRef]
- Mackenzie, R.; Pedrós-Alió, C.; Díez, B. Bacterial Composition of Microbial Mats in Hot Springs in Northern Patagonia: Variations with Seasons and Temperature. Extremophiles 2013, 17, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.R.; Castenholz, R.W.; Pedersen, D. Phylogeography of the Thermophilic Cyanobacterium Mastigocladus laminosus. Appl. Environ. Microbiol. 2007, 73, 4751–4759. [Google Scholar] [CrossRef]
- Muster, P.; Binder, A.; Schneider, K.; Bachofen, R. Influence of Temperature and PH on the Growth of the Thermophilic Cyanobacterium Mastigocladus laminosus in Continuous Culture. Plant Cell Physiol. 1983, 24, 273–280. [Google Scholar] [CrossRef]
- Stal, L.J. The Effect of Oxygen Concentration and Temperature on Nitrogenase Activity in the Heterocystous Cyanobacterium Fischerella Sp. Sci. Rep. 2017, 7, 5402. [Google Scholar] [CrossRef]
- Fish, A.; Danieli, T.; Ohad, I.; Nechushtai, R.; Livnah, O. Structural Basis for the Thermostability of Ferredoxin from the Cyanobacterium Mastigocladus laminosus. J. Mol. Biol. 2005, 350, 599–608. [Google Scholar] [CrossRef]
- Schirmer, T.; Bode, W.; Huber, R.; Sidler, W.; Zuber, H. X-ray Crystallographic Structure of the Light-Harvesting Biliprotein C-Phycocyanin from the Thermophilic Cyanobacterium Mastigocladus laminosus and Its Resemblance to Globin Structures. J. Mol. Biol. 1985, 184, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Glauser, M.; Bryant, D.A.; Frank, G.; Wehrli, E.; Rusconi, S.S.; Sidler, W.; Zuber, H. Phycobilisome Structure in the Cyanobacteria Mastigocladus laminosus and Anabaena Sp. PCC 7120. Eur. J. Biochem. 1992, 205, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Nies, M.; Wehrmeyer, W. Biliprotein Assembly in the Hemidiscoidal Phycobilisomes of the Thermophilic Cyanobacterium Mastigocladus laminosus Cohn. Characterization of Dissociation Products with Special Reference to the Peripheral Phycoerythrocyanin-Phycocyanin Complexes. Arch. Microbiol. 1981, 129, 374–379. [Google Scholar] [CrossRef]
- Reuter, W.; Nickel-Reuter, C. Molecular assembly of the phycobilisomes from the cyanobacterium Mastigocladus laminosus. J. Photochem. Photobiol. B 1992, 18, 51–66. [Google Scholar] [CrossRef]
- Duerring, M.; Huber, R.; Bode, W.; Ruembeli, R.; Zuber, H. Refined Three-Dimensional Structure of Phycoerythrocyanin from the Cyanobacterium Mastigocladus laminosus at 2.7 Å. J. Mol. Biol. 1990, 211, 633–644. [Google Scholar] [CrossRef]
- Reuter, W.; Wiegand, G.; Huber, R.; Than, M.E. Structural Analysis at 2.2 Å of Orthorhombic Crystals Presents the Asymmetry of the Allophycocyanin–Linker Complex, AP⋅LC7.8, from Phycobilisomes of Mastigocladus laminosus. Proc. Natl. Acad. Sci. USA 1999, 96, 1363–1368. [Google Scholar] [CrossRef] [PubMed]
- Kurisu, G.; Zhang, H.; Smith, J.L.; Cramer, W.A. Structure of the Cytochrome B6f Complex of Oxygenic Photosynthesis: Tuning the Cavity. Science 2003, 302, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Baniulis, D.; Yamashita, E.; Zhang, H.; Hasan, S.S.; Cramer, W.A. Structure-Function of the Cytochrome B6f Complex. Photochem. Photobiol. 2008, 84, 1349–1358. [Google Scholar] [CrossRef] [PubMed]
- Almog, O.; Shoham, G.; Michaeli, D.; Nechushtai, R. Monomeric and Trimeric Forms of Photosystem I Reaction Center of Mastigocladus laminosus: Crystallization and Preliminary Characterization. Proc. Natl. Acad. Sci. USA 1991, 88, 5312–5316. [Google Scholar] [CrossRef]
- Gisriel, C.; Shen, G.; Kurashov, V.; Ho, M.-Y.; Zhang, S.; Williams, D.; Golbeck, J.H.; Fromme, P.; Bryant, D.A. The Structure of Photosystem I Acclimated to Far-Red Light Illuminates an Ecologically Important Acclimation Process in Photosynthesis. Sci. Adv. 2020, 6, eaay6415. [Google Scholar] [CrossRef]
- Komenda, J.; Hassan, H.A.G.; Diner, B.A.; Debus, R.J.; Barber, J.; Nixon, P.J. Degradation of the Photosystem II D1 and D2 Proteins in Different Strains of the Cyanobacterium Synechocystis PCC 6803 Varying with Respect to the Type and Level of PsbA Transcript. Plant Mol. Biol. 2000, 42, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Barbato, R.; Friso, G.; Rigoni, F.; Dalla Vecchia, F.; Giacometti, G.M. Structural Changes and Lateral Redistribution of Photosystem II during Donor Side Photoinhibition of Thylakoids. J. Cell Biol. 1992, 119, 325–335. [Google Scholar] [CrossRef]
- Cardona, T.; Murray, J.W.; Rutherford, A.W. Origin and Evolution of Water Oxidation before the Last Common Ancestor of the Cyanobacteria. Mol. Biol. Evol. 2015, 32, 1310–1328. [Google Scholar] [CrossRef]
- Sheridan, K.J.; Duncan, E.J.; Eaton-Rye, J.J.; Summerfield, T.C. The Diversity and Distribution of D1 Proteins in Cyanobacteria. Photosynth. Res. 2020, 145, 111–128. [Google Scholar] [CrossRef]
- Kós, P.B.; Deák, Z.; Cheregi, O.; Vass, I. Differential Regulation of PsbA and PsbD Gene Expression, and the Role of the Different D1 Protein Copies in the Cyanobacterium Thermosynechococcus elongatus BP-1. Biochim. Biophys. Acta (BBA)-Bioenerg. 2008, 1777, 74–83. [Google Scholar] [CrossRef]
- Zhang, A.; Carroll, A.L.; Atsumi, S. Carbon Recycling by Cyanobacteria: Improving CO2 Fixation through Chemical Production. FEMS Microbiol. Lett. 2017, 364, fnx165. [Google Scholar] [CrossRef]
- Cooley, J.W.; Vermaas, W.F.J. Succinate Dehydrogenase and Other Respiratory Pathways in Thylakoid Membranes of Synechocystis Sp. Strain PCC 6803: Capacity Comparisons and Physiological Function. J. Bacteriol. 2001, 183, 4251–4258. [Google Scholar] [CrossRef]
- Nogales, J.; Gudmundsson, S.; Knight, E.M.; Palsson, B.O.; Thiele, I. Detailing the Optimality of Photosynthesis in Cyanobacteria through Systems Biology Analysis. Proc. Natl. Acad. Sci. USA 2012, 109, 2678–2683. [Google Scholar] [CrossRef] [PubMed]
- Sano, E.B.; Wall, C.A.; Hutchins, P.R.; Miller, S.R. Ancient Balancing Selection on Heterocyst Function in a Cosmopolitan Cyanobacterium. Nat. Ecol. Evol. 2018, 2, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.R.; Longley, R.; Hutchins, P.R.; Bauersachs, T. Cellular Innovation of the Cyanobacterial Heterocyst by the Adaptive Loss of Plasticity. Curr. Biol. 2020, 30, 344–350.e4. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Gan, F.; Shen, G.; Bryant, D.A. RfpA, RfpB, and RfpC Are the Master Control Elements of Far-Red Light Photoacclimation (FaRLiP). Front. Microbiol. 2015, 6, 1303. [Google Scholar] [CrossRef]
- Nürnberg, D.J.; Morton, J.; Santabarbara, S.; Telfer, A.; Joliot, P.; Antonaru, L.A.; Ruban, A.V.; Cardona, T.; Krausz, E.; Boussac, A.; et al. Photochemistry beyond the Red Limit in Chlorophyll f–Containing Photosystems. Science 2018, 360, 1210–1213. [Google Scholar] [CrossRef]
- Gisriel, C.J.; Cardona, T.; Bryant, D.A.; Brudvig, G.W. Molecular Evolution of Far-Red Light-Acclimated Photosystem II. Microorganisms 2022, 10, 1270. [Google Scholar] [CrossRef]
- Hirose, Y.; Fujisawa, T.; Ohtsubo, Y.; Katayama, M.; Misawa, N.; Wakazuki, S.; Shimura, Y.; Nakamura, Y.; Kawachi, M.; Yoshikawa, H.; et al. Complete Genome Sequence of Cyanobacterium Fischerella Sp. NIES-3754, Providing Thermoresistant Optogenetic Tools. J. Biotechnol. 2016, 220, 45–46. [Google Scholar] [CrossRef]
- Hirose, Y.; Ohtsubo, Y.; Misawa, N.; Yonekawa, C.; Nagao, N.; Shimura, Y.; Fujisawa, T.; Kanesaki, Y.; Katoh, H.; Katayama, M.; et al. Genome Sequencing of the NIES Cyanobacteria Collection with a Focus on the Heterocyst-Forming Clade. DNA Res. 2021, 28, dsab024. [Google Scholar] [CrossRef]
- Sato, S.; Ikeuchi, M.; Nakamoto, H. Expression and Function of a GroEL Paralog in the Thermophilic Cyanobacterium Thermosynechococcus elongatus under Heat and Cold Stress. FEBS Lett. 2008, 582, 3389–3395. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; MacRae, T.H. Small Heat Shock Proteins: Molecular Structure and Chaperone Function. Cell. Mol. Life Sci. 2005, 62, 2460–2476. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, H.; Chaurasia, A.K.; Apte, S.K. Cyanobacterial Heat-Shock Response: Role and Regulation of Molecular Chaperones. Microbiology 2014, 160, 647–658. [Google Scholar] [CrossRef]
- Tanaka, N.; Hiyama, T.; Nakamoto, H. Cloning, Characterization and Functional Analysis of GroESL Operon from Thermophilic Cyanobacterium Synechococcus vulcanus. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzym. 1997, 1343, 335–348. [Google Scholar] [CrossRef]
- Suzuki, I.; Simon, W.J.; Slabas, A.R. The Heat Shock Response of Synechocystis Sp. PCC 6803 Analysed by Transcriptomics and Proteomics. J. Exp. Bot. 2006, 57, 1573–1578. [Google Scholar] [CrossRef] [PubMed]
- Downs, C.A.; Ryan, S.L.; Heckathorn, S.A. The Chloroplast Small Heat-Shock Protein: Evidence for a General Role in Protecting Photosystem II against Oxidative Stress and Photoinhibition. J. Plant Physiol. 1999, 155, 488–496. [Google Scholar] [CrossRef]
- Preiss, J. Bacterial Glycogen Synthesis and Its Regulation. Annu. Rev. Microbiol. 1984, 38, 419–458. [Google Scholar] [CrossRef]
- Lea-Smith, D.J.; Bombelli, P.; Vasudevan, R.; Howe, C.J. Photosynthetic, Respiratory and Extracellular Electron Transport Pathways in Cyanobacteria. Biochim. Biophys. Acta (BBA)-Bioenerg. 2016, 1857, 247–255. [Google Scholar] [CrossRef]
- Karp, P.D.; Billington, R.; Caspi, R.; Fulcher, C.A.; Latendresse, M.; Kothari, A.; Keseler, I.M.; Krummenacker, M.; Midford, P.E.; Ong, Q.; et al. The BioCyc Collection of Microbial Genomes and Metabolic Pathways. Brief. Bioinform. 2019, 20, 1085–1093. [Google Scholar] [CrossRef]
- Padhi, B.; Chauhan, G.; Kandoi, D.; Stirbet, A.; Tripathy, B.C.; Govindjee, G. A Comparison of Chlorophyll Fluorescence Transient Measurements, Using Handy Pea and Fluorpen Fluorometers. Photosynthetica 2021, 59, 399–408. [Google Scholar] [CrossRef]
- Ranglová, K.; Lakatos, G.E.; Manoel, J.A.C.; Grivalský, T.; Masojídek, J. Rapid Screening Test to Estimate Temperature Optima for Microalgae Growth Using Photosynthesis Activity Measurements. Folia Microbiol. 2019, 64, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Stirbet, A.; Lazár, D.; Papageorgiou, G.C.; Govindjee, G. Chlorophyll a Fluorescence in Cyanobacteria: Relation to Photosynthesis. In Cyanobacteria: From Basic Science to Applications; Academic Press: Cambridge, MA, USA, 2018; pp. 79–130. ISBN 9780128146682. [Google Scholar]
- Sabath, N.; Ferrada, E.; Barve, A.; Wagner, A. Growth Temperature and Genome Size in Bacteria Are Negatively Correlated, Suggesting Genomic Streamlining during Thermal Adaptation. Genome Biol. Evol. 2013, 5, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Zhang, S.; Rockwell, N.C.; Martin, S.S.; Lagarias, J.C.; Bryant, D.A. Extensive Remodeling of a Cyanobacterial Photosynthetic Apparatus in Far-Red Light. Science 2014, 345, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Shen, G.; Bryant, D.A. Occurrence of Far-Red Light Photoacclimation (FaRLiP) in Diverse Cyanobacteria. Life 2015, 5, 4–24. [Google Scholar] [CrossRef]
- Cherepanov, D.A.; Shelaev, I.V.; Gostev, F.E.; Aybush, A.V.; Mamedov, M.D.; Shen, G.; Nadtochenko, V.A.; Bryant, D.A.; Semenov, A.Y.; Golbeck, J.H. Evidence That Chlorophyll f Functions Solely as an Antenna Pigment in Far-Red-Light Photosystem I from Fischerella thermalis PCC 7521. Biochim. Biophys. Acta (BBA)-Bioenerg. 2020, 1861, 148184. [Google Scholar] [CrossRef]
- Ho, M.-Y.; Shen, G.; Canniffe, D.P.; Zhao, C.; Bryant, D.A. Light-Dependent Chlorophyll f Synthase Is a Highly Divergent Paralog of PsbA of Photosystem II. Science 2016, 353, aaf9178. [Google Scholar] [CrossRef]
- Wada, H.; Murata, N. The Essential Role of Phosphatidylglycerol in Photosynthesis. Photosynth. Res. 2007, 92, 205–215. [Google Scholar] [CrossRef]
- Endo, K.; Kobayashi, K.; Wang, H.-T.; Chu, H.-A.; Shen, J.-R.; Wada, H. Site-Directed Mutagenesis of Two Amino Acid Residues in Cytochrome B559 α Subunit That Interact with a Phosphatidylglycerol Molecule (PG772) Induces Quinone-Dependent Inhibition of Photosystem II Activity. Photosynth. Res. 2019, 139, 267–279. [Google Scholar] [CrossRef]
- Narusaka, Y.; Murakami, A.; Saeki, M.; Kobayashi, H.; Satoh, K. Preliminary Characterization of a Photo-Tolerant Mutant of Synechocystis Sp. PCC 6803 Obtained by in Vitro Random Mutagenesis of PsbA2. Plant Sci. 1996, 115, 261–266. [Google Scholar] [CrossRef]
- Gong, H.; Nilsen, S. Effect of Temperature on Photoinhibition of Photosynthesis, Recovery, Turnover of the 32 KD Chloroplast Protein in Lemna gibba. J. Plant Physiol. 1989, 135, 9–14. [Google Scholar] [CrossRef]
- Bohler, M.C.; Binder, A. Photosynthetic Activities of a Membrane Preparation of the Thermophilic Cyanobacterium Mastigocladus laminosus. Arch. Microbiol. 1980, 124, 155–160. [Google Scholar] [CrossRef]
- Radway, J.C.; Weissman, J.C.; Wilde, E.W.; Benemann, J.R. Exposure of Fischerella [Mastigocladus] to High and Low Temperature Extremes: Strain Evaluation for a Thermal Mitigation Process. J. Appl. Phycol. 1992, 4, 67–77. [Google Scholar] [CrossRef]
- Kopf, M.; Klähn, S.; Scholz, I.; Matthiessen, J.K.F.; Hess, W.R.; Voß, B. Comparative Analysis of the Primary Transcriptome of Synechocystis Sp. PCC 6803. DNA Res. 2014, 21, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Mulo, P.; Sakurai, I.; Aro, E.-M. Strategies for PsbA Gene Expression in Cyanobacteria, Green Algae and Higher Plants: From Transcription to PSII Repair. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 247–257. [Google Scholar] [CrossRef]
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A Toolkit to Classify Genomes with the Genome Taxonomy Database. Bioinformatics 2020, 36, 1925–1927. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Edgar, R.C. Muscle: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Li, Z.; Sun, Y.; Aluko, O.O.; Wu, X.; Wang, Q.; Liu, G. MG2C: A User-Friendly Online Tool for Drawing Genetic Maps. Mol. Hortic. 2021, 1, 16. [Google Scholar] [CrossRef]
- Liles, C.; Kumar, M.A.; Weinshenker, D. Use of Gene-Specific Primer Cocktails for First-Strand CDNA Synthesis With a Reverse Transcriptase Kit. Am. Biotechnol. Lab. 2004, 22, 20–21. [Google Scholar]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Tran, T.D.C.; Bernard, C.; Ammar, M.; Chaouch, S.; Comte, K. Heat Shock Transcriptional Responses in an MC-Producing Cyanobacterium (Planktothrix Agardhii) and Its MC-Deficient Mutant under High Light Conditions. PLoS ONE 2013, 8, e73198. [Google Scholar] [CrossRef]
- Pinto, F.; Pacheco, C.C.; Ferreira, D.; Moradas-ferreira, P.; Tamagnini, P. Selection of Suitable Reference Genes for RT-QPCR Analyses in Cyanobacteria. PLoS ONE 2012, 7, e34983. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. EggNOG-Mapper v2: Functional Annotation, Orthology Assignments, and Domain Prediction at the Metagenomic Scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Bolstad, B. PreprocessCore: A Collection of Pre-Processing Functions. Available online: https://github.com/bmbolstad/preprocessCore (accessed on 11 June 2022).
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Morris, J.H.; Apeltsin, L.; Newman, A.M.; Baumbach, J.; Wittkop, T.; Su, G.; Bader, G.D.; Ferrin, T.E. ClusterMaker: A Multi-Algorithm Clustering Plugin for Cytoscape. BMC Bioinform. 2011, 12, 436. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Pennec, E.L.; Slowikowski, K. A Word Cloud Geom for Ggplot2, Ggwordcloud. Available online: https://lepennec.github.io/ggwordcloud/ (accessed on 29 September 2022).
- Zhang, S.; Bryant, D.A. The Tricarboxylic Acid Cycle in Cyanobacteria. Science 2011, 334, 1551–1554. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; RStudio: Vienna, Austria, 2022. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | F-Value | p-Value |
|---|---|---|
| Temperature | 14.0456 | <0.0001 |
| DCMU | 1.5343 | 0.2188 |
| Temperature/DCMU | 2.6381 | 0.0286 |
| Temperature (°C) | Diff | Lower | Upper | Adj. p-Value |
|---|---|---|---|---|
| 45–40 | 0.0390115 | −0.009507 | 0.0875305 | 0.1740 |
| 50–40 | −0.018833 | −0.070941 | 0.0332745 | 0.8500 |
| 55–40 | −0.026833 | −0.078941 | 0.0252745 | 0.6040 |
| 60–40 | −0.089833 | −0.141941 | −0.037725 | <0.0001 |
| 50–45 | −0.057845 | −0.102187 | −0.013503 | 0.0043 |
| 55–45 | −0.065845 | −0.110187 | −0.021503 | 0.0008 |
| 60–45 | −0.128845 | −0.173187 | −0.084503 | <0.0001 |
| 55–50 | −0.008 | −0.056242 | 0.0402425 | 0.9900 |
| 60–50 | −0.071 | −0.119242 | −0.022758 | 0.0009 |
| 60–55 | −0.063 | −0.111242 | −0.014758 | 0.0043 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vergara-Barros, P.; Alcorta, J.; Casanova-Katny, A.; Nürnberg, D.J.; Díez, B. Compensatory Transcriptional Response of Fischerella thermalis to Thermal Damage of the Photosynthetic Electron Transfer Chain. Molecules 2022, 27, 8515. https://doi.org/10.3390/molecules27238515
Vergara-Barros P, Alcorta J, Casanova-Katny A, Nürnberg DJ, Díez B. Compensatory Transcriptional Response of Fischerella thermalis to Thermal Damage of the Photosynthetic Electron Transfer Chain. Molecules. 2022; 27(23):8515. https://doi.org/10.3390/molecules27238515
Chicago/Turabian StyleVergara-Barros, Pablo, Jaime Alcorta, Angélica Casanova-Katny, Dennis J. Nürnberg, and Beatriz Díez. 2022. "Compensatory Transcriptional Response of Fischerella thermalis to Thermal Damage of the Photosynthetic Electron Transfer Chain" Molecules 27, no. 23: 8515. https://doi.org/10.3390/molecules27238515
APA StyleVergara-Barros, P., Alcorta, J., Casanova-Katny, A., Nürnberg, D. J., & Díez, B. (2022). Compensatory Transcriptional Response of Fischerella thermalis to Thermal Damage of the Photosynthetic Electron Transfer Chain. Molecules, 27(23), 8515. https://doi.org/10.3390/molecules27238515