

Recent Advances in Herbal-Derived Products with Skin Anti-Aging Properties and Cosmetic Applications

Abstract
1. Introduction
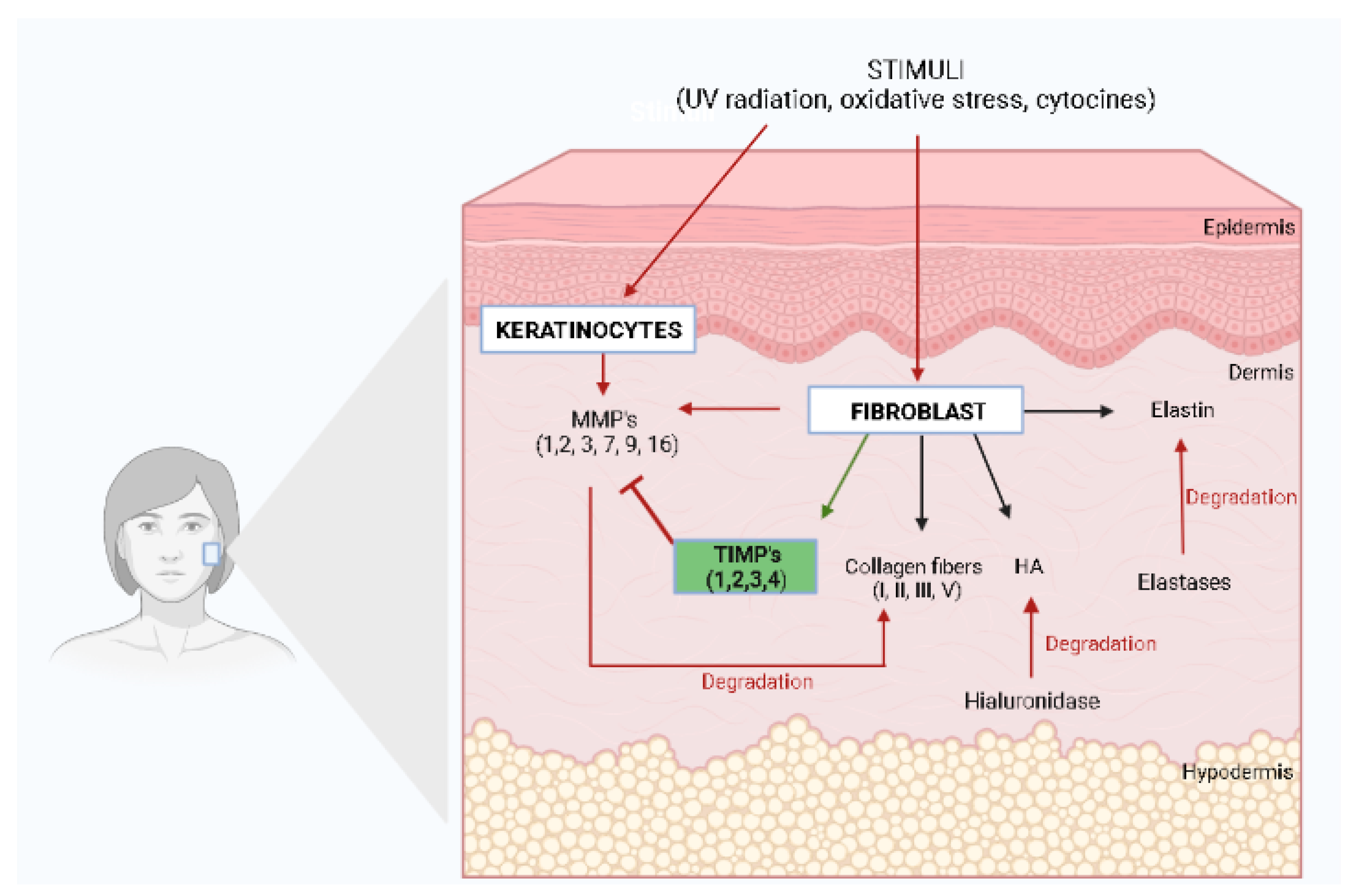
2. Skin Anti-Aging Natural Products by Acting on the Skin Elasticity and Wrinkle Formation
- Elastase inhibitory activity was determined after the use of the plant extracts prepared with Eugenia dysenterica [26], Gastrodia elata [27], Litchi sinensis [28], Magnolia officinalis [29], Malaxis acuminata [30], Manilkara zapota [14], Nephelium lappaceum [28], Phyllanthus emblica [14], Sclerocarya birrea [31], Sylibum marianum [14], Spatholobus suberectus [32], Tamarindus indica [28], and a polyherbal formulation containing Nyctanthes arbor-tristis leaf, unripe and ripe Aegle marmelos fruit pulp, and the terminal meristem of Musa paradisiaca extracts [33];
- An increase in pro-collagen expression or synthesis was observed after treatments with plant extracts prepared from Alchemilla mollis [36], Azadirachta indica [37], Camellia sinensis [38], Citrus junus [39], Trapa japonica [40], and a mixture of plant extracts of Kochia scoparia and Rosa multiflora [41];
- The downregulation of MMP-1 expression was described after treatments with plant extracts of Alchemilla mollis [36], Allium sativum [49], Azadirachta indica [37], Camellia sinensis [38], Gastrodia elata [27], Kochia scoparia [41], Magnolia officinalis [29], Passiflora tarminiana [45], Penthorium chinense [47], Rosa multiflora ([41], and Syzygium aromaticum [50];
- The inhibition of the unspecific collagenase activity was observed after treatments with plant extracts of Cassia fistula [43], Curcuma heyneana [52], Eugenia dysenterica [26], Hibiscus sabdariffa [53], Litchi chinens [38], Magnolia officinalis [29], Malaxis acuminate [30], Manilkara zapota [14], Nephelium lappaceum [28], Passiflora tarminiana [45], Phyllanthus emblica [14], Piper cambodianum [34], Sclerocarya birrea [31], Sylibum marianum [14], and Tamarindus indica [28].
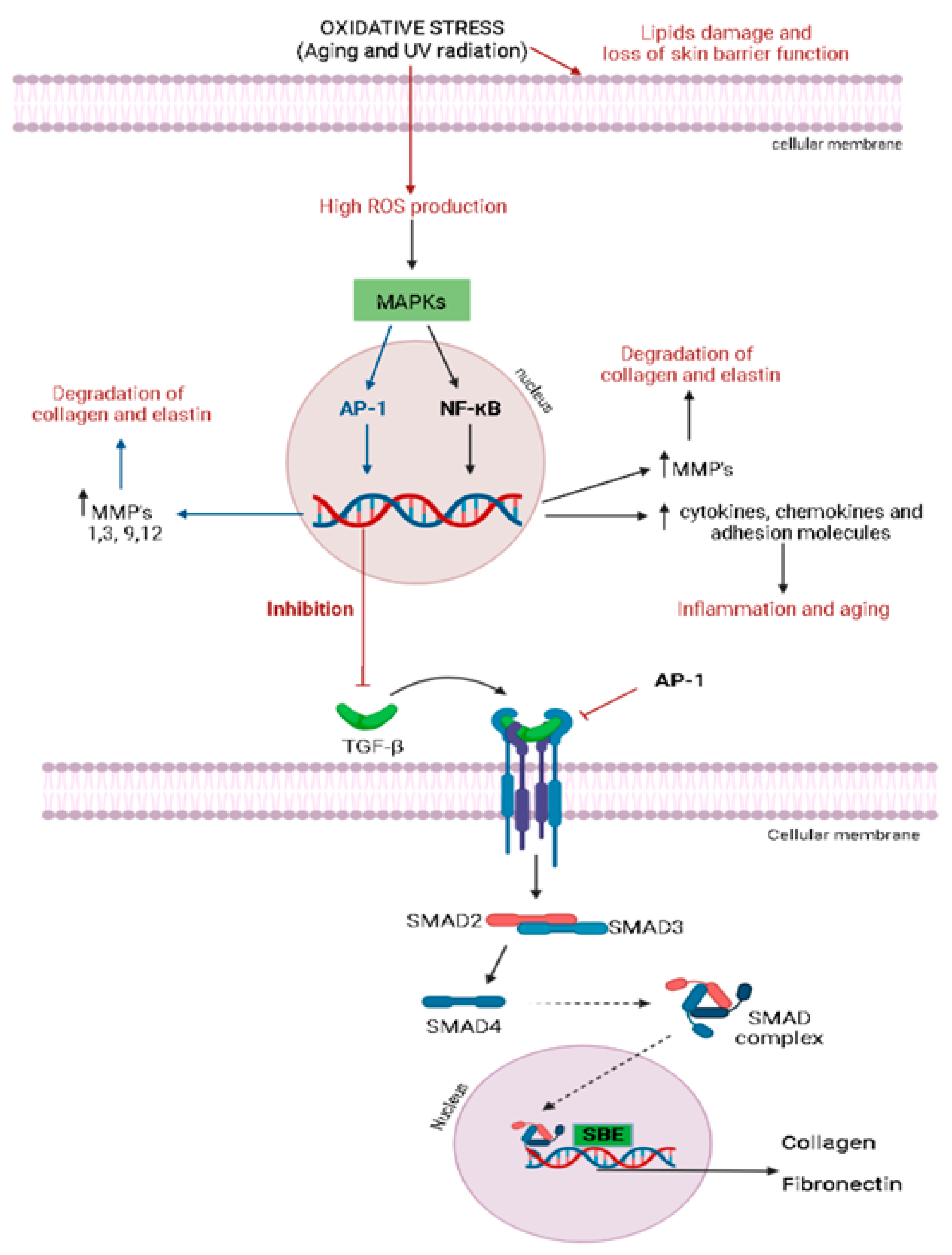
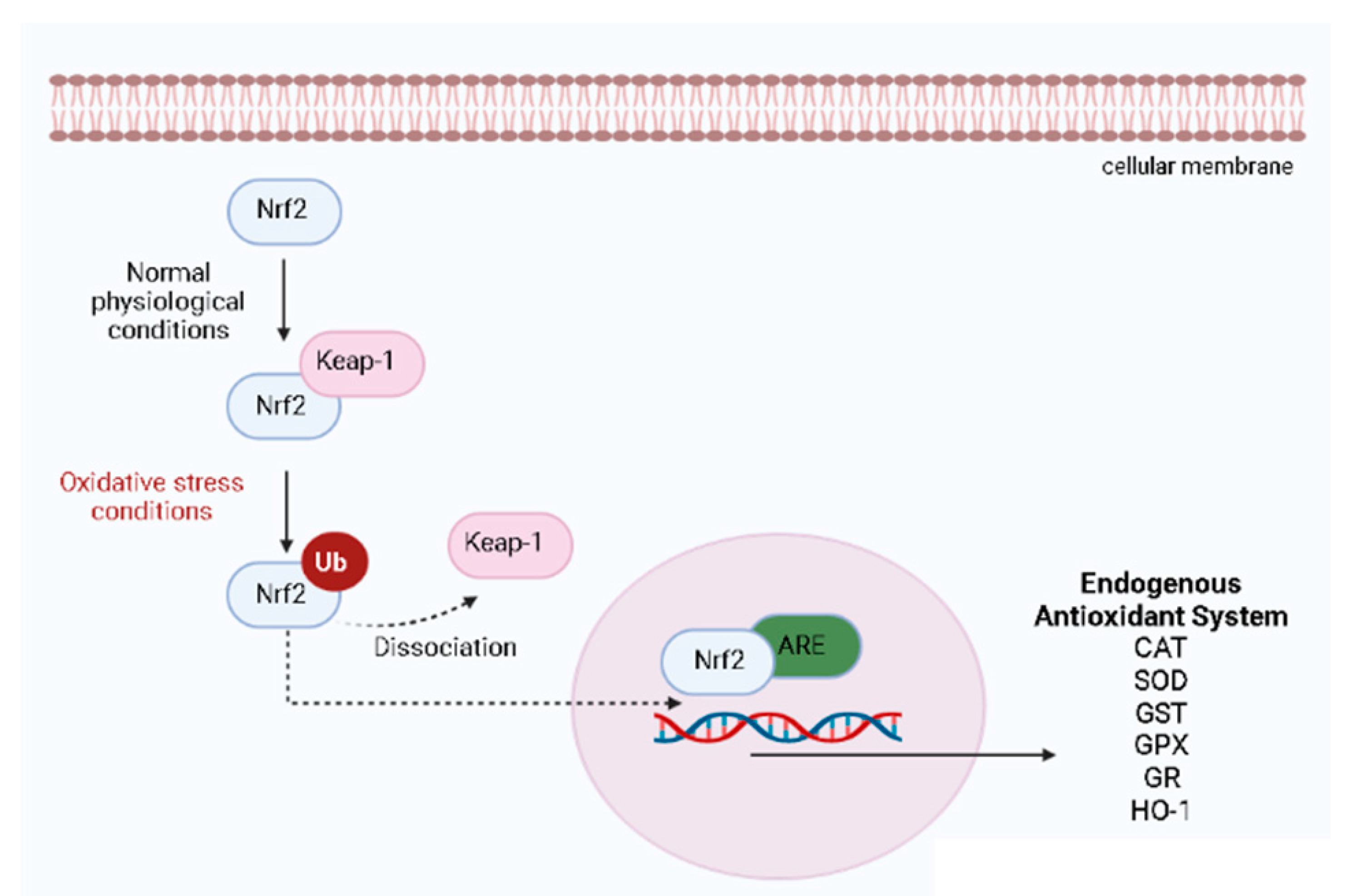
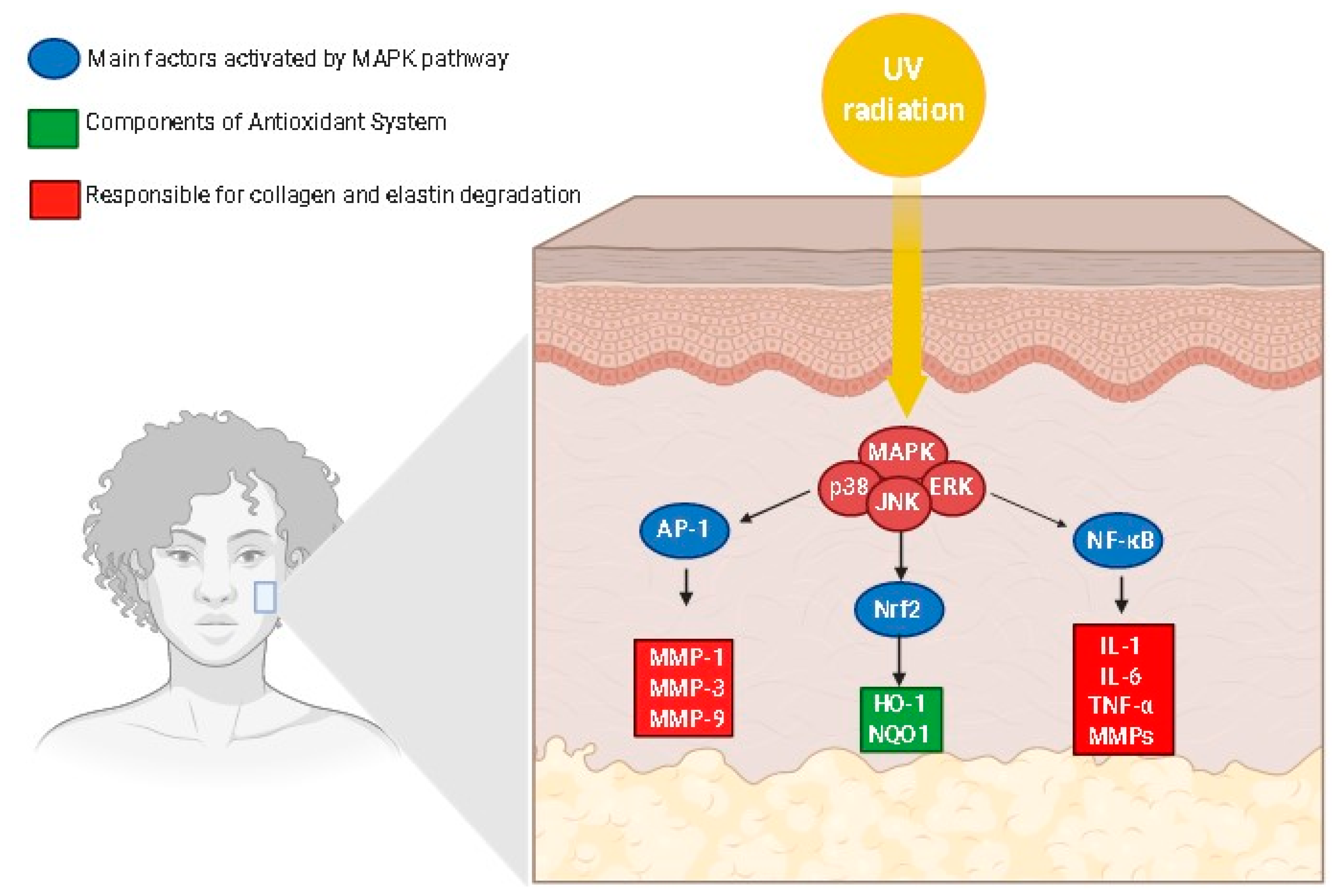
3. Skin Anti-Aging Natural Products by Acting on the Skin Oxidative Stress
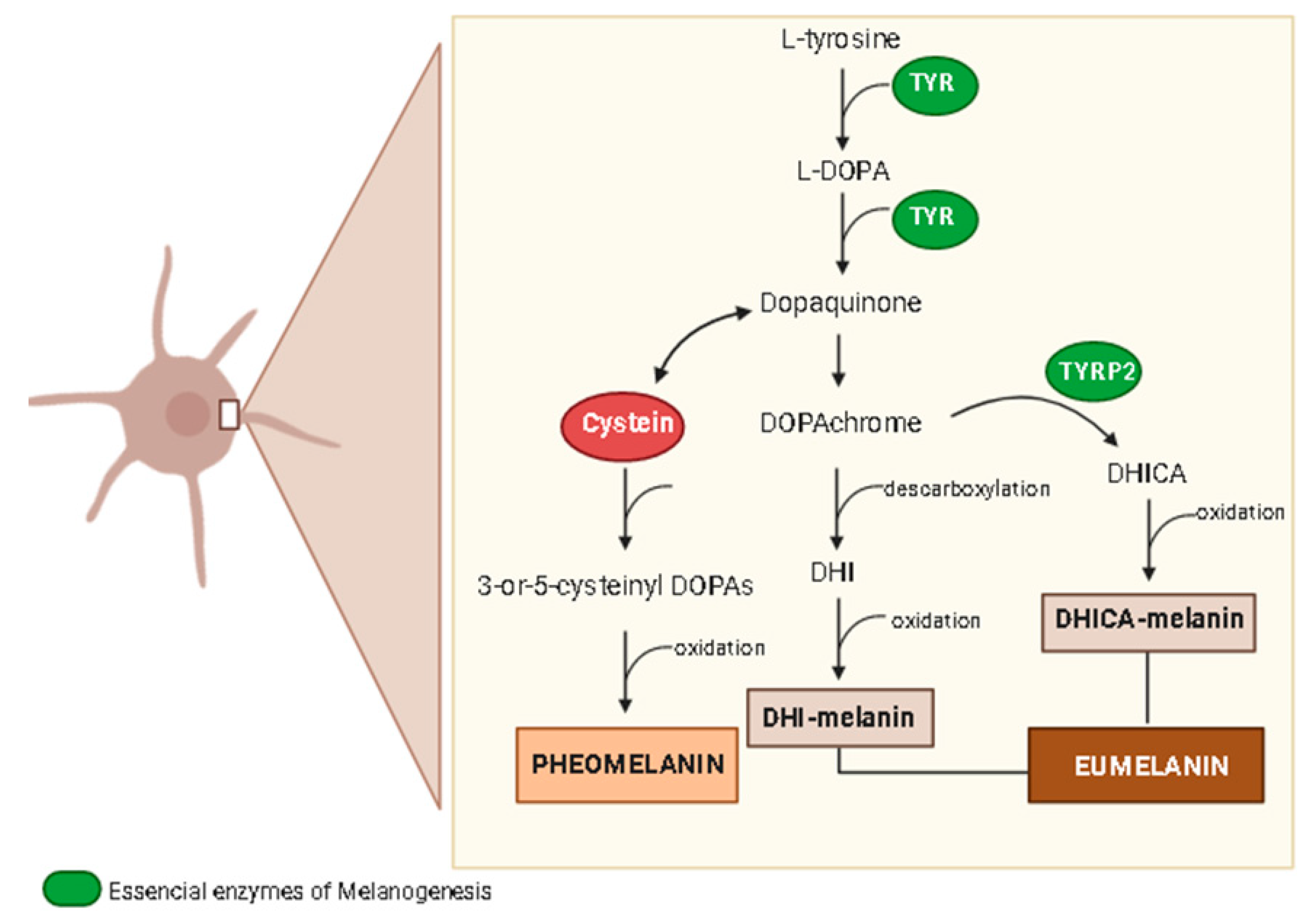
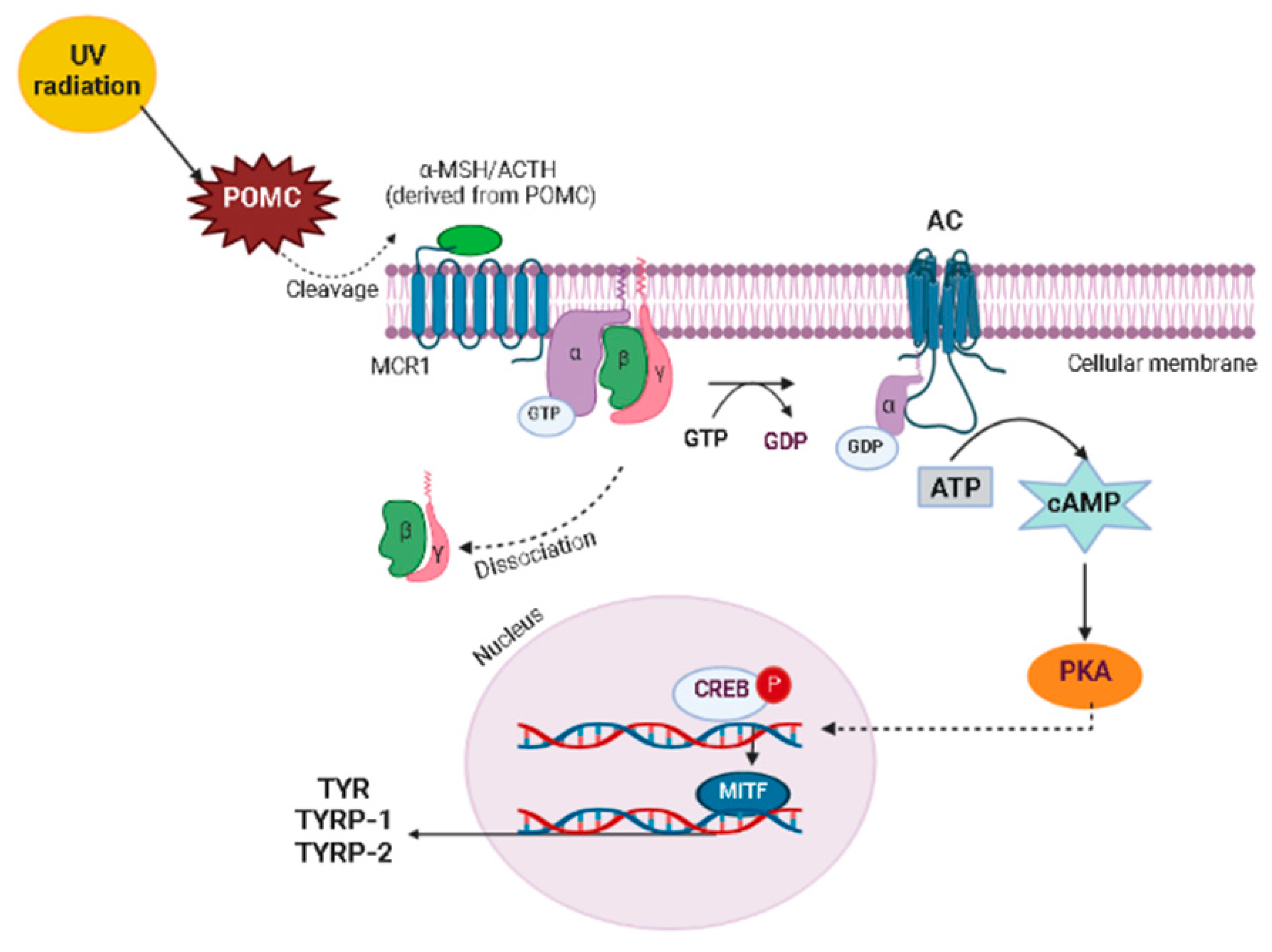
4. Skin Anti-Aging Natural Products by Acting on the Skin Pigmentation
5. Skin Anti-Aging Natural Products by Acting on the Skin Hydration
6. Skin Anti-Aging Natural Products by Acting on other Signaling and Transcriptional Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants/Products | Effects | Refs |
|---|---|---|
| Alchemilla mollis | In vitro (NHDF cells) inhibition of AP-1 activation, c-Jun, and c-Fos levels, and increase in Nrf2 pathway | [36] |
| Allium sativum | In vitro (HaCaT cells) inhibition of UV-induced increase in SA-β-gal levels and UV-induced decrease in SIRT1 activity | [49] |
| Andrographis paniculata | In vitro (Human epidermal stem cells) increase VEGF production and upregulation of integrin β1 | [42] |
| Azadirachta indica | In vitro (NHDF cells) downregulation of c-Jun and c-Fos proteins and upregulation of TGF-β | [37] |
| Camelia sinensis | In vitro (HaCat cells) downregulation of HO-1 and upregulation of Nrf2 via phosphorylation of p38 and ERK | [38] |
| Catechin from Leontopodium alpinum | In vitro (HaCaT cells) suppression of p65 (NF-κB) phosphorylation | [73] |
| Kochia scoparia | In vitro (NHDF cells) upregulation of PPAR-γ and PPAR-α | [41] |
| Polysaccharides from Panax ginseng | In vitro (HaCaT cells) decrease UV-induced AP-1 transactivation | [24] |
| Pourthiaea villosa | In vitro (NHDF cells) inhibition of AP-1 activation-related JNK and p38 MAPKs | [51] |
| Rosa multiflora | In vitro (NHDF cells) upregulation of PPAR-γ and PPAR-α | [41] |
| Spatholobus suberectus | In vitro (HaCaT cells) downregulation of NF-κB and AP-1 pathways, inhibition of phosphorylated ERK1/2 and p38 | [32] |
| Solanum tuberosum | In vitro (NHDF cells) induction of TGF-β signaling pathway, increment of phosphorylation of Akt and ERk1/2 | [13] |
| Syzygium aromaticum | In vitro (NHDF cells) reduction of AP-1 signaling pathway via reduction in c-Jun and c-Fos levels, suppression of NF-κB expression, and upregulation of Nrf2 | [50] |
| Trapa japonica | In vitro (NHDF and HaCaT cells) activation of the TGF-β/GSK-3β/β-catenin pathway | [40] |
| Thymus vulgaris | In vitro (3T3-L1 embryonic fibroblasts) upregulation of PPARγ and increase in adiponectin production | [4] |
| Ulmus macrocarpa | In vitro (HDF cells) blockade of JNK and p38 MAPK signaling | [61] |
| Verbascoside from Syringa vulgaris | In vitro (HaCaT cells) suppression of p65 (NF-κB) phosphorylation | [73] |
| Vitis vinifera | In vitro (NHDF cells) induction of SIRT 1 expression | [74] |
| Xanthones from Garcinia mangostana | In vivo (hairless mice) suppression of UVB-induced phosphorylation of MAPKs (ERK, P38, and JNK) and downregulation of IL-1β, IL-6, and TNF-α | [19] |
7. Promising Natural Compounds and Herbal Preparation for the Development of New Skin Anti-Aging Cosmetics
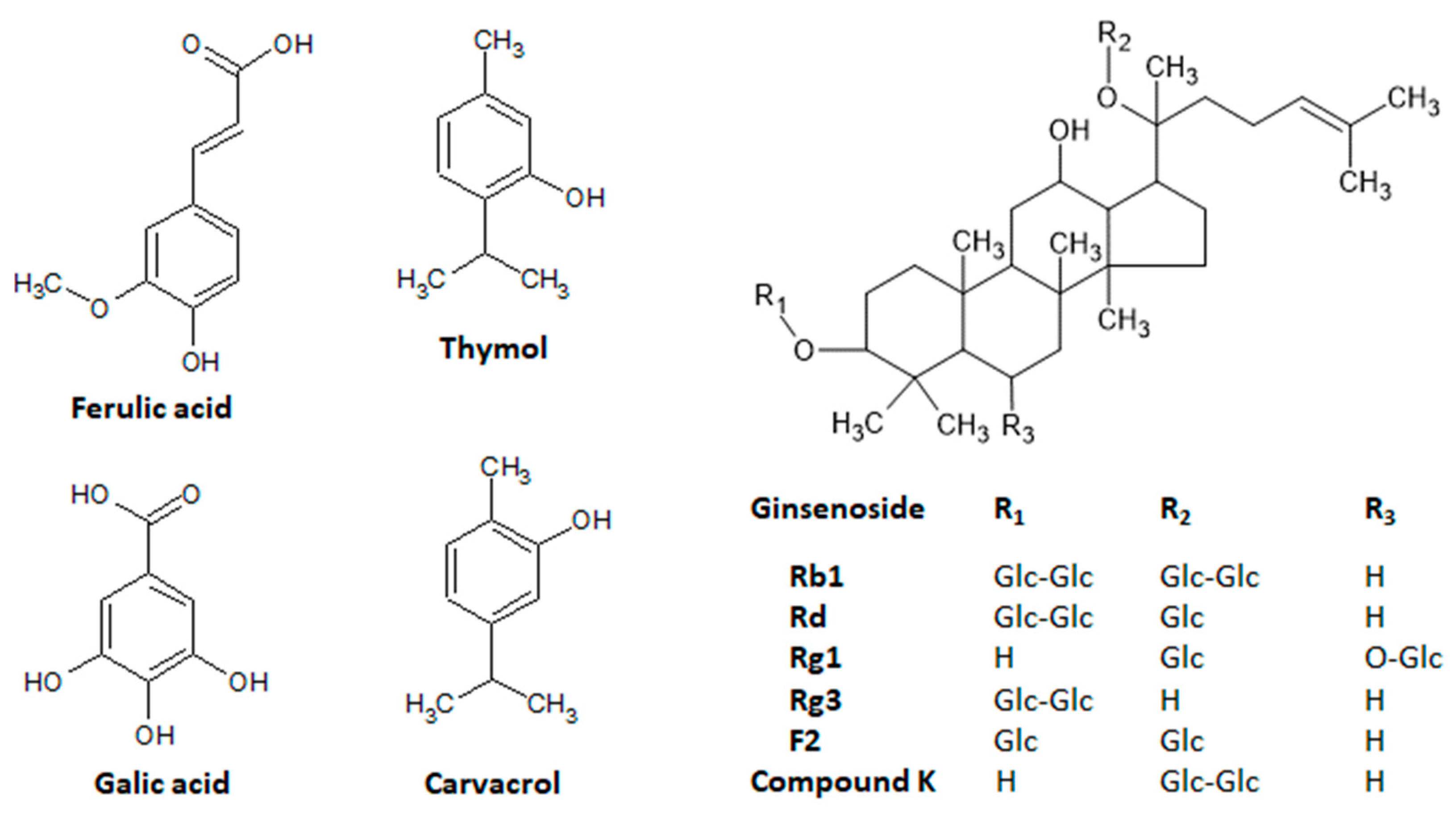
7.1. Ferulic Acid
7.2. Gallic Acid
7.3. Thymus vulgaris
7.4. Panax ginseng
7.5. Triticum aestivum L.
7.6. Andrographis paniculata
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Li, Z.; Zhang, Z.; Ren, Y.; Wang, Y.; Fang, J.; Yue, H.; Ma, S.; Guan, F. Aging and age-related diseases: From mechanism to therapeutic strategies. Biogerontology 2021, 22, 165–187. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Garagnani, P.; Morsiani, C.; Conte, M.; Santoro, A.; Grignolio, A.; Monti, D.; Capri, M.; Salvioli, S. The continuum of aging and age-related diseases: Common mechanisms but different rates. Front. Med. 2018, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, J.P.; Stevens, M.; Thornton, D. The business of anti-aging science. Trends Biotechnol. 2017, 35, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Caversan, J.; Mussi, L.; Sufi, B.; Padovani, G.; Nazaro, L.; Camargo-Junior, F.B.; Magalhães, W.V.; Di Stasi, L.C. A new phytocosmetic preparation from Thymus vulgaris stimulates adipogenesis and controls skin aging process: In vitro studies and topical effects in a double-blind placebo-controlled clinical trial. J. Cosmet. Dermatol. 2021, 20, 2190–2202. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.A.; Mikail, M.A.; Zamakshshari, N.; Abdulah, A.S. Natural anti-aging skincare: Role and potential. Biogerontology 2020, 21, 293–310. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Elewa, R.; Makrantonaki, E. Aesthetic aspects of skin aging, prevention, and local treatment. Clin. Dermatol. 2019, 37, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Dai, B.; Pelton, L.E. Exploring consumer’s skincare retails patronage. J. Retail. Cons. Serv. 2018, 43, 269–277. [Google Scholar] [CrossRef]
- AMR (Allied Market Research). Cosmetic Market by Category, Gender and Distribution Channel: Opportunity, Analysis and Industry Forecast, 2021–2027. Available online: https:///www.alliedmarketresearch.com/cosmetics-market (accessed on 4 July 2022).
- Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Makrantonaki, E.; Zoubolis, C.C. Skin anti-aging strategies. Dermato-Endocrinology 2012, 4, 308–319. [Google Scholar] [CrossRef]
- Di Stasi, L.C. Coumarin derivatives in Inflammatory Bowel Diseases. Molecules 2021, 26, 422. [Google Scholar] [CrossRef]
- Shin, J.W.; Kwon, S.H.; Choi, J.Y.; Na, J.I.; Huh, C.H.; Choi, H.R.; Park, K.C. Molecular mechanisms of dermal aging and antiaging approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef]
- Tracy, L.E.; Minasian, R.A.; Caterson, E.J. Extracellular matrix and dermal fibroblast function in the healing wound. Adv. Wound Care 2016, 5, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Suto, M.; Masutomi, H.; Ishihara, K.; Masaki, H. A potato peel extract stimulates type I collagen synthesis via Akt and ERK signaling in normal human dermal fibroblasts. Biol. Pharm. Bull. 2019, 42, 1510–1516. [Google Scholar] [CrossRef] [PubMed]
- Pientaweeratch, S.; Panapisal, V.; Tansirikongkol, A. Antioxidant, anti-collagenase and anti-elastase activities of Phyllanthus emblica, Manilkara zapota and sylimarin: An in vitro comparative study for anti-aging applications. Pharmac. Biol. 2016, 54, 1865–1872. [Google Scholar] [CrossRef]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and biological attributes of matrix metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [PubMed]
- Choi, E.H. Aging of the skin barrier. Clin. Dermatol. 2019, 37, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef] [PubMed]
- Krutmann, J.; Bouloc, A.; Sore, G.; Bernad, B.A.; Passeron, T. The skin aging exposome. J. Dermatol. Sci. 2017, 85, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Im, A.R.; Kim, Y.M.; Chin, Y.C.; Chae, S. Protective effects of compounds from Garcinia mangostana L. (mangosteen) against UVB damage in HaCaT cells and hairless mice. Int. J. Mol. Med. 2017, 40, 1941–1949. [Google Scholar] [CrossRef][Green Version]
- Ma, R.J.; Yang, L.; Bai, X.; Yuan, M.Y.; Wang, Y.K.; Xie, Y.; Hu, J.M.; Zhou, J. Phenolic constituents with antioxidative, tyrosinase inhibitory and anti-aging activities from Dendrobium loddigesii Rolfe. Nat. Prod. Bioprosp. 2019, 9, 329–336. [Google Scholar] [CrossRef]
- Morikawa, T.; Nagatomo, A.; Kitazawa, K.; Muraoka, O.; Kikuchi, T.; Yamada, T.; Tanaka, R.; Ninomiya, K. Collagen synthesis-promoting effects of andiroba oil and its limonoid constituents in normal human dermal fibroblasts. J. Oleo Sci. 2018, 67, 1271–1277. [Google Scholar] [CrossRef]
- Tan, H.; Sonam, T.; Shimizu, K. The potential of triterpenoids from loquat leaves (Eryobotrya japonica) for prevention and treatment of skin disorder. Int. J. Mol. Sci. 2017, 18, 1030. [Google Scholar] [CrossRef] [PubMed]
- Henriet, E.; Jäger, S.; Tran, C.; Bastien, P.; Michelet, J.F.; Minondo, A.M.; Formanek, F.; Dalko-Csiba, M.; Lortat-Jacob, H.; Breton, L.; et al. A jasmonic acid derivative improves skin healing and induces changes in proteoglycan expression and glycosaminoglycan structure. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2250–2260. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Jang, S.G.; Lee, C.H.; Lee, J.Y.; Park, H.; Kim, J.H.; Lee, S.; Kim, S.H.; Park, E.Y.; Lee, K.W.; et al. Beneficial effects on skin health using polysaccharides from red ginseng by-product. J. Food Biochem. 2019, 43, e12961. [Google Scholar] [CrossRef]
- Zdunska-Peciak, K.; Rotsztejn, H. The effectiveness of ferulic acid and microneedling in reducing signs of photoaging: A split-face comparative study. Dermatol. Ther. 2020, 33, e14000. [Google Scholar]
- Moreira, L.C.; Ávila, R.I.; Veloso, D.F.M.C.; Pedrosa, T.N.; Lima, E.S.; Couto, R.O.; Lima, E.M.; Batista, A.C.; Paula, J.R.; Valadares, M.C. In vitro safety and efficacy of a complex botanical mixture of Eugenia dysenterica Dc. (Myrtaceae): Prospects for developing a new dermocosmetic product. Toxicol. Vitr. 2017, 45, 394–408. [Google Scholar] [CrossRef] [PubMed]
- Song, E.; Chung, H.; Shin, E.; Jeong, J.K.; Han, B.K.; Choi, H.J.; Hwang, J. Gastrodea elata Blume extract modulates antioxidant activity and ultraviolet A-irradiated skin aging in human dermal fibroblast cells. J. Med. Food 2016, 19, 1057–1064. [Google Scholar] [CrossRef]
- Lourith, N.; Kanlayavattanakul, M.; Chaikul, P.; Chansriniyom, C.; Bunwatcharaphansakun, P. In vitro and cellular activities of the selected fruit residues for skin aging treatment. An. Acad. Bras. Ciências 2017, 89, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Chen, C.; Cheng, C.; Dai, H.; Ai, Y.; Lin, C.; Chung, Y. Evaluation of tyronisase inhibitory, antioxidant, antimicrobial, and antiaging activities of Magnolia officinalis extracts after Aspergillus niger fermentation. Biomed. Res. Int. 2018, 2018, 5201786. [Google Scholar] [CrossRef] [PubMed]
- Bose, B.; Choudhury, H.; Tandon, P.; Kumaria, S. Studies on secondary metabolite profiling, anti-inflammatory potential, in vitro photoprotective and skin-aging related enzyme inhibitory activities of Malaxis acuminate, a threatened orchid of nutraceutical importance. J. Photochem. Photobiol. B 2017, 173, 686–695. [Google Scholar] [CrossRef]
- Shoko, T.; Naharaj, V.J.; Naidoo, D.; Tselanyane, M.; Nthambeleni, R.; Khorombi, E.; Apostolides, Z. Anti-aging potential of extracts from Sclerocarya birrea (A. Rich.) Hochst and its chemical profiling by UPLC-Q-TOF-MS. BMC Complem. Alt. Med. 2018, 18, 54. [Google Scholar] [CrossRef]
- Kwon, K.R.; Alam, M.B.; Park, J.H.; Kim, T.H.; Lee, S.H. Attenuation of UVB-induced photo-aging by polyphenolic-rich Spatholobus suberectus stem extract via modulation of MAPK/AP-1/MMPs signaling in human keratinocytes. Nutrients 2019, 11, 1341. [Google Scholar] [CrossRef] [PubMed]
- Sundaran, I.K.; Sarangi, D.D.; Sundararajan, V.; George, S.; Mohideen, S.S. Poly herbal formulation with anti-elastase and antioxidant properties for skin anti-aging. BMC Complem. Alt. Med. 2018, 18, 33. [Google Scholar]
- Lee, H.; Hong, Y.; Kwon, S.H.; Park, J.; Park, J. Anti-aging effects of Piper cambodianum P. Fourn. extract on normal human dermal fibroblast cells and a wound-healing model in mice. Clin. Interv. Aging 2016, 11, 1017–1026. [Google Scholar] [PubMed]
- Dieament, G.; Pereda, M.D.C.V.; Nogueira, C.; Eberlin, S.; Facchini, G.; Checon, J.T.; Cesar, C.K.; Mussi, L.; Polezel, M.A.; Oliveira-Junior, D.M.; et al. Antiageing mechanisms of a standardized supercritical CO2 preparation of Black Jack (Bidens Pilosa L.) in human fibroblasts and skin fragments. Altern. Med. 2015, 2015, 280529. [Google Scholar]
- Hwang, E.; Ngo, H.T.T.; Seo, S.A.; Park, B.; Zhang, M.; Yi, T.H. Protective effect of dietary Alchemilla mollis on UVB-irradiated premature skin aging through regulation of transcription factor NFATc1 and Nrf2/ARE pathways. Phytomedicine 2018, 39, 125–136. [Google Scholar] [CrossRef]
- Ngo, H.T.T.; Hwang, E.; Seo, S.A.; Park, B.; Sun, Z.W.; Zhang, M.; Shin, Y.K.; Yi, T.H. Topical application of neem leaves prevents wrinkles formation in UVB-exposed hairless mice. J. Photochem. Photobiol. B 2017, 169, 161–170. [Google Scholar] [CrossRef]
- Zhao, P.; Alam, M.B.; Lee, S.H. Protection of UVB-induced photoaging by Fuzhuan-brick tea aqueous extract via MAPKs/Nrf2-mediated down-regulation of MMP-1. Nutrients 2019, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Panthi, V.K.; Pangeni, R.; Kim, H.J.; Park, J.W. Preparation, characterization, and biological activities of topical anti-aging ingredients in a Citrus junos callus extract. Molecules 2017, 22, 2198. [Google Scholar] [CrossRef]
- Nam, G.H.; Kawk, H.W.; Kim, S.Y.; Kim, Y.M. Solvent fraction of fermented Trapa japonica fruit extract stimulates collagen synthesis through TGF-β1/GSK-3β/β-catenin pathway in human dermal fibroblasts. J. Cosmet. Dermatol. 2020, 19, 226–233. [Google Scholar] [CrossRef]
- Jeon, H.; Kim, D.H.; Nho, Y.H.; Park, J.E.; Kim, S.N.; Choi, E.H. A mixture of Kochia scoparia and Rosa multiflora with PPAR α/γ dual agonistic effects prevents photoaging in hairless mice. Int. J. Mol. Sci. 2016, 17, 1919. [Google Scholar] [CrossRef]
- You, J.; Roh, K.B.; Li, Z.; Liu, G.; Tang, J.; Shin, S.; Park, D.; Jung, E. The antiaging properties of Andrographs paniculata by activation epidermal cell stemness. Molecules 2015, 20, 17557–17569. [Google Scholar] [CrossRef] [PubMed]
- Limtrakul, P.; Yodkeeree, S.; Thippraphan, P.; Punfa, W.; Srisomboom, J. Anti-aging and tyrosinase inhibition effects of Cassia fistula flower butanolic extract. BMC Compl. Altern. Med. 2016, 16, 497. [Google Scholar] [CrossRef] [PubMed]
- Pakravan, N.; Mahmoudi, E.; Hashemi, S.A.; Kamali, J.; Hajiaghayi, R.; Rahimzadeh, M. Cosmeceutical effect of ethyl acetate fraction of kombucha tea by intradermal administration in the skin of aged mice. J. Cosmet. Dermatol. 2017, 17, 1216–1224. [Google Scholar] [CrossRef] [PubMed]
- Bravo, K.; Duque, L.; Ferreres, F.; Moreno, D.A.; Osorio, E. Passiflora tarminiana fruits reduce UVB-induced photoaging in human skin fibroblasts. J. Photochem. Photobiol. B 2017, 168, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Cicchetti, E.; Duroure, L.; Le Borgne, E.; Laville, R. Upregulation of skin-aging biomarkers in aged NHDF cells by a sucrose ester extract from agroindustrial waste of Physalis peruviana calyces. J. Nat. Prod. 2018, 81, 1946–1955. [Google Scholar] [CrossRef]
- Jeong, D.; Lee, J.; Park, S.H.; Kim, Y.A.; Park, B.J.; Oh, J.; Sung, G.H.; Aravithan, A.; Kim, J.H.; Kang, H.; et al. Antiphotoaging and antimelanogenic effects of Penthorum chinense pursh ethanol extract due to antioxidant- and autophagy-inducing properties. Oxid. Med. Cell Long. 2019, 2019, 9679731. [Google Scholar] [CrossRef] [PubMed]
- Khare, R.; Upmanyu, N.; Jha, M. Exploring the potential of methanolic extract of Salvia officinalis against UV exposed skin aging: In vivo and in vitro model. Curr. Aging Sci. 2021, 12, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K. Protective effect of garlic on cellular senescence in UVB-exposed HaCaT human keratinocytes. Nutrient 2016, 8, 464. [Google Scholar] [CrossRef]
- Hwang, E.; Lin, P.; Ngo, T.T.; Yi, T.H. Clove attenuates UVB-induced photodamage and repairs skin barrier function in hairless mice. Food Nutr. 2018, 9, 4936–4947. [Google Scholar] [CrossRef]
- Choi, S.I.; Lee, J.S.; Lee, S.; Cho, B.Y.; Choi, S.H.; Han, X.; Sin, W.S.; Kim, Y.C.; Lee, B.Y.; Kang, I.J.; et al. Protective effects and mechanisms of Pourthiaea villosa (Thumb.) Decne. extract on hydrogen peroxide-induced skin aging in human dermal fibroblasts. J. Med. Food 2019, 22, 841–850. [Google Scholar] [CrossRef]
- Kusumawati, I.; Kurniawan, K.O.; Rullyansyah, S.; Prijo, T.A.; Widyowati, R.; Ekowati, J.; Hestianah, E.P.; Maat, S.; Matsunami, K. Anti-aging of Curcuma heyneana Valeton & Zipj: A scientific approach use in Javanese tradition. J. Ethnopharmacol. 2018, 225, 64–70. [Google Scholar] [PubMed]
- Li, J.; Lu, Y.R.; Lin, I.F.; Kang, W.; Chen, H.B.; Lu, H.F.; Wang, H.M.D. Reversing UVB-induced photoaging with Hibiscus sabdariffa calyx aqueous extract. J. Sci. Food Agric. 2020, 100, 672–681. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Oh, J.H.; Shin, M.K.; Lee, S.R. Beneficial effects of blood group antigen synthesis-increasing natural plant extracts and monosaccharides on extracellular matrix protein production in vivo. J. Dermatol. Sci. 2015, 80, 152–155. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, T.; Tsunenaga, M.; Miyanaga, M.; Ueda, U.; Ogo, M. Oral intake of lingonberry and amla fruit extract improves skin conditions in healthy female subjects: A randomized, double-blind placebo-controlled clinical trial. Biotechnol. Appl. Biochem. 2018, 66, 870–879. [Google Scholar] [CrossRef] [PubMed]
- Davinelli, S.; Bertoglio, J.C.; Polimeni, A.; Scapagnini, G. Cytroprotective polyphenols Against chronological skin aging and cutaneous photodamage. Curr. Pharmac. Design 2018, 24, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Petruk, G.; Giudice, R.D.; Rigano, M.M.; Monti, D.M. Antioxidants from plants protect against skin photoaging. Oxid. Med. Cell. Long. 2018, 2018, 1454936. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.H.; Park, J.; Jun, W.; Kim, D.; Ko, J.A.; El-Aty, A.M.A.; Choi, J.Y.; Kim, D.I.; Yang, K.Y. Transglycosylation of gallic acid by using Leuconostoc glucansucrase and its characterization as a functional cosmetic agent. AMB Express 2017, 7, 224. [Google Scholar] [CrossRef]
- Manosroi, A.; Jantrawut, P.; Akihisa, T.; Manosroi, W.; Manosroi, J. In vitro and in vivo skin anti-aging evaluation of gel containing niosomes loaded with a semi-purified fraction containing gallic acid from Terminalia chebula galls. Pharmac. Biol. 2011, 49, 1190–1203. [Google Scholar] [CrossRef]
- Goindi, S.; Guleria, A.; Aggarwal, N. development and evaluation of solid lipid nanoparticles of N-6-furfuryl adenine for prevention of photoaging. J. Biomed. Nanotechnol. 2015, 11, 1734–1746. [Google Scholar] [CrossRef]
- Choi, S.I.; Lee, J.H.; Kim, J.M.; Jung, T.D.; Cho, B.Y.; Choi, S.H.; Lee, D.W.; Kim, J.; Kim, J.Y.; Lee, O.H. Ulmus macrocarpa Hance extracts attenuated H2O2 and UVB-induced skin photo-aging by activating antioxidant enzymes and inhibiting MAPK Pathways. Int. J. Mol. Sci. 2017, 18, 1200. [Google Scholar] [CrossRef]
- Michael, J.S.; Kalirajan, A.; Padmalatha, C.; Ranjit Singh, A.J.A. In vitro antioxidant evaluation and total phenolics of methanolic extracts of Nyctanthes arbor-tristis L. Chin. J. Nat. Med. 2013, 11, 484–487. [Google Scholar] [CrossRef]
- Mohammad, I.S.; Khan, H.M.S.; Akhtar, N.; Rasool, F. Biological potential and phytochemical evaluation of Prosopis cineraria. World App. Sci. J. 2013, 27, 1489–1494. [Google Scholar]
- Dzialo, M.; Mierziak, J.; Korzun, U.; Preisner, M.; Zsopa, J.; Kulma, A. The potential of plant phenolics in prevention and therapy os skin disorders. Int. J. Mol. Sci. 2016, 17, 160. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Jung, S.H. Recent development of signaling pathways inhibitors of melanogenesis. Cell. Signal. 2017, 40, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Serre, C.; Busuttil, V.; Botto, J.M. Intrinsic and extrinsic regulations of human skin melanogenesis and pigmentation. Int. J. Cosmet. Sci. 2018, 40, 328–347. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.R.; El Chamil, C.; O’Neill, C.A.; Watson, R.E.B. Osmolyte transporter expression is reduced in photoaged human skin: Implications for skin hydration in aging. Aging Cell 2020, 19, e13058. [Google Scholar] [CrossRef]
- Boisnic, S.; Keophiphath, M.; Serandour, A.L.; Branchet, M.C.; Le Breton, S.; Lamour, I.; Gaillard, E. polar lipids from wheat extract oil improve skin damages induced by aging: Evidence from a randomized, placebo-controlled clinical trial in women and an ex vivo study on human skin explant. J. Cosmet. Dermatol. 2019, 18, 2027–2036. [Google Scholar] [CrossRef]
- Markiewicz, A.; Zasda, M.; Erkiert-Polguj, A.; Wieckowska-Szakiel, M.; Budzisz, E. An evaluation of the antiaging properties of strawberry hydrolysate treatment enriched with l-ascorbic acid applied with microneedle mesotherapy. J. Cosmet. Dermatol. 2019, 18, 129–135. [Google Scholar] [CrossRef]
- Mercurio, D.G.; Wagemaker, T.A.L.; Alves, V.M.; Benevenutto, C.G.; Gaspar, L.R.; Maia Campos, P.M.B.G. In vivo photoprotective effects of cosmetic formulations containing UV filters, vitamins, ginkgo biloba and red algae extracts. J. Photochem. Photobiol. B 2015, 153, 121–126. [Google Scholar] [CrossRef]
- An, S.; Cha, H.J.; Ko, J.M.; Han, H.; Kim, S.Y.; Kim, K.S.; Lee, S.J.; An, I.S.; Kim, S.; Youn, H.J.; et al. Kinetin improves barrier function of the skin by modulating keratinocyte differentiation markers. Ann. Dermatol. 2017, 29, 6–12. [Google Scholar] [CrossRef]
- Katekawa, E.; Caversan, J.; Mussi, L.; Camargo-Junior, F.B.; Sufi, B.; Padovani, G.; Nazato, L.; Nogueira, C.; Magalhães, W.V.; Di Stasi, L.C. Novel topical skin hydration agent containing Anadenanthera colubrine polysaccharide-standardized herbal preparation. J. Cosmet. Dermatol. 2019, 19, 1691–1698. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, V.; Potapovich, A.; Albuhaydar, A.R.; Mayer, W.; De Luca, C.; Korkina, L. Natural substances for prevention of skin photoaging: Screening systems in the development of sunscreen and rejuvenation cosmetics. Rejuvenation Res. 2018, 21, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Letsiou, S.; Kapazoglou, A.; Tsaftaris, A. Transcriptional and epigenetic effects of Vitis vinifera L. leaf extract on UV-stressed human dermal fibroblasts. Mol. Biol. Rep. 2020, 47, 5763–5772. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, C.; Santangelo, R. Ferulic acid: Pharmacological and toxicological aspects. Food Chem. Toxicol. 2014, 65, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Basak, P.; Dutta, S.; Chowdhury, S.; Sil, P.C. New insights into the ameliorative effects of ferulic acid in pathophysiological conditions. Food Chem. Toxicol. 2017, 103, 41–55. [Google Scholar] [CrossRef]
- Barone, E.; Calabrese, V.; Mancuso, C. Ferulic acid and its therapeutic potential as a hormetin for age-related diseases. Biogerontology 2009, 10, 97–108. [Google Scholar] [CrossRef]
- Kim, K.H.; Jung, J.H.; Chung, W.S.; Lee, C.H.; Jang, H.J. Ferulic acid induces keratin 6α via inhibition of nuclear β-catenin accumulation and activation of Nrf2 in wound-induced inflammation. Biomedicines 2021, 9, 459. [Google Scholar] [CrossRef]
- Calabrese, V.; Calafato, S.; Puleo, E.; Cornelius, C.; Sapienza, M.; Morganti, P.; Mancuso, C. redox regulation of cellular stress response by ferulic acid ethyl ester in human dermal fibroblasts: Role of vitagenes. Clin. Dermatol. 2008, 26, 358–363. [Google Scholar] [CrossRef]
- Hahn, H.J.; Kim, K.B.; Bae, S.; Choi, B.G.; An, S.; Ahn, K.J.; Kim, S.Y. Pretreatment of ferulic acid protects human dermal fibroblasts against ultraviolet A irradiation. Ann. Dermatol. 2016, 28, 740–748. [Google Scholar] [CrossRef]
- Zhou, Z.; Shi, T.; Hou, J.; Li, M. Ferulic acid alleviates atopic dermatitis-like symptoms in mice via its potent anti-inflammatory effect. Immunopharmacol. Immunotoxicol. 2020, 42, 156–164. [Google Scholar] [CrossRef]
- Peres, D.D.; Sarruf, F.D.; Oliveira, C.A.; Velasco, M.V.R.; Baby, A.R. Ferulic acid photoprotective properties in association with UV filters: Multifunctional sunscreen with improved SPF and UVA-PF. J. Photochem. Photobiol. B 2018, 185, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Cho, J.H.; Hong, S.H.; Kim, D.H.; Jung, H.Y.; Kang, I.K.; Cho, D.H. Whitening and anti-wrinkle activities of ferulic acid isolated from Tetragonia tetragonoides in B16F10 melanoma and CCD- 986sk fibroblast cells. J. Nat. Med. 2018, 72, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Alias, L.M.; Manoharam, S.; Vellaichamy, L.; Balakrishnan, S.; Ramachandran, C.R. Protective effect of ferulic acid on 7,12-dimethylbenz[a]anthracene-induced skin carcinogenesis in Swiss albino mice. Exp. Toxicol. Patol. 2009, 61, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Saija, A.; Tomaino, A.; Trombetta, D.; De Pasquale, A.; Uccella, N.; Barbuzzi, T.; Paolino, D.; Bonina, F. In vitro and in vivo evaluation of caffeic and ferulic acids as topical photoprotective agentes. Int. J. Pharmac. 2000, 199, 39–47. [Google Scholar] [CrossRef]
- Amic, A.; Markovic, J.M.D.; Markovic, Z.; Mienkovic, D.; Milanovic, Z.; Antonijevic, M.; Cargadová, D.M.; Pedregal, J.R.G. Theoretical study of radical inactivation, LOX inhibition, and iron chelation: The role of ferulic acid in skin protection against UVA induced oxidative stress. Antioxidants 2021, 10, 1303. [Google Scholar] [CrossRef]
- Lin, F.H.; Lin, J.Y.; Gupta, R.D.; Tournas, J.A.; Burch, J.A.; Selim, M.A.; Monteiro-Riviere, N.A.; Grichnik, J.M.; Zielinski, J.; Pinnell, S.R. Ferulic acid stabilizes a solution of vitamins C and E and doubles its photoprotection of skin. J. Investig. Dermatol. 2005, 125, 826–832. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Y.; Zhao, Z.; Qiu, J. Oxidative stress in the skin: Impact and related protection. Int. J. Cosm. Sci. 2021, 43, 495–509. [Google Scholar] [CrossRef]
- Rattan, S.I.S. Hormesis in aging. Ageing Res. Rev. 2008, 7, 63–78. [Google Scholar] [CrossRef]
- Zdunska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant properties of ferulic acid and its possible application. Skin Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef]
- Chaikul, P.; Khat-udomkiri, N.; Iangthanarat, K.; Manosroi, J.; Manosroi, A. Characteristics and in vitro anti-skin aging activity of gallic acid loaded in cationic CTAB noisome. Eur. J. Pharmac. Sci. 2019, 131, 39–49. [Google Scholar] [CrossRef]
- Alonso, C.; Martí, M.; Barba, C.; Lis, M.; Rubio, L.; Coderch, L. Skin penetration and antioxidant effects of cosmeto-textiles with gallic acid. J. Photochem. Photobiol. B 2016, 156, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Khmaladze, I.; Österlund, C.; Smiljanic, S.; Hrapovic, N.; Lafon-Kolb, V.; Amini, B.N.; Xi, L.; Fabre, S. A novel multifunctional skin care formulation with a unique blend of antipollution, brightening and antiaging active complexes. J. Cosmet. Dermatol. 2019, 19, 1415–1425. [Google Scholar] [CrossRef] [PubMed]
- Monteiro e Silva, S.A.; Calixto, G.M.F.; Cajado, J.; Carvalho, P.C.A.; Rodero, C.F.; Chorilli, M.; Leonardi, G.R. Gallic acid-lçoaded gel formulation combats skin oxidative stress: Development, characterization and ex vivo biological assays. Polymers 2017, 9, 391. [Google Scholar] [CrossRef]
- Wu, Y.Z.; Tsai, Y.Y.; Chang, L.S.; Chen, Y.J. Evaluation of gallic acid-coated gold nanoparticles as an anti-aging ingredient. Pharmaceuticals 2021, 14, 1071. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Zhou, X. Gallic acid ameliorates atopic dermatitis-like skin inflammation through immune regulation in a mouse model. Clin. Cosmet. Inv. Dermatol. 2021, 14, 1675–1683. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, X.; Wei, J.; Chen, H.; Lu, Y.; Li, K.L.; Han, L.; Lu, C. Gallic acid inhibits the expression od keratin 16 and keratin 17 through Nrf2 in psoriasis-like skin disease. Int. Immunopharmacol. 2018, 65, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, K.; Ruan, M.; Yang, J.; Gao, Z. Gallic acid inhibits fibroblasts growth and migration in keloids through the AKT/ERK signaling pathway. Acta Biochim. Biophys. Sin. 2018, 50, 1114–1120. [Google Scholar] [CrossRef]
- Chen, Y.J.; Lee, Y.C.; Huang, C.H.; Chang, L.S. Gallic acid-capped gold nanoparticles inhibit EGF-induced MMP-9 expression through suppression of p300 stabilization and NFκB/c-Jun activation in breast MDA-MB-231 cells. Toxicol. Appl. Pharmacol. 2016, 310, 98–107. [Google Scholar] [CrossRef]
- Cheng, Y.J.; Chang, L.S. Gallic acid downregulates matrix metalloproteinase-2 (MMP-2) and MMP-9 in human leukemia cells with expressed Bcr/Abl. Mol. Nutr. Food Res. 2012, 56, 1398–1402. [Google Scholar] [CrossRef]
- Zguizzato, M.; Valacchi, G.; Pecorelli, A.; Boldrini, P.; Similière, F.; Huang, N.; Cortesi, R.; Esposito, E. Gallic acid loaded poloxamer gel as a new adjuvant strategy for melanoma: A preliminary study. Colloids Surf. Biointerf. 2020, 185, 110613. [Google Scholar] [CrossRef]
- Meeran, M.F.N.; Javed, H.; Taee, H.A.; Azimullah, S.; Ojha, S.K. Pharmacological properties and molecular mechanisms of thymol: Prospects for its therapeutic potential and pharmaceutical development. Front. Pharmacol. 2017, 8, 380. [Google Scholar] [CrossRef] [PubMed]
- Hotta, M.; Nakata, R.; Katsukawa, M.; Hori, K.; Takahashi, S.; Inoue, H. Carvacrol, a component of thyme oil, activates PPARα and γ and suppresses COX-2 expression. J. Lip. Res. 2010, 51, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.I.; Jeong, N.H.; Jun, S.H.; Son, J.H.; Kim, S.; Jeon, H.; Kang, S.C.; Kim, S.H.; Lee, J.C. Thymol attenuates the worsening of atopic dermatitis induced by Staphylococcus aureus membrane vesicles. Int. Immunopharmacol. 2018, 59, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Pivetta, T.P.; Simões, S.; Araújo, M.M.; Carvalho, T.; Arruda, C.; Marcato, P.D. development of nanoparticles from natural lipids for topical delivery of thymol: Investigation of its anti-inflammatory properties. Coll. Surf. Biointerf. 2018, 164, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Folle, C.; Díaz-Carrido, N.; Sánchez-López, E.; Marqués, A.M.; Badia, J.; Baldomá, L.; Espina, M.; Calpena, A.C.; García, M.L. Surface-modified multifunctional thymol-loaded biodegradable nanoparticles for topical acne treatment. Pharmaceuticals 2021, 13, 1501. [Google Scholar] [CrossRef] [PubMed]
- Folle, C.; Marqués, A.M.; Díaz-Carrido, N.; Espina, M.; Sánchez-López, E.; Badia, J.; Baldoma, L.; Calpena, A.C.; García, M.L. Thymol-loaded PLGA nanoparticles: An efficient approach for acne treatment. J. Nanotechnol. 2021, 19, 359. [Google Scholar] [CrossRef] [PubMed]
- Aristatile, B.; Al-Assaf, A.H.; Pugalendi, K.V. Carvacrol ameliorates the PPAR-α and cytochrome P450 expression on d-galactosamine induced hepatotoxicity rats. Afr. J. Tradit. Complement. Alt. Med. 2014, 11, 118–123. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Varoni, M.; Iriti, M.; Martorell, M.; Setzer, W.N.; Contreras, M.M.; Salehi, B.; Soltani-Nejad, A.; Rajabi, S.; Tajbakhsh, M.; et al. Carvacrol and human health: A comprehensive review. Phytother. Res. 2017, 32, 1675–1687. [Google Scholar] [CrossRef]
- Suntres, Z.E.; Coccimiglio, J.; Alipour, M. The bioactivity and toxicological actions of carvacrol. Crit. Rev. Food Sci. Nutr. 2015, 55, 304–318. [Google Scholar] [CrossRef]
- Khoury, R.E.; Jubeli, R.M.; Beurouthy, M.E.; Guffroy, A.B.; Rizk, T.; Tfayli, A.; Lteif, R.J. Phytochemical screening and antityrosinase activity of carvacrol, thymoquinone, and four essential oils of Lebanese plants. Cosmet. Dermatol. 2018, 18, 944–952. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.; Xue, C.; Chen, H.; Xue, Y.; Zhao, F.; Zhu, M.X.; Cao, Z. TRPV3 enhances skin keratinocytes proliferation through EGFR-dependent signaling pathways. Cell Biol. Toxicol. 2021, 37, 313–330. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, D.H.; Park, S.J.; Kim, J.M.; Ryu, J.H. Ginseng in traditional herbal presriptions. J. Ginseng Res. 2012, 36, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Kim, K. Effect of ginseng and ginsenosides on melanogenesis and their mechanism of action. J. Ginseng Res. 2015, 38, 1–6. [Google Scholar] [CrossRef]
- Hwang, E.; Park, S.Y.; Jo, H.; Lee, D.G.; Kim, H.T.; Kim, Y.M.; Yin, C.S.; Yi, T.H. Efficacy and safety of enzyme-modified Panax ginseng for anti-wrinkle therapy in health skin: A single-center, randomized, double-blind, placebo-controlled study. Rejuvenation Res. 2015, 18, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.G.; Lee, C.C.; Dong, Z.; Lee, K.W. Ginsenosides and their metabolites: A review of their pharmacological activities in the skin. Arch. Dermatol. Res. 2015, 307, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kim, D.; Yoo, S.; Hong, Y.H.; Han, S.Y.; Jeong, S.; Jeong, D.; Kim, J.H.; Cho, J.Y.; Park, J. The skin protective effects of compound K, a metabolite of ginsenoside Rb1 from Panax ginseng. J. Ginseng Res. 2018, 42, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Xu, X.; Sun, Z.; Wang, F.; Ma, R.; Feng, K.; Li, T.; Sun, L. Protective effects of ginseng proteins on photoaging of muse fibroblasts induced by UVA. Photochem. Photobiol. 2020, 96, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Han, N.R.; Ko, S.G.; Moon, P.D.; Park, H.J. Ginsenoside Rg3 attenuates skin disorders via down-regulation of MDM2/HIF1α signaling pathway. J. Ginseng Res. 2021, 45, 610–616. [Google Scholar] [CrossRef]
- Xu, X.; Sun, G.; Liu, J.; Zhou, J.; Li, J.; Sun, Z.; Li, X.; Cheng, H.; Zhao, D.; Jiang, R.; et al. Akt activation-dependent protective effect of wild ginseng adventitious root protein against UVA-induced NIH-3T3 cell damage. Wound Repair Regen. 2021, 29, 1006–1016. [Google Scholar] [CrossRef]
- Jiang, R.; Xu, X.H.; Wang, K.; Yang, X.Z.; Bi, Y.F.; Yan, Y.; Liu, J.Z.; Chen, X.N.; Wang, Z.Z.; Gui, X.L.; et al. Ethyl acetate extract from Panax ginseng C.A. Myer and its main constituents inhibit α-melanocyte-stimulating hormone-induced melanogenesis by suppressing oxidative stress in B16 mouse melanoma cells. J. Ethnopharmacol. 2017, 208, 149–156. [Google Scholar] [CrossRef]
- Lee, J.O.; Kim, E.; Kim, J.H.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Kim, J.; Kim, S.H.; Parl, C.; Seo, D.B.; et al. Antimelanogenesis and skin-protective activities of Panax ginseng calyx ethanol extract. J. Ginseng Res. 2018, 42, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Kim, H.G.; Lee, Y.G.; Kim, J.H.; Lee, J.W.; Choi, B.R.; Jang, I.B.; Kim, G.S.; Bael, N.I. Isolation and quantification of ginsenoside Rh23, a new anti-melanogenic compound from the leaves of Panax ginseng. Molecules 2018, 23, 267. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Xiao, Y.K.; Hwang, E.; Haeng, J.J.; Yi, T.H. Antiphotoaging and antimelanogenesis properties of ginsenoside C-Y, a ginsenoside Rb2 metabolite from American ginseng PDD-ginsenoside. Photochem. Photobiol. 2019, 95, 1412–1423. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, X.; Jiang, R.; Sun, L.; Zhao, D. Vanillic acid in Panax ginseng root extract inhibits melanogenesis in B16F10 cells via inhibition of the NO/PKG signaling pathway. Biosci. Biotechnol. Biochem. 2019, 83, 1205–1215. [Google Scholar] [CrossRef]
- Lee, H.R.; Jung, J.M.; Seo, J.Y.; Chang, S.E.; Song, Y. Anti-melanogenic property of ginsenoside Rf from Panax ginseng via inhibition of CREB/MITF pathway in melanocytes and ex vivo human skin. J. Ginseng Res. 2021, 45, 555–564. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, R.; Zhou, J.; Xu, X.; Sun, Z.; Li, J.; Chen, X.; Li, Z.; Yan, X.; Zhao, D.; et al. Salicylic acid in ginseng root alleviates skin hyperpigmentation disorders by inhibiting melanogenesis and melanosome transport. Eur. J. Pharmacol. 2021, 910, 174458. [Google Scholar] [CrossRef]
- Lorz, L.R.; Kim, M.Y.; Cho, J.Y. Medicinal potential of Panax ginseng and its ginsenosides in atopic dermatitis treatment. J. Ginseng Res. 2020, 44, 8–13. [Google Scholar] [CrossRef]
- Choi, B.Y. Hair-growth potential of ginseng and its major metabolites: A review on its molecular mechanisms. Int. J. Mol. Sci. 2018, 19, 2703. [Google Scholar] [CrossRef]
- Park, K.S.; Park, D.H. The effect of Korean red ginseng on full-thickness skin wound healing in rats. J. Ginseng Res. 2019, 43, 226–235. [Google Scholar] [CrossRef]
- Faltermaier, A.; Waters, D.; Becker, T.; Arendt, E.; Gastl, M. Common wheat (Triticum aestivum L.) and its use asa brewing cereal—A review. J. Inst. Brew. 2014, 120, 1–15. [Google Scholar] [CrossRef]
- Hattarki, S.A.; Bogar, C. Triticum aestivum (wheat grass), a powerhouse plant—A review. Dent. J. Adv. Stud. 2017, 5, 25–29. [Google Scholar]
- Sutar-kapashikar, P.S.; Gawali, T.R.; Kolli, S.R.; Khot, A.S.; Dehankar, S.P.; Patil, P.D. Phenolic content in Triticum aestivum: A review. Int. J. New Technol. Res. 2018, 4, 1–2. [Google Scholar] [CrossRef]
- Lee, J.H.; Ki, H.H.; Kim, D.K.; Lee, Y.M. Triticum aestivum sprout extract attenuates 2,3-dinitrobenzene-induced atopic dermatitis-like skin lesions in mice and the expression of chemokines in human keratinocytes. Mol. Med. Rep. 2018, 18, 3461–3468. [Google Scholar] [PubMed]
- Son, D.J.; Jun, J.C.; Choi, Y.M.; Ryu, H.Y.; Lee, S.; Davis, B.A. Wheat extract oil (WEO) attenuates YVB-induced photoaging via collagen synthesis in human keratinocytes and hairless mice. Nutrients 2020, 12, 300. [Google Scholar] [CrossRef] [PubMed]
- Tito, A.; Minale, M.; Riccio, S.; Grieco, F.; Colucci, M.G.; Apone, G. A Triticum vulgare extract exhibits regenerating activity during the wound healing process. Clin. Cosmet. Investig. Dermatol. 2020, 13, 21–30. [Google Scholar] [CrossRef]
- Sahin, F.; Koçak, P.; Günes, M.Y.; Özkan, I.; Yildirim, E.; Kala, E.Y. In vitro wound healing activity of wheat-derived nanovesicles. Appl. Biochem. Biotechnol. 2019, 188, 381–394. [Google Scholar] [CrossRef]
- Sui, H.; Wang, F.; Weng, Z.; Song, H.; Fang, Y.; Tang, X.; Shen, X. A wheat germ-derived peptide YDWPGGRN facilitates skin wound-healing processes. Biochem. Biophys. Res. Commun. 2020, 524, 943–950. [Google Scholar] [CrossRef]
- Morretta, E.; D’Agostino, A.; Cassese, E.; Maglione, B.; Petrella, A.; Schiraldi, C.; Monti, M.C. Label-free quantitative proteomics to explore the action mechanism of the pharmaceutical-grade Triticum vulgare extract in speeding up keratinocyte healing. Molecules 2022, 27, 1108. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, B.; Bajpai, V. Andrographis paniculata (Brum.f.) Nees: Traditional uses, phytochemistry, pharmacological properties and quality control/quality assurance. J. Ethnopharmacol. 2021, 275, 114054. [Google Scholar] [CrossRef]
- Hossain, S.; Urbi, Z.; Sule, A.; Rahman, K.M.H. Andrographis paniculata (Burm.f.) Wall. Ex Nees: A review of ethnobotany, phytochemistry, and pharmacology. Sci. World J. 2014, 2014, 274905. [Google Scholar] [CrossRef]
- Mehta, S.; Sharma, A.K.; Sing, R.K. Pharmacological activities and molecular mechanisms of pure and crude extract of Andrographis paniculate: An update. Phytomedicine Plus 2021, 1, 100085. [Google Scholar] [CrossRef]
- Fardiyah, Q.; Ersam, T.; Suyanta; Slamet, A.; Suprapto; Kurniawan, F. New potential and characterization of Andrographis paniculata L. Ness plant extracts as photoprotective agent. Arab. J. Chem. 2020, 13, 8888–8897. [Google Scholar] [CrossRef]







| Compounds | Test | Effects | Refs |
|---|---|---|---|
| α-mangostin from Garcinia mangostana | In vitro (keratinocytes) In vivo (hairless mice) | Downregulation of UV-induced MMP-1/MMP-9 Reduction in UV-induced wrinkles | [19] |
| Batatasin III from Dendrobium loddigesii | In vitro (fibroblasts) | Increase in collagen synthesis | [20] |
| Ferulic acid from several plant origin | Clinical trial | Enhancement of skin elasticity by bleaching, anti-redness, smoothing, and moisturizing activities | [25] |
| Jasmonate derivative | In vitro (keratinocytes) | Upregulation of proteoglycans Increase in glycosaminoglycan production | [23] |
| Limonoids from Carapa guianensis | In vitro (fibroblasts) | Increase in collagen synthesis | [21] |
| Polysaccharides from Panax ginseng | In vitro (keratinocytes) | Downregulated UV-induced MMP-1 expression | [24] |
| Triterpenoids from Eriobotrya japonica | In vitro (fibroblasts) | Increase in collagen synthesis | [22] |
| Plants | Effects | Refs |
|---|---|---|
| Alchemilla mollis | DPPH and ABTS radical scavenging activity | [36] |
| Allium sativum | DPPH radical scavenging activity Reduction in NO production Decrease in UVB-induced ROS generation | [49] |
| Camelia sinensis | DPPH, ABTS, CUPRAC, FRAP, and ORAC radical scavenging activity Decrease in UVB-induced ROS generation in fibroblasts Upregulation of SOD, CAT, and GPX in fibroblasts Increase in Nrf2 transcriptional level and nuclear translocation in fibroblasts | [38] |
| Cassia fistula | DPPH and ABTS radical scavenging activity | [43] |
| Curcuma heyneana | DPPH radical scavenging activity | [52] |
| Gastrodia elata | DPPH and ABTS radical scavenging activity Increase in SOD activity | [27] |
| Hibiscus sabdariffa | DPPH and FRAP radical scavenging activity Decrease in UVB-induced ROS generation | [53] |
| Litchi chinensis | DPPH, superoxide, and ABTS radical scavenging activity | [28] |
| Magnolia officinalis | DPPH and FRAP radical scavenging activity | [29] |
| Malaxis acuminata | DPPH and ABTS radical scavenging activity | [30] |
| Manikaria zapota | DPPH and ABTS radical scavenging activity | [14] |
| Nephelium lappaceum | DPPH, superoxide, and ABTS radical scavenging activity | [28] |
| Nictanthes arbor-tristis | DPPH radical scavenging activity Reduction in UVB-induced ROS production in human fibroblasts | [62] |
| Passiflora tarminiana | ORAC radical scavenging activity | [45] |
| Penthorum chinense | DPPH radical scavenging activity | [47] |
| Phyllanthus emblica | DPPH and ABTS radical scavenging activity | [14] |
| Piper cambodianum | Reduction in UVB-induced ROS production in human fibroblasts | [34] |
| Pourthiaea villosa | DPPH and ABTS+ radical scavenging activity Inhibition of H2O2-induced intracellular ROS production Increase in SOD1 and SOD2 proteins levels Inhibition of H2O2-induced premature cellular senescence | [51] |
| Prosopis cineraria | DPPH radical scavenging activity | [63] |
| Salvia officinalis | DPPH radical scavenging activity | [48] |
| Spatholobus suberectus | Reduction in UVB-induced ROS production in human fibroblasts | [32] |
| Silybum marianum | DPPH and ABTS radical scavenging activity | [14] |
| Syzygium aromaticum | Reduction in UVB-induced ROS production in human fibroblasts | [50] |
| Tamarindus indica | DPPH, superoxide, and ABTS radical scavenging activity | [28] |
| Ulmus macrocarpa | DPPH and ABTS+ radical scavenging activity Inhibition of H2O2-induced intracellular ROS production Increase in SOD1, SOD2, and protein levels Inhibition of H2O2-induced premature cellular senescence | [61] |
| Plants | Effects | Refs |
|---|---|---|
| Cassia fistula | In vitro inhibition of l-DOPA oxidation by tyrosinase | [43] |
| Citrus junus | In vitro inhibition of l-DOPA oxidation by tyrosinase Inhibition of melanin synthesis in melanoma cells | [39] |
| Curcuma heyneana | In vitro inhibition of l-DOPA oxidation by tyrosinase | [52] |
| Eugenia dysenterica | In vitro inhibition of l-DOPA oxidation by tyrosinase | [26] |
| Hibiscus sabdariffa | In vitro inhibition of tyrosinase activity Downregulation of MITF, tyrosinase, TRP-1, and TRP-2 gene expression Reduction in MITF, tyrosinase, TRP-1, and TRP-2 levels | [53] |
| Litchi chinensis | Reduction in melanin content in human fibroblasts Inhibition of tyrosinase activity in melanoma cells Inhibition of TRP-2 activity in melanoma cells | [28] |
| Magnolia officinalis | In vitro reduction in melanin content in CCD-966KS cells | [29] |
| Malaxis acuminata | In vitro inhibition of l-DOPA oxidation by tyrosinase | [30] |
| Penthorum chinense | Inhibition of melanin content in B16F10 cells | [47] |
| Prosopis cineraria | In vitro inhibition of l-DOPA oxidation by tyrosinase Reduction in melanin content in a clinical trial | [63] |
| Tamarindus indica | In vitro inhibition of melanin synthesis in melanoma cells Decrease in tyrosine mRNA and protein level | [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, E.F.; Magalhães, W.V.; Di Stasi, L.C. Recent Advances in Herbal-Derived Products with Skin Anti-Aging Properties and Cosmetic Applications. Molecules 2022, 27, 7518. https://doi.org/10.3390/molecules27217518
Costa EF, Magalhães WV, Di Stasi LC. Recent Advances in Herbal-Derived Products with Skin Anti-Aging Properties and Cosmetic Applications. Molecules. 2022; 27(21):7518. https://doi.org/10.3390/molecules27217518
Chicago/Turabian StyleCosta, Erika F., Wagner V. Magalhães, and Luiz C. Di Stasi. 2022. "Recent Advances in Herbal-Derived Products with Skin Anti-Aging Properties and Cosmetic Applications" Molecules 27, no. 21: 7518. https://doi.org/10.3390/molecules27217518
APA StyleCosta, E. F., Magalhães, W. V., & Di Stasi, L. C. (2022). Recent Advances in Herbal-Derived Products with Skin Anti-Aging Properties and Cosmetic Applications. Molecules, 27(21), 7518. https://doi.org/10.3390/molecules27217518