



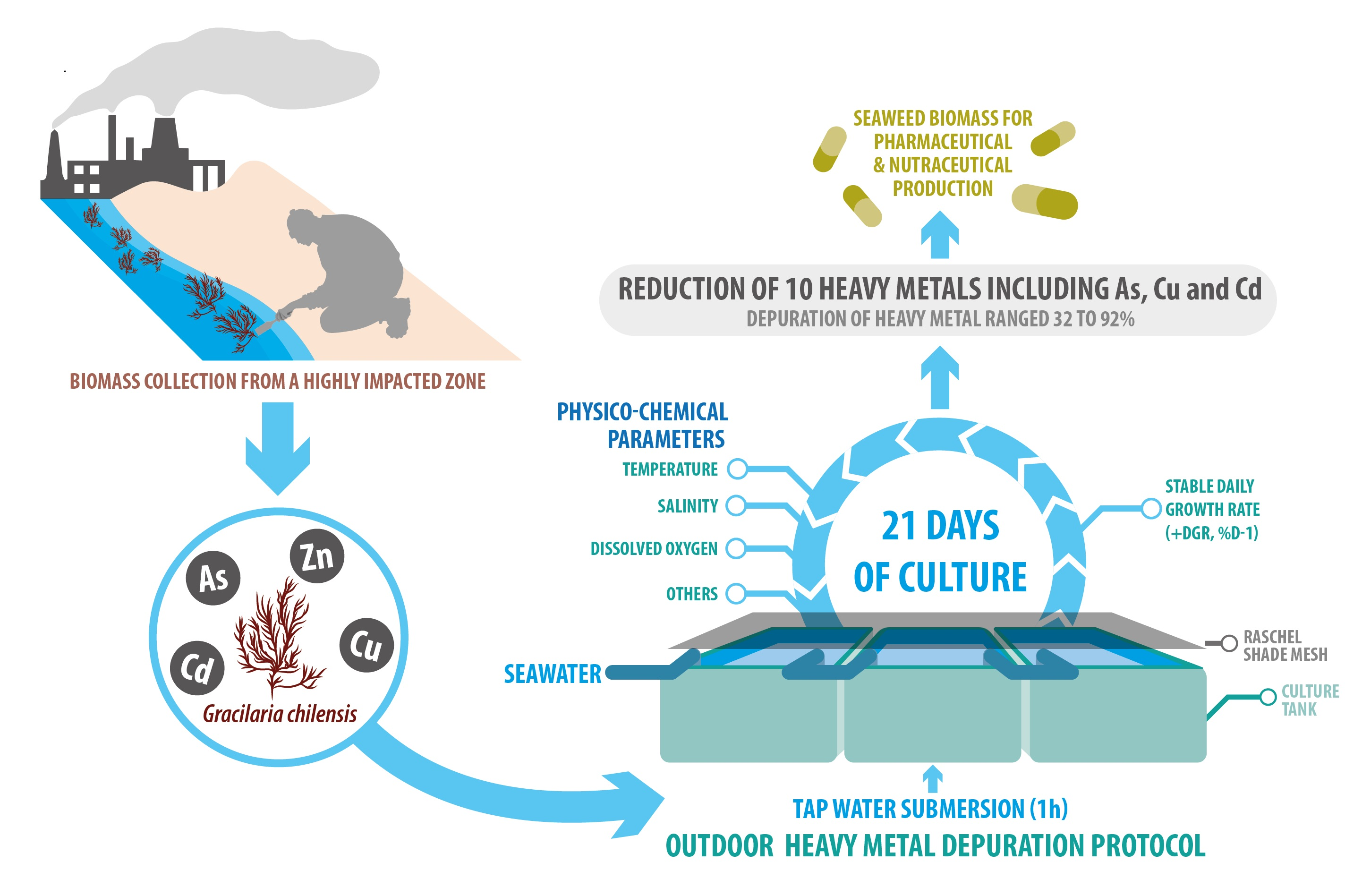
Heavy Metal Depuration Steps for Gracilaria chilensis in Outdoor Culture Systems

 ,
, 

Abstract

1. Introduction
2. Results
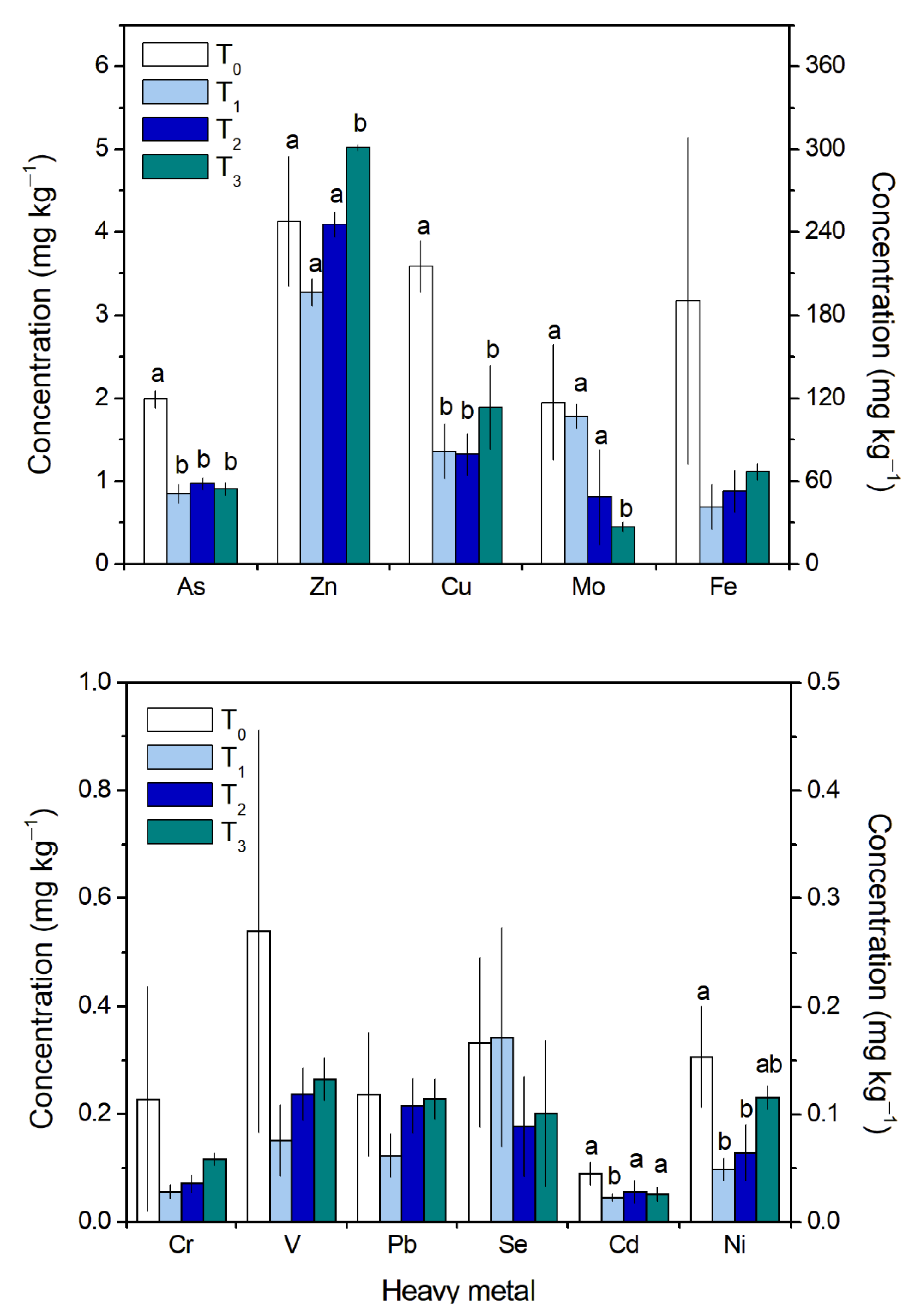
2.1. Heavy Metal Bioconcentration in Gracilaria chilensis Biomass
2.2. Percentage of Depuration of Heavy Metals
2.3. Normative Comparison
2.4. Daily Growth Rates of Gracilaria chilensis Biomass
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Seaweed Collection
5.2. Experimental Setup, Open-System Culture Conditions, and Measured Parameters
5.3. Percentage of Depuration (PD) of Heavy Metals
5.4. Daily Growth Rate (DGR)
5.5. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Anis, M.; Ahmed, S.; Hasan, M.M. Algae as nutrition, medicine and cosmetic: The forgotten history, present status and future trends. World J. Pharm. Pharm. Sci. 2017, 6, 1934–1959. [Google Scholar] [CrossRef]
- Ganesan, A.R.; Tiwari, U.; Rajauria, G. Seaweed nutraceuticals and their therapeutic role in disease prevention. Food Sci. Hum. Wellness 2019, 8, 252–263. [Google Scholar] [CrossRef]
- Rimmer, M.A.; Larson, S.; Lapong, I.; Purnomo, A.H.; Pong-Masak, P.R.; Swanepoel, L.; Paul, N.A. Seaweed Aquaculture in Indonesia Contributes to Social and Economic Aspects of Livelihoods and Community Wellbeing. Sustainability. 2021, 13, 10946. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture; Towards Blue Transformation; Food & Agriculture Organization: Rome, Italy, 2022; p. 45. [Google Scholar] [CrossRef]
- SERNAPESCA. Anuario Estadístico de Pesca y Acuicultura. Servicio Nacional de Pesca, Valparaíso. 2021. Available online: http://www.sernapesca.cl/informacion-utilidad/anuarios-estadisticos-de-pesca-y-acuicultura. (accessed on 25 May 2022).
- Arakaki, N.; Schmidt, W.E.; Carbajal, P.; Fredericq, S. First occurrence of Gracilaria chilensis, and distribution of Gracilariopsis lemaneiformis (Gracilariaceae, Gracilariales) in Peru on the basis of rbcL sequence analysis. Phytotaxa. 2015, 208, 175–181. [Google Scholar] [CrossRef]
- Brito, N. El Oro Negro de Pitipalena. In Rescate Patrimonial de la Extracción del Pelillo (Gracilaria chilensis) en Puerto Raúl Marín Balmaceda; Ediciones Ñire Negro Coyhaique: Coyhaique, Chile, 2019; p. 78. [Google Scholar]
- Dillehay, T.D.; Ramírez, C.; Collins, M.B.; Rossen, J.; Pino-Navarro, J.D. Monte Verde: Seaweed, food, medicine, and the peopling of South America. Science 2008, 320, 784–786. [Google Scholar] [CrossRef]
- Torres, P.; Santos, J.P.; Chow, F.; dos Santos, D.Y.A.C. A comprehensive review of traditional uses, bioactivity potential, and chemical diversity of the genus Gracilaria (Gracilariales, Rhodophyta). Algal Res. 2019, 37, 288–306. [Google Scholar] [CrossRef]
- Le, Y.; Wang, B.; Xue, M. Nutraceuticals use and type 2 diabetes mellitus. Curr. Opin. Pharmacol. 2022, 62, 168–176. [Google Scholar] [CrossRef]
- Rajasekaran, A. 1.05—Nutraceuticals. In Comprehensive Medicinal Chemistry III; Chackalamannil, S., Rotella, D., Ward, S.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 107–134. [Google Scholar]
- Hafting, J.T.; Craigie, J.S.; Stengel, D.B.; Loureiro, R.R.; Buschmann, A.H.; Yarish, C.; Edwards, M.D.; Critchley, A.T. Prospects and challenges for industrial production of seaweed bioactives. J. Phycol. 2015, 51, 821–837. [Google Scholar] [CrossRef]
- Pinto, C.; Ibáñez, M.R.; Loyola, G.; León, L.; Salvatore, Y.; González, C.; Barraza, V.; Castañeda, F.; Aldunate, R.; Contreras-Porcia, L.; et al. Characterization of an Agarophyton chilense Oleoresin Containing PPARgamma Natural Ligands with Insulin-Sensitizing Effects in a C57Bl/6J Mouse Model of Diet-Induced Obesity and Antioxidant Activity in Caenorhabditis elegans. Nutrients 2021, 13, 1828. [Google Scholar] [CrossRef]
- Guillemin, M.-L.; Faugeron, S.; Destombe, C.; Viard, F.; Correa, J.A.; Valero, M. Genetic Variation in Wild and Cultivated Population of the Haploid-diploid red Alga Gracilaria chilensis: How farming practices favor asexual reproduction and heterozygosity. Evolution 2008, 62, 1500–1519. [Google Scholar] [CrossRef]
- Westermeier, R.; Rivera, P.J.; Gomez, I. El uso de mangas de polietileno como sustrato en el repoblamiento de Gracilaria sp. (Rhodophyta, Gigartinales) en el sur de Chile. Gayana 1988, 45, 95–106. [Google Scholar]
- Avila, M.; Aroca, G.; Rodríguez, D.; Riquelme, R.; Piel, M.I.; Ramírez, M.E.; De Zarate, C. Manual de Buenas Prácticas para el Cultivo de Pelillo (Agarophyton chilensis ex Gracilaria chilensis); Universidad Arturo Prat, ICYT: Iquique, Chile, 2019; pp. 1–11. [Google Scholar]
- Saavedra, S.; Henríquez, L.; Leal, P.; Galleguillos, F.; Cook, S.; Cárcamo, F. Cultivo de Macroalgas: Diversificación de la Acuicultura de Pequeña Escala en Chile; Convenio de Desempeño, Subsecretaría de Economía y Empresas de Menor Tamaño; Instituto de Fomento Pesquero: Valparaíso, Chile, 2019; pp. 1–106. [Google Scholar]
- Campbell, I.; Macleod, A.; Sahlmann, C.; Neves, L.; Funderud, J.; Øverland, M.; Hughes, A.D.; Stanley, M. The Environmental Risks Associated with the Development of Seaweed Farming in Europe—Prioritizing Key Knowledge Gaps. Front. Mar. Sci. 2019, 6, 107. [Google Scholar] [CrossRef]
- Walkinshaw, C.; Lindeque, P.K.; Thompson, R.; Tolhurst, T.; Cole, M. Microplastics and seafood: Lower trophic organisms at highest risk of contamination. Ecotoxicol. Environ. Saf. 2020, 190, 110066. [Google Scholar] [CrossRef]
- Yokoya, N.; Oliveira, E.C. Temperature responses of economically important red algae and their potential for mariculture in Brazilian waters. J. Appl. Phycol. 1992, 4, 339–345. [Google Scholar] [CrossRef]
- Yokoya, N.; Oliveira, E.C. Effects of salinity on the growth rate, morphology and water content of some Brazilian red algae of economic importance. Cienc. Mar. 1992, 18, 49–64. [Google Scholar] [CrossRef][Green Version]
- Rivas, J.; Núñez, A.; Piña, F.; Erazo, F.; Castañeda, F.; Araya, M.; Meynard, A.; Contreras-Porcia, L. Indoor culture scaling of Gracilaria chilensis (Florideophyceae, Rhodophyta): The effects of nutrients by means of different culture media. Revista de Biología Marina y Oceanografía 2021, 56, 186–199. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V. Seaweed cultivation: Methods and problems. Russ. J. Mar. Biol. 2010, 36, 227–242. [Google Scholar] [CrossRef]
- Rees, A. Safety factors and nutrient uptake by seaweeds. Mar. Ecol. Prog. Ser. 2003, 263, 29–42. [Google Scholar] [CrossRef]
- Harrison, P.; Hurd, C. Nutrient physiology of seaweeds: Application of concepts to aquaculture. Cah. Biol. Mar. 2001, 42, 71–82. [Google Scholar]
- Reef, R.; Pandolfi, J.M.; Lovelock, C. The effect of nutrient enrichment on the growth, nucleid acid concentrations, and elemental stoichiometry of coral reef macroalgae. Ecol. Evol. 2012, 2, 1985–1995. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E. Bioaccumulation of non-essential hazardous heavy metals and metalloids in freshwater fish. Risk to human health. Environ. Chem. Lett. 2018, 16, 903–917. [Google Scholar] [CrossRef]
- Latorre-Padilla, N.; Meynard, A.; Oyarzun, F.X.; Contreras-Porcia, L. Ingestion of contaminated kelps by the herbivore Tetraphygus niger: Negative effects on food intake, growth, fertility, and early development. Mar. Pollut. Bull. 2021, 167, 112365. [Google Scholar] [CrossRef]
- Latorre-Padilla, N.; Meynard, A.; Rivas, J.; Contreras-Porcia, L. Transfer of Pollutants from Macrocystis pyrifera to Tetraphygus niger in a Highly Impacted Coastal Zone of Chile. Toxics 2021, 9, 244. [Google Scholar] [CrossRef]
- Peng, Z.; Guo, Z.; Wang, Z.; Zhang, R.; Wu, Q.; Gao, H.; Wang, Y.; She, Z.; Lek, S.; Xiao, J. Species-specific bioaccumulation and health risk assessment of heavy metal in seaweeds in tropic coasts of South China Sea. Sci. Total Environ. 2022, 832, 155031. [Google Scholar] [CrossRef]
- Garnham, G.W.; Codd, G.A.; Gadd, G.M. Accumulation of Cobalt, Zinc and Manganese by the Estuarine Green Microalga Chlorella salina Inmobilized in Alginate Microbreads. Environ. Sci. Technol. 1992, 26, 1764–1770. [Google Scholar] [CrossRef]
- Andrade, S.; Contreras, L.; Moffett, J.W.; Correa, J.A. Kinetics of copper accumulation in Lessonia nigrescens (Phaeophyceae) under conditions of environmental oxidative stress. Aquat. Toxicol. 2006, 78, 398–401. [Google Scholar] [CrossRef]
- Contreras, L.; Moenne, A.; Correa, J.A. Antioxidant responses in Scytosiphon lomentaria (Phaeophyceae) Inhabiting Cooper-Enriched coastal environments. J. Phycol. 2005, 41, 1184–1195. [Google Scholar] [CrossRef]
- Sordet, C.; Contreras-Porcia, L.; Lovazzano, C.; Goulitquer, S.; Andrade, S.; Potin, P.; Correa, J.A. Physiological plasticity of Dictyota kunthii (Phaeophyceae) to Cooper excess. Aquat. Toxicol. 2014, 150, 220–228. [Google Scholar] [CrossRef]
- Contreras-Porcia, L.; Meynard, A.; López-Cristoffanini, C.; Latorre, N.; Kumar, M. Marine Metal Pollution and Effects on Seaweed Species. Syst. Biol. Mar. Ecosyst. 2017, 1, 35–48. [Google Scholar] [CrossRef]
- Mohamed, S.F.; Borik, R.M. Modern Trends in Using Marine Algae for Treatment of Aquatic Pollution. Int. J. ChemTech Res. 2013, 5, 2863–2889. [Google Scholar]
- Araya, M.; Rivas, J.; Sepúlveda, G.; Espinoza-González, C.; Lira, S.; Meynard, A.; Blanco, E.; Escalona, N.; Ginocchio, R.; Garrido-Ramírez, E.; et al. Effect of Pyrolysis Temperature on Copper Aqueous Removal Capability of Biochar Derived from the Kelp Macrocystis pyrifera. Appl. Sci. 2021, 11, 9223. [Google Scholar] [CrossRef]
- Sudharsan, S.; Seedevi, P.; Ramasamy, P.; Subhapradha, N.; Vairamani, S.; Shanmugam, A. Heavy metal accumulation in seaweeds and sea grasses along southeast coast of India. J. Chem. Pharm. Res. 2012, 4, 4240–4244. [Google Scholar]
- Wang, Z.; Wang, X.; Ke, C. Bioaccumulation of trace metals by the live macroalga Gracilaria lemaneiformis. J. Appl. Phycol. 2013, 26, 1889–1897. [Google Scholar] [CrossRef]
- Luo, H.; Wang, Q.; Zhang, C.; Zhang, L.; Yang, Y. Bioaccumulation and release of heavy metals during growth and decomposition of cultivated Gracilaria lemaneiformis. Mar. Pollut. Bull. 2021, 173, 113130. [Google Scholar] [CrossRef]
- Hu, S.; Tang, C.H.; Wu, M. Cadmium accumulation by several seaweeds. Sci. Total Environ. 1996, 187, 65–71. [Google Scholar] [CrossRef]
- Sun, X.; Liu, Z.; Jiang, Q.; Yang, Y. Concentrations of various elements in seaweed and seawater from Shen’ao bay, Nan’ao Island, Guangdong coast, China: Environmental monitoring and the bioremediation potential of the seaweed. Sci. Total Environ. 2019, 659, 632–639. [Google Scholar] [CrossRef]
- Oyarzo-Miranda, C.; Latorre, N.; Meynard, A.; Rivas, J.; Bulboa, C.; Contreras-Porcia, L. Coastal pollution from the industrial park Quintero Bay of central Chile: Effects on abundance, morphology, and development of the kelp Lessonia spicata (Phaeophyceae). PLoS ONE 2020, 15, e0240581. [Google Scholar] [CrossRef]
- Jara-Yáñez, R.; Meynard, A.; Acosta, G.; Latorre-Padilla, N.; Oyarzo-Miranda, C.; Castañeda, F.; Piña, F.; Rivas, J.; Bulboa, C.; Contreras-Porcia, L. Negative consequences on the Growth, Morphometry and Community Structure of the Kelp Macrocystis pyrifera (Phaeophyceae, Ochrophyta) by a Short Pollution Pulse of Heavy Metals and PAHs. Toxics 2021, 9, 190. [Google Scholar] [CrossRef]
- Ali, A.Y.A.; Idris, M.A.; Eltayeb, M.A.H.; El-Zahhar, A.A.; Ashraf, I.M. Bioaccumulation and health risk assessment of toxic metals in red algae in Sudanese Red Sea coast. Toxin Rev. 2021, 40, 1327–1337. [Google Scholar] [CrossRef]
- Benabdallah, N.; Harrache, D.; Mir, A.; de la Guardia, M.; Benhachem, F.Z. Bioaccumulation of trace metals by red alga Corallina elongata in the coast of Beni Saf, west coast, Algeria. Chem. Int. 2017, 3, 320–331. [Google Scholar]
- Foday, E.H., Jr.; Bo, B.; Xu, X. Removal of Toxic Heavy Metals from Contaminated Aqueous Solutions Using Seaweeds: A Review. Sustainability 2021, 13, 12311. [Google Scholar] [CrossRef]
- Stengel, D.B.; Macken, A.; Morrison, L.; Morley, N. Zinc concentrations in marine macroalgae and a lichen from western Ireland in relation to phylogenetic grouping, habitat and morphology. Mar. Pollut. Bull. 2004, 48, 902–909. [Google Scholar] [CrossRef]
- Anilkumar, B.; Babu, N.; Kavitha, G. Biosorption of Zinc on to Gracilaria corticata (Red Algae) Powder and Optimization using Central Composite Design. J. Appl. Sci. Eng. Methodol. 2016, 2, 412–425. [Google Scholar]
- Rosemary, T.; Arulkumar, A.; Paramasivam, S.; Mondragon-Portocarrero, A.; Miranda, J.M. Biochemical, Micronutrient and Physicochemical Properties of the Dried Red Seaweeds Gracilaroa edulis and Gracilaria corticata. Molecules 2019, 24, 2225. [Google Scholar] [CrossRef]
- Jayasankar, R.; Paliwal, K. Seasonal variation in the elemental composition of Gracilaria species of the Gulf of Mannar, Tamil Nadu coast. Seaweed Res. Util. 2002, 24, 55–59. [Google Scholar]
- García-Casal, M.N.; Pereira, A.C.; Leets, I.; Ramírez, J.; Quiroga, M.F. High Iron Content and Bioavailability in Humans from Four Species of Marine Algae. J. Nutr. 2007, 137, 2691–2695. [Google Scholar] [CrossRef]
- Evans, L.K.; Edwards, M.S. Bioaccumulation of cooper and zinc by the giant kelp Macrocystis pyrifera. Algae 2011, 26, 265–275. [Google Scholar] [CrossRef]
- Lares, M.L.; Flores-Muñoz, G.; Lara-Lara, R. Temporal variability of bioavaliable Cd, Hg, Zn, Mn and Al in an upwelling regime. Environ. Pollut. 2002, 120, 595–608. [Google Scholar] [CrossRef]
- Wang, W.-X.; Dei, R.C.H. Kinetic measurements of metal accumulation in two marine macroalgae. Mar. Biol. 1999, 135, 11–23. [Google Scholar] [CrossRef]
- Capo, T.R.; Jaramillo, J.C.; Boyd, A.E.; Lapointe, B.E.; Serafy, J.E. Sustained high yields of Gracilaria (Rhodophyta) grown in intensive large-scale culture. J. Appl. Phycol. 1999, 11, 143–147. [Google Scholar] [CrossRef]
- Yu, J.; Yang, J.-F. Physiological and biochemical response of seaweed Gracilaria lemaneiformis to concentration changes of N and P. J. Exp. Mar. Biol. Ecol. 2008, 367, 142–148. [Google Scholar] [CrossRef]
- Larsson, N. The brutal reality of life inside one of the world’s most polluted cities. Wired Magazine. 2020. Available online: https://www.wired.co.uk/article/chile-quintero-pollution (accessed on 1 April 2022).
- Gayo, E.M.; Muñoz, A.A.; Maldonado, A.; Lavergne, C.; Francois, J.P.; Rodríguez, D.; Klock-Barría, K.; Sheppard, P.R.; Aguilera-Betti, I.; Alonso-Hernández, C.; et al. A Cross-Cutting Approach for Relating Anthropocene, Environmental Injustice and Sacrifice Zones. Earth’s Future 2022, 10, e2021EF002217. [Google Scholar] [CrossRef]
- Yong, Y.S.; Yong, W.T.L.; Anton, A. Analysis of formulae for determination of seaweed growth rate. J. Appl. Phycol. 2013, 25, 1831–1834. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Ranges of Percentage of Depuration | ||||||
|---|---|---|---|---|---|---|
| T1 | T2 | T3 | ||||
| Min | Max | Min | Max | Min | Max | |
| As | 54 | 60 | 49 | 55 | 53 | 57 |
| Zn | −5 | 32 | −25 | 18 | −4 | −55 |
| Cu | 54 | 67 | 58 | 69 | 36 | 56 |
| Cr | −38 | 90 | −29 | 87 | −106 | 73 |
| Mo | −45 | 39 | 20 | 94 | 69 | 81 |
| Pb | −32 | 75 | −116 | 48 | −158 | 33 |
| Se | −84 | 39 | −38 | 76 | 30 | 56 |
| V | 8 | 92 | 1 | 78 | −43 | 74 |
| Fe | 54 | 92 | 42 | 87 | 41 | 81 |
| Cd | 31 | 58 | 13 | 50 | 37 | 51 |
| Ni | 65 | 71 | 34 | 72 | −6 | 40 |
| Metal | Concentration at 21 Days (mg kg−1) * | Maximum Metal Levels According to Canadian Guidelines for Chemical Contaminants and Toxins in Fish and Fish Products | Maximum Metal Levels According to Commission Regulation (EC) No. 1881/2006, European Union | Maximum Metal Levels According to CXS_193e (Codex Alimentarius) | Decree 997 Approves Food Health Regulations (Chile) |
|---|---|---|---|---|---|
| As | 0.90 ± 0.07 | 3.5 a | n.i. | 0.1–0.5 c | 0.01–0.5 e |
| Zn | 5.02 ± 0.03 | n.i. | n.i. | n.i. | 100.0 f |
| Cu | 1.9 ± 0.5 | n.i. | n.i. | n.i. | 10.0 f |
| Cr | 0.12 ± 0.01 | n.i. | n.i. | n.i. | n.i. |
| Mo | 0.45 ± 0.05 | n.i. | n.i. | n.i. | n.i. |
| Pb | 0.23 ± 0.04 | 0.5 a | 0.3–1.5 b | 0.3 d | 2.0 g |
| Se | 0.20 ± 0.01 | n.i. | n.i. | n.i. | 0.30 h |
| V | 0.27 ± 0.04 | n.i. | n.i. | n.i. | n.i. |
| Fe | 66.8 ± 5.7 | n.i. | n.i. | n.i. | 0.2–50 e |
| Cd | ± 0.01 | n.i. | 0.05–1.0 b | 2.0 d | 0.01–0.5 i |
| Ni | 0.12 ± 0.01 | n.i. | n.i. | n.i. | n.i. |
| DF | Sum Sq | Mean Sq | F | p Value | |
|---|---|---|---|---|---|
| Time (days of culture) | 2 | 0.339 | 0.1694 | 0.185 | 0.84 |
| Residuals | 6 | 5.504 | 0.9174 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivas, J.; Piña, F.; Araya, M.; Latorre-Padilla, N.; Pinilla-Rojas, B.; Caroca, S.; Bronfman, F.C.; Contreras-Porcia, L. Heavy Metal Depuration Steps for Gracilaria chilensis in Outdoor Culture Systems. Molecules 2022, 27, 6832. https://doi.org/10.3390/molecules27206832
Rivas J, Piña F, Araya M, Latorre-Padilla N, Pinilla-Rojas B, Caroca S, Bronfman FC, Contreras-Porcia L. Heavy Metal Depuration Steps for Gracilaria chilensis in Outdoor Culture Systems. Molecules. 2022; 27(20):6832. https://doi.org/10.3390/molecules27206832
Chicago/Turabian StyleRivas, Jorge, Florentina Piña, Matías Araya, Nicolás Latorre-Padilla, Benjamín Pinilla-Rojas, Sofía Caroca, Francisca C. Bronfman, and Loretto Contreras-Porcia. 2022. "Heavy Metal Depuration Steps for Gracilaria chilensis in Outdoor Culture Systems" Molecules 27, no. 20: 6832. https://doi.org/10.3390/molecules27206832
APA StyleRivas, J., Piña, F., Araya, M., Latorre-Padilla, N., Pinilla-Rojas, B., Caroca, S., Bronfman, F. C., & Contreras-Porcia, L. (2022). Heavy Metal Depuration Steps for Gracilaria chilensis in Outdoor Culture Systems. Molecules, 27(20), 6832. https://doi.org/10.3390/molecules27206832