
Evaluation of Antibacterial and Antifungal Properties of Low Molecular Weight Chitosan Extracted from Hermetia illucens Relative to Crab Chitosan

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Obtaining Chitin and Chitosan
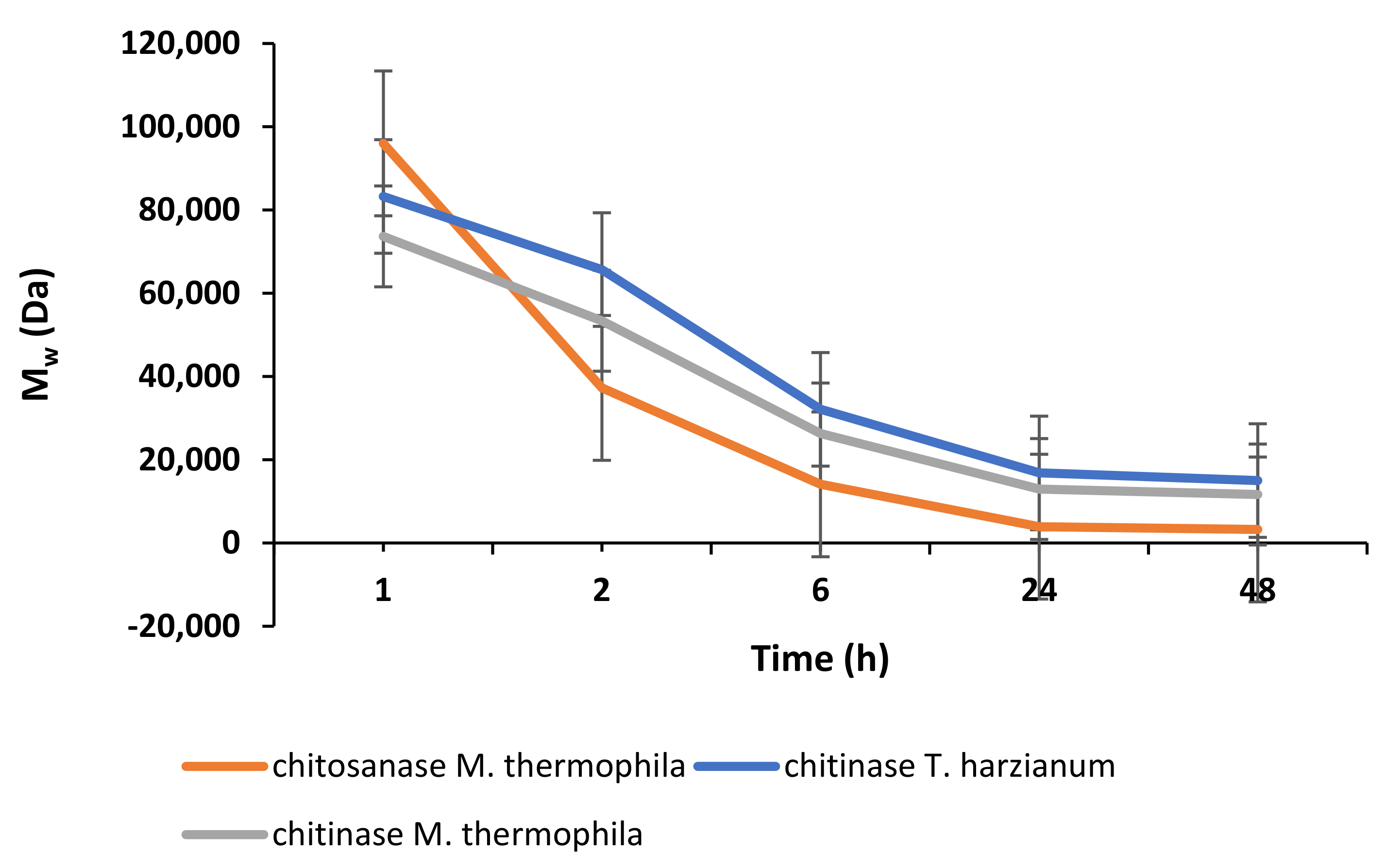
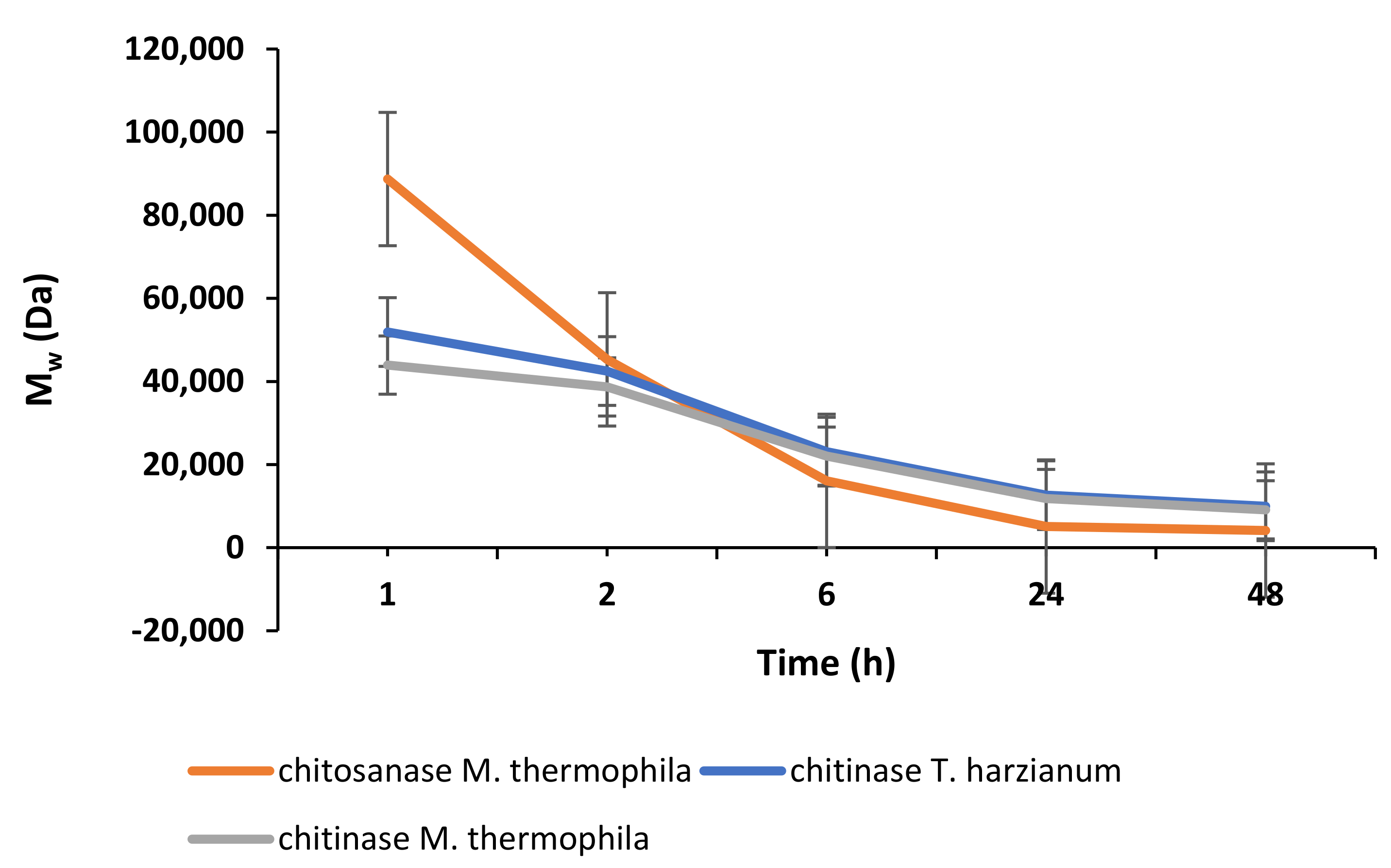
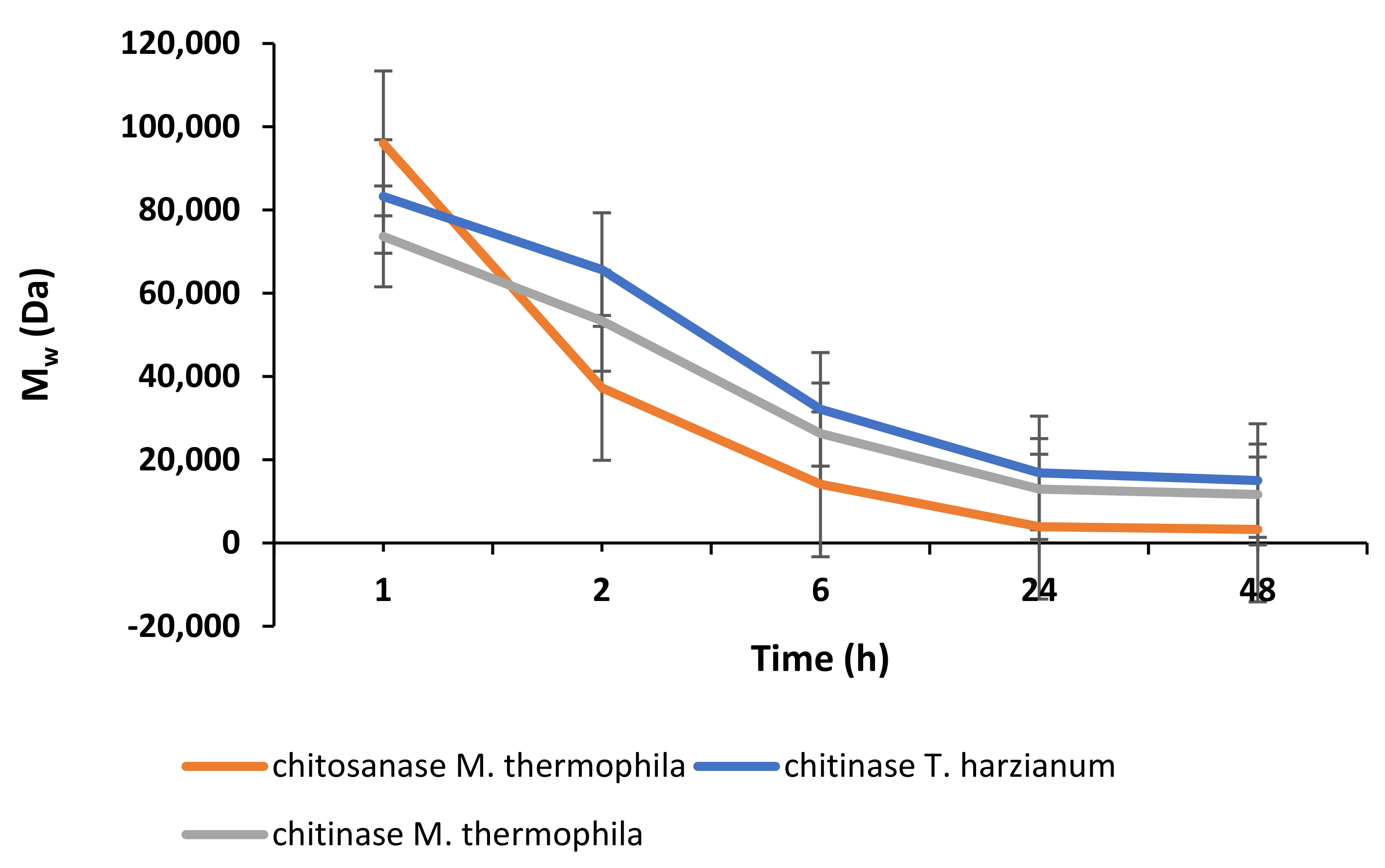
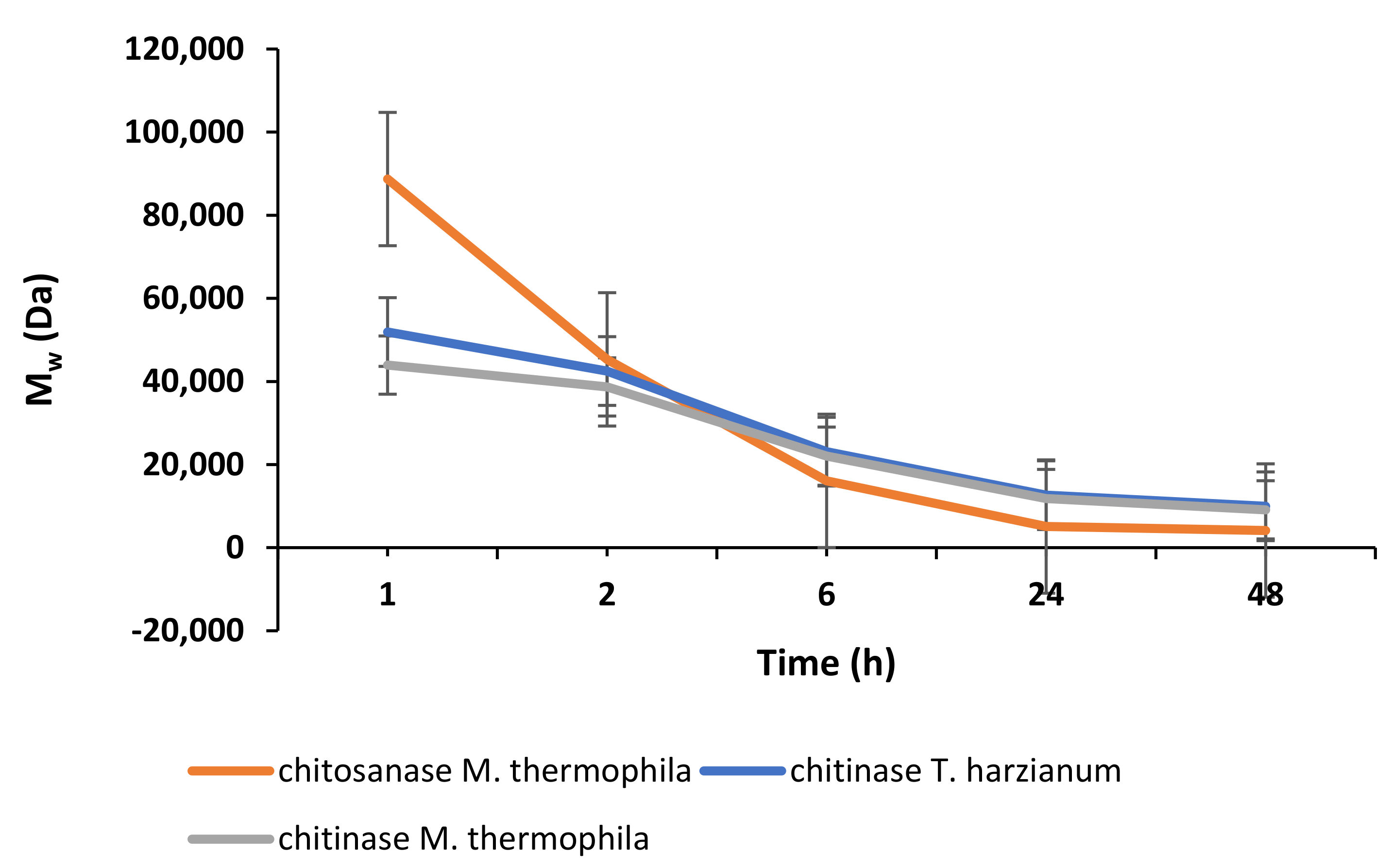
2.2. Enzymatic Depolymerisation of Chitosan
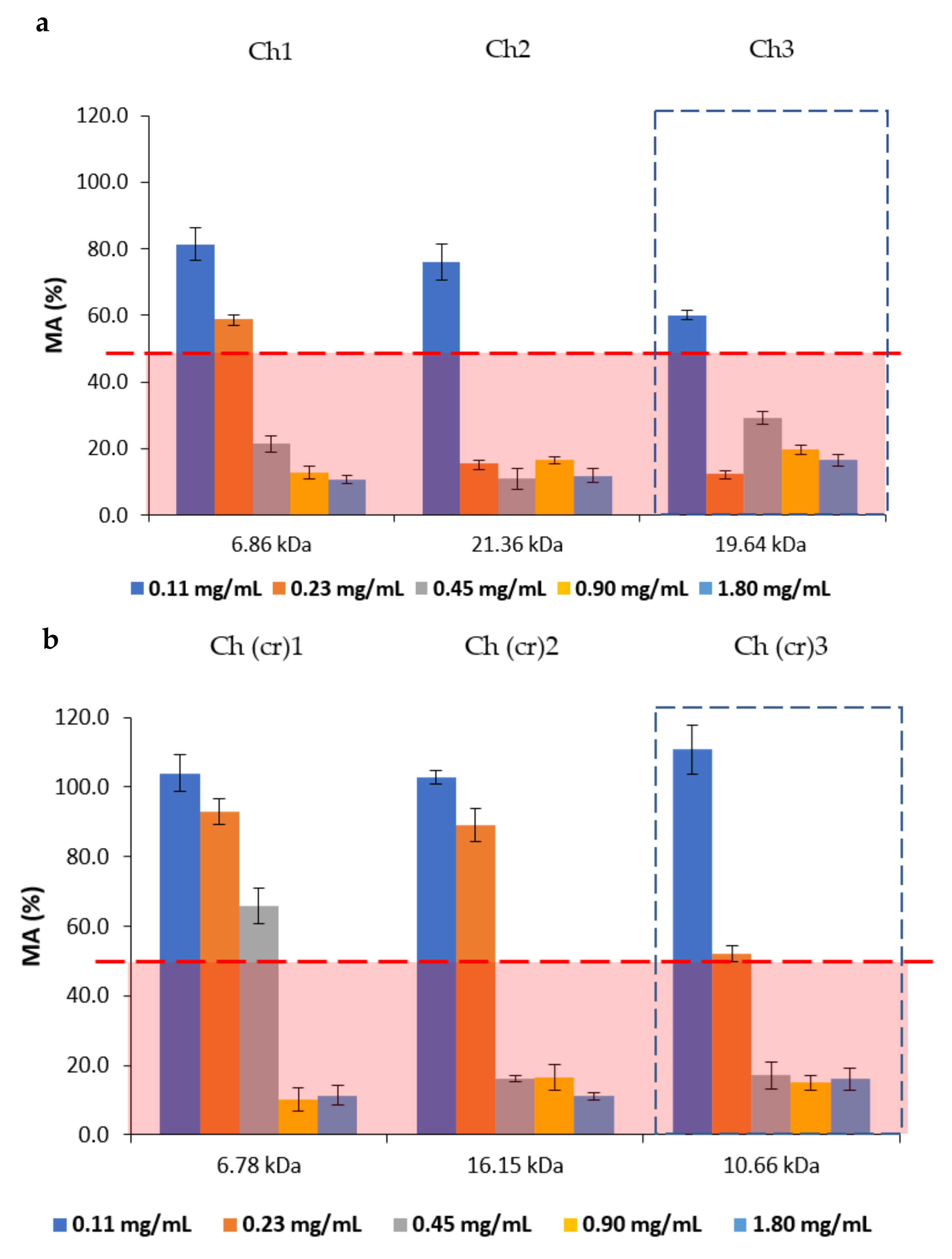
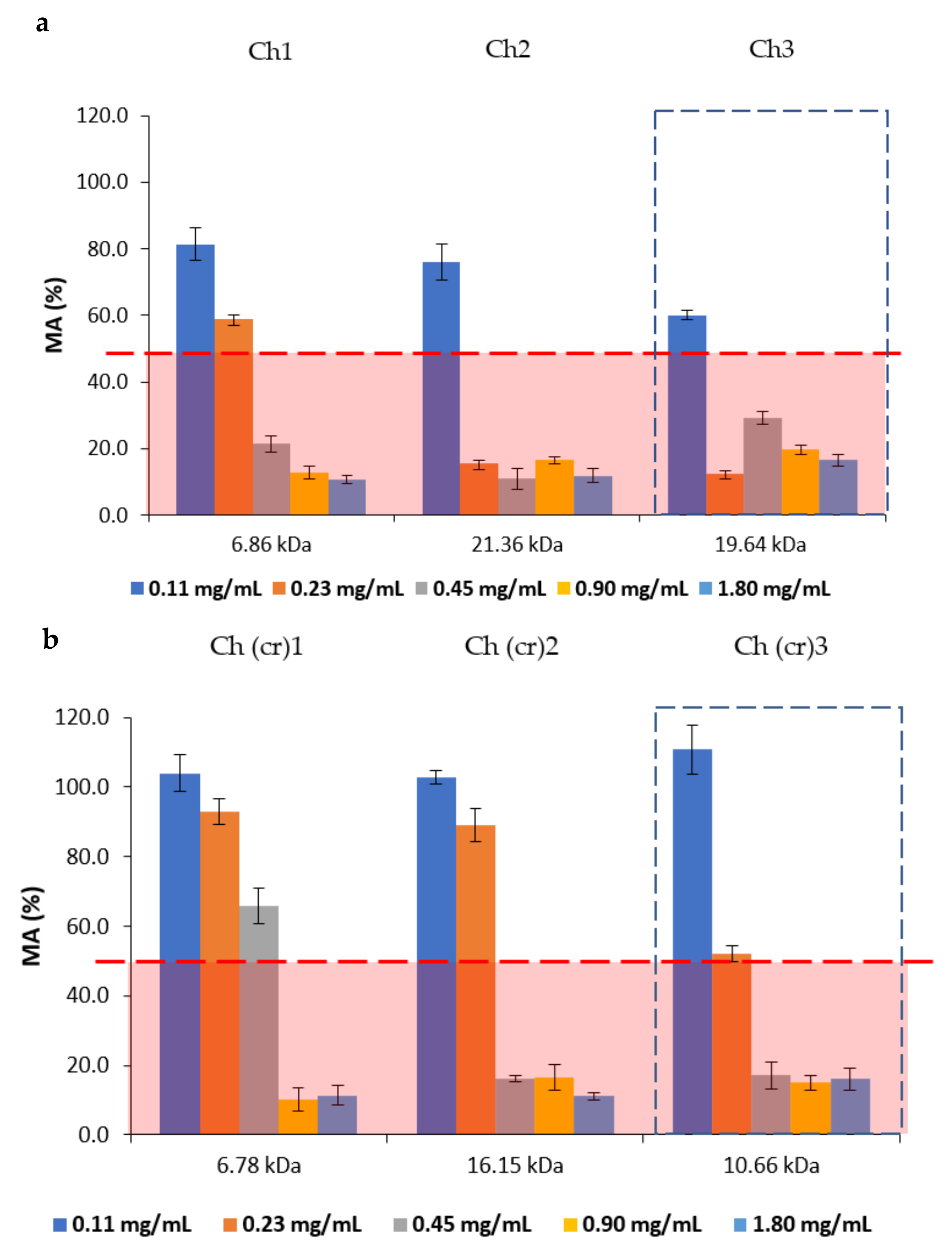
2.3. Antibacterial and Antifungal Activities
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Chitin Extraction
4.3. Chitosan Preparation
4.4. Enzyme Preparations for Chitosan Depolymerisation
4.5. Enzymatic Depolymerisation of Chitosans
4.6. Characterisation of Chitosan
4.6.1. Elemental Analysis
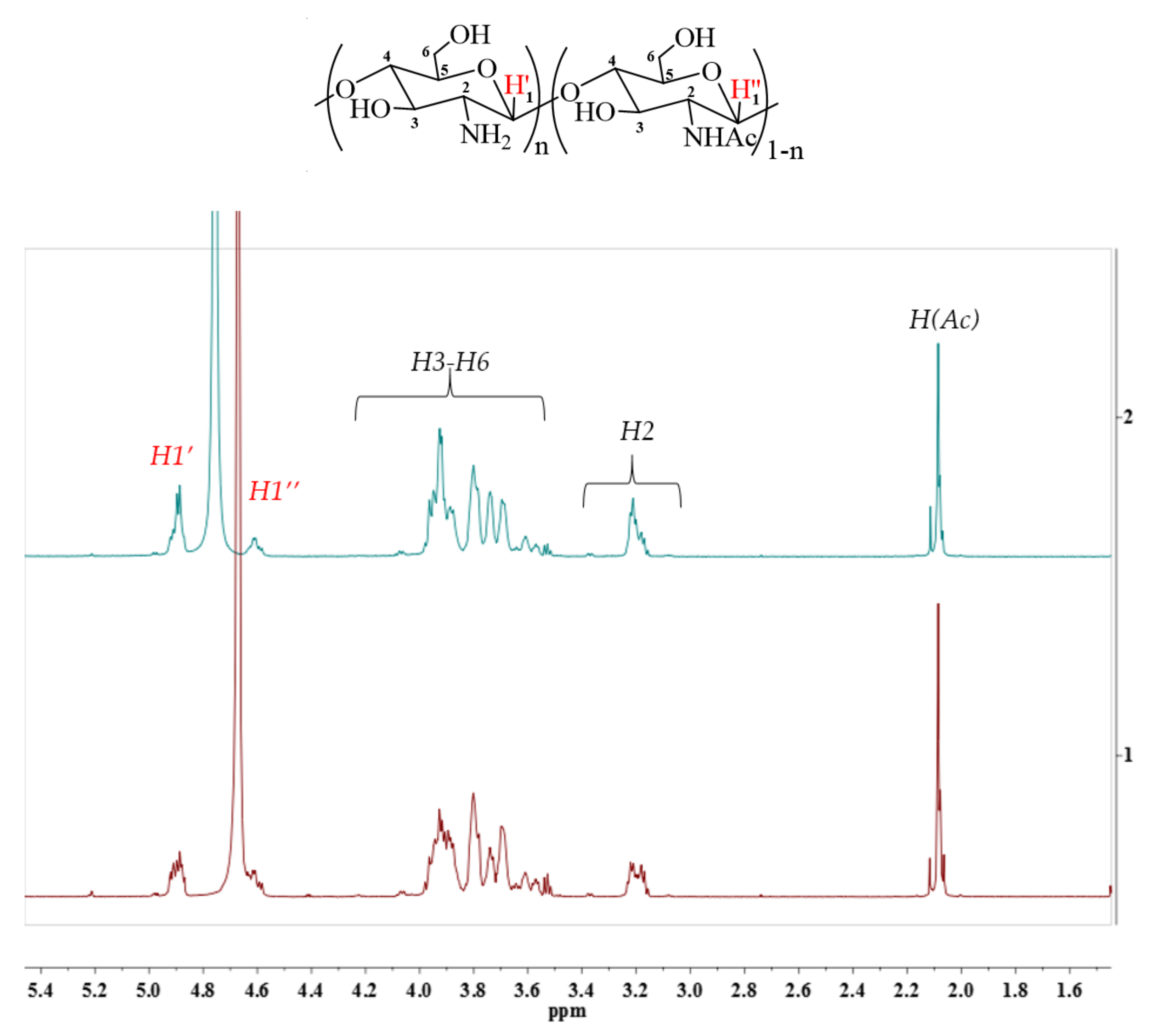
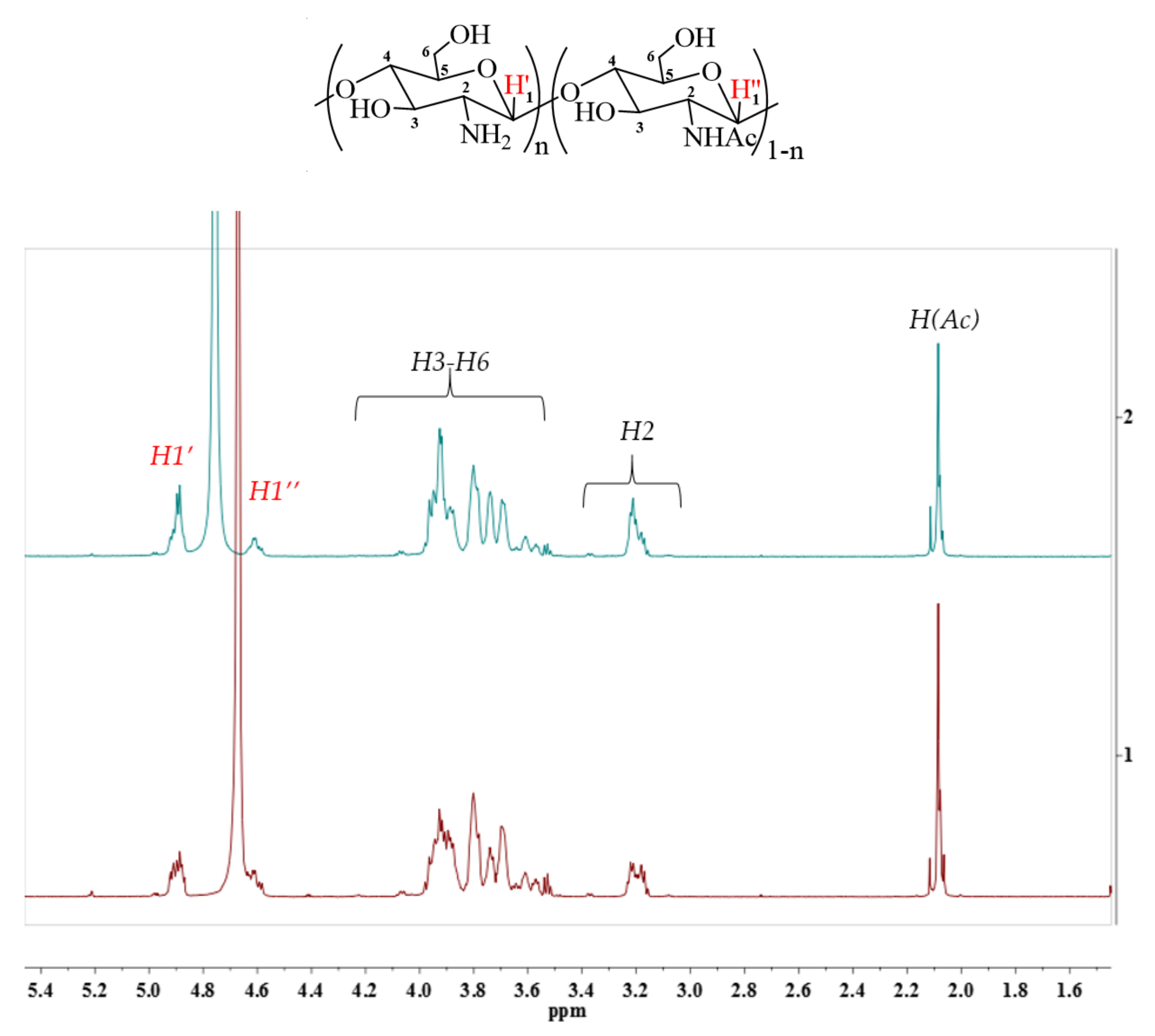
4.6.2. 1H NMR Analysis
4.6.3. Determination of the Degree of Deacetylation (DDA)
4.6.4. High Performance Liquid Chromatography (HPLC)
4.7. Microbiological Assays
4.7.1. Evaluation of the Antibacterial Activity
4.7.2. Evaluation of the Antifungal Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Azmana, M.; Mahmood, S.; Hilles, A.R.; Rahman, A.; Arifin, M.A.B.; Ahmed, S. A review on chitosan and chitosan-based bionanocomposites: Promising material for combatting global issues and its applications. Int. J. Biol. Macromol. 2021, 185, 832–848. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, Z.; Eini, M.; Rastegari, A.; Tehrani, M.R. Chitosan as a machine for biomolecule delivery: A review. Carbohydr. Polym. 2021, 256, 117414. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, R.; Parseh, B.; Du, G. Prospects and challenges of anticancer agents’ delivery via chitosan-based drug carriers to combat breast cancer: A review. Carbohydr. Polym. 2021, 268, 118192. [Google Scholar] [CrossRef] [PubMed]
- Pal, P.; Pal, A.; Nakashima, K.; Yadav, B.K. Applications of chitosan in environmental remediation: A review. Chemosphere 2021, 266, 128934. [Google Scholar] [CrossRef]
- Kostag, M.; El Seoud, O.A. Sustainable biomaterials based on cellulose, chitin and chitosan composites—A review. Carbohydr. Polym. Technol. Appl. 2021, 2, 100079. [Google Scholar] [CrossRef]
- Wong, C.Y.; Rosli, S.S.; Uemura, Y.; Ho, Y.C.; Leejeerajumnean, A.; Kiatkittipong, W.; Cheng, C.K.; Lam, M.K.; Lim, J.W. Potential protein and biodiesel sources from black soldier fly larvae: Insights of larval harvesting instar and fermented feeding medium. Energies 2019, 12, 1570. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.; Wolf, D.; Gutzeit, H.O. The black soldier fly, Hermetia illucens—A promising source for sustainable production of proteins, lipids and bioactive substances. Zeitschrift fur Naturforsch.—Sect. C J. Biosci. 2017, 72, 351–363. [Google Scholar] [CrossRef]
- Dumas, A.; Raggi, T.; Barkhouse, J.; Lewis, E.; Weltzien, E. The oil fraction and partially defatted meal of black soldier fly larvae (Hermetia illucens) affect differently growth performance, feed efficiency, nutrient deposition, blood glucose and lipid digestibility of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 492, 24–34. [Google Scholar] [CrossRef]
- Mertenat, A.; Diener, S.; Zurbrügg, C. Black Soldier Fly biowaste treatment – Assessment of global warming potential. Waste Manag. 2019, 84, 173–181. [Google Scholar] [CrossRef]
- Khayrova, A.; Lopatin, S.; Varlamov, V. Obtaining chitin, chitosan and their melanin complexes from insects. Int. J. Biol. Macromol. 2021, 167, 1319–1328. [Google Scholar] [CrossRef]
- Kaczmarek, M.B.; Struszczyk-Swita, K.; Li, X.; Szczęsna-Antczak, M.; Daroch, M. Enzymatic modifications of chitin, chitosan, and chitooligosaccharides. Front. Bioeng. Biotechnol. 2019, 7, 243. [Google Scholar] [CrossRef] [Green Version]
- Montilla, A.; Ruiz-Matute, A.I.; Corzo, N.; Giacomini, C.; Irazoqui, G. Enzymatic generation of chitooligosaccharides from chitosan using soluble and immobilized glycosyltransferase (Branchzyme). J. Agric. Food Chem. 2013, 61, 10360–10367. [Google Scholar] [CrossRef]
- Duan, B.; Huang, Y.; Lu, A.; Zhang, L. Recent advances in chitin based materials constructed via physical methods. Prog. Polym. Sci. 2018, 82, 1–33. [Google Scholar] [CrossRef]
- Liang, S.; Sun, Y.; Dai, X. A review of the preparation, analysis and biological functions of chitooligosaccharide. Int. J. Mol. Sci. 2018, 19, 2197. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Han, J.; Xue, Y.; Dai, Y.; Liu, J.; Gan, L.; Xie, R.; Long, M. Hydrogen peroxide pretreatment efficiently assisting enzymatic hydrolysis of chitosan at high concentration for chitooligosaccharides. Polym. Degrad. Stab. 2019, 164, 177–186. [Google Scholar] [CrossRef]
- Poshina, D.N.; Raik, S.V.; Poshin, A.N.; Skorik, Y.A. Accessibility of chitin and chitosan in enzymatic hydrolysis: A review. Polym. Degrad. Stab. 2018, 156, 269–278. [Google Scholar] [CrossRef]
- Hackman, R.H. Chemistry of insect cuticle. III. Hardening and darkening of the cuticle. Biochem. J. 1953, 54, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Beament, J.W.L. Wax Secretion in Insects. Nature 1951, 167, 652–653. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The epicuticle in an insect, Rhodnius prolixus (Hemiptera). Proc. R. Soc. Lond. Ser. B Biol. Sci. 1947, 134, 163–181. [Google Scholar]
- Park, Y.; Kim, M.H.; Park, S.C.; Cheong, H.; Jang, M.K.; Nah, J.W.; Hahm, K.S. Investigation of the antifungal activity and mechanism of action of LMWS-chitosan. J. Microbiol. Biotechnol. 2008, 18, 1729–1734. [Google Scholar]
- Roncal, T.; Oviedo, A.; López de Armentia, I.; Fernández, L.; Villarán, M.C. High yield production of monomer-free chitosan oligosaccharides by pepsin catalyzed hydrolysis of a high deacetylation degree chitosan. Carbohydr. Res. 2007, 342, 2750–2756. [Google Scholar] [CrossRef]
- Cabrera, J.; Cutsem, P.V. Preparation of chitooligosaccharides with degree of polymerization higher than 6 by acid or enzymatic degradation of chitosan. Biochem. Eng. J. 2005, 25, 165–172. [Google Scholar] [CrossRef]
- Ribas Fonseca, L.; Porto Santos, T.; Czaikoski, A.; Lopes Cunha, R. Modulating properties of polysaccharides nanocomplexes from enzymatic hydrolysis of chitosan. Food Res. Int. 2020, 137, 109642. [Google Scholar] [CrossRef]
- Miguez, N.; Kidibule, P.; Santos-Moriano, P.; Ballesteros, A.O.; Fernandez-Lobato, M.; Plou, F.J. Enzymatic synthesis and characterization of different families of chitooligosaccharides and their bioactive properties. Appl. Sci. 2021, 11, 3212. [Google Scholar] [CrossRef]
- Kim, S.-K.; Je, J.-Y. Continuous Production of Chitooligosaccharides by Enzymatic Hydrolysis. In Chitin, Chitosan, Oligosaccharides and Their Derivatives; CRC Press: Boca Raton, FL, USA, 2010; pp. 47–51. [Google Scholar]
- Abdel-Aziz, S.M.; Kahil, T.; Keera, A.A. Kinetic behavior of free and in situ immobilized chitosanases produced by the fungus Mucor rouxii. World Appl. Sci. J. 2014, 30, 1–9. [Google Scholar]
- Saharan, V.; Mehrotra, A.; Khatik, R.; Rawal, P.; Sharma, S.S.; Pal, A. Synthesis of chitosan based nanoparticles and their in vitro evaluation against phytopathogenic fungi. Int. J. Biol. Macromol. 2013, 62, 677–683. [Google Scholar] [CrossRef]
- Xing, K.; Shen, X.; Zhu, X.; Ju, X.; Miao, X.; Tian, J.; Feng, Z.; Peng, X.; Jiang, J.; Qin, S. Synthesis and in vitro antifungal efficacy of oleoyl-chitosan nanoparticles against plant pathogenic fungi. Int. J. Biol. Macromol. 2016, 82, 830–836. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; Rabea, E.I. Characterization and antimicrobial activity of water-soluble N-(4-carboxybutyroyl) chitosans against some plant pathogenic bacteria and fungi. Carbohydr. Polym. 2012, 87, 250–256. [Google Scholar] [CrossRef]
- Rabea, E.I.; Badawy, M.E.I.; Steurbaut, W.; Stevens, C.V. In vitro assessment of N-(benzyl)chitosan derivatives against some plant pathogenic bacteria and fungi. Eur. Polym. J. 2009, 45, 237–245. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; Rabea, E.I.; Eid, A.R.; Badr, M.M.; Marei, G.I.K. Structure and antimicrobial comparison between N-(benzyl) chitosan derivatives and N-(benzyl) chitosan tripolyphosphate nanoparticles against bacteria, fungi, and yeast. Int. J. Biol. Macromol. 2021, 186, 724–734. [Google Scholar] [CrossRef]
- Binesh, N.; Farhadian, N.; Mohammadzadeh, A. Enhanced stability of salt-assisted sodium ceftriaxone-loaded chitosan nanoparticles: Formulation and optimization by 32-full factorial design and antibacterial effect study against aerobic and anaerobic bacteria. Colloids Surfaces A Physicochem. Eng. Asp. 2021, 618, 126429. [Google Scholar] [CrossRef]
- Hussein, M.A.M.; Grinholc, M.; Dena, A.S.A.; El-Sherbiny, I.M.; Megahed, M. Boosting the antibacterial activity of chitosan–gold nanoparticles against antibiotic–resistant bacteria by Punicagranatum L. extract. Carbohydr. Polym. 2021, 256, 117498. [Google Scholar] [CrossRef] [PubMed]
- Etemadi, S.; Barhaghi, M.H.S.; Leylabadlo, H.E.; Memar, M.Y.; Mohammadi, A.B.; Ghotaslou, R. The synergistic effect of turmeric aqueous extract and chitosan against multidrug-resistant bacteria. New Microbes New Infect. 2021, 41, 100861. [Google Scholar] [CrossRef] [PubMed]
- Kole, S.; Qadiri, S.S.N.; Shin, S.-M.; Kim, W.-S.; Lee, J.; Jung, S.-J. Nanoencapsulation of inactivated-viral vaccine using chitosan nanoparticles: Evaluation of its protective efficacy and immune modulatory effects in olive flounder (Paralichthys olivaceus) against viral haemorrhagic septicaemia virus (VHSV) infection. Fish Shellfish Immunol. 2019, 91, 136–147. [Google Scholar] [CrossRef]
- Jia, S.; Zhou, K.; Pan, R.; Wei, J.; Liu, Z.; Xu, Y. Oral immunization of carps with chitosan–alginate microcapsule containing probiotic expressing spring viremia of carp virus (SVCV) G protein provides effective protection against SVCV infection. Fish Shellfish Immunol. 2020, 105, 327–329. [Google Scholar] [CrossRef]
- Modak, C.; Jha, A.; Sharma, N.; Kumar, A. Chitosan derivatives: A suggestive evaluation for novel inhibitor discovery against wild type and variants of SARS-CoV-2 virus. Int. J. Biol. Macromol. 2021, 187, 492–512. [Google Scholar] [CrossRef]
- Sharma, N.; Modak, C.; Singh, P.K.; Kumar, R.; Khatri, D.; Singh, S.B. Underscoring the immense potential of chitosan in fighting a wide spectrum of viruses: A plausible molecule against SARS-CoV-2? Int. J. Biol. Macromol. 2021, 179, 33–44. [Google Scholar] [CrossRef]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef] [Green Version]
- Marei, N.; Elwahy, A.H.M.; Salah, T.A.; El Sherif, Y.; El-Samie, E.A. Enhanced antibacterial activity of Egyptian local insects’ chitosan-based nanoparticles loaded with ciprofloxacin-HCl. Int. J. Biol. Macromol. 2019, 126, 262–272. [Google Scholar] [CrossRef]
- Matica, A.; Menghiu, G.; Ostafe, V. Antibacterial Properties of Chitin and Chitosans. New Front. Chem. 2017, 26, 39–54. [Google Scholar]
- Kim, J.Y.; Lee, J.K.; Lee, T.S.; Park, W.H. Synthesis of chitooligosaccharide derivative with quaternary ammonium group and its antimicrobial activity against Streptococcus mutans. Int. J. Biol. Macromol. 2003, 32, 23–27. [Google Scholar] [CrossRef]
- Mei, Y.X.; Dai, X.Y.; Yang, W.; Xu, X.W.; Liang, Y. xiang Antifungal activity of chitooligosaccharides against the dermatophyte Trichophyton rubrum. Int. J. Biol. Macromol. 2015, 77, 330–335. [Google Scholar] [CrossRef]
- Li, J.; Zhuang, S. Antibacterial activity of chitosan and its derivatives and their interaction mechanism with bacteria: Current state and perspectives. Eur. Polym. J. 2020, 138, 109984. [Google Scholar] [CrossRef]
- Vishu Kumar, A.B.; Varadaraj, M.C.; Gowda, L.R.; Tharanathan, R.N. Characterization of chito-oligosaccharides prepared by chitosanolysis with the aid of papain and Pronase, and their bactericidal action against Bacillus cereus and Escherichia coli. Biochem. J. 2005, 391, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Zhang, J.; Tan, W.; Wang, G.; Li, Q.; Dong, F.; Guo, Z. Antifungal activity of double Schiff bases of chitosan derivatives bearing active halogeno-benzenes. Int. J. Biol. Macromol. 2021, 179, 292–298. [Google Scholar] [CrossRef]
- Boddy, L. Chapter 8—Pathogens of Autotrophs. In The Fungi, 3rd ed.; Watkinson, S.C., Boddy, L., Money, N.P., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 245–292. [Google Scholar]
- Jacobs, K.; Wingfield, M.J.; Gibbs, J.N. PATHOLOGY|Diseases of Forest Trees. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Oxford, UK, 2004; pp. 766–772. [Google Scholar]
- Sinitsyna, O.A.; Rubtsova, E.A.; Sinelnikov, I.G.; Osipov, D.O.; Rozhkova, A.M.; Matys, V.Y.; Bubnova, T.V.; Nemashkalov, V.A.; Sereda, A.S.; Tcsherbakova, L.A.; et al. Creation of Chitinase Producer and Disruption of Micromycete Cell Wall with the Obtained Enzyme Preparation. Biochem. 2020, 85, 717–724. [Google Scholar] [CrossRef]
- Sinitsyn, A.P.; Okunev, O.N.; Chernoglazov, V.M.; Sinitsyna, O.A.; Popov, V.O. Strain of Mycelial Mushroom Penicillium Verruculosum—Producer of Complex of Cellulase, Xylanase and Xyloglucanase and Way of Reception of Fermental Preparation of Complex of Cellulase, Xylanase and Xyloglucanase for Hydrolysis of Cellulose and Hemicellulose. Patent RU 2361918, 20 July 2009. [Google Scholar]
- Sinitsyn, A.P.; Rozhkova, A.M.; Sinitsyna, O.A.; Fedorova, E.A.; Okunev, O.N.; Bekkarevich, A.O.; Sokolova, L.M.; Matys, V.J.; Koshelev, A.V.; Vinetskij, J.P.; et al. Genetic Maker to Ensure Expression of Target Homologous and Heterologous Genes in Cells of Filamentous Fungus Penicillium Verruculosum; Used as Host, Method for Making Strain of Fungus Penicillium Verruculosum and Method for Making Enzymatic Agent. Patent RU 2378372, 20 January 2010. [Google Scholar]
- Sinitsyn, A.P.; Korotkova, O.G.; Rubtsova, E.A.; Sinitsyna, O.A.; Kondrat’eva, E.G.; Sereda, A.S.; Zorov, I.N.; Rojkova, A.M. Construction of Recombinant Producers of Enzyme Preparations for Fodder Production by Means of Expression System Based on Penicillium verruculosum Fungus. Biotechnology 2019, 35, 6–14. [Google Scholar] [CrossRef]
- Friedemann, T.E.; Weber, C.W.; Witt, N.F. Determination of reducing sugars by oxidation in alkaline ferricyanide solution. Anal. Biochem. 1962, 4, 358–377. [Google Scholar] [CrossRef]
- Lyalina, T.; Zubareva, A.; Lopatin, S.; Zubov, V.; Sizova, S.; Svirshchevskaya, E. Correlation Analysis of Chitosan Physicochemical Parameters Determined by Different Methods. Org. Med. Chem. Int. J. 2017, 1, 555562. [Google Scholar]
- Lopatin, S.A.; Derbeneva, M.S.; Kulikov, S.N.; Varlamov, V.P.; Shpigun, O.A. Fractionation of chitosan by ultrafiltration. J. Anal. Chem. 2009, 64, 648–651. [Google Scholar] [CrossRef]
- Shagdarova, B.; Lunkov, A.; Il’ina, A.; Varlamov, V. Investigation of the properties of N-[(2-hydroxy-3-trimethylammonium) propyl] chloride chitosan derivatives. Int. J. Biol. Macromol. 2019, 124, 994–1001. [Google Scholar] [CrossRef]
- Karpova, N.V.; Shagdarova, B.T.; Lyalina, T.S.; Il’ina, A.V.; Tereshina, V.M.; Varlamov, V.P. Influence of the Main Characteristics of Low Weight Chitosan on the Growth of the Plant Pathogenic Fungus Botrytis cinerea. Appl. Biochem. Microbiol. 2019, 55, 405–413. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | %C | %H | %N |
|---|---|---|---|
| Crab | 39.48 | 6.91 | 6.67 |
| H. illucens | 41.28 | 6.87 | 7.09 |
| Sample | Ch1 | Ch (cr)1 | Ch2 | Ch (cr)2 | Ch3 | Ch (cr)3 |
|---|---|---|---|---|---|---|
| Mw, kDa | 6.86 | 6.78 | 21.36 | 16.15 | 19.64 | 10.66 |
| PDI | 1.67 | 1.54 | 2.14 | 1.95 | 1.96 | 1.71 |
| MIC, µg/mL | 62.5 | 125 | 62.5 | 62.5 | 62.5 | 62.5 |
| Enzyme Preparation Number | Recombinant Enzyme | Activity by Chitosan (1000 kDa), U/g * |
|---|---|---|
| # 3-432.5 | chitosanase Chi 402 from M. thermophila (Mw 44 kDa) | 600 |
| # 3-458.1 | chitinase Chi 418 from T. harzianum (Mw 42 kDa) | 2000 |
| # 3-544.H | chitinase Chi 403 from M. thermophila (Mw 43 kDa) | 1500 |
| Sample | MW, kDa | PDI | Enzyme |
|---|---|---|---|
| Ch1 | 6.86 | 1.67 | # 3-432.5 Chi 402—chitosanase M. thermophila |
| Ch (cr)1 | 6.78 | 1.54 | |
| Ch2 | 21.36 | 2.14 | |
| Ch (cr)2 | 16.15 | 1.95 | |
| Ch3 | 19.64 | 1.96 | # 3-544.H Chi 403—chitinase M. thermophila |
| Ch (cr)3 | 10.66 | 1.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khayrova, A.; Lopatin, S.; Shagdarova, B.; Sinitsyna, O.; Sinitsyn, A.; Varlamov, V. Evaluation of Antibacterial and Antifungal Properties of Low Molecular Weight Chitosan Extracted from Hermetia illucens Relative to Crab Chitosan. Molecules 2022, 27, 577. https://doi.org/10.3390/molecules27020577
Khayrova A, Lopatin S, Shagdarova B, Sinitsyna O, Sinitsyn A, Varlamov V. Evaluation of Antibacterial and Antifungal Properties of Low Molecular Weight Chitosan Extracted from Hermetia illucens Relative to Crab Chitosan. Molecules. 2022; 27(2):577. https://doi.org/10.3390/molecules27020577
Chicago/Turabian StyleKhayrova, Adelya, Sergey Lopatin, Balzhima Shagdarova, Olga Sinitsyna, Arkady Sinitsyn, and Valery Varlamov. 2022. "Evaluation of Antibacterial and Antifungal Properties of Low Molecular Weight Chitosan Extracted from Hermetia illucens Relative to Crab Chitosan" Molecules 27, no. 2: 577. https://doi.org/10.3390/molecules27020577
APA StyleKhayrova, A., Lopatin, S., Shagdarova, B., Sinitsyna, O., Sinitsyn, A., & Varlamov, V. (2022). Evaluation of Antibacterial and Antifungal Properties of Low Molecular Weight Chitosan Extracted from Hermetia illucens Relative to Crab Chitosan. Molecules, 27(2), 577. https://doi.org/10.3390/molecules27020577