
Moco Carrier and Binding Proteins

Institute of Plant Biology, Technische Universität Braunschweig, 38106 Braunschweig, Germany
Molecules 2022, 27(19), 6571; https://doi.org/10.3390/molecules27196571
Submission received: 31 August 2022
/
Revised: 25 September 2022
/
Accepted: 26 September 2022
/
Published: 4 October 2022
(This article belongs to the Special Issue State-of-the-Art in Molybdenum Cofactor Research)
Abstract
The molybdenum cofactor (Moco) is the active site prosthetic group found in numerous vitally important enzymes (Mo-enzymes), which predominantly catalyze 2 electron transfer reactions. Moco is synthesized by an evolutionary old and highly conserved multi-step pathway, whereby the metal insertion reaction is the ultimate reaction step here. Moco and its intermediates are highly sensitive towards oxidative damage and considering this, they are believed to be permanently protein bound during synthesis and also after Moco maturation. In plants, a cellular Moco transfer and storage system was identified, which comprises proteins that are capable of Moco binding and release but do not possess a Moco-dependent enzymatic activity. The first protein described that exhibited these properties was the Moco carrier protein (MCP) from the green alga Chlamydomonas reinhardtii. However, MCPs and similar proteins have meanwhile been described in various plant species. This review will summarize the current knowledge of the cellular Moco distribution system.
1. Introduction
The molybdenum cofactor (Moco) biosynthesis pathway comprises several strictly conserved reaction steps, ultimately yielding Moco. Within the first step of the pathway, GTP is converted to cyclic pyranopterin monophosphate (cPMP, [1,2]) a reaction thatinvolves the formation of the instable intermediate 3′,8-cH2GTP [3,4]. Recent work suggests that it is the first step of eukaryotic Moco biosynthesis which is under regulatory control as deduced from the finding that in the filamentous fungus Neurospora crassa (N. crassa, [5]) the 3′,8-cH2GTP synthesizing enzyme is co-regulated (precisely the formation of one particular splice variant) with the major N. crassa Moco user enzyme (i.e. nitrate reductase). In the second step of the pathway, cPMP is converted to molybdopterin (MPT) in a stoichiometric reaction which is catalyzed by the heterotetrameric MPT synthase complex. This reaction is best characterized by the fact that the MPT dithiolene sulfurs are incorporated here to assemble the metal-free Moco scaffold [6,7,8]. In the subsequent third and fourth reaction steps, molybdate is incorporated into the MPT dithiolene moiety (for a recent review, see [9] in this Special Issue). Initially, MPT binds to the molybdenum insertase (Mo-insertase) G-domain where it becomes adenylylated, yielding MPT-AMP (termed adenylated MPT, [10]). In the following, MPT-AMP is used as the substrate from the Mo-insertase E-domain, which catalyzes the insertion of molybdate, yielding Moco-AMP [11]. The subsequent hydrolysis of the Moco-AMP phophoanhydride-bond then releases Moco (Figure 1, [11]), which can now be transferred either to the Moco-dependent enzymes or to the cell’s Moco transfer and storage system [12].
For a recent review of Moco biosynthesis, see Ralf Mendel’s review in this Special Issue [13]. The most likely candidate proteins for the cellular transfer and storage system have been identified and characterized in plants (i.e., Moco carrier protein (MCP, [14,15,16,17]) and Moco binding proteins (MoBP, [18,19]). However, these proteins possess considerably different Moco binding and stabilizing properties [12,18,19], as documented by the finding that plant MCP proteins can be co-purified with significant amounts of Moco [16,17], while MoBP proteins can’t [18,19]. Notably, the hitherto sole known prokaryotic MCP was found to be co-purified with comparably low amounts of Moco [20], thus classifying the Moco binding properties of this protein to be similar to the A. thaliana MoBP proteins.
Next to the biochemical characterization, the results coming from in vivo approaches also classify the MoBP proteins to function within the cellular Moco transfer and storage system. Members of the MoBP protein family were shown to interact with both the molybdenum insertase [21], the Mo-enzyme nitrate reductase [18,21], and the Moco sulfurase ABA3 in plants [21]. However, for MCP proteins, a similar detailed in vivo characterization is missing, but these were shown to reconstitute nitrate reductase activity both in fully defined in vitro systems and in the nit-1-based reconstitution assay [14,16,22]. Within this review, the current view on the function of MCP and MoBP proteins for the cellular Moco transfer will be summarized (see Figure 2 for an overview).
2. Identification and Occurance of Moco Carrier and Binding Proteins
The first incidence ofthe existence of a Moco carrier protein (MCP) was reported for the prokaryote E. coli in 1979 [24]. However, indications for the existence of an MCP protein were also later on reported for eukaryotes [14,25], while—similar to E. coli—he identity of the protein was not elucidated in the initial study. However, for C. reinhardtii [14] following work revealed the identity of the MCP amino acid sequence and the encoding gene [15]. Work with the recombinant C. reinhardtii MCP protein then characterized its Moco-binding properties [15,16] and revealed its structure [16]. Having hands on the structure was of great importance since, at that time, the identification of C. reinhardtii MCP homologs was restrained to the use of structure-based homology searches, as no meaningful results were obtained by using the C. reinhardtii MCP amino acid sequence as a query in pBLAST searches [18]. Structure-based homology searches then revealed two homologous eukaryotic (plant, Arabidopsis thaliana) structures [18] and three of bacterial origin [16]. However, the identified A. thaliana proteins were found to be part of a protein family [26] and were termed Moco binding proteins (MoBP) to indicate their — as compared to C. reinhardtii MCP—different Moco-binding properties. Notably, next to the existence of an MCP in C. reinhardtii also a putative MoBP was identified here, which is not member of a protein family [19]. The following work took advantage ofthe ongoing progress in the field of genome sequencing and annotation (Table 1, [17,20]), which allowed it to identify potential C. reinhardtii MCP homologs amino acid sequence-based in the algae Chlamydomonas eustigma, Haematococcus lacustris, Gonium pectoral and Volvox carteri [17]. Likewise, C. reinhardtii MCP homologs were identified in various bacteria [20], with candidate proteins from the phylum Cyanobacteria and α-Proteobacteria showing the highest degree of sequence identity to CrMCP (ranging from 50–60%, [20]).
3. Biochemical and Structural Characterization of Moco Carrier and Binding Proteins
Upon identification of the C. reinhardtii MCP encoding gene (mcp1) it became possible to express and purify the recombinant protein, facilitating the biochemical characterization of MCP [15,16]. Doing so revealed the recombinant protein to be a tetramer in solution, and further, C. reinhardtii MCP was found to be co-purified with Moco upon expression in the E. coli TP1000 strain [27], while no MPT was co-purified with recombinant C. reinhardtii MCP upon expression in the MPT accumulating E. coli RK5206 strain [28]. Most importantly, next to the capability to bind Moco, the recombinant C. reinhardtii MCP was found to serve as a Moco donor for Moco-free nitrate reductase in nit-1 cell extracts without preceding heat or acidic treatment [16]. As this renders C. reinhardtii MCP to bind Moco reversible, it was concluded that within the cell the carrier protein could serve as a Moco storage protein to buffer a varying Moco demand [15,16]. To give a first insight into the molecular detail behind C. reinhardtii MCP Moco-binding, a putative Moco binding site was identified and a docking approach suggested the positioning of Moco within here. Adjacent to the putative binding site, a structural disorder loop is located. Upon the sequence-based identification of various other putative eukaryotic MCPs, within the N-terminal part of the MCP amino acid sequence, a sequence stretch of low homology (lhr) was identified [17]. For C. reinhardii MCP, this sequence was found to form the aforementioned structurally disordered loop and it was suggested that the observed differences in sequence result in varying properties of the binding site, which were suggested to be required for the adaption of each particular MCP to the varying cellular properties of the respective host [17]. Consistently, the (eukaryotic) MCP from Volvox carteri (V. carteri) was found to adapt the Moco binding properties from the prokaryotic MCP from Rippkaea orientalis, when the V. carteri lhr sequence is replaced with the R. orientalis lhr sequence [17]. Next to recombinant MCPs, also the MoBP proteins from A. thaliana were characterized biochemically, and contrary to the recombinant C. reinhardtii and V. carteri MCPs [16,17], their characterization revealed none of them to be co-purified with significant amounts of Moco, albeit in vitro loading of Moco to MoBP proteins was possible [18]. The Moco KD values determined for the recombinant MoBP proteins were found to lay in the low µM range thus being in the range of the Moco KD value determined for recombinantCnx1E [29]. As both, recombinant Cnx1E and recombinant MoBP proteins cannot be co-purified with significant amounts of Moco, it can be concluded that the Moco KD values determined do not allow the co-purification of Moco with any of these proteins. Next to the rather low Moco KD values recorded [18], the fact that member(s) of the MoBP protein family were found to interact with the Mo-insertase Cnx1 (catalyzing the final step of Moco biosynthesis [18,30] and downstream Moco user enzymes (i.e., nitrate reductase [18] and Moco sulfurase [30]) led to the conclusion that these proteins have a function for Moco transfer in the cell but not for Moco storage (see also [19]). Likewise, for C. reinhardtii MCP data were reported that also suggests a protein–protein interaction here with a Moco user enzyme (i.e., nitrate reductase, [14]) which was shown to be of functional importance. Consequently, MCP proteins were suggested to be involved in both, Moco transfer and storage [19]. Clearly, one of the major differences between MoBP and MCP proteins is the varying affinity for Moco binding, which could be seen as an adaption to fulfill the respective cellular function(s) these proteins are assumed to have.
The first structurally characterized Moco carrier protein was the MCP from C. reinhardtii [16]. The C. reinhardtii MCP monomers are arranged in a Rossmann-like fold and the symmetric homo-tetramer assembled from these was described as dimers of dimers, which was traced back to the fact that each monomer contributes to two distinct intermolecular interactions here [16]. Each monomer houses one putative Moco binding site. Recently the structure of the first prokaryotic (Rippkaea orientalis, R. orientalis) MCP has been reported [20], which was found to be highly similar to that of C. reinhardtii MCP. Similar to C. reinhardtii MCP and also for R. orientalis MCP, a positively charged surface depression was identified as a putative Moco binding site and consistent with the docking results reported for C. reinhardtii MCP [16], likewise for R. orientalis MCP, the phosphate moiety of Moco was suggested to reside deeply buried within the binding pocket while the molybdenum center was suggested to be rather surface exposed. The C. reinhardtii monomer was found to be structurally highly similar to that of the MoBP monomer, while other than MCP proteins, MoBP proteins form dimers both in the structure and in solution [18] and contrary to MCP, Moco binding to Arabidopsis MoBP proteins was traced back to the C-terminus of the protein [18], which potentially could explain the differences in Moco binding displayed by MoBP and plant MCP proteins.
4. Conclusions
The fate of Moco upon synthesis can only be traced back to limited detail. This is due to the finding that—contrary to Moco biosynthesis—no mutants of the cellular Moco distribution and storage system are known. However, candidate proteins were identified in plants and bacteria, while hitherto, in the animal and fungal kingdoms homologs to the plant proteins have not been revealed and characterized so far. Therefore it can be concluded that though Moco biosynthesis is strictly conserved amongst the kingdoms of life, it´s trafficking and most likely also the processes underlying the insertion of Moco into the cognate molybdenum enzymes is not. It is tempting to assume that the presence of MCP/MoBP proteins in plants is linked to the outstanding importance of nitrate reductase here. However, fungi also possess the nitrate assimilation pathway but lack functional homologs to the known Moco carrier and binding proteins, suggesting that here the Moco carrier/binding proteins were lost during evolution. This could be explained by the fact that Moco biosynthesis is tightly regulated here, thus rendering a Moco storage system needless.
Funding
T.K. acknowledges the support of this research by the Deutsche Forschungsgemeinschaft (GRK 2223/1). Open access publication was funded by Publikations fonds of TU Braunschweig.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I would like to extend my sincere gratitude to Ralf Mendel for his experience, support and advice.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wuebbens, M.M.; Rajagopalan, K.V. Structural characterization of a molybdopterin precursor. J. Biol. Chem. 1993, 268, 13493–13498. [Google Scholar] [CrossRef]
- Santamaria-Araujo, J.A.; Fischer, B.; Otte, T.; Nimtz, M.; Mendel, R.R.; Wray, V.; Schwarz, G. The tetrahydropyranopterin structure of the sulfur-free and metal-free molybdenum cofactor precursor. J. Biol. Chem. 2004, 279, 15994–15999. [Google Scholar] [CrossRef] [PubMed]
- Hover, B.M.; Loksztejn, A.; Ribeiro, A.A.; Yokoyama, K. Identification of a cyclic nucleotide as a cryptic intermediate in molybdenum cofactor biosynthesis. J. Am. Chem. Soc. 2013, 135, 7019–7032. [Google Scholar] [CrossRef] [PubMed]
- Hover, B.M.; Tonthat, N.K.; Schumacher, M.A.; Yokoyama, K. Mechanism of pyranopterin ring formation in molybdenum cofactor biosynthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 6347–6352. [Google Scholar] [CrossRef]
- Wajmann, S.; Hercher, T.W.; Buchmeier, S.; Hansch, R.; Mendel, R.R.; Kruse, T. The First Step of Neurospora crassa Molybdenum Cofactor Biosynthesis: Regulatory Aspects under N-Derepressing and Nitrate-Inducing Conditions. Microorganisms 2020, 8, 534. [Google Scholar] [CrossRef]
- Pitterle, D.M.; Johnson, J.L.; Rajagopalan, K.V. In vitro synthesis of molybdopterin from precursor Z using purified converting factor. Role of protein-bound sulfur in formation of the dithiolene. J. Biol. Chem. 1993, 268, 13506–13509. [Google Scholar] [CrossRef]
- Wuebbens, M.M.; Rajagopalan, K.V. Mechanistic and Mutational Studies of Escherichia coli Molybdopterin Synthase Clarify the Final Step of Molybdopterin Biosynthesis. J. Biol. Chem. 2003, 278, 14523–14532. [Google Scholar] [CrossRef]
- Rudolph, M.J.; Wuebbens, M.M.; Turque, O.; Rajagopalan, K.V.; Schindelin, H. Structural studies of molybdopterin synthase provide insights into its catalytic mechanism. J. Biol. Chem. 2003, 278, 14514–14522. [Google Scholar] [CrossRef]
- Kruse, T. Function of Molybdenum Insertases. Molecules 2022, 27, 5372. [Google Scholar] [CrossRef]
- Kuper, J.; Llamas, A.; Hecht, H.J.; Mendel, R.R.; Schwarz, G. Structure of the molybdopterin-bound Cnx1G domain links molybdenum and copper metabolism. Nature 2004, 430, 803–806. [Google Scholar] [CrossRef]
- Probst, C.; Yang, J.; Krausze, J.; Hercher, T.W.; Richers, C.P.; Spatzal, T.; Kc, K.; Giles, L.J.; Rees, D.C.; Mendel, R.R.; et al. Mechanism of molybdate insertion into pterin-based molybdenum cofactors. Nat. Chem. 2021, 13, 758–765. [Google Scholar] [CrossRef]
- Mendel, R.R. The molybdenum cofactor. J. Biol. Chem. 2013, 288, 13165–13172. [Google Scholar] [CrossRef]
- Mendel, R.R. The History of the Molybdenum Cofactor—A Personal View. Molecules 2022, 27, 4934. [Google Scholar] [CrossRef]
- Aguilar, M.; Kalakoutskii, K.; Cardenas, J.; Fernandez, E. Direct transfer of molybdopterin cofactor to apo nitrate reductase from a carrier protein in Chlamydomonas reinhardtii. FEBS Lett. 1992, 307, 162–163. [Google Scholar] [CrossRef]
- Ataya, F.S.; Witte, C.P.; Galvan, A.; Igeno, M.I.; Fernandez, E. Mcp1 Encodes the Molybdenum Cofactor Carrier Protein in Chlamydomonas reinhardtii and Participates in Protection, Binding, and Storage Functions of the Cofactor. J. Biol. Chem. 2003, 278, 10885–10890. [Google Scholar] [CrossRef]
- Fischer, K.; Llamas, A.; Tejada-Jimenez, M.; Schrader, N.; Kuper, J.; Ataya, F.S.; Galvan, A.; Mendel, R.R.; Fernandez, E.; Schwarz, G. Function and structure of the molybdenum cofactor carrier protein from Chlamydomonas reinhardtii. J. Biol. Chem. 2006, 281, 30186–30194. [Google Scholar] [CrossRef]
- Hercher, T.W.; Krausze, J.; Yang, J.; Kirk, M.L.; Kruse, T. Identification and characterisation of the Volvox carteri Moco carrier protein. Biosci. Rep. 2020, 40, BSR20202351. [Google Scholar] [CrossRef]
- Kruse, T.; Gehl, C.; Geisler, M.; Lehrke, M.; Ringel, P.; Hallier, S.; Hansch, R.; Mendel, R.R. Identification and biochemical characterization of molybdenum cofactor-binding proteins from Arabidopsis thaliana. J. Biol. Chem. 2010, 285, 6623–6635. [Google Scholar] [CrossRef]
- Llamas, A.; Tejada-Jimenez, M.; Fernandez, E.; Galvan, A. Molybdenum metabolism in the alga Chlamydomonas stands at the crossroad of those in Arabidopsis and humans. Metallomics 2011, 3, 578–590. [Google Scholar] [CrossRef]
- Krausze, J.; Hercher, T.W.; Archna, A.; Kruse, T. The structure of the Moco carrier protein from Rippkaea orientalis. Acta Crystallogr. F Struct. Biol.Commun. 2020, 76, 453–463. [Google Scholar] [CrossRef]
- Kaufholdt, D.; Baillie, C.K.; Meyer, M.H.; Schwich, O.D.; Timmerer, U.L.; Tobias, L.; van Thiel, D.; Hansch, R.; Mendel, R.R. Identification of a protein-protein interaction network downstream of molybdenum cofactor biosynthesis in Arabidopsis thaliana. J. Plant Physiol. 2016, 207, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Ringel, P.; Krausze, J.; van den Heuvel, J.; Curth, U.; Pierik, A.J.; Herzog, S.; Mendel, R.R.; Kruse, T. Biochemical characterization of molybdenum cofactor-free nitrate reductase from Neurospora crassa. J. Biol. Chem. 2013, 288, 14657–14671. [Google Scholar] [CrossRef] [PubMed]
- Llamas, A.; Otte, T.; Multhaup, G.; Mendel, R.R.; Schwarz, G. The Mechanism of nucleotide-assisted molybdenum insertion into molybdopterin. A novel route toward metal cofactor assembly. J. Biol. Chem. 2006, 281, 18343–18350. [Google Scholar] [CrossRef] [PubMed]
- Amy, N.K.; Rajagopalan, K.V. Characterization of molybdenum cofactor from Escherichia coli. J. Bacteriol. 1979, 140, 114–124. [Google Scholar] [CrossRef]
- Kalakoutskii, K.; Fernandez, E. Different forms of molybdenum cofactor in Vicia faba seeds: The presence of molybdenum cofactor carrier protein and its purification. Planta 1996, 201, 64–70. [Google Scholar] [CrossRef]
- Palmer, T.; Santini, C.-L.; Iobbi-Nivol, C.; Eaves, D.J.; Boxer, D.H.; Giordano, G. Involvement of the narJ and mob gene products in distinct steps in the biosynthesis of the molybdoenzyme nitrate reductase in Escherichia coli. Mol. Microbiol. 1996, 20, 875–884. [Google Scholar] [CrossRef]
- Stewart, V.; MacGregor, C.H. Nitrate reductase in Escherichia coli K-12: Involvement of chlC, chlE, and chlG loci. J. Bacteriol. 1982, 151, 788–799. [Google Scholar] [CrossRef]
- Schwarz, G.; Boxer, D.H.; Mendel, R.R. Molybdenum cofactor biosynthesis. The plant protein Cnx1 binds molybdopterin with high affinity. J. Biol. Chem. 1997, 272, 26811–26814. [Google Scholar] [CrossRef]
- Kaufholdt, D.; Baillie, C.K.; Bikker, R.; Burkart, V.; Dudek, C.A.; von Pein, L.; Rothkegel, M.; Mendel, R.R.; Hansch, R. The molybdenum cofactor biosynthesis complex interacts with actin filaments via molybdenum insertase Cnx1 as anchor protein in Arabidopsis thaliana. Plant Sci. 2016, 244, 8–18. [Google Scholar] [CrossRef]
- Jeon, W.B.; Allard, S.T.; Bingman, C.A.; Bitto, E.; Han, B.W.; Wesenberg, G.E.; Phillips, G.N., Jr. X-ray crystal structures of the conserved hypothetical proteins from Arabidopsis thaliana gene loci At5g11950 and AT2g37210. Proteins 2006, 65, 1051–1054. [Google Scholar] [CrossRef]
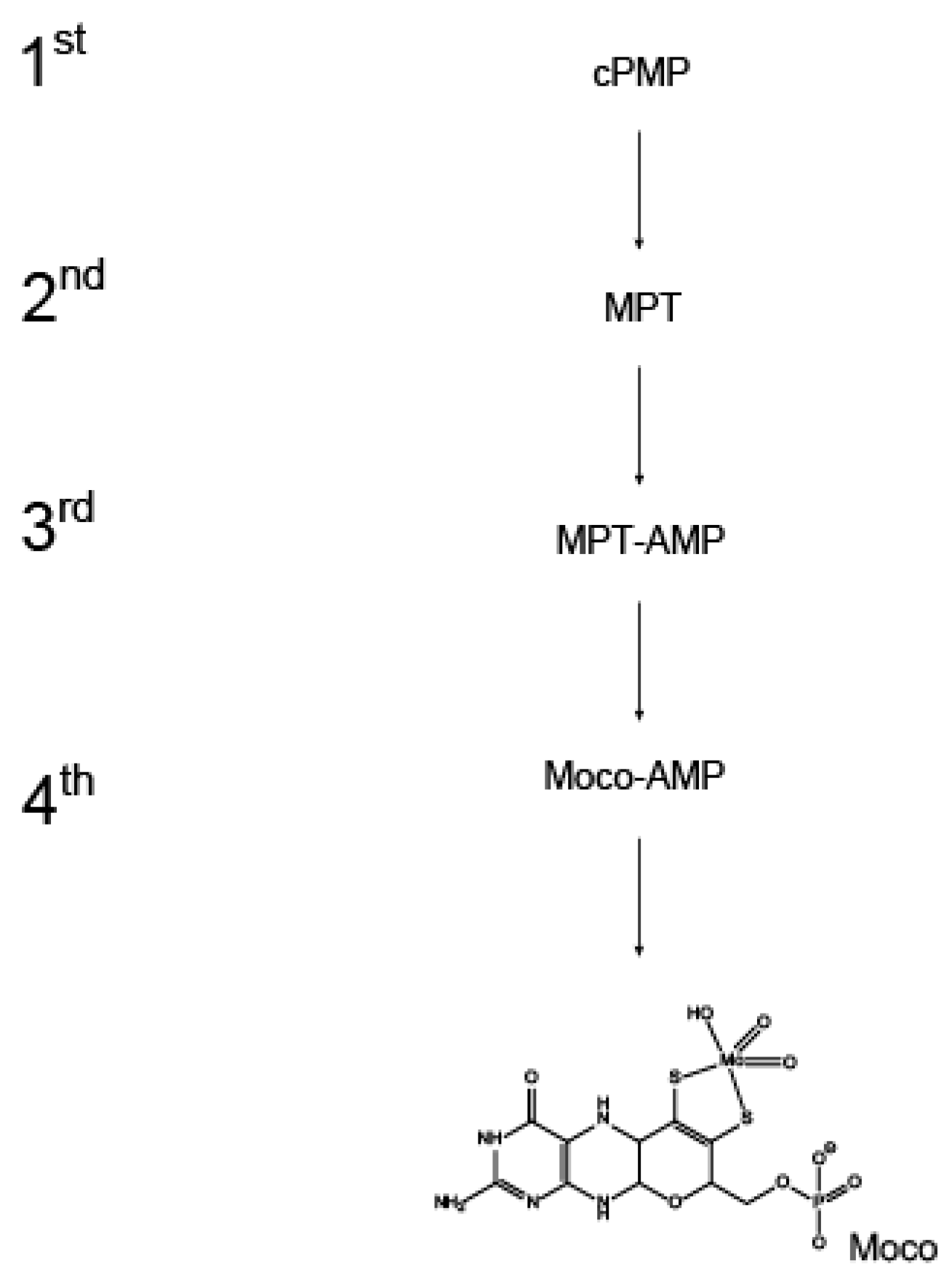
Figure 1.
The eukaryotic molybdenum cofactor biosynthesis. Molybdenum cofactor (Moco) biosynthesis comprises four subsequent reaction steps, each giving yield to the intermediates shown: The reaction product of the first reaction step is cyclic pyranopterin monophosphate (cPMP) which is subsequently converted to molybdopterin (MPT) in the second reaction step. MPT is adenylyated, yielding MPT-AMP that is then converted to Moco-AMP. In the ultimate, fourth reaction step, the phosphoanhydride-bond within Moco-AMP is hydrolyzed and Moco is released.
Figure 1.
The eukaryotic molybdenum cofactor biosynthesis. Molybdenum cofactor (Moco) biosynthesis comprises four subsequent reaction steps, each giving yield to the intermediates shown: The reaction product of the first reaction step is cyclic pyranopterin monophosphate (cPMP) which is subsequently converted to molybdopterin (MPT) in the second reaction step. MPT is adenylyated, yielding MPT-AMP that is then converted to Moco-AMP. In the ultimate, fourth reaction step, the phosphoanhydride-bond within Moco-AMP is hydrolyzed and Moco is released.

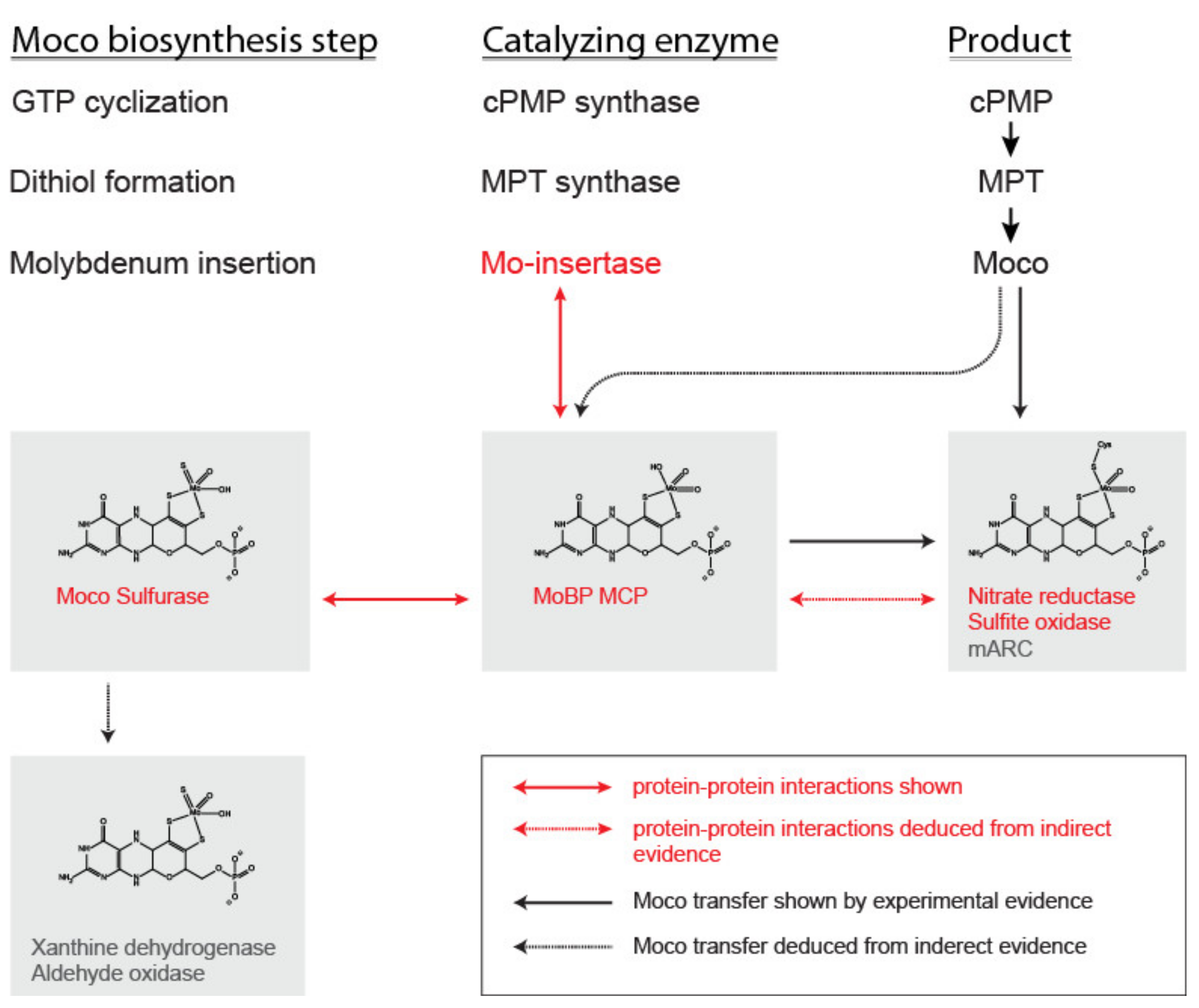
Figure 2.
Molybdenum cofactor synthesis and transfer. The molybdenum cofactor (Moco) biosynthesis reaction steps, catalyzing enzymes, and products are given in the upper third of the figure. The molybdenum insertase (Mo-insertase) is the platform enzyme of the biosynthesis complex and interacts with the cellular transfer system (i.e., Moco binding proteins (MoBP) and putatively also with Moco carrier proteins (MCP)) as well as with nitrate reductase and sulfite oxidase [21]. The interactions of the Mo-insertase with MoBP proteins and the interaction of these proteins with the Moco sulfurase and nitrate reductase [18] are indicated by red arrows. The interaction of MoBP proteins with nitrate reductase and the plant Mo-insertase Cnx1 has been studied in [18]. Dashed lines indicate indirect evidence for the interaction, while solid lines indicate direct evidence. The transfer of Moco from source to sink is indicated by black arrows. Again dashed lines indicate indirect evidence, while solid lines indicate direct evidence for Moco transfer.The interaction network of Arabidopsis thaliana (A. thaliana) MoBP proteins within the cell has been studied in [21], while the interaction of MCP with the Mo-insertase has not been studied as of yet. Upon synthesis, Moco can be transferred on plant sulfite oxidase in vitro [23]. Indirect evidence for the interaction of C. reinhardtii MCP with Moco free nitrate reductase, present in the nit-1 system was described in [14]. Moco binding proteins were shown to enhance the formation of functional nitrate reductase assembly in the nit-1 system [18], while C. reinhardtii as well as V. carteri MCP proteins were found to serve as Moco donors here [16,17]. No evidence for the transfer of Moco from MoBP proteins to the (plant) Moco sulfurase ABA3 has been described in the literature as yet. The transfer of sulfurated Moco from ABA3 to the enzymes of the Xanthine oxidase family can be assumed.
Figure 2.
Molybdenum cofactor synthesis and transfer. The molybdenum cofactor (Moco) biosynthesis reaction steps, catalyzing enzymes, and products are given in the upper third of the figure. The molybdenum insertase (Mo-insertase) is the platform enzyme of the biosynthesis complex and interacts with the cellular transfer system (i.e., Moco binding proteins (MoBP) and putatively also with Moco carrier proteins (MCP)) as well as with nitrate reductase and sulfite oxidase [21]. The interactions of the Mo-insertase with MoBP proteins and the interaction of these proteins with the Moco sulfurase and nitrate reductase [18] are indicated by red arrows. The interaction of MoBP proteins with nitrate reductase and the plant Mo-insertase Cnx1 has been studied in [18]. Dashed lines indicate indirect evidence for the interaction, while solid lines indicate direct evidence. The transfer of Moco from source to sink is indicated by black arrows. Again dashed lines indicate indirect evidence, while solid lines indicate direct evidence for Moco transfer.The interaction network of Arabidopsis thaliana (A. thaliana) MoBP proteins within the cell has been studied in [21], while the interaction of MCP with the Mo-insertase has not been studied as of yet. Upon synthesis, Moco can be transferred on plant sulfite oxidase in vitro [23]. Indirect evidence for the interaction of C. reinhardtii MCP with Moco free nitrate reductase, present in the nit-1 system was described in [14]. Moco binding proteins were shown to enhance the formation of functional nitrate reductase assembly in the nit-1 system [18], while C. reinhardtii as well as V. carteri MCP proteins were found to serve as Moco donors here [16,17]. No evidence for the transfer of Moco from MoBP proteins to the (plant) Moco sulfurase ABA3 has been described in the literature as yet. The transfer of sulfurated Moco from ABA3 to the enzymes of the Xanthine oxidase family can be assumed.


{kind=link}
{kind=link}
Table 1.
Moco carrier and Moco binding proteins. The accession numbers of known and characterized Moco carrier (MCP) and Moco binding proteins (MoBP) and the corresponding publication where these were first described are tabulated. Ataya 2003 [15], Kruse 2010 [18], Krausze 2020 [20], Hercher 2020 [17].
Table 1.
Moco carrier and Moco binding proteins. The accession numbers of known and characterized Moco carrier (MCP) and Moco binding proteins (MoBP) and the corresponding publication where these were first described are tabulated. Ataya 2003 [15], Kruse 2010 [18], Krausze 2020 [20], Hercher 2020 [17].
| Accession Number | Annotation | Publication | Organism |
|---|---|---|---|
| AY039706 | CeMcp1 | [15] | Chlamydomonas reinhardtii |
| At2g28305 | MoBP1 | [18] | Arabidopsis thaliana |
| At5g11950 | MoBP2 | ||
| At2g37210 | MoBP3 | ||
| At4g35190 | MoBP4 | ||
| At3g53450 | MoBP5 | ||
| At5g03270 | MoBP6 | ||
| WP_012595913 | RoMCP | [20] | Rippkaea orientalis |
| XP_002954772.1 | VcMCP | [17] | Volvox carteri |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kruse, T. Moco Carrier and Binding Proteins. Molecules 2022, 27, 6571. https://doi.org/10.3390/molecules27196571
AMA Style
Kruse T. Moco Carrier and Binding Proteins. Molecules. 2022; 27(19):6571. https://doi.org/10.3390/molecules27196571
Chicago/Turabian StyleKruse, Tobias. 2022. "Moco Carrier and Binding Proteins" Molecules 27, no. 19: 6571. https://doi.org/10.3390/molecules27196571
APA StyleKruse, T. (2022). Moco Carrier and Binding Proteins. Molecules, 27(19), 6571. https://doi.org/10.3390/molecules27196571