
From Myricetin to the Discovery of Novel Natural Human ENPP1 Inhibitors: A Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and MM/GBSA Study
Abstract
:1. Introduction
2. Results
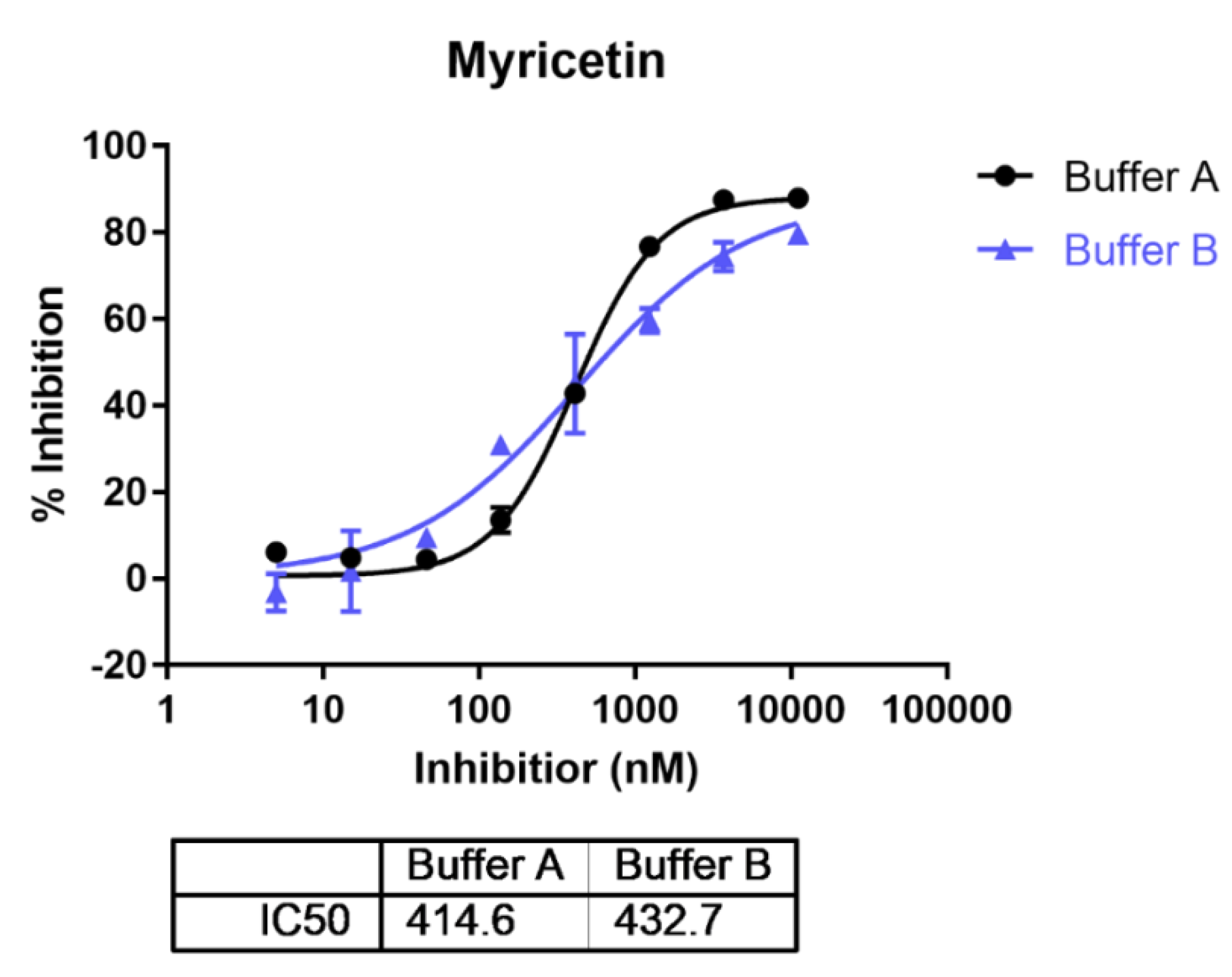
2.1. In Vitro ENPP1 Inhibition by Myricetin and pH Does Not Influence Its Activity
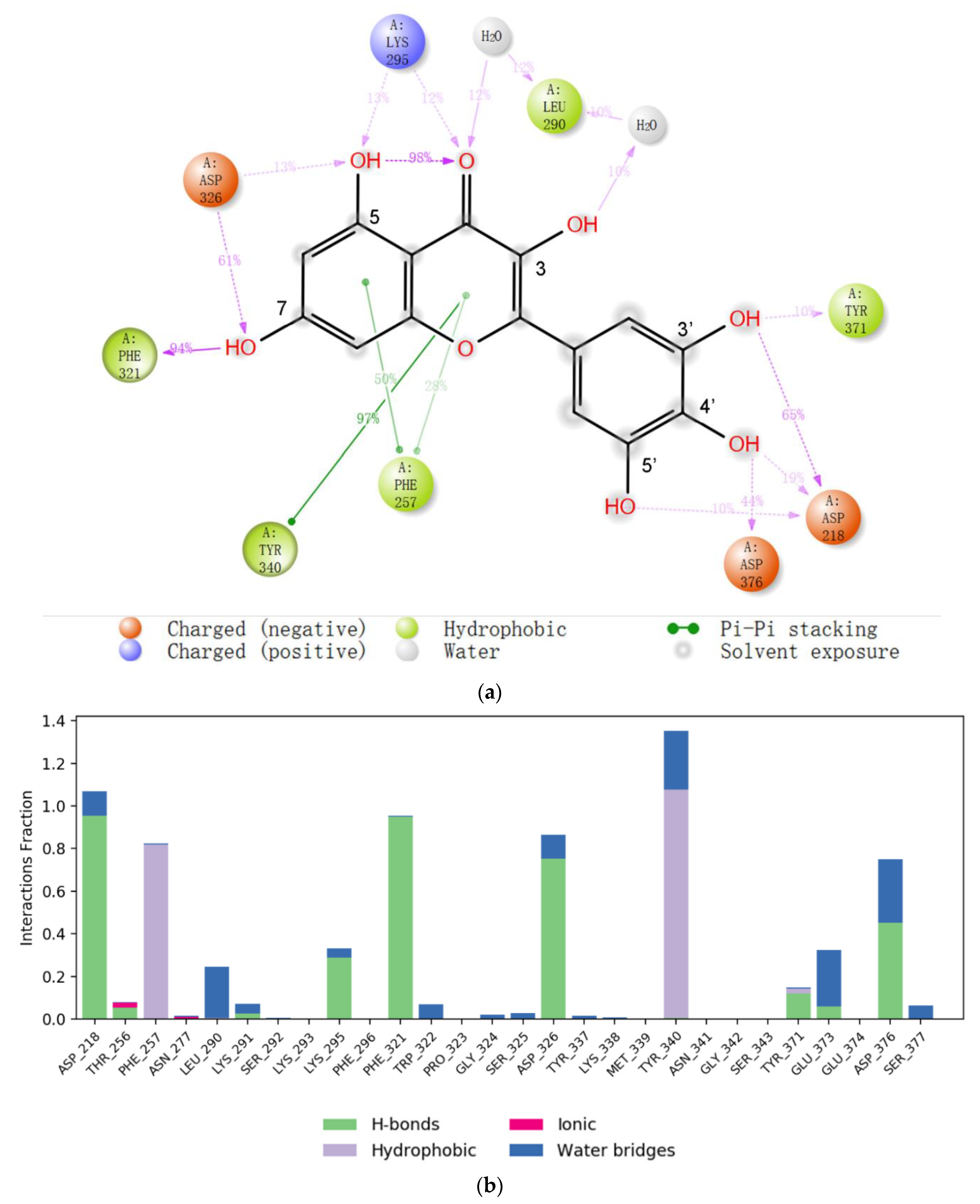
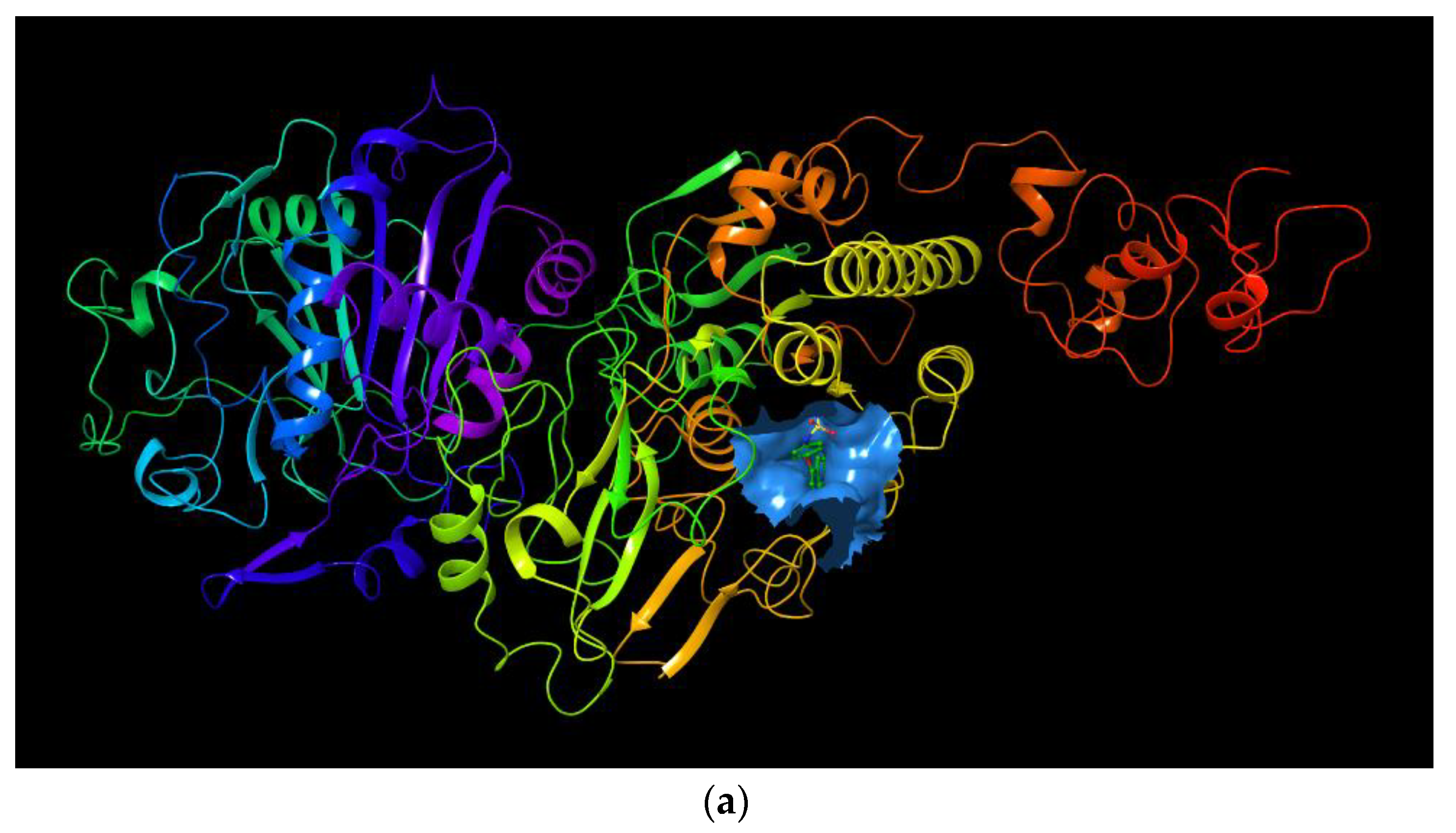
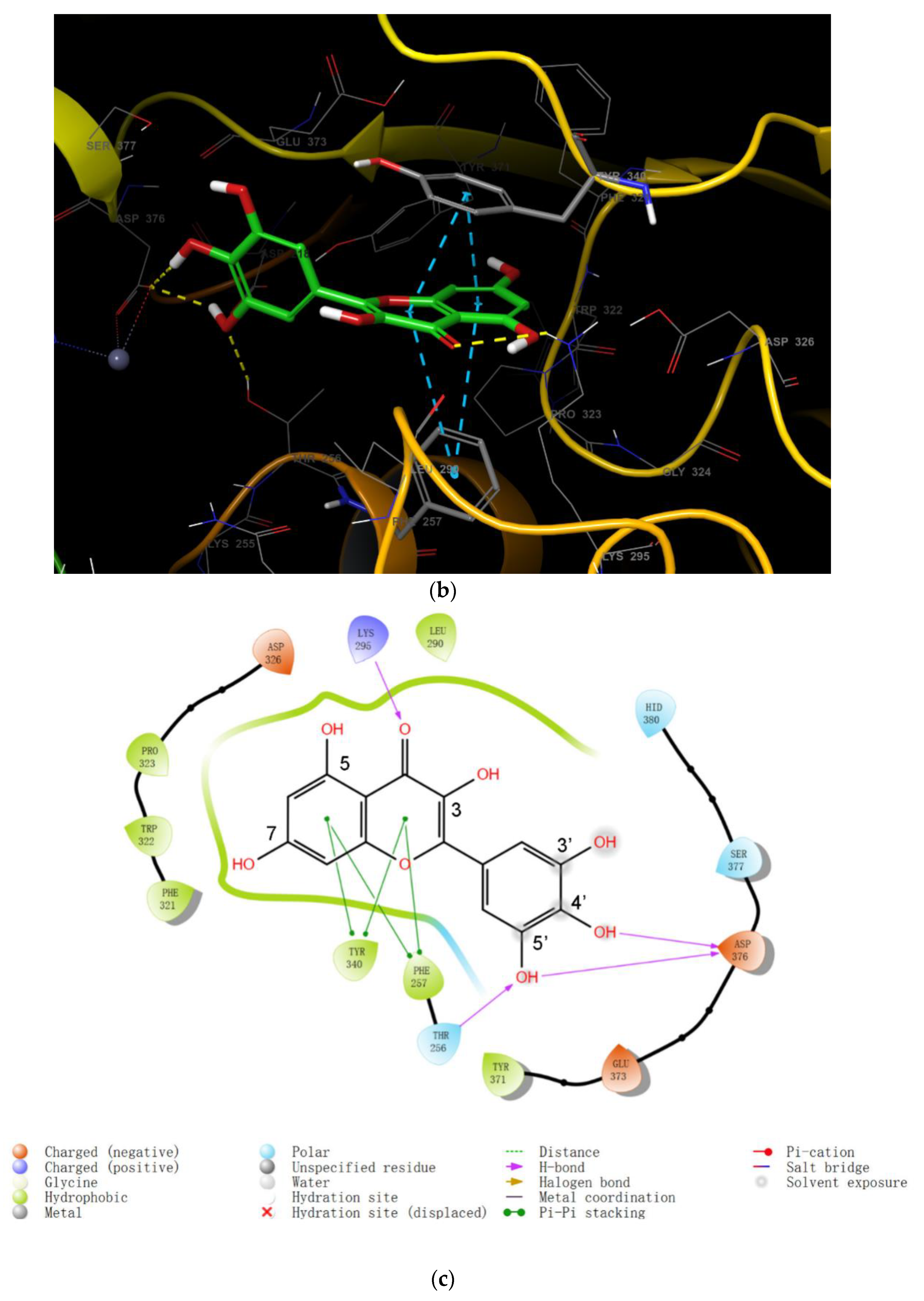
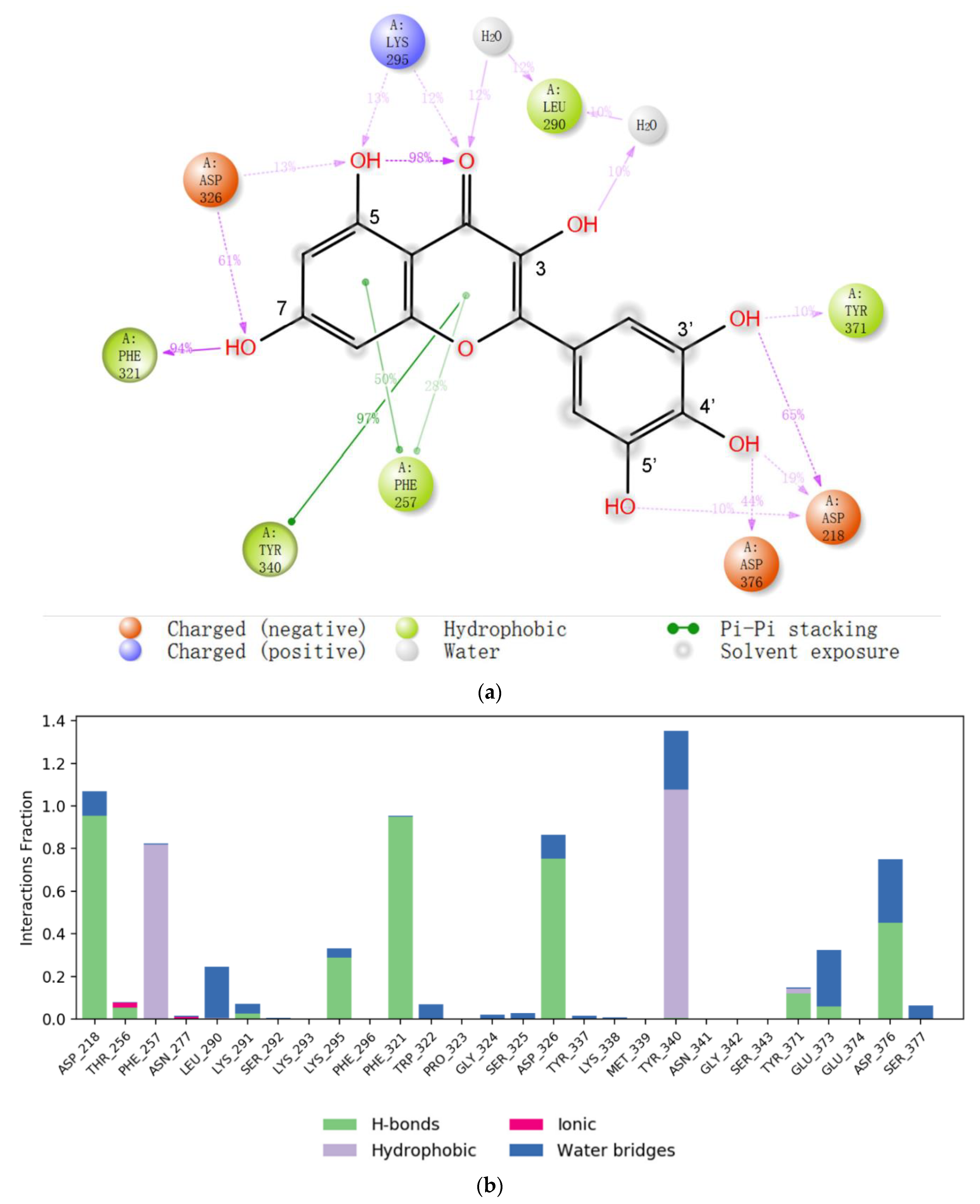
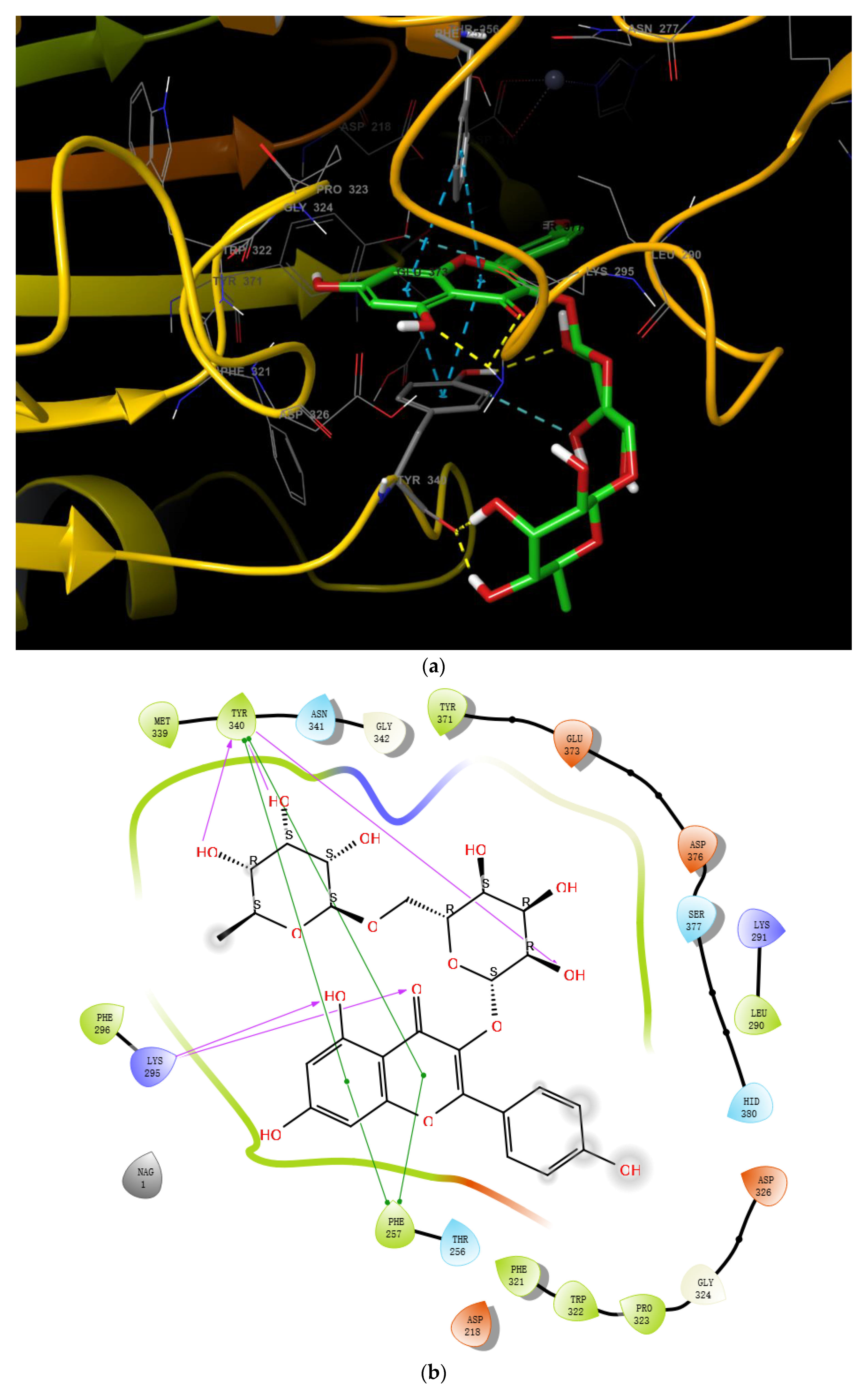
2.2. Molecular Docking of Myricetin with ENPP1 and Further Validations
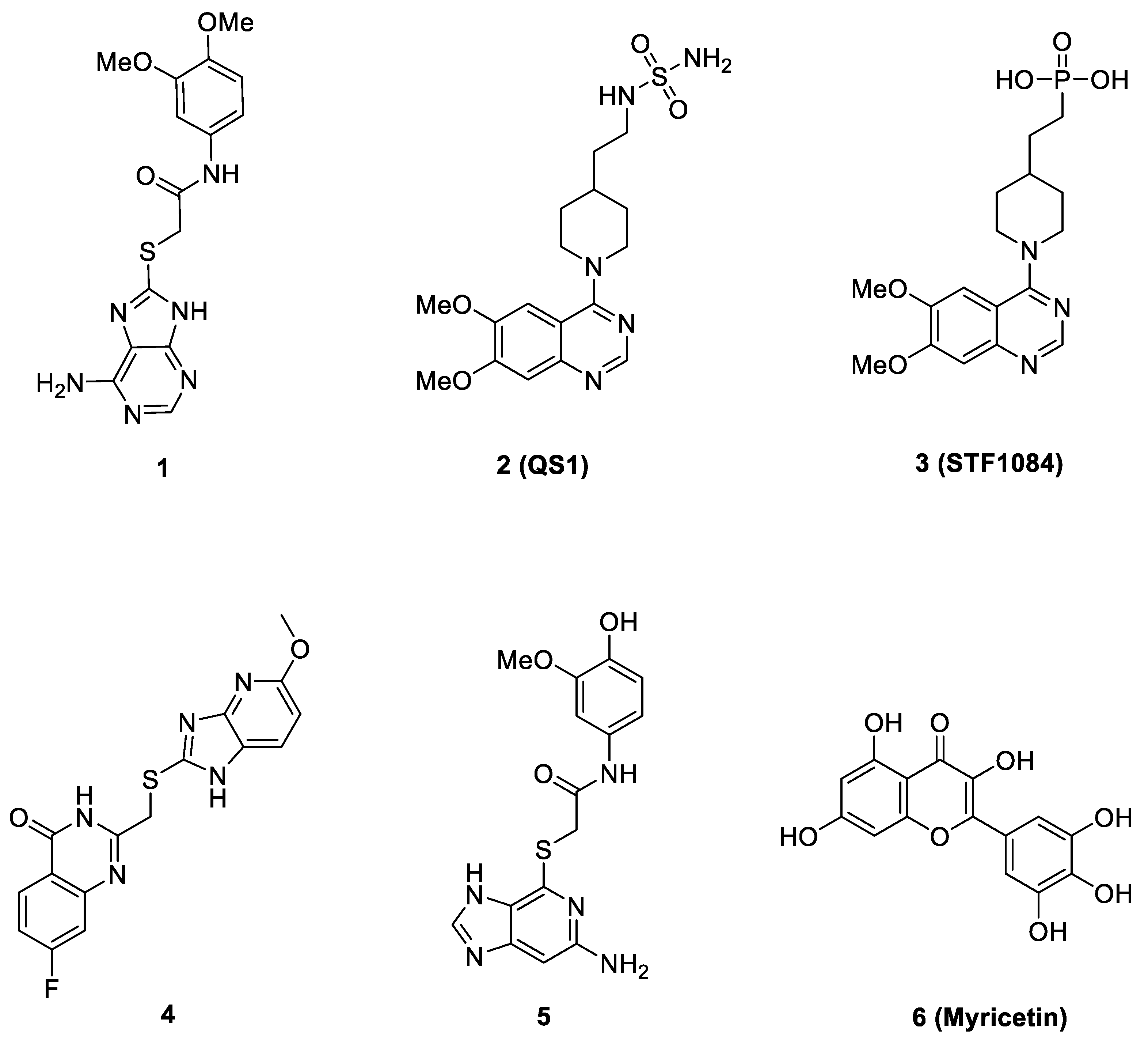
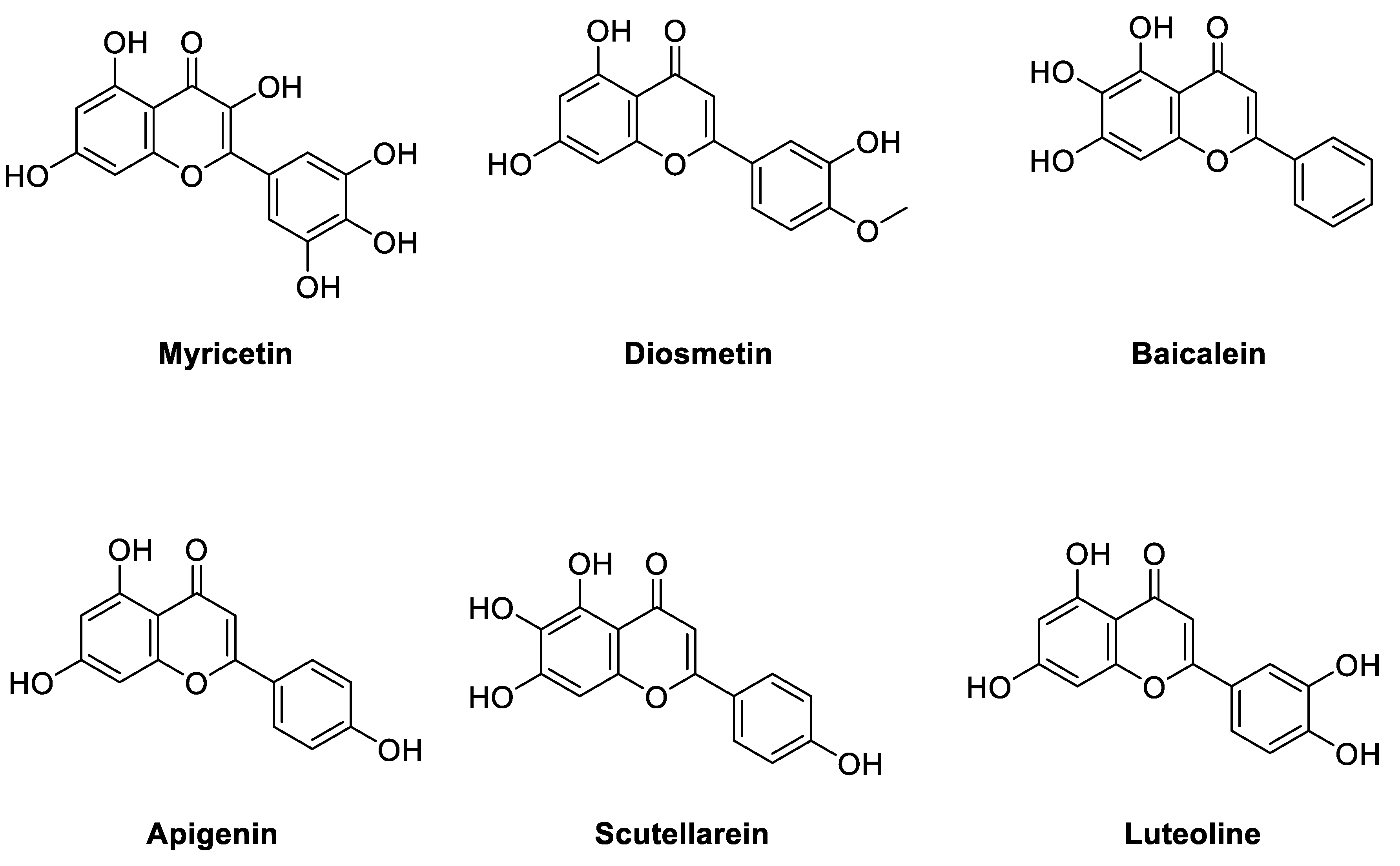
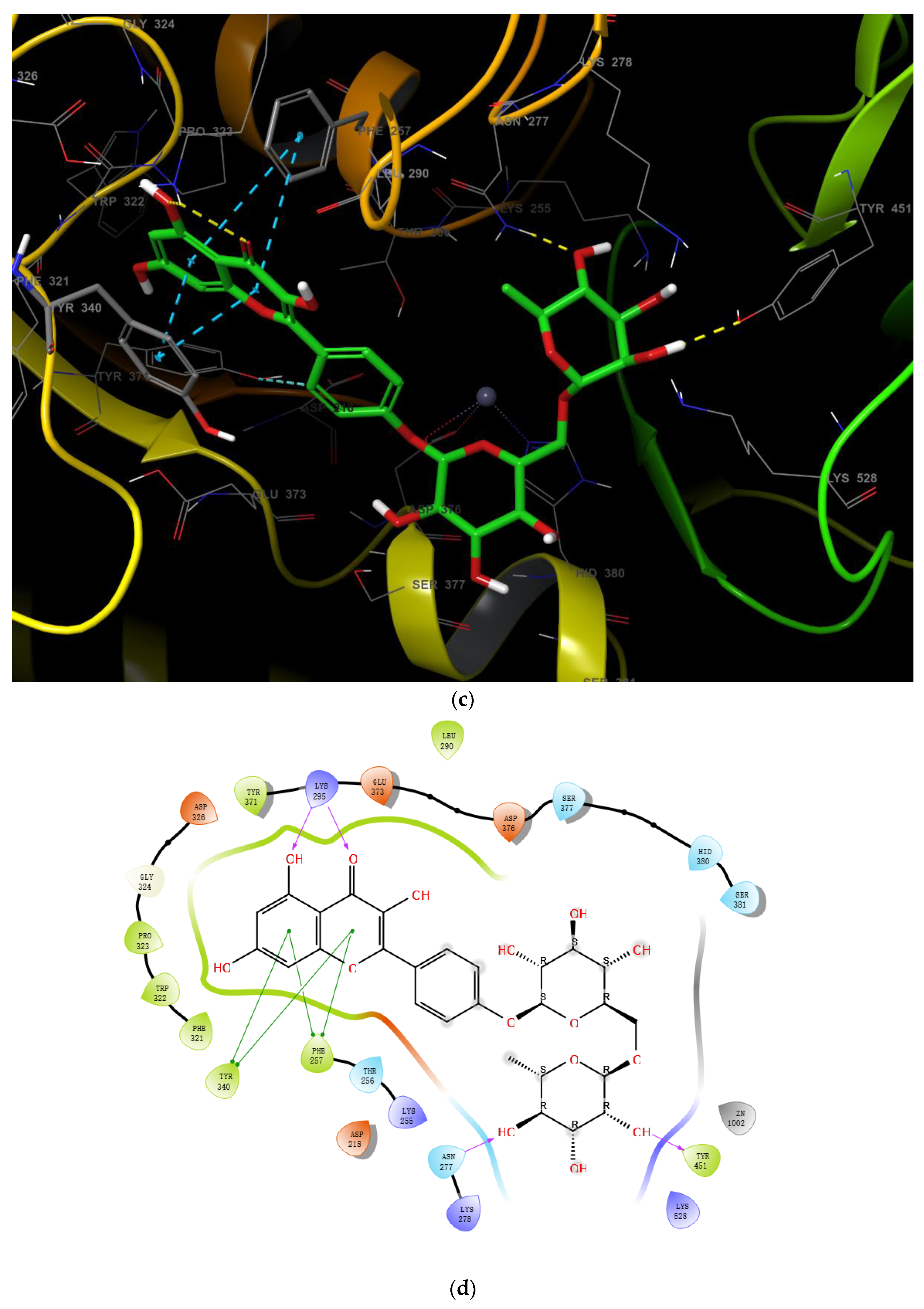
2.3. Virtual Screening of Novel ENPP1 Inhibitors
3. Discussion
4. Materials and Methods
4.1. Molecular Docking and Structure-Based Virtual Screening
4.1.1. Receptor Preparation
4.1.2. Receptor Grid Generation
4.1.3. Preparation of Flavonoid Natural Products Database
4.1.4. Molecular Docking Studies of Myricetin
4.1.5. Identification of Novel Human ENPP1 Inhibitors by Structure-Based Virtual Screening of FNPD
4.2. Molecular Dynamics Simulation and Analysis
4.3. In Vitro ENPP1 Enzyme Inhibition Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Borza, R.; Salgado-Polo, F.; Moolenaar, W.H.; Perrakis, A. Structure and function of the ecto-nucleotide pyrophosphatase/phosphodiesterase (ENPP) family: Tidying up diversity. J. Biol. Chem. 2022, 298, 101526. [Google Scholar] [CrossRef] [PubMed]
- Perez-Valencia, J.A.; Prosdocimi, F.; Cesari, I.M.; da Costa, I.R.; Furtado, C.; Agostini, M.; Rumjanek, F.D. Angiogenesis and evading immune destruction are the main related transcriptomic characteristics to the invasive process of oral tongue cancer. Sci. Rep. 2018, 8, 2007. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, N.C.; Zhu, D.; Milne, E.M.; van’t Hof, R.; Martin, A.; Quarles, L.D.; Millan, J.L.; Farquharson, C.; MacRae, V.E. Altered bone development and an increase in FGF-23 expression in Enpp1(-/-) mice. PLoS ONE 2012, 7, e32177. [Google Scholar] [CrossRef]
- Korostishevsky, M.; Cohen, Z.; Malkin, I.; Ermakov, S.; Yarenchuk, O.; Livshits, G. Morphological and biochemical features of obesity are associated with mineralization genes’ polymorphisms. Int. J. Obes. 2010, 34, 1308–1318. [Google Scholar] [CrossRef]
- Wang, H.; Ye, F.; Zhou, C.; Cheng, Q.; Chen, H. High expression of ENPP1 in high-grade serous ovarian carcinoma predicts poor prognosis and as a molecular therapy target. PLoS ONE 2021, 16, e0245733. [Google Scholar] [CrossRef]
- Ledderose, C.; Hefti, M.M.; Chen, Y.; Bao, Y.; Seier, T.; Li, L.; Woehrle, T.; Zhang, J.; Junger, W.G. Adenosine arrests breast cancer cell motility by A3 receptor stimulation. Purinergic Signal. 2016, 12, 673–685. [Google Scholar] [CrossRef]
- Li, N.; Pan, H.; Cui, M.; Li, Q. Association of K121Q Polymorphism in Ectonucleotide Pyrophosphatase/Phosphodiesterase 1 with Clinical Characteristics of Metabolic Syndrom. Exp. Clin. Endocrinol. Diabetes 2016, 124, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Nishimasu, H.; Okudaira, S.; Mihara, E.; Ishitani, R.; Takagi, J.; Aoki, J.; Nureki, O. Crystal structure of Enpp1, an extracellular glycoprotein involved in bone mineralization and insulin signaling. Proc. Natl. Acad. Sci. USA 2012, 109, 16876–16881. [Google Scholar] [CrossRef] [PubMed]
- Di Paola, R.; Caporarello, N.; Marucci, A.; Dimatteo, C.; Iadicicco, C.; del Guerra, S.; Prudente, S.; Sudano, D.; Miele, C.; Parrino, C.; et al. ENPP1 affects insulin action and secretion: Evidences from in vitro studies. PLoS ONE 2011, 6, e19462. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.N.; Dallas-Yang, Q.; Liu, F.; Ho, T.; Ellsworth, K.; Fischer, P.; Natasha, T.; Ireland, C.; Lu, P.; Li, C.; et al. Evidence that inhibition of insulin receptor signaling activity by PC-1/ENPP1 is dependent on its enzyme activity. Eur. J. Pharmacol. 2009, 606, 17–24. [Google Scholar] [CrossRef]
- Szeri, F.; Niaziorimi, F.; Donnelly, S.; Fariha, N.; Tertyshnaia, M.; Patel, D.; Lundkvist, S.; van de Wetering, K. The Mineralization Regulator ANKH Mediates Cellular Efflux of ATP, Not Pyrophosphate. J. Bone Miner. Res. 2022, 37, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Huitema, L.F.; Apschner, A.; Logister, I.; Spoorendonk, K.M.; Bussmann, J.; Hammond, C.L.; Schulte-Merker, S. Entpd5 is essential for skeletal mineralization and regulates phosphate homeostasis in zebrafish. Proc. Natl. Acad. Sci. USA 2012, 109, 21372–21377. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Duran, M.A.; Dhanota, N.; Chatila, W.K.; Bettigole, S.E.; Kwon, J.; Sriram, R.K.; Humphries, M.P.; Salto-Tellez, M.; James, J.A.; et al. Metastasis and Immune Evasion from Extracellular cGAMP Hydrolysis. Cancer Discov. 2021, 11, 1212–1227. [Google Scholar] [CrossRef]
- Ritchie, C.; Carozza, J.A.; Li, L. Biochemistry, Cell Biology, and Pathophysiology of the Innate Immune cGAS-cGAMP-STING Pathway. Annu. Rev. Biochem. 2022, 91, 599–628. [Google Scholar] [CrossRef] [PubMed]
- Carozza, J.A.; Bohnert, V.; Nguyen, K.C.; Skariah, G.; Shaw, K.E.; Brown, J.A.; Rafat, M.; von Eyben, R.; Graves, E.E.; Glenn, J.S.; et al. Extracellular cGAMP is a cancer cell-produced immunotransmitter involved in radiation-induced anti-cancer immunity. Nat. Cancer 2020, 1, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Gangar, M.; Goyal, S.; Raykar, D.; Khurana, P.; Martis, A.M.; Goswami, A.; Ghoshal, I.; Patel, K.V.; Nagare, Y.; Raikar, S.; et al. Design, synthesis and biological evaluation studies of novel small molecule ENPP1 inhibitors for cancer immunotherapy. Bioorg. Chem. 2022, 119, 105549. [Google Scholar] [CrossRef]
- Chang, L.; Lee, S.Y.; Leonczak, P.; Rozenski, J.; de Jonghe, S.; Hanck, T.; Muller, C.E.; Herdewijn, P. Imidazopyridine- and purine-thioacetamide derivatives: Potent inhibitors of nucleotide pyrophosphatase/phosphodiesterase 1 (NPP1). J. Med. Chem. 2014, 57, 10080–10100. [Google Scholar] [CrossRef]
- Patel, S.D.; Habeski, W.M.; Cheng, A.C.; de la Cruz, E.; Loh, C.; Kablaoui, N.M. Quinazolin-4-piperidin-4-methyl sulfamide PC-1 inhibitors: Alleviating hERG interactions through structure based design. Bioorg. Med. Chem. Lett. 2009, 19, 3339–3343. [Google Scholar] [CrossRef]
- Wang, X.; Lu, X.; Yan, D.; Zhou, Y.; Tan, X. Development of Novel Ecto-Nucleotide Pyrophosphatase/Phosphodiesterase 1 (ENPP1) Inhibitors for Tumor Immunotherapy. Int. J. Mol. Sci. 2022, 23, 7104. [Google Scholar] [CrossRef]
- Li, S.; Yokota, T.; Wang, P.; Hoeve, J.T.; Ma, F.; Le, T.M.; Abt, E.R.; Zhou, Y.; Wu, R.; Nanthavongdouangsy, M.; et al. Cardiomyocytes disrupt pyrimidine biosynthesis in nonmyocytes to regulate heart repair. J. Clin. Investig. 2022, 132, e156296. [Google Scholar] [CrossRef]
- Prem, P.N.; Sivakumar, B.; Boovarahan, S.R.; Kurian, G.A. Recent advances in potential of Fisetin in the management of myocardial ischemia-reperfusion injury—A systematic review. Phytomedicine 2022, 101, 154123. [Google Scholar] [CrossRef] [PubMed]
- Prem, P.N.; Kurian, G.A. Fisetin attenuates renal ischemia/reperfusion injury by improving mitochondrial quality, reducing apoptosis and oxidative stress. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2022, 395, 547–561. [Google Scholar] [CrossRef]
- Wu, J.; Chen, H.; Qin, J.; Chen, N.; Lu, S.; Jin, J.; Li, Y. Baicalin Improves Cardiac Outcome and Survival by Suppressing Drp1-Mediated Mitochondrial Fission after Cardiac Arrest-Induced Myocardial Damage. Oxidative Med. Cell. Longev. 2021, 2021, 8865762. [Google Scholar] [CrossRef] [PubMed]
- Ashrafizadeh, M.; Samarghandian, S.; Hushmandi, K.; Zabolian, A.; Shahinozzaman, M.; Saleki, H.; Esmaeili, H.; Raei, M.; Entezari, M.; Zarrabi, A.; et al. Quercetin in Attenuation of Ischemic/Reperfusion Injury: A Review. Curr. Mol. Pharmacol. 2021, 14, 537–558. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.A.; Aminuddin, A.; Yunus, M.H.M.; Murthy, J.K.; Hui, C.K.; Ugusman, A. Antioxidative and anti-inflammatory activities of Polygonum minus: A review of literature. Rev. Cardiovasc. Med. 2020, 21, 275–287. [Google Scholar]
- Li, J.; Gu, Z.; Liu, Y.; Wang, Y.; Zhao, M. Astilbin attenuates cerebral ischemia/reperfusion injury by inhibiting the TLR4/MyD88/NF-kappaB pathway. Toxicol. Res. 2019, 8, 1002–1008. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Wang, X.; Xu, Z.; Liu, Z.; Ni, Q.; Chu, X.; Qiu, M.; Zhao, A.; Jia, W. Protective effect of flavonoids from Scutellaria baicalensis Georgi on cerebral ischemia injury. J. Ethnopharmacol. 2006, 108, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Bak, I.; Lekli, I.; Juhasz, B.; Nagy, N.; Varga, E.; Varadi, J.; Gesztelyi, R.; Szabo, G.; Szendrei, L.; Bacskay, I.; et al. Cardioprotective mechanisms of Prunus cerasus (sour cherry) seed extract against ischemia-reperfusion-induced damage in isolated rat hearts. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1329–H1336. [Google Scholar] [CrossRef]
- Zenebe, W.; Pechanova, O.; Bernatova, I. Protective effects of red wine polyphenolic compounds on the cardiovascular system. Exp. Clin. Cardiol. 2001, 6, 153–158. [Google Scholar]
- Song, X.; Tan, L.; Wang, M.; Ren, C.; Guo, C.; Yang, B.; Ren, Y.; Cao, Z.; Li, Y.; Pei, J. Myricetin: A review of the most recent research. Biomed. Pharmacother. 2021, 134, 111017. [Google Scholar] [CrossRef]
- Salimi, A.; Jamali, Z.; Shabani, M. Antioxidant Potential and Inhibition of Mitochondrial Permeability Transition Pore by Myricetin Reduces Aluminium Phosphide-Induced Cytotoxicity and Mitochondrial Impairments. Front. Pharmacol. 2021, 12, 719081. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Lin, C.H.; Liu, R.; Li, C.; Jiao, S.; Yi, X.; Walker, M.J.; Xu, X.M.; Zhao, T.; Huang, P.C.; et al. Myricetin against myocardial injury in rat heat stroke model. Biomed. Pharmacother. 2020, 127, 110194. [Google Scholar] [CrossRef] [PubMed]
- Giri, A.K.; Ianevski, A. High-throughput screening for drug discovery targeting the cancer cell-microenvironment interactions in hematological cancers. Expert Opin. Drug Discov. 2022, 17, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Gavriilidou, A.F.M.; Sokratous, K.; Yen, H.Y.; de Colibus, L. High-Throughput Native Mass Spectrometry Screening in Drug Discovery. Front. Mol. Biosci. 2022, 9, 837901. [Google Scholar] [CrossRef] [PubMed]
- Vasina, M.; Velecky, J.; Planas-Iglesias, J.; Marques, S.M.; Skarupova, J.; Damborsky, J.; Bednar, D.; Mazurenko, S.; Prokop, Z. Tools for computational design and high-throughput screening of therapeutic enzymes. Adv. Drug Deliv. Rev. 2022, 183, 114143. [Google Scholar] [CrossRef]
- Cetin, Y.; Aydinlik, S.; Gungor, A.; Kan, T.; Avsar, T.; Durdagi, S. Review on in silico Methods, High-throughput Screening Techniques, and Cell Culture Based In Vitro Assays for SARS-CoV-2. In Current Medicinal Chemistry; Bentham Science Publishers: Sharjah, United Arab Emirates, 2022. [Google Scholar]
- Lu, S.; He, X.; Yang, Z.; Chai, Z.; Zhou, S.; Wang, J.; Rehman, A.U.; Ni, D.; Pu, J.; Sun, J.; et al. Activation pathway of a G protein-coupled receptor uncovers conformational intermediates as targets for allosteric drug design. Nat. Commun. 2021, 12, 4721. [Google Scholar] [CrossRef]
- Li, L.; Yin, Q.; Kuss, P.; Maliga, Z.; Millan, J.L.; Wu, H.; Mitchison, T.J. Hydrolysis of 2’3’-cGAMP by ENPP1 and design of nonhydrolyzable analogs. Nat. Chem. Biol. 2014, 10, 1043–1048. [Google Scholar] [CrossRef]
- Yuki, H.; Tanaka, Y.; Hata, M.; Ishikawa, H.; Neya, S.; Hoshino, T. Implementation of pi-pi interactions in molecular dynamics simulation. J. Comput. Chem. 2007, 28, 1091–1099. [Google Scholar] [CrossRef]
- Gupta, G.; Siddiqui, M.A.; Khan, M.M.; Ajmal, M.; Ahsan, R.; Rahaman, M.A.; Ahmad, M.A.; Arshad, M.; Khushtar, M. Current Pharmacological Trends on Myricetin. Drug Res. 2020, 70, 448–454. [Google Scholar] [CrossRef]
- Lu, S.; Chen, Y.; Wei, J.; Zhao, M.; Ni, D.; He, X.; Zhang, J. Mechanism of allosteric activation of SIRT6 revealed by the action of rationally designed activators. Acta Pharm. Sin. B 2021, 11, 1355–1361. [Google Scholar] [CrossRef]
- Lu, S.; Ni, D.; Wang, C.; He, X.; Lin, H.; Wang, Z.; Zhang, J. Deactivation Pathway of Ras GTPase Underlies Conformational Substates as Targets for Drug Design. ACS Catal. 2019, 9, 7188–7196. [Google Scholar] [CrossRef]
- Ni, D.; Wei, J.; He, X.; Rehman, A.U.; Li, X.; Qiu, Y.; Pu, J.; Lu, S.; Zhang, J. Discovery of cryptic allosteric sites using reversed allosteric communication by a combined computational and experimental strategy. Chem. Sci. 2020, 12, 464–476. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Sun, H.; Wang, J.; Wang, Z.; Liu, H.; Zhang, J.Z.H.; Hou, T. End-Point Binding Free Energy Calculation with MM/PBSA and MM/GBSA: Strategies and Applications in Drug Design. Chem. Rev. 2019, 119, 9478–9508. [Google Scholar] [CrossRef] [PubMed]
- Dennis, M.L.; Newman, J.; Dolezal, O.; Hattarki, M.; Surjadi, R.N.; Nuttall, S.D.; Pham, T.; Nebl, T.; Camerino, M.; Khoo, P.S.; et al. Crystal structures of human ENPP1 in apo and bound forms. Acta Crystallogr. Sect. D Struct. Biol. 2020, 76, 889–898. [Google Scholar] [CrossRef]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; Hermans, J. Interaction Models for Water in Relation to Protein Hydration. In Intermolecular Forces, Proceedings of the Fourteenth Jerusalem Symposium on Quantum Chemistry and Biochemistry, Jerusalem, Israel, 13–16 April 1981; Pullman, B., Ed.; Springer: Dordrecht, The Netherlands, 1981; pp. 331–342. [Google Scholar]
- Lu, C.; Wu, C.; Ghoreishi, D.; Chen, W.; Wang, L.; Damm, W.; Ross, G.A.; Dahlgren, M.K.; Russell, E.; von Bargen, C.D.; et al. OPLS4: Improving Force Field Accuracy on Challenging Regimes of Chemical Space. J. Chem. Theory Comput. 2021, 17, 4291–4300. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cas No. | Chemical Structures | Docking Score | MM/GBSA ∆G Bind (kcal/mol) |
|---|---|---|---|
| 1397173-50-0 | 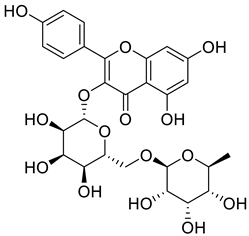 | −12.055 | −27.18 |
| 1169835-58-8 | 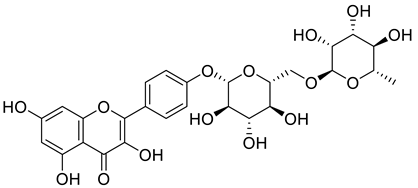 | −12.324 | −19.73 |
| 529-44-2 (Myricetin) | 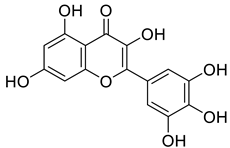 | −9.574 | −11.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, S.; Shao, Z. From Myricetin to the Discovery of Novel Natural Human ENPP1 Inhibitors: A Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and MM/GBSA Study. Molecules 2022, 27, 6175. https://doi.org/10.3390/molecules27196175
Song S, Shao Z. From Myricetin to the Discovery of Novel Natural Human ENPP1 Inhibitors: A Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and MM/GBSA Study. Molecules. 2022; 27(19):6175. https://doi.org/10.3390/molecules27196175
Chicago/Turabian StyleSong, Shaohan, and Zhiyu Shao. 2022. "From Myricetin to the Discovery of Novel Natural Human ENPP1 Inhibitors: A Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and MM/GBSA Study" Molecules 27, no. 19: 6175. https://doi.org/10.3390/molecules27196175
APA StyleSong, S., & Shao, Z. (2022). From Myricetin to the Discovery of Novel Natural Human ENPP1 Inhibitors: A Virtual Screening, Molecular Docking, Molecular Dynamics Simulation, and MM/GBSA Study. Molecules, 27(19), 6175. https://doi.org/10.3390/molecules27196175