
Root-Associated Microbiomes of Panax notoginseng under the Combined Effect of Plant Development and Alpinia officinarum Hance Essential Oil


Abstract
:1. Introduction
2. Results
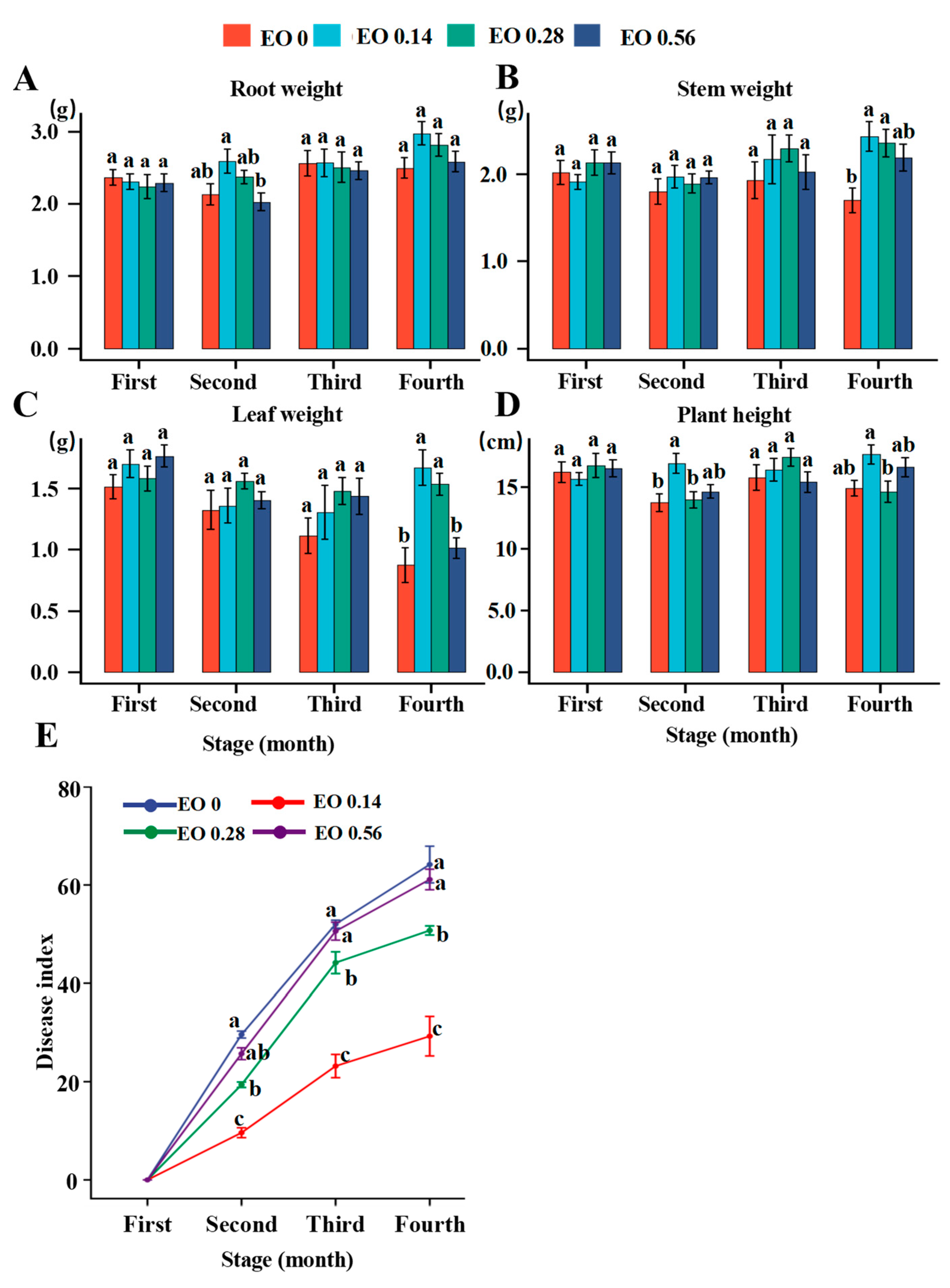
2.1. Effects of EO on Plant Growth, Disease Occurrence and Saponin Content
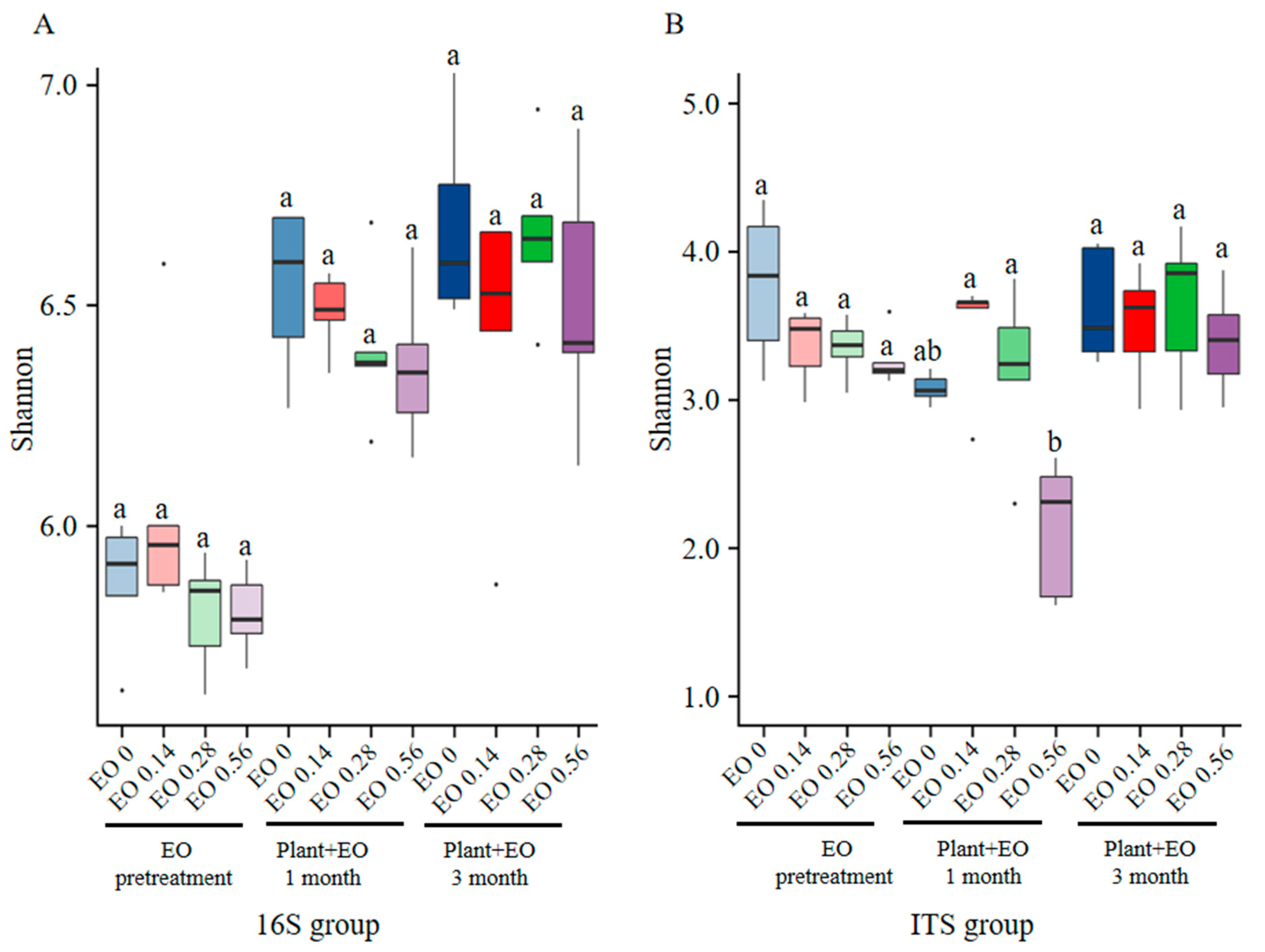
2.2. Different Responses of Bacterial and Fungal Diversity to Plant Development and EO Treatment
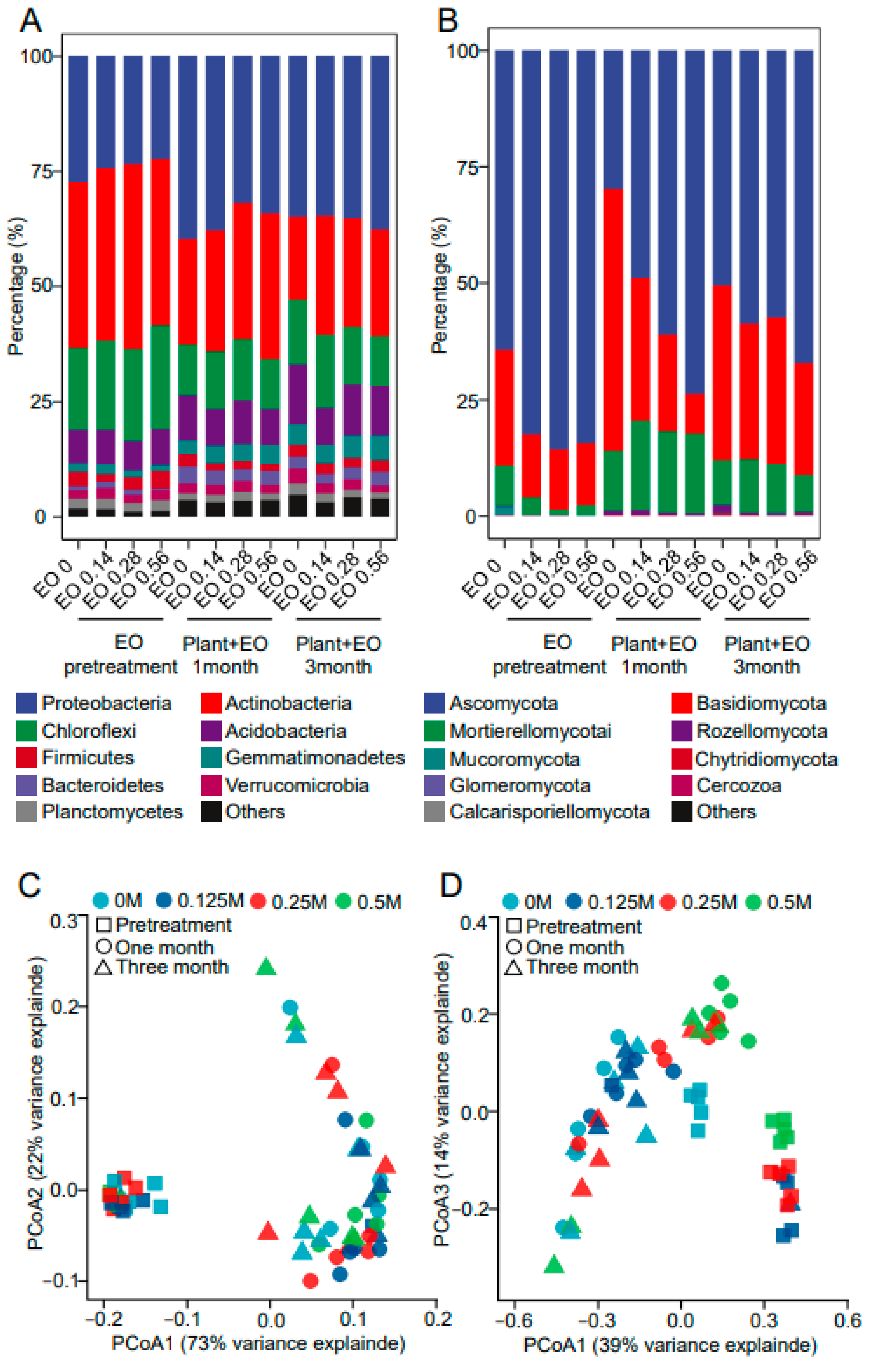
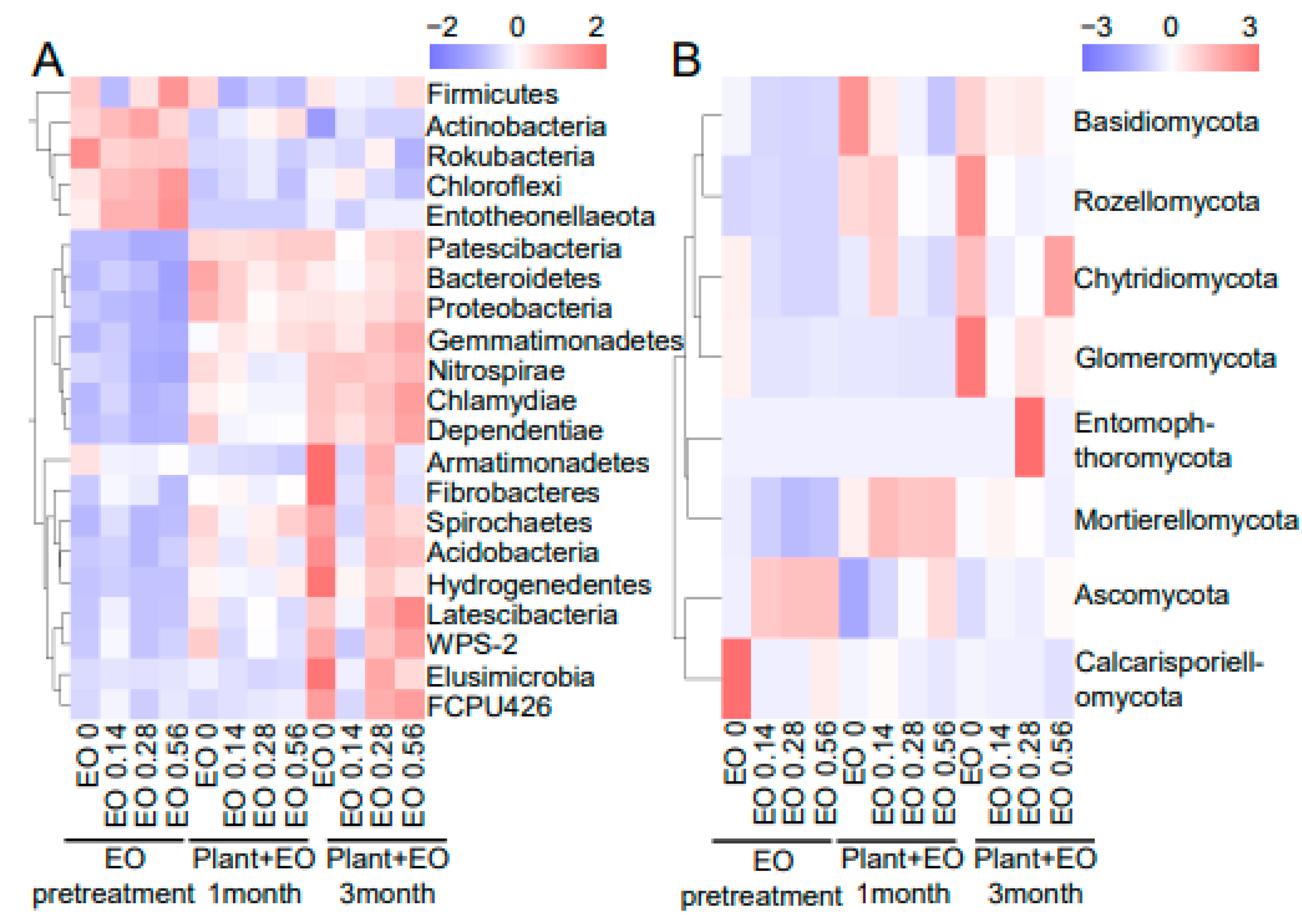
2.3. Bacterial and Fungal Community Composition in Response to Plant Advancement and EO Treatment
2.4. Correlations of Plant Biomass, Disease Index, and Saponin Content with Bacteria and Fungi
3. Materials and Methods
3.1. General Experimental Design
3.2. Preparation of EOs
3.3. Pot Experiment and Sample Collection
3.4. Determination of Biomass, Disease Index, and Saponin Content
3.5. DNA Extraction and Amplicon Sequencing
3.6. Analysis of Sequencing Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, T.; Xiu, M.C.; Zong, H.S.; Zhao, K.J.; Zhao, N.J.; Lo, C.K.; Tsim, K. Chemical assessment of roots of Panax notoginseng in China: Regional and seasonal variations in its active constituents. J. Agric. Food Chem. 2003, 51, 4617–4623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.L.; Wang, W.Q. Progress on formation mechanism and control measurements of continuous cropping obstacles in plants. J. Biol. 2010, 27, 69–72. [Google Scholar]
- Han, X.-B.; Zhao, J.; Cao, J.-M.; Zhang, C.-S. Essential oil of Chrysanthemum indicum L.: Potential biocontrol agent against plant pathogen Phytophthora nicotianae. Environ. Sci. Pollut. Res. Int. 2019, 26, 7013–7023. [Google Scholar] [CrossRef]
- Isman, M.B.; Miresmailli, S.; Machial, C. Commercial opportunities for pesticides based on plant essential oils in agriculture, industry and consumer products. Phytochem. Rev. 2011, 10, 197–204. [Google Scholar] [CrossRef]
- Radulovic, N.S.; Blagojevic, P.D.; Stojanovic-Radic, Z.Z.; Stojanovic, N.M. Antimicrobial plant metabolites: Structural diversity and mechanism of action. Curr. Med. Chem. 2013, 20, 932–952. [Google Scholar] [PubMed]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Minerdi, D.; Bossi, S.; Gullino, M.L.; Garibaldi, A. Volatile organic compounds: A potential direct long-distance mechanism for antagonistic action of Fusarium oxysporum strain MSA 35. Environ. Microbiol. 2010, 11, 844–854. [Google Scholar] [CrossRef]
- Tomislav, C.; Henry, M.; Aschenbrenner, I.A.; Martin, G.; Gabriele, B. Analyzing the antagonistic potential of the lichen microbiome against pathogens by bridging metagenomic with culture studies. Front. Microbiol. 2015, 6, 620. [Google Scholar]
- Adam, O.; Sylwia, J.; Paolina, G.; Rita, G. The antimicrobial volatile power of the rhizospheric isolate Pseudomonas donghuensis P482. PLoS ONE 2017, 12, e0174362. [Google Scholar]
- Paolina, G.; Cornelis, H.; Saskia, G.; Wietse, D.B. Volatiles produced by the mycophagous soil bacterium Collimonas. FEMS Microbiol. Ecol. 2014, 87, 639–649. [Google Scholar]
- Yuan, X.; Hong, S.; Xiong, W.; Raza, W.; Shen, Z.; Wang, B.; Li, R.; Ruan, Y.; Shen, Q.; Dini-Andreote, F. Development of fungal-mediated soil suppressiveness against Fusarium wilt disease via plant residue manipulation. Microbiome 2021, 9, 200. [Google Scholar] [CrossRef] [PubMed]
- Attaran, E.; Rostas, M.J. Pseudomonas syringae Elicits Emission of the Terpenoid (E,E)-4,8,12-Trimethyl-1,3,7,11-Tridecatetraene in Arabidopsis Leaves Via Jasmonate Signaling and Expression of the Terpene Synthase TPS4. Mol. Plant-Microbe Interact. 2008, 21, 1482–1497. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, R.; Lee, S.; Ryu, C. Microbe-induced plant volatiles. New Phytol. 2017, 220, 684–691. [Google Scholar] [CrossRef]
- Deangelis, K.M.; Brodie, E.L.; Desantis, T.Z.; Andersen, G.L.; Firestone, M.K. Selective progressive response of soil microbial community to wild oat roots. ISME J. 2009, 3, 168–178. [Google Scholar] [CrossRef] [Green Version]
- Clemensson-Lindell, A.; Persson, H. Effects of freezing on rhizosphere and root nutrient content using two soil sampling methods. Plant Soil 1992, 139, 39–45. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Mago, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Aronesty, E. Ea-Utils: Command-Line Tools for Processing Biological Sequencing Data. 2011. Available online: http://code.google.com/p/ea-utils (accessed on 1 October 2015).
- Yang, N.; Li, A. Advances in the research of plant essential oils for pest control. Plant Prot. 2007, 33, 16. [Google Scholar]
- Grande-Tovar, C.D.; Chaves-Lopez, C.; Serio, A.; Rossi, C.; Paparella, A. Chitosan coatings enriched with essential oils: Effects on fungi involved in fruit decay and mechanisms of action. Trends Food Sci. Technol. 2018, 78, 61–71. [Google Scholar] [CrossRef]
- Jian, J.; Xie, Y.; Guo, Y.; Cheng, Y.; He, Q.; Yao, W. The inhibitory effect of plant essential oils on foodborne pathogenic bacteria in food. Crit. Rev. Food Sci. Nutr. 2019, 59, 3281–3292. [Google Scholar]
- Zhang, Y.-M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Bagchi, B.; Pal, K.; Bhandary, S.; Kool, A.; Hoque, N.A.; Biswas, P.; Thakur, P.; Das, K.; Karmakar, P.; et al. Essential oil impregnated luminescent hydroxyapatite: Antibacterial and cytotoxicity studies. Mater. Sci. Eng. C 2020, 116, 12. [Google Scholar] [CrossRef] [PubMed]
- An, P.; Yang, X.; Yu, J.; Qi, J.; Ren, X.; Kong, Q. α-terpineol and terpene-4-ol, the critical components of tea tree oil, exert antifungal activities in vitro and in vivo against Aspergillus niger in grapes by inducing morphous damage and metabolic changes of fungus. Food Control 2019, 98, 42–53. [Google Scholar] [CrossRef]
- Khaleil, M.M.; Alnoman, M.M.; Elrazik, E.S.A.; Zagloul, H.; Khalil, A.M.A. Essential oil of Foeniculum vulgare Mill. as a green fungicide and defense-inducing agent against Fusarium root rot disease in Vicia faba L. Biology 2021, 10, 696. [Google Scholar] [CrossRef] [PubMed]
- Landi, L.; Peralta-Ruiz, Y.; Chaves-López, C.; Romanazzi, G. Chitosan Coating Enriched with Ruta graveolens L. Essential Oil Reduces Postharvest Anthracnose of Papaya (Carica papaya L.) and Modulates Defense-Related Gene Expression. Front. Plant Sci. 2021, 12, 765806. [Google Scholar] [CrossRef]
- Chantal, L.M.; Janette, N.; Mathias, D.; Robert, P.; Birgit, P. mVOC: A database of microbial volatiles. Nucleic Acids Res. 2014, 42, D744–D748. [Google Scholar]
- Effmert, U.; Kalderás, J.; Warnke, R.; Piechulla, B. Volatile Mediated Interactions Between Bacteria and Fungi in the Soil. J. Chem. Ecol. 2012, 38, 665–703. [Google Scholar] [CrossRef]
- Bitas, V.; Kim, H.S.; Bennett, J.W.; Kang, S. Sniffing on microbes: Diverse roles of microbial volatile organic compounds in plant health. Mol. Plant-Microbe Interact. 2013, 26, 835–843. [Google Scholar] [CrossRef]
- Walker, T.S.; Bais, H.P.; Vivanco, G. Root Exudation and Rhizosphere Biology. Plant Physiol. 2003, 132, 44–51. [Google Scholar] [CrossRef]
- Chen, S.; Waghmode, T.R.; Sun, R.; Kuramae, E.E.; Hu, C.; Liu, B. Root-associated microbiomes of wheat under the combined effect of plant development and nitrogen fertilization. Microbiome 2019, 7, 136. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Physiol. 2012, 64, 807–838. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.L.; Li, X.G.; Zhu, F.; Lei, C.L. Structural characterization of nanoparticles loaded with garlic essential oil and their insecticidal activity against Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Agric. Food Chem. 2009, 57, 10156–10162. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.M.; Ma, Y.N.; Yin, Y.J.; Chen, C.J.; Cheng, Y.X. Effects of essential Oils from Zingiberaceae plants on root-rot disease of Panax notoginseng. Molecules 2018, 23, 1021. [Google Scholar] [CrossRef]
- Ma, Y.-N.; Xu, F.-R.; Chen, C.-J.; Li, Q.-Q.; Wang, M.-Z.; Cheng, Y.-X.; Dong, X. The beneficial use of essential oils from buds and fruit of Syzygium aromaticum to combat pathogenic fungi of Panax notoginseng. Ind. Crop. Prod. 2019, 133, 185–192. [Google Scholar] [CrossRef]
- Misra, G.; Pavlostathis, S.G. Biodegradation kinetics of monoterpenes in liquid and soil-slurry systems. Appl. Microbiol. Biotechnol. 1997, 47, 572–577. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential Oils in Insect Control: Low-Risk Products in a High-Stakes World. Annu. Rev. Entomol. 2012, 57, 405. [Google Scholar] [CrossRef] [PubMed]
- Hammerbacher, A.; Coutinho, T.A.; Gershenzon, J. Roles of plant volatiles in defence against microbial pathogens and microbial exploitation of volatiles. Plant Cell Environ. 2019, 42, 2827–2843. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | One Month | Three Months | ||||||
|---|---|---|---|---|---|---|---|---|
| R1 | Rb1 | Rd | Rg1 | R1 | Rb1 | Rd | Rg1 | |
| EO 0 | 2.61 ± 0.25 b | 10.06 ± 1.68 c | 10.82 ± 1.03 b | 17.79 ± 2.12 b | 3.81 ± 0.06 a | 10.43 ± 0.61 b | 8.66 ± 1.13 b | 23.54 ± 4.5 a |
| EO 0.14 | 3.70 ± 0.16 a | 20.28 ± 1.55 a | 34.39 ± 12.9 a | 23.45 ± 1.39 ab | 7.15 ± 3.44 a | 15.73 ± 0.51 a | 17.04 ± 1.63 a | 24.87 ± 1.44 a |
| EO 0.28 | 2.45 ± 0.31 b | 14.93 ± 1.10 b | 20.79 ± 7.96 ab | 22.98 ± 1.58 ab | 4.55 ± 0.80 a | 10.07 ± 1.09 b | 7.70 ± 0.87 b | 20.30 ± 1.24 a |
| EO 0.56 | 3.05 ± 0.64 ab | 17.59 ± 0.64 b | 26.81 ± 1.89 ab | 25.94 ± 3.49 a | 2.79 ± 0.36 a | 11.61 ± 0.9 b | 8.64 ± 2.66 b | 23.36 ± 1.83 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.-L.; Liu, X.-Y.; Zhou, C.; Han, S.-F.; Xu, F.-R.; Dong, X. Root-Associated Microbiomes of Panax notoginseng under the Combined Effect of Plant Development and Alpinia officinarum Hance Essential Oil. Molecules 2022, 27, 6014. https://doi.org/10.3390/molecules27186014
Zhou H-L, Liu X-Y, Zhou C, Han S-F, Xu F-R, Dong X. Root-Associated Microbiomes of Panax notoginseng under the Combined Effect of Plant Development and Alpinia officinarum Hance Essential Oil. Molecules. 2022; 27(18):6014. https://doi.org/10.3390/molecules27186014
Chicago/Turabian StyleZhou, Han-Lin, Xiao-Yun Liu, Chao Zhou, Si-Fang Han, Fu-Rong Xu, and Xian Dong. 2022. "Root-Associated Microbiomes of Panax notoginseng under the Combined Effect of Plant Development and Alpinia officinarum Hance Essential Oil" Molecules 27, no. 18: 6014. https://doi.org/10.3390/molecules27186014
APA StyleZhou, H.-L., Liu, X.-Y., Zhou, C., Han, S.-F., Xu, F.-R., & Dong, X. (2022). Root-Associated Microbiomes of Panax notoginseng under the Combined Effect of Plant Development and Alpinia officinarum Hance Essential Oil. Molecules, 27(18), 6014. https://doi.org/10.3390/molecules27186014