
Identification of Gedunin from a Phytochemical Depository as a Novel Multidrug Resistance-Bypassing Tubulin Inhibitor of Cancer Cells


 ,
, 
Abstract
:1. Introduction
2. Results
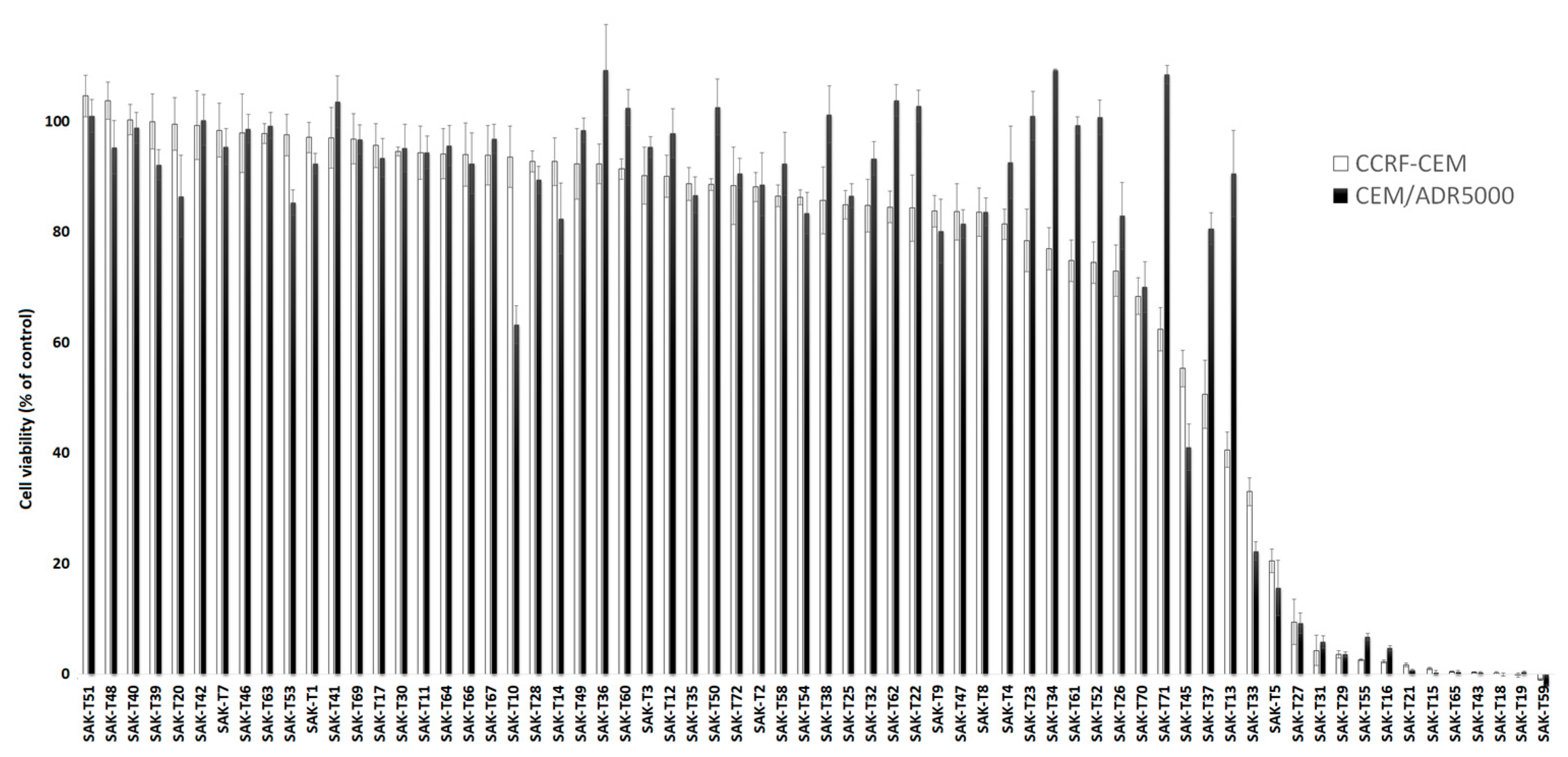
2.1. Activity Screening of 69 Phytochemicals in Leukemia Cells
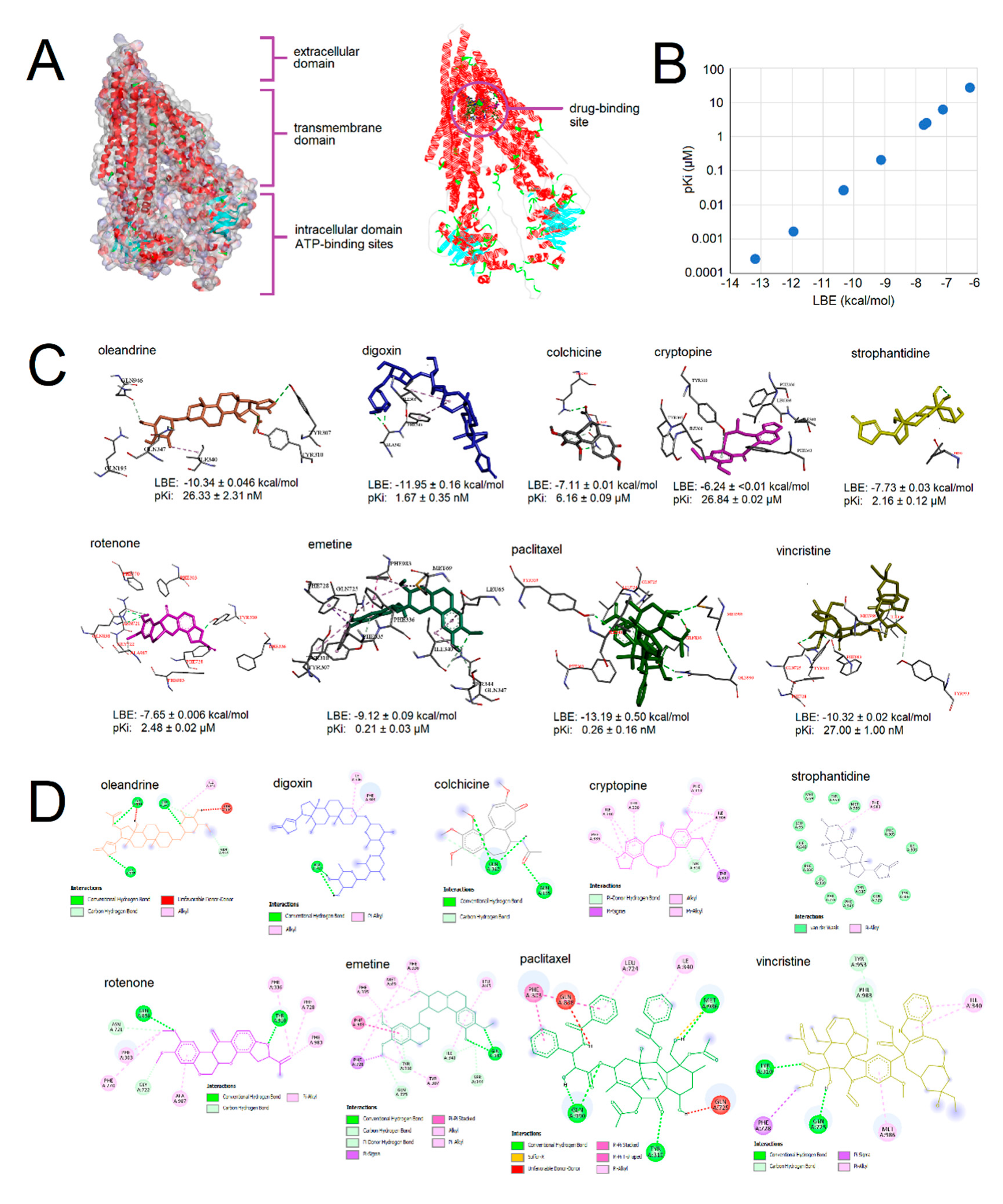
2.2. Molecular Docking of Selected Phytochemicals to P-Glycoprotein
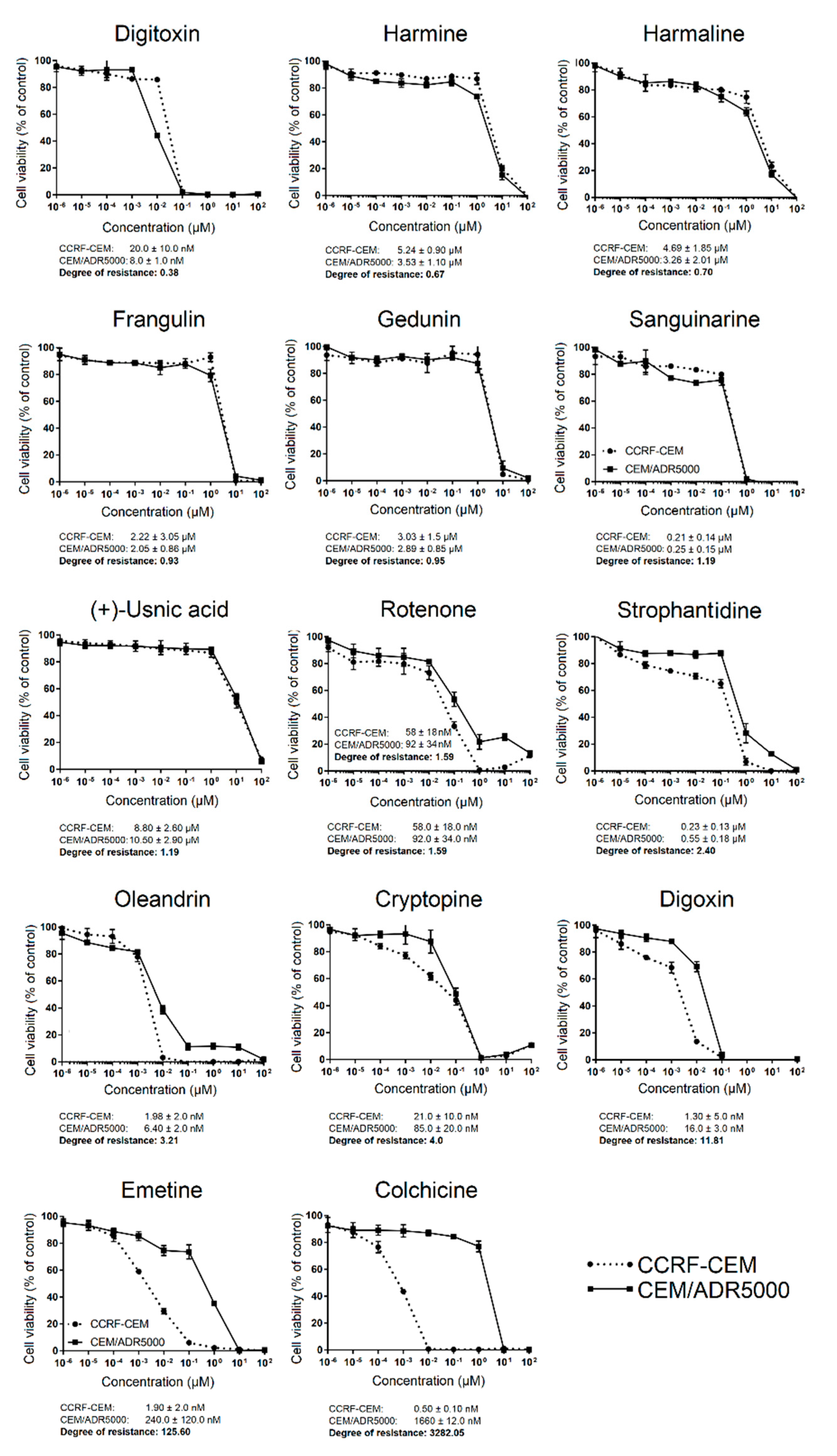
2.3. Activity Profiling of Selected Phytochemicals in Cell Lines of Diverse Tumor Types
2.4. Acute and Chronic Toxicity of Selected Phytochemicals
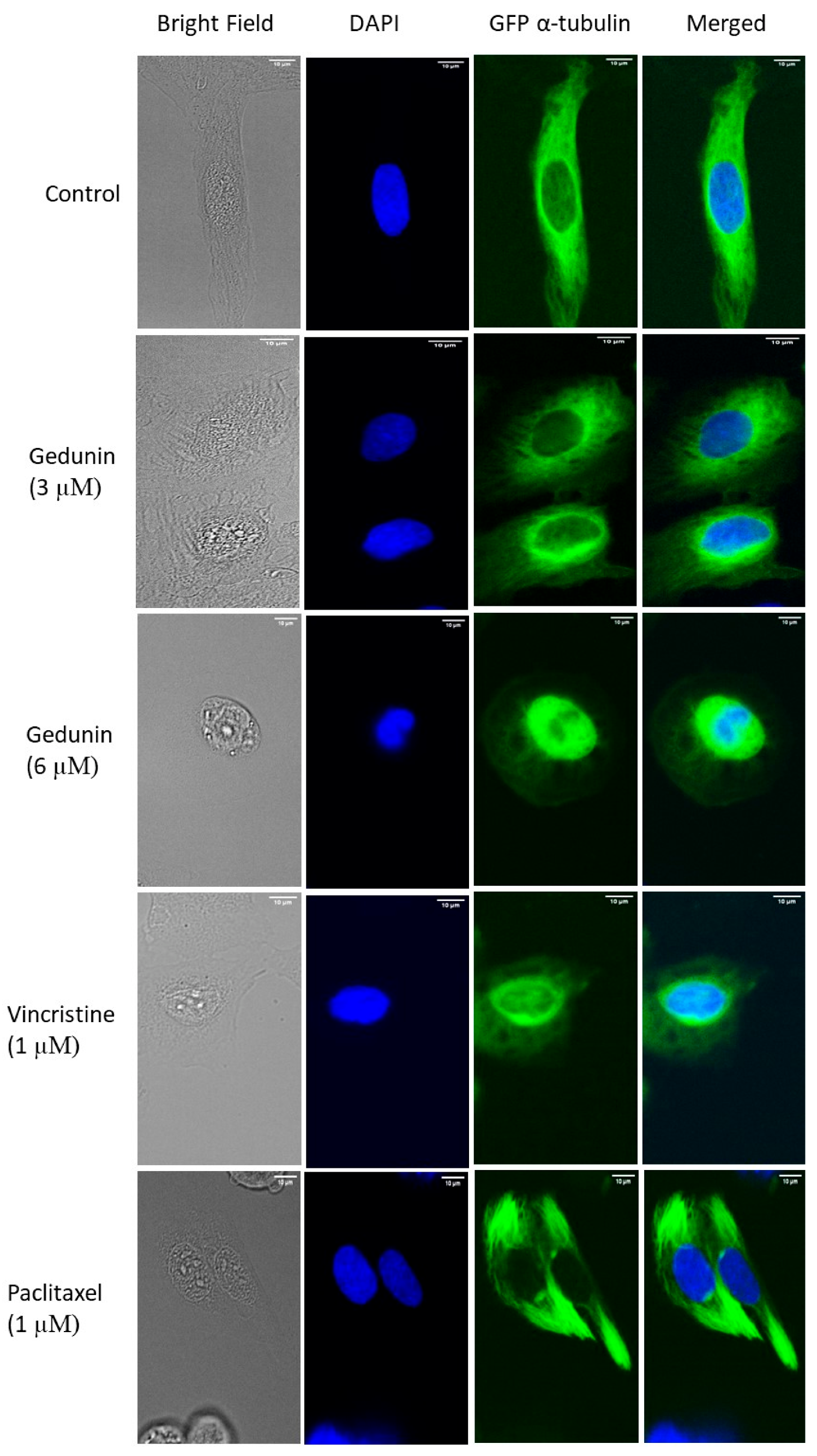
2.5. Effect of Gedunin on the Microtubule Network
2.6. Molecular Docking of Gedunin to β-Tubulin
2.7. Proteome Profiling of Gedunin
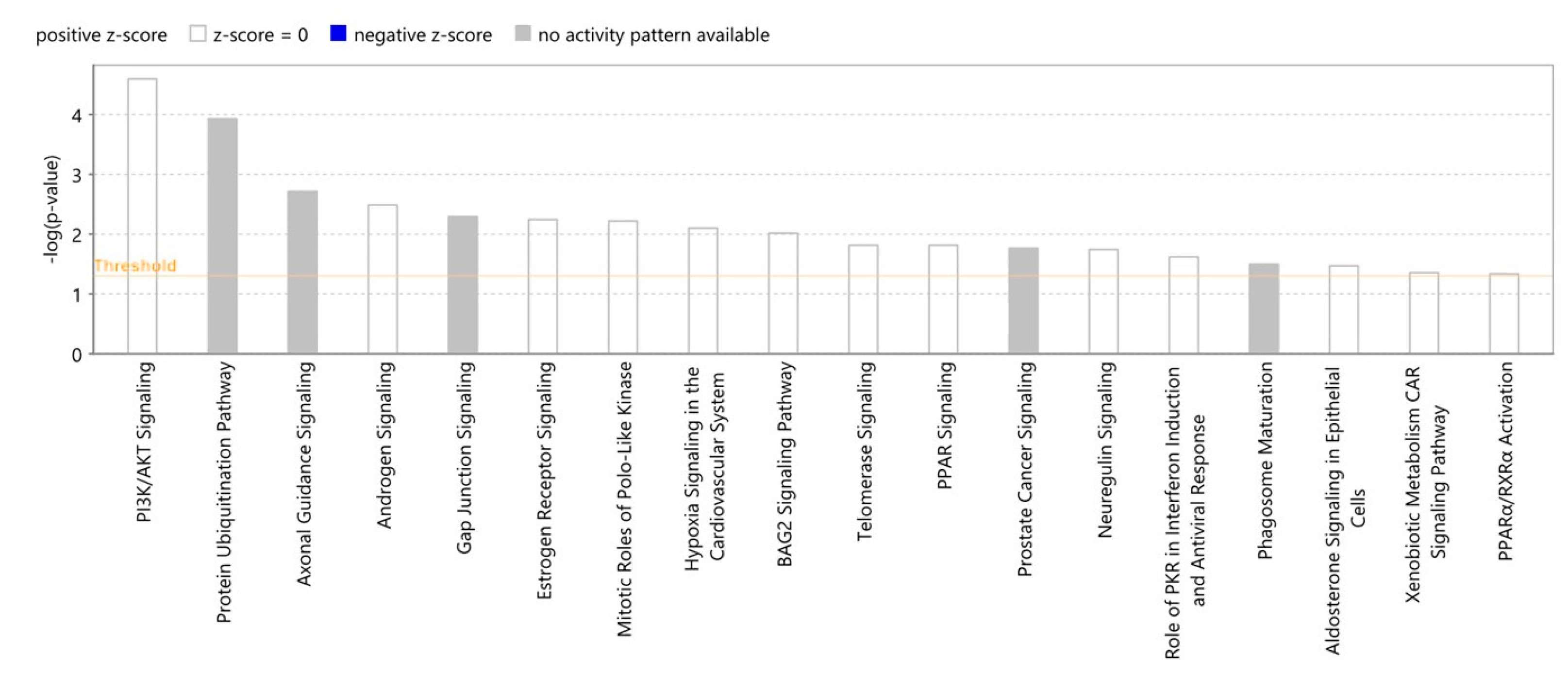
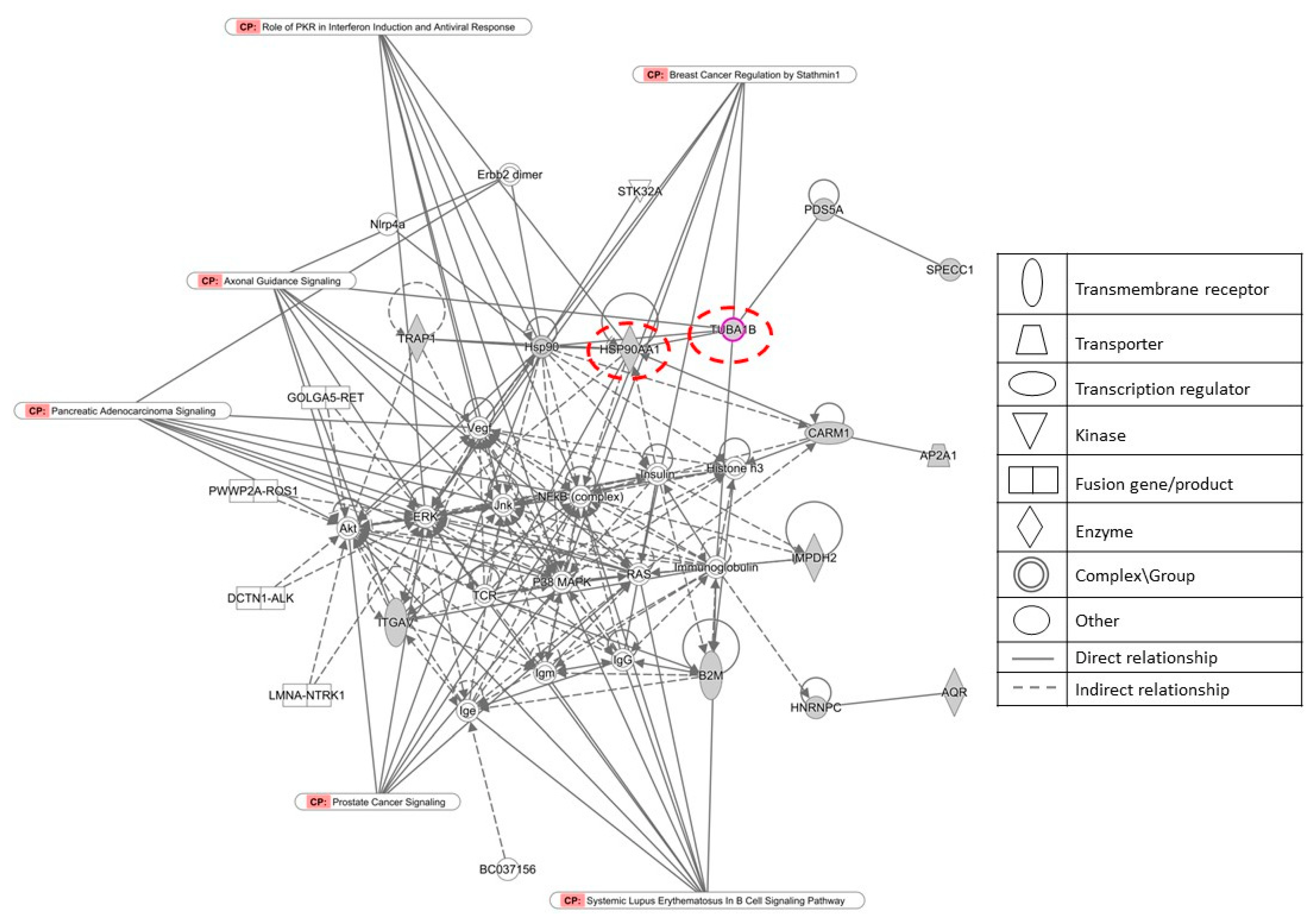
2.8. Proteomic Canonical Pathway and Network Analyses of Gedunin
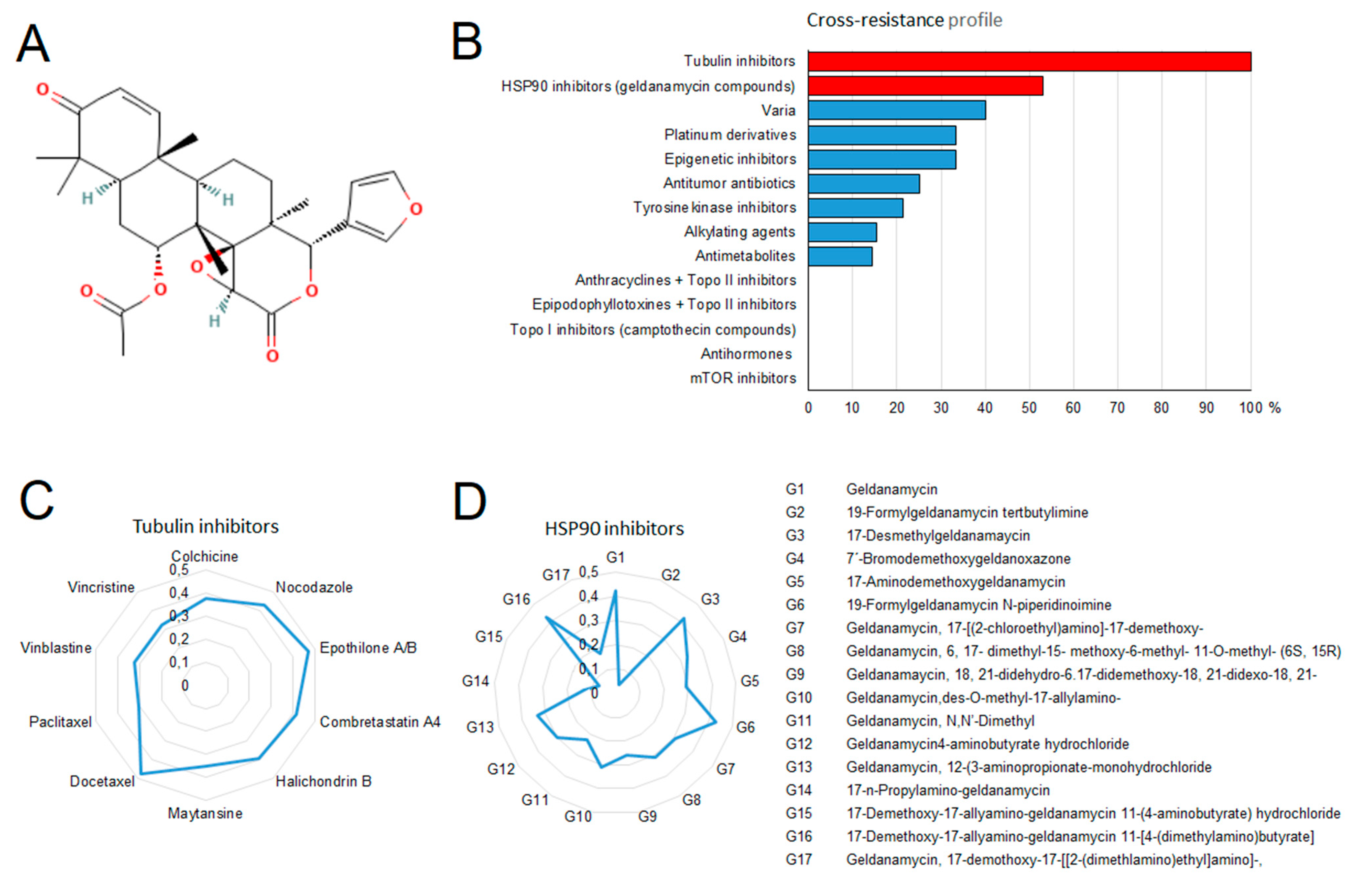
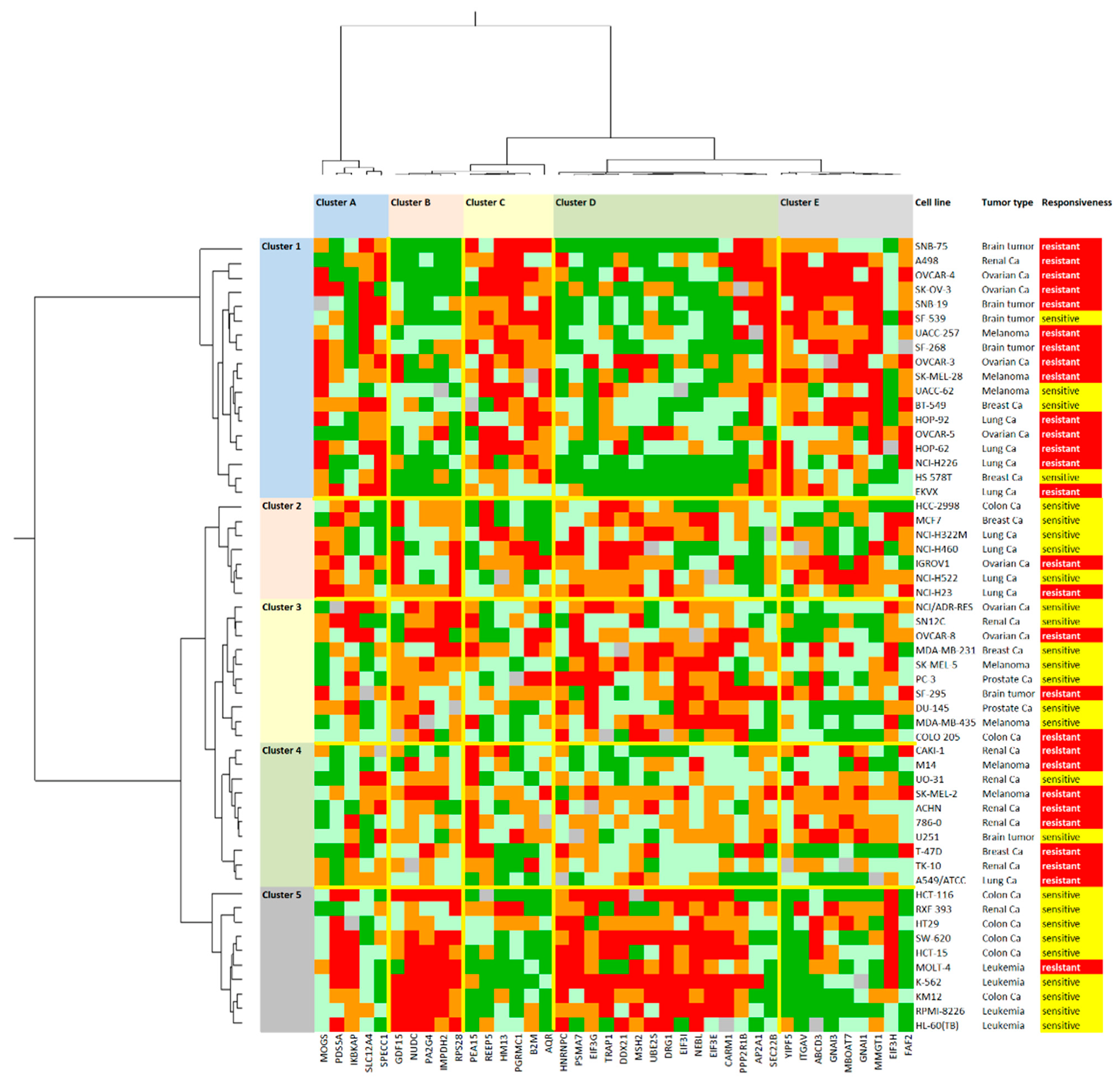
2.9. Drug Resistance Profiling of Gedunin
3. Discussion
4. Materials and Methods
4.1. Isolation of the Characterization of Gedunin
4.2. Cell Lines
4.3. Resazurin Assay
4.4. Agglomerative Cluster Analyses of Proteomic Expression Data
4.5. Confocal Microscopy of the Microtubule Network
4.6. Molecular Docking
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Devereaux, A.L.; Mercer, S.L.; Cunningham, C.W. DARK classics in chemical neuroscience: Morphine. ACS Chem. Neurosci. 2018, 9, 2395–2407. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO traditional medicine strategy 2014–2024. 2013. Available online: https://apps.who.int/iris/bitstream/handle/10665/92455/9789241506090_eng.pdf?sequence=1 (accessed on 13 July 2022).
- Tan, G.; Gyllenhaal, C.; Soejarto, D.D. Biodiversity as a source of anticancer drugs. Curr. Drug Targets 2006, 7, 265–277. [Google Scholar] [CrossRef] [PubMed]
- McChesney, J.D.; Venkataraman, S.K.; Henri, J.T. Plant natural products: Back to the future or into extinction? Phytochemistry 2007, 68, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Kinghorn, A.D.; Pan, L.; Fletcher, J.N.; Chai, H. The relevance of higher plants in lead compound discovery programs. J. Nat. Prod. 2011, 74, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T. Resistance to Targeted ABC Transporters in Cancer; Springer International Publishing: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Efferth, T.; Volm, M. Multiple resistance to carcinogens and xenobiotics: P-glycoproteins as universal detoxifiers. Arch. Toxicol. 2017, 91, 2515–2538. [Google Scholar] [CrossRef]
- Patwardhan, C.A.; Fauq, A.; Peterson, L.B.; Miller, C.; Blagg, B.S.; Chadli, A. Gedunin inactivates the co-chaperone p23 protein causing cancer cell death by apoptosis. J. Biol. Chem. 2013, 288, 7313–7325. [Google Scholar] [CrossRef]
- Campbell, M.J.; Hamilton, B.; Shoemaker, M.; Tagliaferri, M.; Cohen, I.; Tripathy, D. Antiproliferative activity of Chinese medicinal herbs on breast cancer cells in vitro. Anticancer Res. 2002, 22, 3843–3852. [Google Scholar]
- Efferth, T.; Kahl, S.; Paulus, K.; Adams, M.; Rauh, R.; Boechzelt, H.; Hao, X.; Kaina, B.; Bauer, R. Phytochemistry and pharmacogenomics of natural products derived from traditional Chinese medicine and Chinese materia medica with activity against tumor cells. Mol. Cancer Ther. 2008, 7, 152–161. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef]
- Braga, T.M.; Rocha, L.; Chung, T.Y.; Oliveira, R.F.; Pinho, C.; Oliveira, A.I.; Morgado, J.; Cruz, A. Biological activities of gedunin—A limonoid from the Meliaceae family. Molecules 2020, 25, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hieronymus, H.; Lamb, J.; Ross, K.N.; Peng, X.P.; Clement, C.; Rodina, A.; Nieto, M.; Du, J.; Stegmaier, K.; Raj, S.M.; et al. Gene expression signature-based chemical genomic prediction identifies a novel class of HSP90 pathway modulators. Cancer Cell 2006, 10, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Koch, E. Complex interactions between phytochemicals. The multi-target therapeutic concept of phytotherapy. Curr. Drug Targets 2011, 12, 122–132. [Google Scholar] [CrossRef]
- Selby, M.; Delosh, R.; Laudeman, J.; Ogle, C.; Reinhart, R.; Silvers, T.; Lawrence, S.; Kinders, R.; Parchment, R.; Teicher, B.A. 3D models of the NCI60 cell lines for screening oncology compounds. SLAS Discov. 2017, 22, 473–483. [Google Scholar] [CrossRef]
- Mofers, A.; Selvaraju, K.; Gubat, J.; D’Arcy, P.; Linder, S. Identification of proteasome inhibitors using analysis of gene expression profiles. Eur. J. Pharmacol. 2020, 889, 173709. [Google Scholar] [CrossRef] [PubMed]
- Sahai, R.; Bhattacharjee, A.; Shukla, V.N.; Yadav, P.; Hasanain, M.; Sarkar, J.; Narender, T.; Mitra, K. Gedunin isolated from the mangrove plant Xylocarpus granatum exerts its anti-proliferative activity in ovarian cancer cells through G2/M-phase arrest and oxidative stress-mediated intrinsic apoptosis. Apoptosis 2020, 25, 481–499. [Google Scholar] [CrossRef] [PubMed]
- Hientz, K.; Mohr, A.; Bhakta-Guha, D.; Efferth, T. The role of p53 in cancer drug resistance and targeted chemotherapy. Oncotarget 2017, 8, 8921–8946. [Google Scholar] [CrossRef]
- Marin, J.J.G.; Briz, O.; Herraez, E.; Lozano, E.; Asensio, M.; Di Giacomo, S.; Romero, M.R.; Osorio-Padilla, L.M.; Santos-Llamas, A.I.; Serrano, M.A.; et al. Molecular bases of the poor response of liver cancer to chemotherapy. Clin. Res. Hepatol. Gastroenterol. 2018, 42, 182–192. [Google Scholar] [CrossRef]
- Erin, N.; Grahovac, J.; Brozovic, A.; Efferth, T. Tumor microenvironment and epithelial mesenchymal transition as targets to overcome tumor multidrug resistance. Drug Resist. Updat. 2020, 53, 100715. [Google Scholar] [CrossRef]
- Scherf, U.; Ross, D.T.; Waltham, M.; Smith, L.H.; Lee, J.K.; Tanabe, L.; Kohn, K.W.; Reinhold, W.C.; Myers, T.G.; Andrews, D.T.; et al. A gene expression database for the molecular pharmacology of cancer. Nat. Genet. 2000, 24, 236–244. [Google Scholar] [CrossRef]
- Staunton, J.E.; Slonim, D.K.; Coller, H.A.; Tamayo, P.; Angelo, M.J.; Park, J.; Scherf, U.; Lee, J.K.; Reinhold, W.O.; Weinstein, J.N.; et al. Chemosensitivity prediction by transcriptional profiling. Proc. Natl. Acad. Sci. USA 2001, 98, 10787–10792. [Google Scholar] [CrossRef] [Green Version]
- Efferth, T.; Oesch, F. Oxidative stress response of tumor cells: Microarray-based comparison between artemisinins and anthracyclines. Biochem. Pharmacol. 2004, 68, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Özenver, N.; Dawood, M.; Fleischer, E.; Klinger, A.; Efferth, T. Chemometric and transcriptomic profiling, microtubule disruption and cell death induction by secalonic acid in tumor cells. Molecules 2020, 25, 3224. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.J.; Dawood, M.; Hult, A.K.; Olsson, M.L.; Efferth, T. Network pharmacology of triptolide in cancer cells: Implications for transcription factor binding. Investig. New Drugs 2021, 39, 1523–1537. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Kouvonen, P.; Koh, C.C.; Gillet, L.C.; Wolski, W.E.; Röst, H.L.; Rosenberger, G.; Collins, B.C.; Blum, L.C.; Gillessen, S.; et al. Rapid mass spectrometric conversion of tissue biopsy samples into permanent quantitative digital proteome maps. Nat. Med. 2015, 21, 407–413. [Google Scholar] [CrossRef]
- Efferth, T.; Grassmann, R. Impact of viral oncogenesis on responses to anti-cancer drugs and irradiation. Crit. Rev. Oncog. 2000, 11, 165–187. [Google Scholar]
- Panossian, A.; Efferth, T. Network pharmacology of adaptogens in the assessment of their pleiotropic therapeutic activity. Pharmaceuticals 2022, 15, 1051. [Google Scholar] [CrossRef]
- Volm, M.; Efferth, T. Prediction of cancer drug resistance and implications for personalized medicine. Front. Oncol. 2015, 5, 282. [Google Scholar] [CrossRef]
- Sarmento-Ribeiro, A.B.; Scorilas, A.; Gonçalves, A.C.; Efferth, T.; Trougakos, I.P. The emergence of drug resistance to targeted cancer therapies: Clinical evidence. Drug Resist. Updat. 2019, 47, 100646. [Google Scholar] [CrossRef]
- Cordon-Cardo, C.; O’Brien, J.P.; Boccia, J.; Casals, D.; Bertino, J.R.; Melamed, M.R. Expression of the multidrug resistance gene product (P-glycoprotein) in human normal and tumor tissues. J. Histochem. Cytochem. 1990, 38, 1277–1287. [Google Scholar] [CrossRef]
- Saeed, M.E.M.; Boulos, J.C.; Machel, K.; Andabili, N.; Marouni, T.; Roth, W.; Efferth, T. Expression of the stem cell marker ABCB5 in normal and tumor tissues. In Vivo 2022, 36, 1651–1666. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.A.; Duddeck, H.; Gonzalez-Sierra, M. Isolation and characterization of an antimalarial agent of the neem tree Azadirachata indica. J. Nat. Prod. 1989, 52, 922–927. [Google Scholar] [CrossRef]
- Kimmig, A.; Gekeler, V.; Neumann, M.; Frese, G.; Handgretinger, R.; Kardos, G.; Diddens, H.; Niethammer, D. Susceptibility of multidrug-resistant human leukemia cell lines to human interleukin 2-activated killer cells. Cancer Res. 1990, 50, 6793–6799. [Google Scholar]
- Gillet, J.P.; Efferth, T.; Steinbach, D.; Hamels, J.; de Longueville, F.; Bertholet, V.; Remacle, J. Microarray-based detection of multidrug resistance in human tumor cells by expression profiling of ATP-binding cassette transporter genes. Cancer Res. 2004, 64, 8987–8993. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Konkimalla, V.B.; Wang, Y.F.; Sauerbrey, A.; Meinhardt, S.; Zintl, F.; Mattern, J.; Volm, M. Prediction of broad-spectrum resistance of tumors towards anticancer drugs. Clin. Cancer Res. 2008, 14, 2405–2412. [Google Scholar] [CrossRef] [PubMed]
- Alley, M.C.; Scudiero, D.A.; Monks, A.; Hursey, M.L.; Czerwinski, M.J.; Fine, D.L.; Abbott, B.J.; Mayo, J.G.; Shoemaker, R.H.; Boyd, M.R. Feasibility of drug screening with panels of human tumor cell lines using a microculture tetrazolium assay. Cancer Res 1988, 48, 589–601. [Google Scholar] [PubMed]
- Adham, A.N.; Abdelfatah, S.; Naqishbandi, A.M.; Mahmoud, N.; Efferth, T. Cytotoxicity of apigenin toward multiple myeloma cell lines and suppression of iNOS and COX-2 expression in STAT1-transfected HEK293 cells. Phytomedicine 2021, 80, 153371. [Google Scholar] [CrossRef]
- Hegazy, M.F.; Dawood, M.; Mahmoud, N.; Elbadawi, M.; Sugimoto, Y.; Klauck, S.M.; Mohamed, N.; Efferth, T. 2α-Hydroxyalantolactone from Pulicaria undulata: Activity against multidrug-resistant tumor cells and modes of action. Phytomedicine 2021, 81, 153409. [Google Scholar] [CrossRef]
- Ferreira, L.; Hitchcock, D.B. A comparison of hierarchical methods for clustering functional data. Commun. Stat.-Simul. Comput. 2009, 38, 1925–1949. [Google Scholar] [CrossRef]
- Boulos, J.C.; Saeed, M.E.M.; Chatterjee, M.; Bülbül, Y.; Crudo, F.; Marko, D.; Munder, M.; Klauck, S.M.; Efferth, T. Repurposing of the ALK inhibitor crizotinib for acute leukemia and multiple myeloma cells. Pharmaceuticals 2021, 14, 1126. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Compound | Occurrence in Plants |
|---|---|---|
| SAK-T-01 | (-)-Scopolamine (hydrochloride) | Brugmansia spec., Datura spec., Hyoscyamus spec., Mandragora officinarum, Duboisia spec., Scopolia carniolica, Physochlaina orientalis |
| SAK-T-02 | (+)-Bicuculline | Dicentra cucullaria, Adlumia fungosa, Corydalis spec, Fumaria spec., Dicentra peregrina |
| SAK-T-03 | (+)-Laudanosine | Papaver somniferum, Glaucium flavum, Hazomalania voyronii |
| SAK-T-04 | (+)-Tubocurarine | Chondrodendron tomentosum |
| SAK-T-05 | (+)-Usnic acid | Ramalina fraxinea, Dimelaena oreina, Nephroma laevigatum, Xanthoparmelia tinctina, diverse species of the genera Usnea, Cladonia, Hypotrachyna, Lecanora, Ramalina, Evernia, Parmelia, and Alectoria |
| SAK-T-07 | 18-β-Glycyrrhetinic acid | Glycyrrhiza glabra |
| SAK-T-08 | 3,5-Dimethoxy-4-hydroxy cinnamic acid | Coffea spec., Prunus cerasus, Posidonia oceanica, Lithospermum erythrorhizon, Zea mays, Avena sativa, Triticum aestivum, Vitis vinifera |
| SAK-T-09 | Alloptaeroxylin | Spathelia sorbifolia |
| SAK-T-10 | Apigenin | Lawsonia inermis, Matricaria chamomilla, Verbascum lycnitis, Carex fraseriana, Vernonanthura nudiflora, Salvia spec., Dahlia spec., Apium spec. |
| SAK-T-11 | Aristolochic acid | Aristolochia spec., Asarum spec., Thottea duchartrei |
| SAK-T-12 | Barbaloin | Aloe spec., Rhamnus purshiana |
| SAK-T-13 | Berberine | Berberis vulgaris, Coptis chinensis, Hydrastis canadensis |
| SAK-T-14 | Boldine | Peumus boldis, Lindera spec., Licaria triandra, Neolitsea acuminatissima, Damburneya salicifolia, Actinodaphne acuminata |
| SAK-T-15 | Colchicine | Colchicum autumnale |
| SAK-T-16 | Cryptopine | Argemone mexicana, Fumara spec., Papaver spec., Arctomecon humilis |
| SAK-T-17 | Cumarin | Melilotus officinalis, Galium odoratum, Prunus mahaleb, Dipteryx odorata, Cinnamomum spec, Phoenix spec., and others |
| SAK-T-18 | Digitoxin | Digitalis purpurea |
| SAK-T-19 | Digoxin | Digitalis lanata |
| SAK-T-20 | Diosgenin | Dioscorea spec., Ophiopogon intermedius, Dracaena draco, Trigonella foenum graecum, Solanum spec., Asparagus officinalis, Allium cepa, Hellenia lacera |
| SAK-T-21 | Emetine (dihydrochloride) | Carapichea ipecacuanha, Hedera helix, Alangium longiflorum |
| SAK-T-22 | Emodin | Polygonum cuspidatum, Polygonum multiflorum, Rheum spec., Rhamnus spec. |
| SAK-T-23 | Emodin anthrone | Rhamnus prinoides, Paeonia emodi, Rumex acetosa |
| SAK-T-25 | Esculetin | Fraxinus spec., Artemisia eriopoda, Euphorbia decipiens, Phellodndron amurense, Salvia euphratica |
| SAK-T-26 | Esculin | Fraxinus spec., Artemisia eriopoda, Euphorbia decipiens, Phellodndron amurense, Salvia euphratica, Arabidopsis thaliana, Galinsoga quadriradiata, Cichorium pumilum |
| SAK-T-27 | Frangulin | Rhamnus spec. |
| SAK-T-28 | Fraxin | Aesculus hippocastanum, Fraxinus spec., Acer nikoense, Tetradium glabrifolium |
| SAK-T-29 | Gedunin | Azadirachta indica, Cedrela odorata, Melia azedarach, Toona sinensis, Xylocarpus granatum, Entandrophragma angolense, Carapa guianesis |
| SAK-T-30 | Gentibiose | Gentiana spec., Gentianopsis spec., diverse stone fruits |
| SAK-T-31 | Harmaline | Peganum harmala, Banisteriopsis caapi, Passiflora spec. |
| SAK-T-32 | Harman | Vestia foetida, Banisteriopsis caapi, Strychnos johnsonii, Ophiopogon spec., Carex brevicollis, Vitis vinifera |
| SAK-T-33 | Harmine | Banisteriopsis caapi, Thalictrum foetidum, Peganum harmala, Passiflora incarnata, Festuca spec. |
| SAK-T-34 | Hesperetin | Citrus peels, Brassica oleracea, Schizonepeta tenuifolia, Salvia spec., Arabidopsis thaliana, Rubus idaeus |
| SAK-T-35 | Hesperidin | Citrus peels, Ficus erecta var. beecheyana, Zanthoxylum caribaeum, Betula pendula |
| SAK-T-36 | Heteropeucenin | Cedrelopsis grevei |
| SAK-T-37 | Khellin | Ammi visnaga, Annona muricata, Allium nutans, Dioscorea spec. |
| SAK-T-38 | Lawsone | Lawsonia inermis, Eichhornia crassipes |
| SAK-T-39 | Lupeol | Craveta nurvala, Betula spec., Ficus septica, Diospyros morrisiana, Paeonia emodi, Mikania haenkeana, Symphoricarpos albus, Avicennia officinalis, Derris trifoliata |
| SAK-T-40 | Morin | Morus tinctoria, Lotus ucrainicus, Psidium guajava, Petasites formosanus, Tilia tomentosa, Maclura tricupidata, Plantago lanceolata, Endosamara racemosa |
| SAK-T-41 | Narceine | Papaver somniferum |
| SAK-T-42 | Narcotine | Papaver spec. |
| SAK-T-43 | Oleandrin | Nerium oleander |
| SAK-T-44 | Ouabain | Acocanthera schimperi, Acocanthera oppositifolia, Strophantus gratus, Cunila spec. |
| SAK-T-45 | Papaverine | Papaver spec., Papaver rhoeas, Sauropus androgynus |
| SAK-T-46 | Peucenin-(5.7-dihydroxy-6-isopentyl-2-methylchromone | Peucedanum ostruthium |
| SAK-T-47 | Physostigmine | Datura stramonium, Physostigma venenosum, Hippomane mancinella |
| SAK-T-48 | Pilocarpine | Pilocarpus spec., |
| SAK-T-49 | Piperine | Macropiper spec., Piper spec. |
| SAK-T-50 | Psoralen | Ficu carica, Psoralea corylifolia, Citrus × limon, Angelica spec., Dianthus spec., Ammi visnaga, Pastinaca sativa, Petoselinum crispum, Levisticum officinale, Foeniculum vulgare, Daucus carota, Apium graveolens |
| SAK-T-51 | Ptaeroxylin | Cedrelopsis gravei |
| SAK-T-52 | Quercetin (dihydrate) | Quercetin is ubiquitous in vegetarian food |
| SAK-T-53 | Quinidine | Stereoisomer of quinine from Cinchona officinalis |
| SAK-T-54 | Quinine (sulphate) | Quinine is from Cinchona officinalis |
| SAK-T-55 | Rotenone | Lonchocarpus nicou, Derris elliptica, Deguelia utilis |
| SAK-T-58 | Rutin | Viola tricolor, Styphnolobium japonicum, Fagopyrum esculentum, Morus alba, Sambucus nigra subsp. canadensis, Petroselinum crispum, Persicaria hydropiper, Hypericum perforatum, Ficus virens, Visnea mocanera, Heliopsis helianthoides var. scabra, Ferulaga sylvatica, Nerium oleander, Polygonum cognatum |
| SAK-T-59 | Sanguinarine | Sanguinaria canadensis, Argemone spec., Chelidonium majus, Macleaya cordata, Glaucium flavum, Fumaria spec., Eschscholzia california |
| SAK-T-60 | Santonin | Artemisia spec., Fossombronia wondraczekii |
| SAK-T-61 | Scopoletin | Scopolia spec., Artemisia spec., Viburnum prunifolium, Solanum nigrum, Urticaria dioica, Brunfelsia americana, Passiflora spec., Datura metel, Mallotus resinosus, Kleinhovia hospita, Trogonella foenum graecum, Taraxacum officinale, Ficus auriculata, Haplophyllum cappadocicum, Lessingianthus mollissimus, Zantoxylum ailanthoides, Tetradium glabrifolium, Phellodendron amurense |
| SAK-T-62 | Silibinin | Silybum marianum, Anastatica hierochuntica, Silybum eburneum |
| SAK-T-63 | Sinigrin (monohydrate) | Sinigrin from Cakile arabica, Erucaria cakiloidea, Brassica spec., Raphanus spec., Hirschfeldia incana |
| SAK-T-64 | Stigmasterol | Physostigma venenosum, Glycine max, Brassica napus, Hebanthe eriantha, Xylopia aromatica, Citrus spec., Ophiopogon japonicus, Mirabilis jalapa |
| SAK-T-65 | Strophanthidin | Strophantus spec., Convallaria majalis, Adonis spec., Apocrynum cannabinum, Descurainia sophia, Antiaris toxicaria, Cryptolepis migriscens, Erysimum inconspicuum |
| SAK-T-66 | Strychnine | Strychnos nux-vomica, Ignatia amara |
| SAK-T-67 | Thebaine alkaloid | Thebaine from Papaver bracteatum |
| SAK-T-68 | Ursolic acid | Rosmarinus officinalis, Thymus vulgaris, Mirabilis jalapa, Malus domestica, Ocimum spec., Vaccinium spec., Sambucus spec., Crataegus spec., Lavandula spica, Origanum vulgare, Mentha × piperita, Prunus domestica, Pimpinella major, Gladiolus italicus, Symphoricarpus albus, Derri trifoliata, Rhizophora mucronata, Avicennia spec., Thymus spec., Nepeta cataria, Eucalyptus gradis, Psidium guajava, Prunella vulgaris, Nerium oleander, Ilex paraguariensis |
| SAK-T-69 | Vincamine | Vinca spec. |
| SAK-T-70 | Yohimbine (hydrochloride) | Corynanthe (Pausinystalia) johimbe, Rauvolfia spec., Aspidosperma quebracho-blanco, Tabernaemontana corymbosa, Alstonia angustifolia, Pouteria spec. |
| SAK-T-71 | β-Escin | Aesculum hippocastanum, Bobgunnia madagascariensis |
| SAK-T-72 | P-Arbutin | Viburnum opulus, Bergenia crassifolia, Schisandra chinensis, Grevillea robusta, Halocarpus biformis, Arabidopsis thaliana, Rhodolia spec., Vitis vinifera, Eriosema tuberosum |
| Phytochemical | Acute Toxicity * | Chronic Toxicity |
|---|---|---|
| Colchicine | Cardiotoxicity, hepatotoxicity, nephrotoxicity, gastrointestinal toxicity, lung toxicity, behavioral toxicity | None |
| Digitoxin | Cardiotoxicity, neurotoxicity, hematotoxicity, gastrointestinal toxicity, behavioral toxicity | None |
| Digoxin | Cardiotoxicity, hepatotoxicity, gastrointestinal toxicity, behavioral toxicity, lung toxicity | Carcinogenesis |
| Emetine | Cardiotoxicity, gastrointestinal toxicity, muscle weakness, dermal toxicity | Mutagenesis |
| Gedunin | Dermal toxicity (irritant) | None |
| Rotenone | Cardiotoxicity, gastrointestinal toxicity, lung toxicity, behavioral toxicity, dermal toxicity, neurotoxicity | None |
| Sanguinarine | Hepatotoxicity, behavioral toxicity | Teratogenesis, carcinogenesis |
| Strophantidine | Cardiotoxicity, lung toxicity, behavioral toxicity | Cardiotoxicity |
| Tubulin Binding Sites | Compounds | Lowest Binding Energy (Kcal/mol) | pKi (µM) | Amino Acids Involved in Ligand Interaction |
|---|---|---|---|---|
| Vinca alkaloid binding site | Vinorelbine | −10.76 ± 0.35 | 0.02 ± 0.01 | THR210, PHE214, PRO222, TYR224. |
| Gedunin | −8.89 ± 0.00 | 0.30 ± 0.00 | GLN11, THR145, ASN206. | |
| Taxane binding site | Paclitaxel | −10.41 ± 0.66 | 0.07 ± 0.04 | LEU217, HIS229, PHE272, LEU275, THR276, GLN281, LEU371. |
| Gedunin | −7.48 ± 0.00 | 3.27 ± 0.01 | LEU217, LEU230, LEU275, LEU371, LYS372. | |
| Colchicine binding site | Colchicine | −7.57 ± 0.04 | 2.82 ± 0.19 | ASP69, ALA180, LEU248, LYS254. |
| Gedunin | −4.49 ± 0.03 | 514.99 ± 2.92 | ALA100, ASN101, THR179, GLU183, TYR224, LYS254. |
| Clusters 2,3,5 | Clusters 1,4 | |||
|---|---|---|---|---|
| Partition (log10IC50) | <−5.125 M | ≥−5.125 M | ||
| Sensitive | 17 | 6 | ||
| Resistant | 10 | 22 | ||
| χ2 test: p = 0.002 | ||||
| Sensitive | Resistant | Suitability for therapy | ||
| Brain tumor | 2 | 4 | no | |
| Renal Ca | 3 | 5 | no | |
| Ovarian Ca | 1 | 6 | no | |
| Melanoma | 3 | 4 | no | |
| Breast Ca | 4 | 1 | yes | |
| Lung Ca | 3 | 6 | no | |
| Colon Ca | 6 | 1 | yes | |
| Prostate Ca | 2 | 0 | yes | |
| Leukemia | 3 | 1 | yes | |
| χ2 test: p = 0.076 | ||||
| Gedunine | Control Drug | ||
|---|---|---|---|
| (log10 IC50, M) | (log10 IC50, M) | ||
| ABCB1 Expression | Epirubicin | ||
| 7q21 (Chromosomal | r-value | −0.078 | * 0.447 |
| locus of ABCB1 gene) | p-value | 0.297 | * 3.55 × 10−4 |
| ABCB1 expression | r-value | −0.097 | * 0.533 |
| (microarray) | p-value | 0.243 | * 6.82 × 10−6 |
| ABCB1 expression | r-value | −0.143 | * 0.410 |
| (RT-PCR) | p-value | 0.168 | * 1.54 × 10−3 |
| Rhodamine 123 | r-value | −0.115 | * 0.526 |
| accumulation | p-value | 0.207 | * 1.12 × 10−5 |
| ABCB5 Expression | Maytansine | ||
| ABCB5 expression | r-value | −0.072 | * 0.454 |
| (microarray) | p-value | 0.301 | * 6.67 × 10−4 |
| ABCB5 expression | r-value | 0.036 | * 0.402 |
| (RT-PCR) | p-value | 0.396 | * 0.0026 |
| ABCC1 Expression | Vinblastine | ||
| DNA gene | r-value | −0.072 | * 0.429 |
| copy number | p-value | 0.301 | * 0.001 |
| ABCC1 expression | r-value | 0.051 | * 0.399 |
| (microarray) | p-value | 0.357 | * 0.003 |
| ABCC1 expression | r-value | 0.061 | 0.299 |
| (RT-PCR) | p-value | 0.346 | * 0.036 |
| ABCG2 Expression | Pancratistatin | ||
| ABCG2 expression | r-value | −0.178 | * 0.329 |
| (microarray) | p-value | 0.099 | * 0.006 |
| ABCG2 expression | r-value | −0.222 | * 0.346 |
| (western blot) | p-value | 0.052 | * 0.004 |
| EGFR Expression | Erlotinib | ||
| EGFR gene | r-value | 0.172 | −0.245 |
| Copy number | p-value | 0.105 | * 0.029 |
| EGFR expression | r-value | 0.229 | * −0.458 |
| (microarray) | p-value | * 0.046 | * 1.15 × 10−4 |
| EGFR expression | r-value | * 0.346 | * −0.379 |
| (PCR slot blot) | p-value | * 0.005 | * 0.002 |
| EGFR expression | r-value | 0.156 | * −0.376 |
| (protein array) | p-value | 0.128 | * 0.001 |
| TP53 Mutation | 5-Fluorouracil | ||
| TP53 mutation | r-value | −0.093 | * −0.502 |
| (cDNA sequencing) | p-value | 0.253 | * 3.50 × 10−5 |
| TP53 function | r-value | −0.115 | * −0.436 |
| (yeast functional assay) | p-value | 0.213 | * 5.49 × 10−4 |
| WT1 Expression | Ifosfamide | ||
| WT1 expression | r-value | −0.155 | * −0.316 |
| (microarray) | p-value | 0.129 | * 0.007 |
| GSTP1 Expression | Etoposide | ||
| GSTP1 expression | r-value | 0.009 | 0.399 |
| (microarray) | p-value | 0.473 | * 9.58 × 10−4 |
| GSTP1 expression | r-value | 0.123 | 0.509 |
| (northern blot) | p-value | 0.185 | * 2.24 × 10−5 |
| HSP90 | Geldanamycin | ||
| HSP90 Expression | r-value | * −0.426 | * −0.392 |
| (microarray) | p-value | * 5.92×10−4 | * 0.001 |
| Proliferation | |||
| Cell doubling | r-value | * 0.456 | * 0.627 |
| p-value | * 2.67 × 10−4 | * 7–14 × 10−6 | |
| N-/K-/H-RAS Mutations | Melphalan | ||
| TP53 mutation | r-value | 0.275 | * 0.367 |
| (cDNA sequencing) | p-value | 0.021 | * 0002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalid, S.A.; Dawood, M.; Boulos, J.C.; Wasfi, M.; Drif, A.; Bahramimehr, F.; Shahhamzehei, N.; Shan, L.; Efferth, T. Identification of Gedunin from a Phytochemical Depository as a Novel Multidrug Resistance-Bypassing Tubulin Inhibitor of Cancer Cells. Molecules 2022, 27, 5858. https://doi.org/10.3390/molecules27185858
Khalid SA, Dawood M, Boulos JC, Wasfi M, Drif A, Bahramimehr F, Shahhamzehei N, Shan L, Efferth T. Identification of Gedunin from a Phytochemical Depository as a Novel Multidrug Resistance-Bypassing Tubulin Inhibitor of Cancer Cells. Molecules. 2022; 27(18):5858. https://doi.org/10.3390/molecules27185858
Chicago/Turabian StyleKhalid, Sami A., Mona Dawood, Joelle C. Boulos, Monica Wasfi, Assia Drif, Faranak Bahramimehr, Nasim Shahhamzehei, Letian Shan, and Thomas Efferth. 2022. "Identification of Gedunin from a Phytochemical Depository as a Novel Multidrug Resistance-Bypassing Tubulin Inhibitor of Cancer Cells" Molecules 27, no. 18: 5858. https://doi.org/10.3390/molecules27185858
APA StyleKhalid, S. A., Dawood, M., Boulos, J. C., Wasfi, M., Drif, A., Bahramimehr, F., Shahhamzehei, N., Shan, L., & Efferth, T. (2022). Identification of Gedunin from a Phytochemical Depository as a Novel Multidrug Resistance-Bypassing Tubulin Inhibitor of Cancer Cells. Molecules, 27(18), 5858. https://doi.org/10.3390/molecules27185858