
Functional Characterization of an Arabidopsis Profilin Protein as a Molecular Chaperone under Heat Shock Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
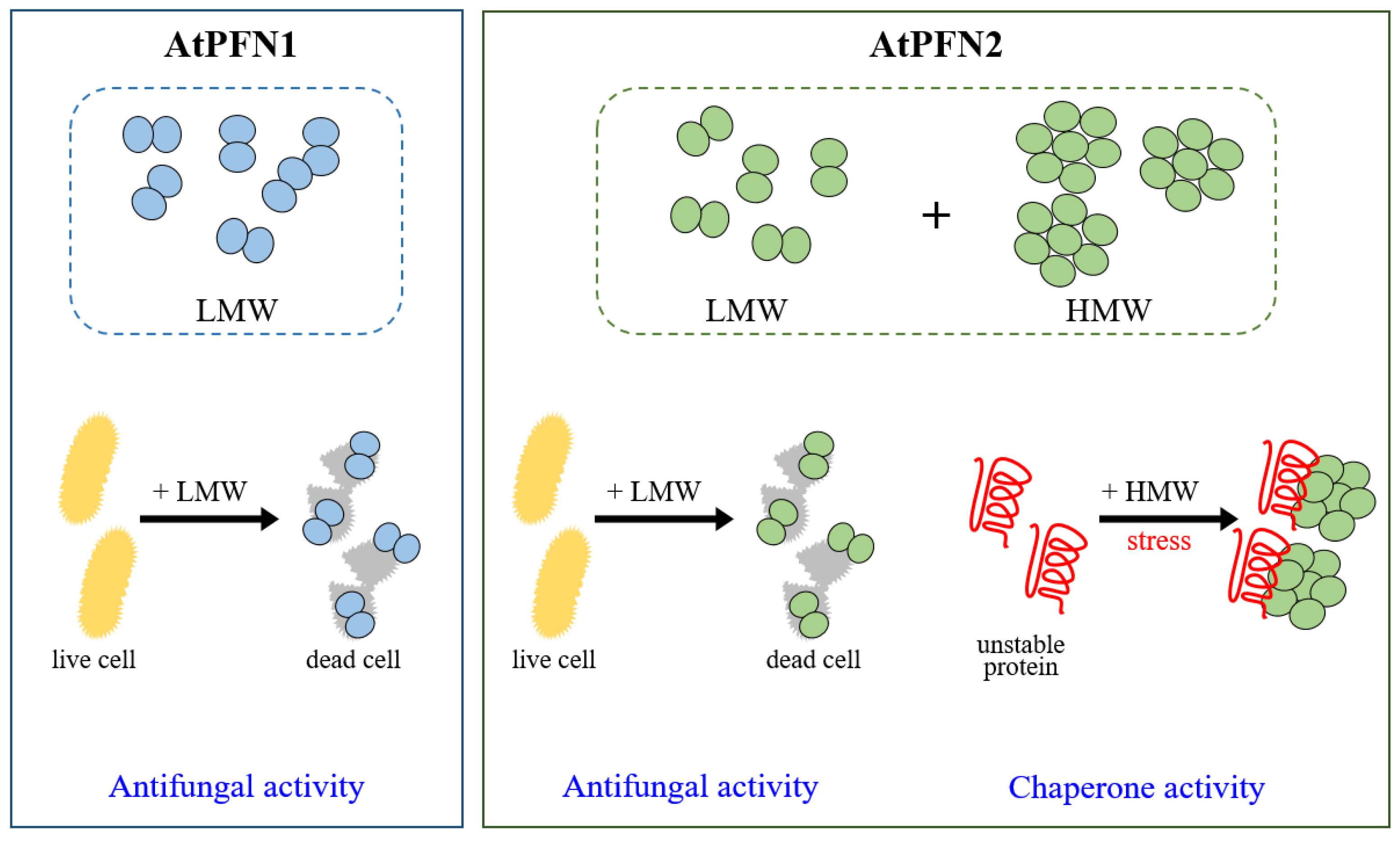
2. Results and Discussion
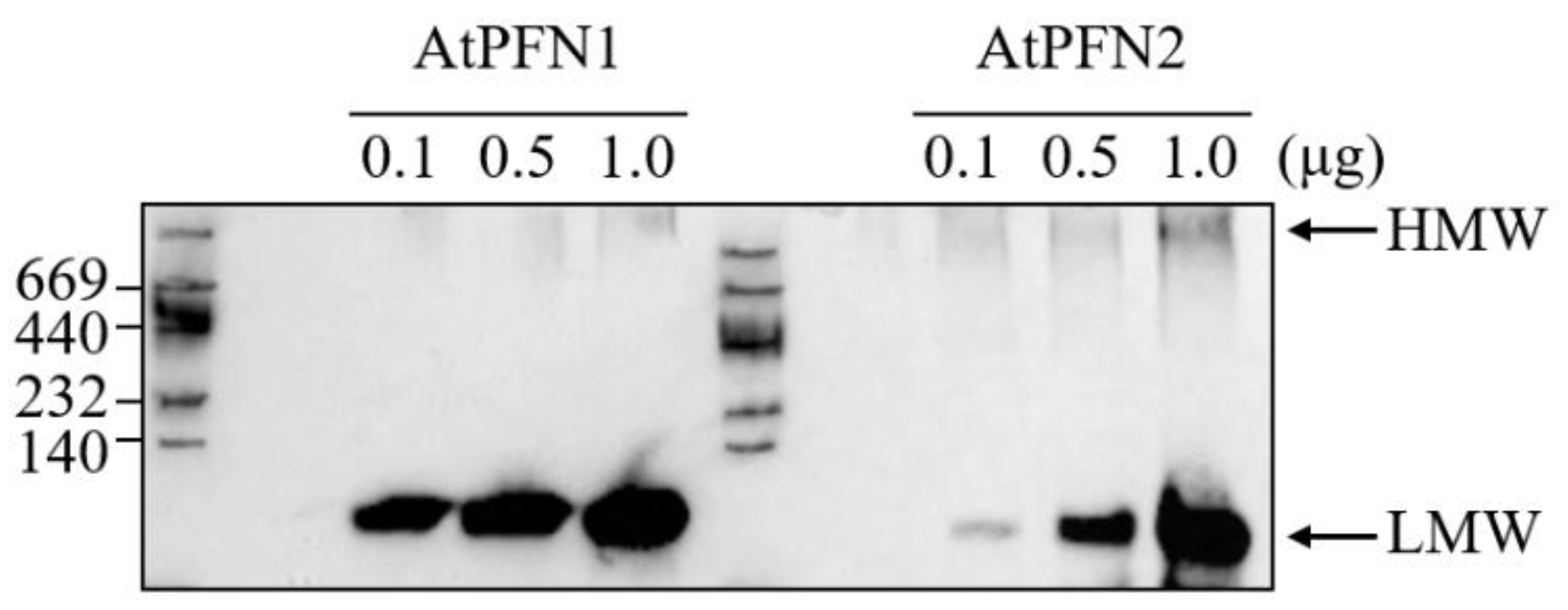
2.1. Stability of Arabidopsis Profilin Proteins in Plant Defense during External Heat Shock
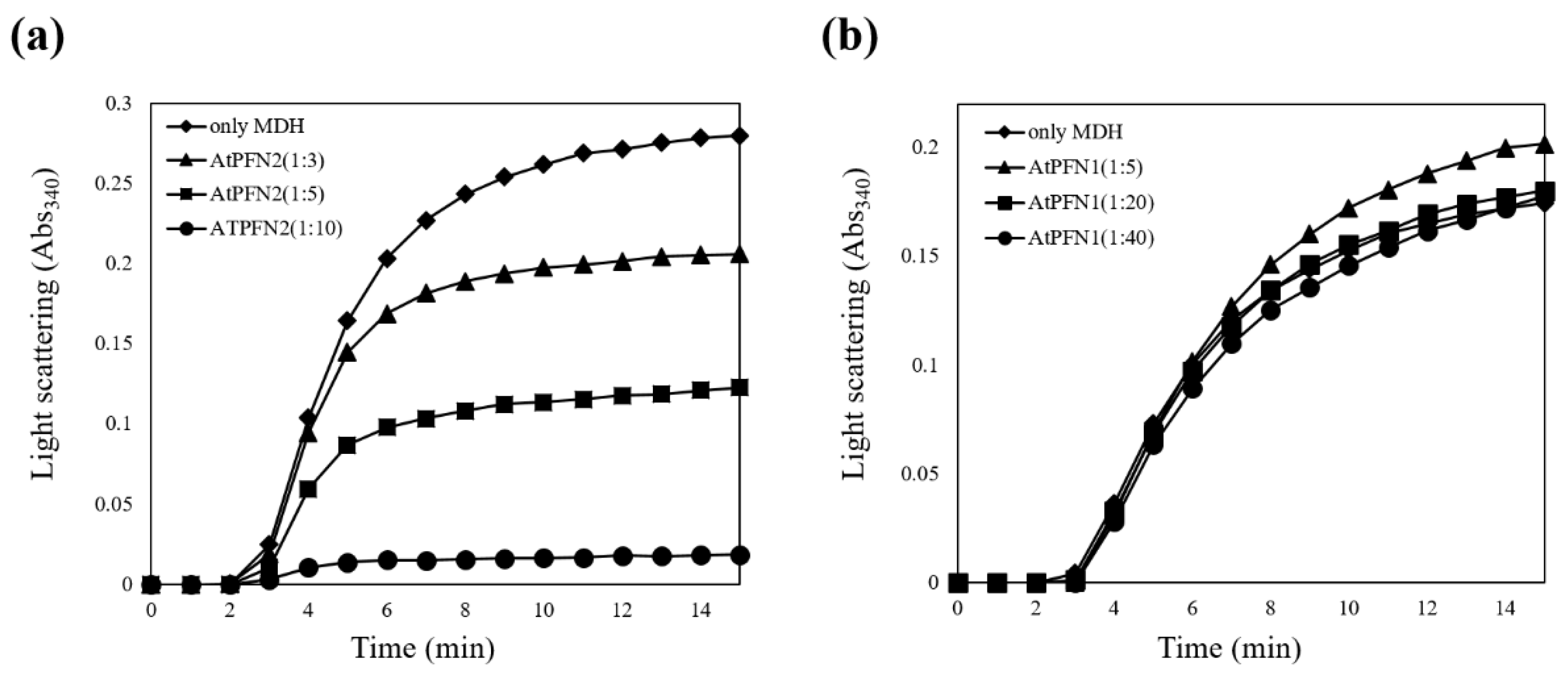
2.2. Molecular Chaperone Function of AtPFN Proteins
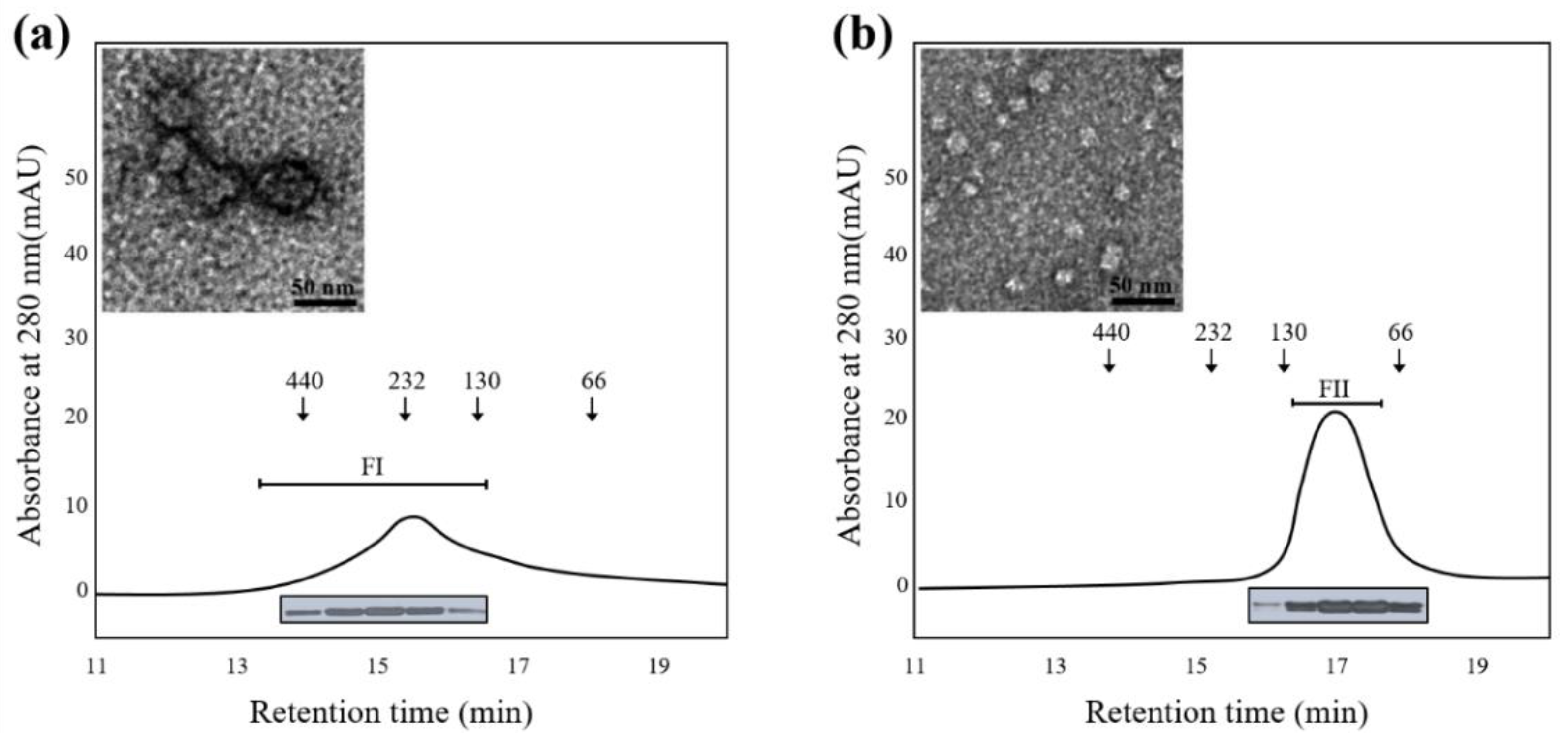
2.3. The Structural Difference between Profilin Isoforms
3. Materials and Methods
3.1. Materials
3.2. Purification and Structural Analysis of the AtPFN Proteins
3.3. Enzymatic Analyses of Molecular Chaperone and Antifungal Activity
3.4. Hydrophobicity Analysis of AtPFNs Using bis-ANS Fluorescence
3.5. Size-Exclusion Chromatography and Polyacrylamide Gel Electrophoresis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bendix, C.; Lewis, J.D. The enemy within: Phloem-limited pathogens. Mol. Plant Pathol. 2018, 19, 238–254. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Celma, J.; Ceballos-Laita, L.; Grusak, M.A.; Abadía, J.; López-Millán, A.-F. Plant fluid proteomics: Delving into the xylem sap, phloem sap, and apoplastic fluid proteomes. BBA-Proteins Proteom. 2016, 1864, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Hipper, C.; Brault, V.; Ziegler-Graff, V.; Revers, F. Viral and Cellular Factors Involved in Phloem Transport of Plant Viruses. Front. Plant Sci. 2013, 4, 154. [Google Scholar] [CrossRef] [PubMed]
- Kappagantu, M.; Collum, T.D.; Dardick, C.; Culver, J.N. Viral Hacks of the Plant Vasculature: The Role of Phloem Alterations in Systemic Virus Infection. Annu. Rev. Virol. 2020, 7, 351–370. [Google Scholar] [CrossRef] [PubMed]
- Turgeon, R.; Wolf, S. Phloem transport: Cellular pathways and molecular trafficking. Ann. Rev. Plant Biol. 2009, 60, 207–221. [Google Scholar] [CrossRef]
- Tetyuk, O.; Benning, U.F.; Hoffmann-Benning, S. Collection and analysis of Arabidopsis phloem exudates using the EDTA-facilitated method. J. Vis. Exp. 2013, 80, e51111. [Google Scholar] [CrossRef]
- Helbig, K.J.; Beard, M.R. The role of viperin in the innate antiviral response. J. Mol. Biol. 2014, 426, 1210–1219. [Google Scholar] [CrossRef]
- Sánchez-Sánchez, H.; Morquecho-Contreras, A. Chemical plant defense against herbivores. In Herbivores; Shields, V.D.C., Ed.; IntechOpen: London, UK, 2017. [Google Scholar]
- Narváez-Vásquez, J.; Pearce, G.; Ryan, C.A. The plant cell wall matrix harbors a precursor of defense signaling peptides. Proc. Natl. Acad. Sci. USA 2005, 102, 12974–12977. [Google Scholar] [CrossRef]
- Mott, G.A.; Middleton, M.A.; Desveaux, D.; Guttman, D.S. Peptides and small molecules of the plant-pathogen apoplastic arena. Front. Plant Sci. 2014, 5, 677. [Google Scholar] [CrossRef]
- Castro, M.S.; Fontes, W. Plant defense and antimicrobial peptides. Protein Pept. Lett. 2005, 12, 11–16. [Google Scholar] [CrossRef]
- Carlsson, L.; Nyström, L.E.; Sundkvist, I.; Markey, F.; Lindberg, U. Actin polymerizability is influenced by profilin, a low-molecular-weight protein in non-muscle cells. J. Mol. Biol. 1977, 115, 465–483. [Google Scholar] [CrossRef]
- Theriot, J.A.; Mitchison, T.J. The three faces of profilin. Cell 1993, 75, 835–838. [Google Scholar] [CrossRef]
- Jockusch, B.; Murk, K.; Rothkegel, M. The profile of profilins. Rev. Physiol. Biochem. Pharmacol. 2007, 159, 131–149. [Google Scholar] [PubMed]
- Kiyota, K.; Yoshimitsu, M.; Satsuki-Murakami, T.; Akutsu, K.; Kajimura, K.; Yamano, T. Detection of the tomato allergen Sola l 1 and evaluation of its reactivity after heat and papain treatment. Food Agric. Immunol. 2017, 28, 1450–1459. [Google Scholar] [CrossRef]
- Baluška, F.; Salaj, J.; Mathur, J.; Braun, M.; Jasper, F.; Šamaj, J.; Chua, N.-H.; Barlow, P.W.; Volkmann, D. Root hair formation: F-actin-dependent tip growth is initiated by local assembly of profilin-supported F-actin meshworks accumulated within expansin-enriched bulges. Dev. Biol. 2000, 227, 618–632. [Google Scholar] [CrossRef]
- Finkel, T.; Theriot, J.A.; Dise, K.R.; Tomaselli, G.F.; Goldschmidt-Clermont, P.J. Dynamic actin structures stabilized by profilin. Proc. Natl. Acad. Sci. USA 1994, 91, 1510–1514. [Google Scholar] [CrossRef]
- Christensen, H.E.; Ramachandran, S.; Tan, C.T.; Surana, U.; Dong, C.H.; Chua, N.H. Arabidopsis profilins are functionally similar to yeast profilins: Identification of a vascular bundle-specific profilin and a pollen-specific profilin. Plant J. 1996, 10, 269–279. [Google Scholar] [CrossRef]
- Huang, S.; McDowell, J.; Weise, M.J.; Meagher, R.B. The Arabidopsis profilin gene family (Evidence for an ancient split between constitutive and pollen-specific profilin genes). Plant Physiol. 1996, 111, 115–126. [Google Scholar] [CrossRef]
- Park, S.-C.; Kim, I.R.; Kim, J.-Y.; Lee, Y.; Kim, E.-J.; Jung, J.H.; Jung, Y.J.; Jang, M.-K.; Lee, J.R. Molecular mechanism of Arabidopsis thaliana profilins as antifungal proteins. BBA-Gen. Subj. 2018, 1862, 2545–2554. [Google Scholar] [CrossRef]
- Krishnan, K.; Moens, P.D. Structure and functions of profilins. Biophys. Rev. 2009, 1, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Babich, M.; Foti, L.R.; Sykaluk, L.L.; Clark, C.R. Profilin forms tetramers that bind to G-actin. Biochem. Biophys. Res. Commun. 1996, 218, 125–131. [Google Scholar] [CrossRef]
- Wopfner, N.; Willeroider, M.; Hebenstreit, D.; Van Ree, R.; Aalbers, M.; Briza, P.; Thalhamer, J.; Ebner, C.; Richter, K.; Ferreira, F. Molecular and immunological characterization of profilin from mugwort pollen. Biol. Chem. 2002, 383, 1779–1789. [Google Scholar] [CrossRef] [PubMed]
- Schmidt von Braun, S.; Schleiff, E. The chloroplast outer membrane protein CHUP1 interacts with actin and profilin. Planta 2008, 227, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Jing, Y.; Wang, Z.; Mao, T.; Šamaj, J.; Yuan, M.; Ren, H. Arabidopsis profilin isoforms, PRF1 and PRF2 show distinctive binding activities and subcellular distributions. J. Integr. Plant Biol. 2009, 51, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Grenklo, S.; Geese, M.; Lindberg, U.; Wehland, J.; Karlsson, R.; Sechi, A.S. A crucial role for profilin–actin in the intracellular motility of Listeria monocytogenes. EMBO Rep. 2003, 4, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Barazorda-Ccahuana, H.L.; Theiss-De-Rosso, V.; Valencia, D.E.; Gómez, B. Heat-stable hazelnut profilin: Molecular dynamics simulations and immunoinformatics analysis. Polymers 2020, 12, 1742. [Google Scholar] [CrossRef]
- Jung, Y.J.; Melencion, S.M.B.; Lee, E.S.; Park, J.H.; Alinapon, C.V.; Oh, H.T.; Yun, D.-J.; Chi, Y.H.; Lee, S.Y. Universal stress protein exhibits a redox-dependent chaperone function in Arabidopsis and enhances plant tolerance to heat shock and oxidative stress. Front. Plant Sci. 2015, 6, 1141. [Google Scholar] [CrossRef]
- Jung, Y.J.; Chi, Y.H.; Chae, H.B.; Shin, M.R.; Lee, E.S.; Cha, J.-Y.; Paeng, S.K.; Lee, Y.; Park, J.H.; Kim, W.Y.; et al. Analysis of Arabidopsis thioredoxin-H isotypes identifies discrete domains that confer specific structural and functional properties. Biochem. J. 2013, 456, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Jung, Y.J.; Lee, J.R.; Lee, Y.M.; Jang, H.H.; Lee, S.S.; Park, J.H.; Kim, S.Y.; Moon, J.C.; Lee, S.Y.; et al. Heat-shock and redox-dependent functional switching of an h-type Arabidopsis thioredoxin from a disulfide reductase to a molecular chaperone. Plant Physiol. 2009, 150, 552–561. [Google Scholar] [CrossRef]
- Lee, J.R.; Lee, S.S.; Jang, H.H.; Lee, Y.M.; Park, J.H.; Park, S.-C.; Moon, J.C.; Park, S.K.; Kim, S.Y.; Lee, S.Y.; et al. Heat-shock dependent oligomeric status alters the function of a plant-specific thioredoxin-like protein, AtTDX. Proc. Natl. Acad. Sci. USA 2009, 106, 5978–5983. [Google Scholar] [CrossRef] [Green Version]
- Haley, D.A.; Horwitz, J.; Stewart, P.L. The small heat-shock protein, αB-crystallin, has a variable quaternary structure. J. Mol. Biol. 1998, 277, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Reddy, G.B.; Reddy, P.Y.; Suryanarayana, P. αA-and αB-crystallins protect glucose-6-phosphate dehydrogenase against UVB irradiation-induced inactivation. Biochem. Biophys. Res. Commun. 2001, 282, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.K.; Kaur, H.; Kumar, G.S.; Kester, K. Interaction of 1,1′-bi (4-anilino) naphthalene-5,5′-disulfonic acid with α-crystallin. J. Biol. Chem. 1998, 273, 8965–8970. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, H.; Jung, Y.J.; Park, S.-C.; Kim, I.R.; Park, J.H.; Jang, M.-K.; Lee, J.R. Functional Characterization of an Arabidopsis Profilin Protein as a Molecular Chaperone under Heat Shock Stress. Molecules 2022, 27, 5771. https://doi.org/10.3390/molecules27185771
Son H, Jung YJ, Park S-C, Kim IR, Park JH, Jang M-K, Lee JR. Functional Characterization of an Arabidopsis Profilin Protein as a Molecular Chaperone under Heat Shock Stress. Molecules. 2022; 27(18):5771. https://doi.org/10.3390/molecules27185771
Chicago/Turabian StyleSon, Hyosuk, Young Jun Jung, Seong-Cheol Park, Il Ryong Kim, Joung Hun Park, Mi-Kyeong Jang, and Jung Ro Lee. 2022. "Functional Characterization of an Arabidopsis Profilin Protein as a Molecular Chaperone under Heat Shock Stress" Molecules 27, no. 18: 5771. https://doi.org/10.3390/molecules27185771
APA StyleSon, H., Jung, Y. J., Park, S.-C., Kim, I. R., Park, J. H., Jang, M.-K., & Lee, J. R. (2022). Functional Characterization of an Arabidopsis Profilin Protein as a Molecular Chaperone under Heat Shock Stress. Molecules, 27(18), 5771. https://doi.org/10.3390/molecules27185771