
Degradative Effect of Nattokinase on Spike Protein of SARS-CoV-2

 , and
, and 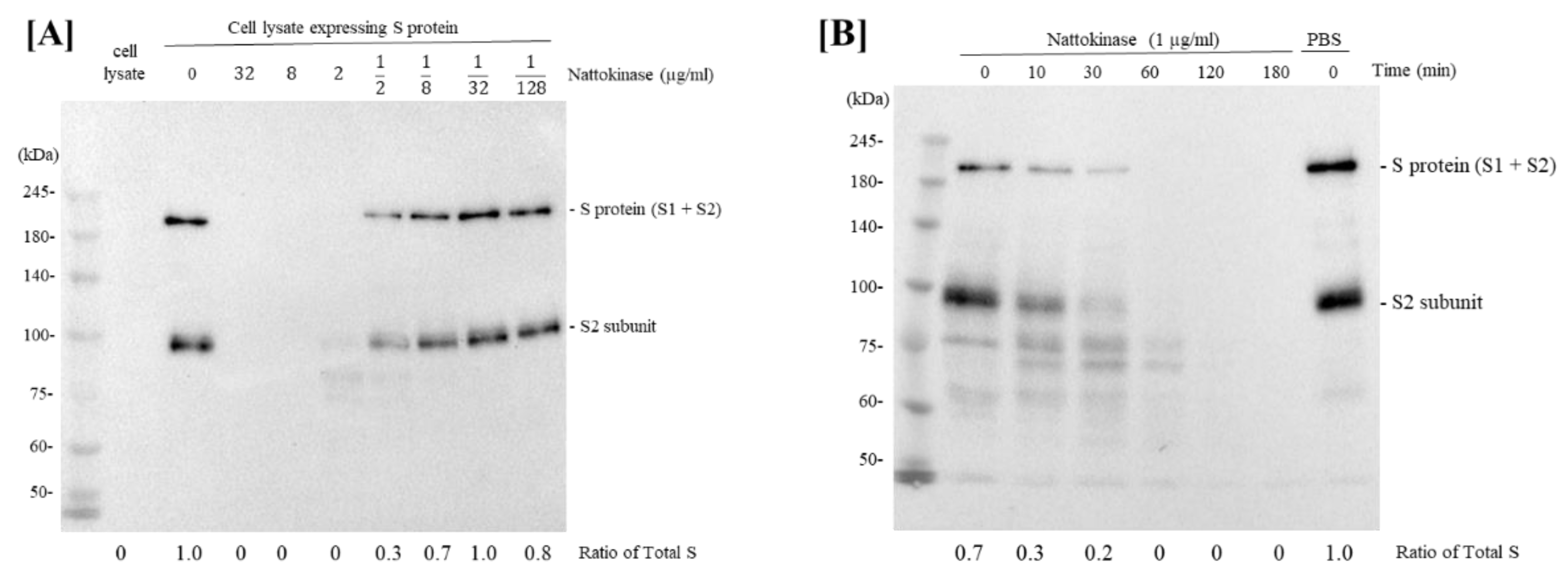{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
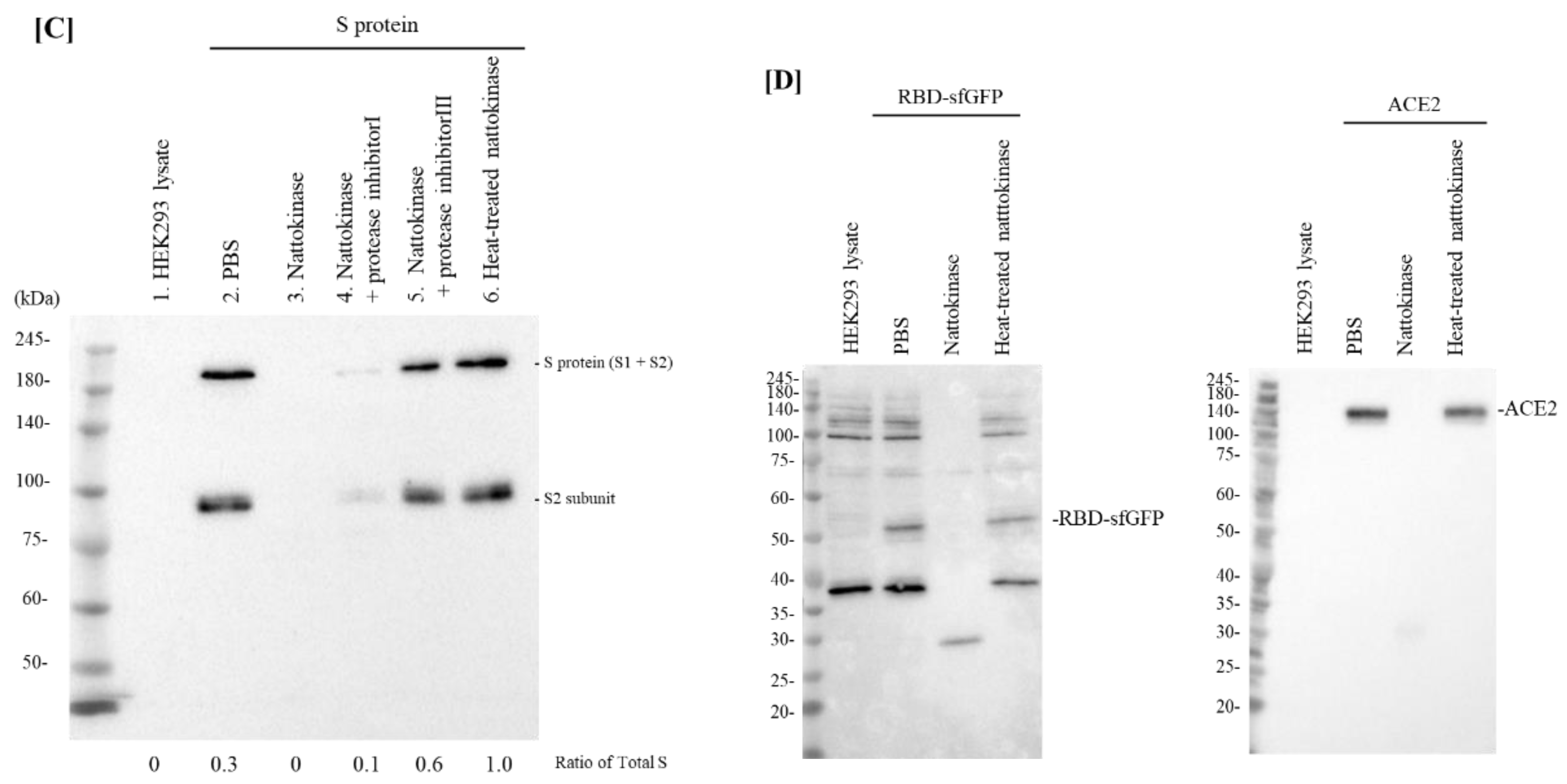
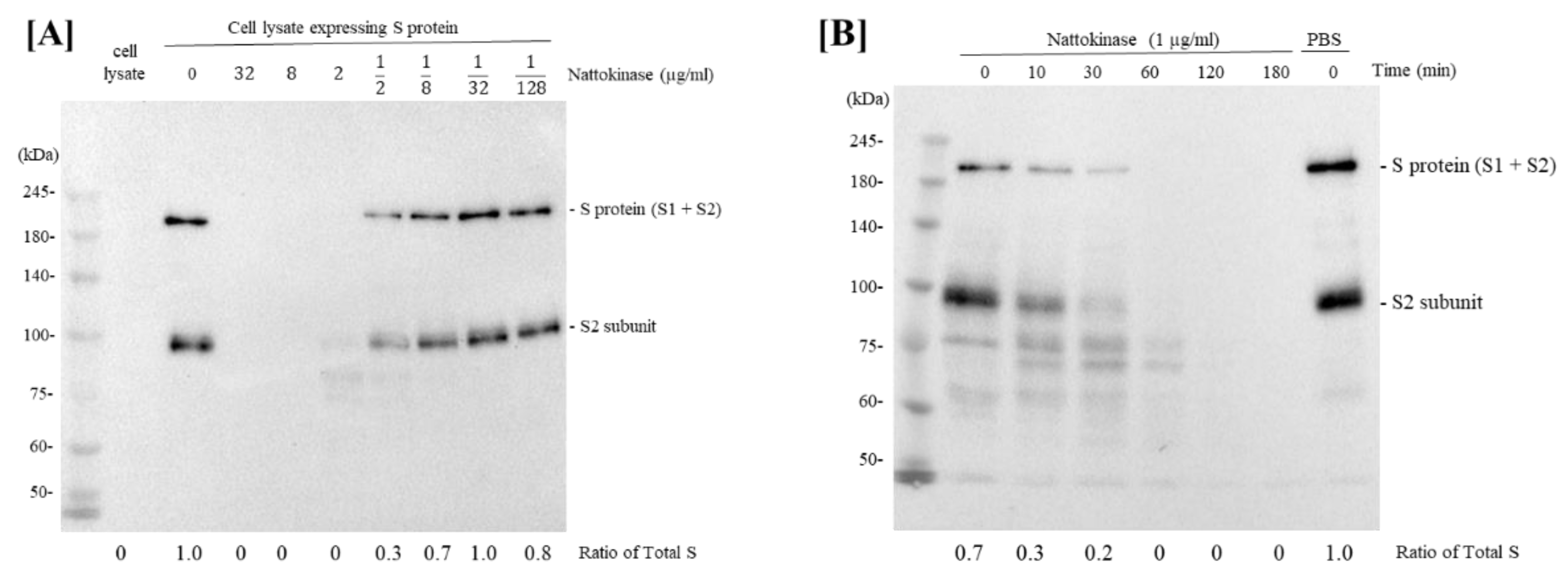
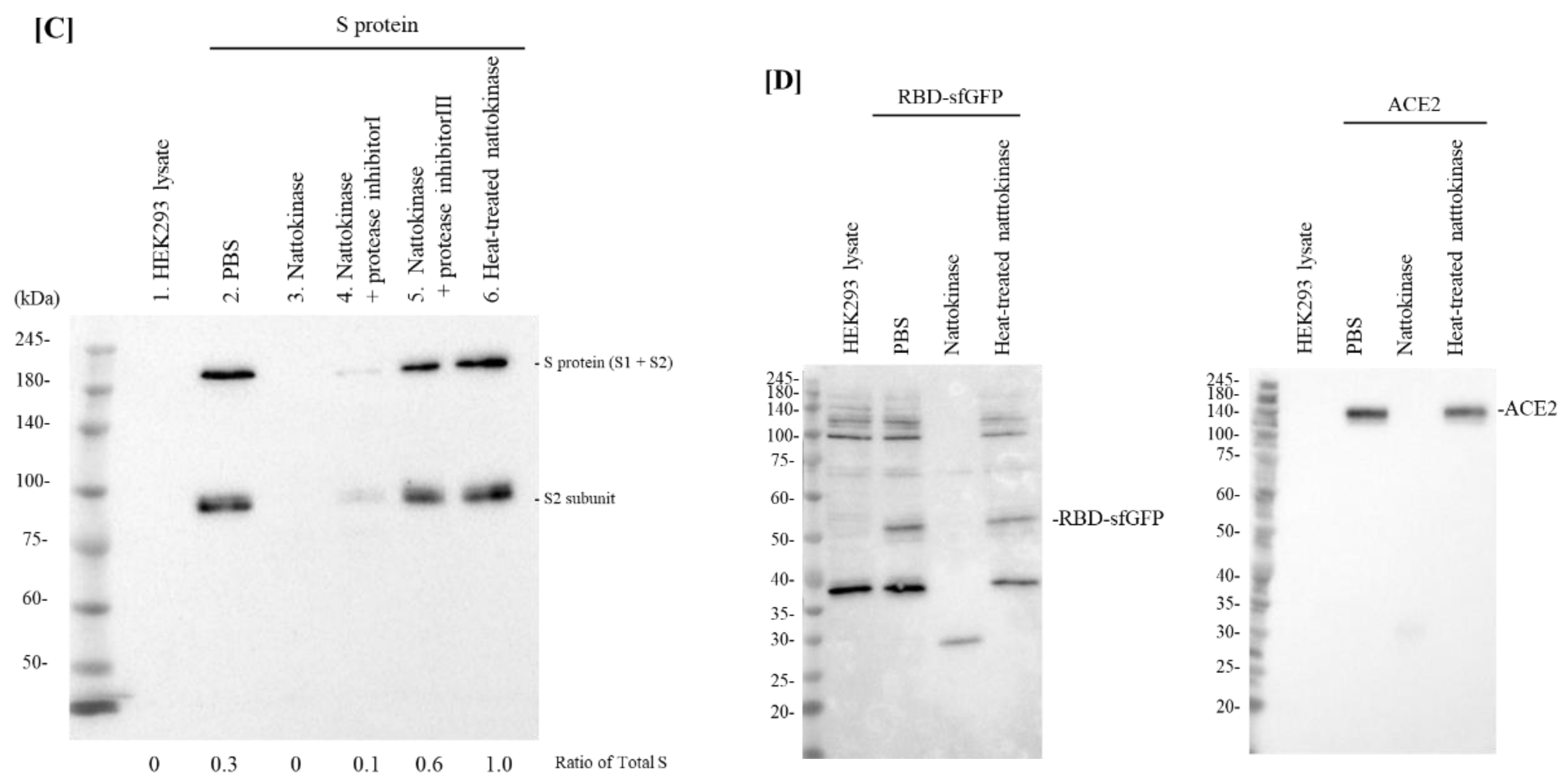
2.1. Degradative Effects of Nattokinase on Spike Protein of SARS-CoV-2 In Vitro
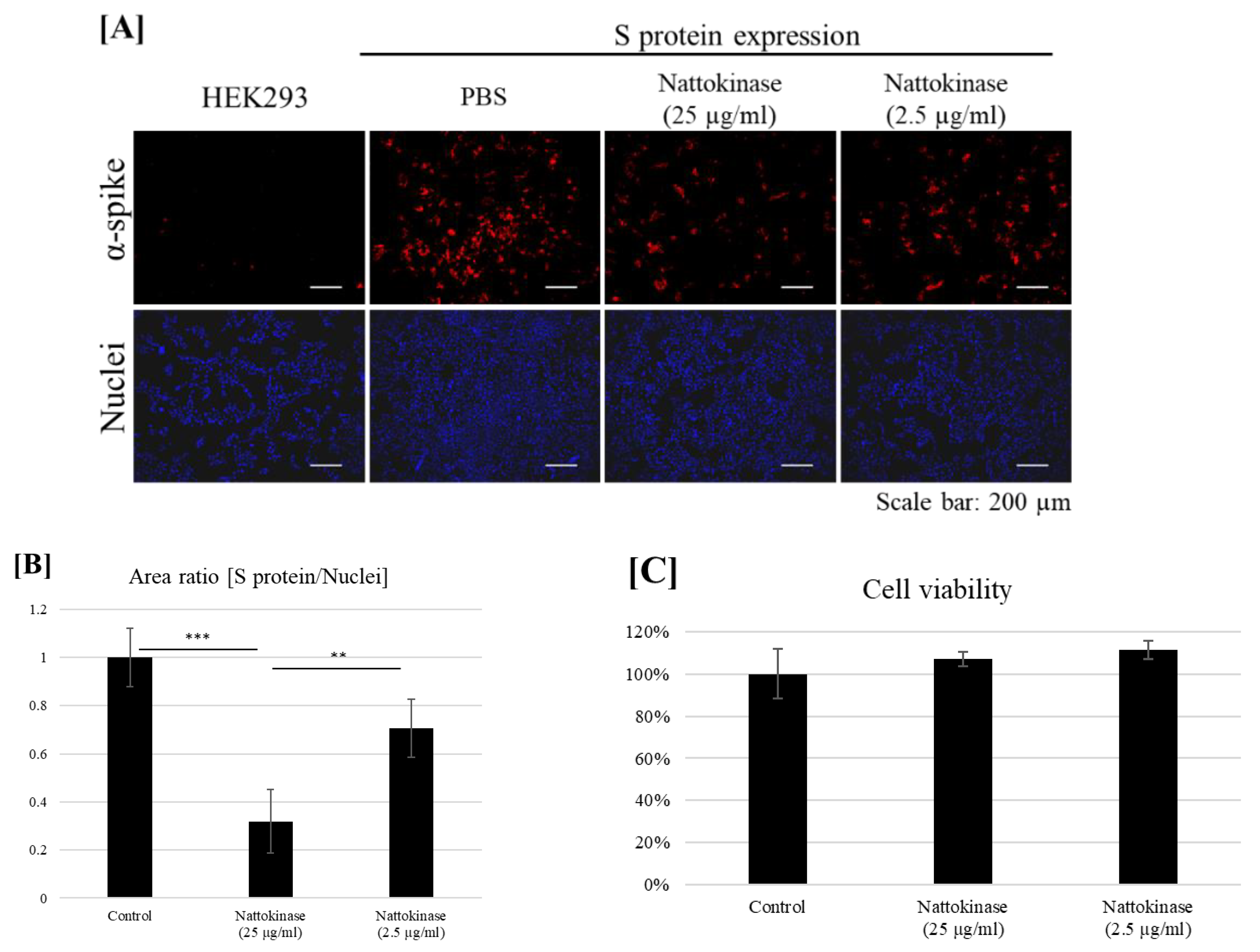
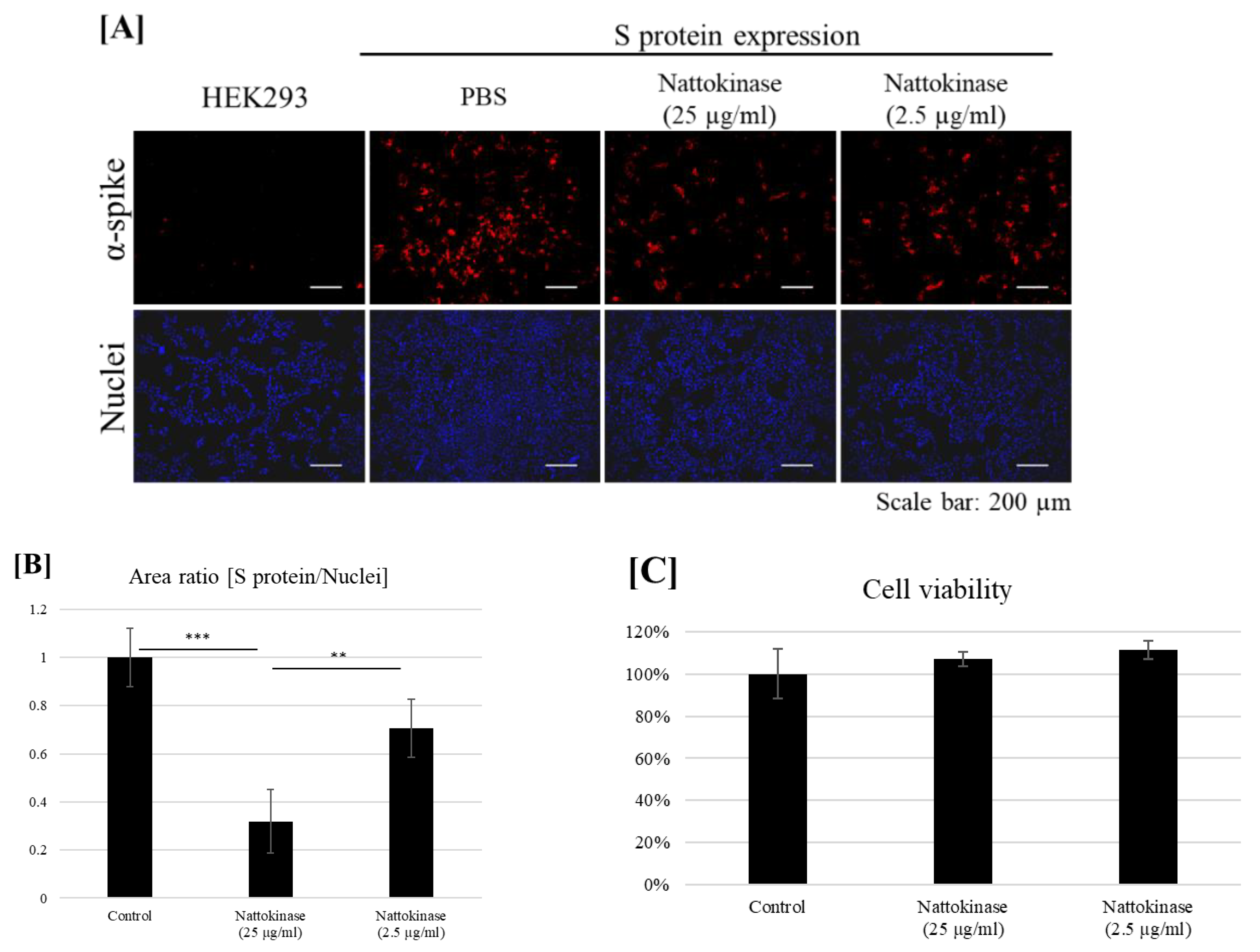
2.2. Degradative Effects of Nattokinase on Spike Protein of SARS-CoV-2 on the Transfected Cell Surface
3. Materials and Methods
3.1. Materials
3.2. Cell Culture and Western Blotting
3.3. Immunofluorescence Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alsobaie, S. Understanding the Molecular Biology of SARS-CoV-2 and the COVID-19 Pandemic: A Review. Infect. Drug Resist. 2021, 14, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Bayati, A.; Kumar, R.; Francis, V.; McPherson, P.S. SARS-CoV-2 infects cells after viral entry via clathrin-mediated endocytosis. J. Biol. Chem. 2021, 296, 100306. [Google Scholar] [CrossRef] [PubMed]
- Bestle, D.; Heindl, M.R.; Limburg, H.; Van Lam van, T.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; et al. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 3, e202000786. [Google Scholar] [CrossRef] [PubMed]
- Sumi, H.; Hamada, H.; Tsushima, H.; Mihara, H.; Muraki, H. A novel fibrinolytic enzyme (nattokinase) in the vegetable cheese Natto; a typical and popular soybean food in the Japanese diet. Experientia 1987, 43, 1110–1111. [Google Scholar] [CrossRef] [PubMed]
- Dabbagh, F.; Negahdaripour, M.; Berenjian, A.; Behfar, A.; Mohammadi, F.; Zamani, M.; Irajie, C.; Ghasemi, Y. Nattokinase: Production and application. Appl. Microbiol. Biotechnol. 2014, 98, 9199–9206. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Yamagata, Y.; Ichishima, E. Nucleotide sequence of the Subtilisin NAT gene, aprN, of Bacillus subtilis (natto). Biosci. Biotech. Bioch. 1992, 56, 1869–1871. [Google Scholar] [CrossRef] [PubMed]
- Urano, T.; Ihara, H.; Umemura, K.; Suzuki, Y.; Oike, M.; Akita, S.; Tsukamoto, Y.; Suzuki, I.; Takada, A. The profibrinolytic enzyme subtilisin NAT purified from Bacillus subtilis Cleaves and inactivates plasminogen activator inhibitor type 1. J. Biol. Chem. 2001, 276, 24690–24696. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kondo, K.; Matsumoto, Y.; Zhao, B.Q.; Otsuguro, K.; Maeda, T.; Tsukamoto, Y.; Urano, T.; Umemura, K. Dietary supplementation of fermented soybean, natto, suppresses intimal thickening and modulates the lysis of mural thrombi after endothelial injury in rat femoral artery. Life Sci. 2003, 73, 1289–1298. [Google Scholar] [CrossRef]
- Hsia, C.H.; Shen, M.C.; Lin, J.S.; Wen, Y.K.; Hwang, K.L.; Cham, T.M.; Yang, N.C. Nattokinase decreases plasma levels of fibrinogen, factor VII, and factor VIII in human subjects. Nutr. Res. 2009, 29, 190–196. [Google Scholar] [CrossRef]
- Weng, Y.; Yao, J.; Sparks, S.; Wang, K.Y. Nattokinase: An Oral Antithrombotic Agent for the Prevention of Cardiovascular Disease. Int. J. Mol. Sci. 2017, 18, 523. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Gum, S.N.; Paik, J.K.; Lim, H.H.; Kim, K.C.; Ogasawara, K.; Inoue, K.; Park, S.; Jang, Y.; Lee, J.H. Effects of nattokinase on blood pressure: A randomized, controlled trial. Hypertens. Res. 2008, 31, 1583–1588. [Google Scholar] [CrossRef]
- Wu, C.; Gao, C.; Lü, S.; Xu, X.; Wen, N.; Zhang, S.; Liu, M. Construction of polylysine dendrimer nanocomposites carrying nattokinase and their application in thrombolysis. J. Biomed. Mater. Res. A 2018, 106, 440–449. [Google Scholar] [CrossRef]
- Lampe, B.J.; English, J.C. Toxicological assessment of nattokinase derived from Bacillus subtilis var natto. Food Chem. Toxicol. 2016, 88, 87–99. [Google Scholar] [CrossRef]
- Kou, Y.; Feng, R.; Chen, J.; Duan, L.; Wang, S.; Hu, Y.; Zhang, N.; Wang, T.; Deng, Y.; Song, Y. Development of a nattokinase-polysialic acid complex for advanced tumor treatment. Eur. J. Pharm. Sci. 2020, 145, 105241. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, Y.; Qian, J.; Wu, S.; Ji, Y.; Liu, Y.; Zeng, J.; Gong, A. Nattokinase Crude Extract Inhibits Hepatocellular Carcinoma Growth in Mice. J. Microbiol. Biotechnol. 2019, 29, 1281–1287. [Google Scholar] [CrossRef]
- Oba, M.; Rongduo, W.; Saito, A.; Okabayashi, T.; Yokota, T.; Yasuoka, J.; Sato, Y.; Nishifuji, K.; Wake, H.; Nibu, Y.; et al. Natto extract, a Japanese fermented soybean food, directly inhibits viral infections including SARS-CoV-2 in vitro. Biochem. Biophys. Res. Commun. 2021, 570, 21–25. [Google Scholar] [CrossRef]
- Smaoui, M.R.; Yahyaoui, H. Unraveling the stability landscape of mutations in the SARS-CoV-2 receptor-binding domain. Sci. Rep. 2021, 11, 9166. [Google Scholar] [CrossRef]
- Carter, P.; Wells, J.A. Dissecting the catalytic triad of a serine protease. Nature 1988, 332, 564–568. [Google Scholar] [CrossRef]
- Yanagisawa, Y.; Chatake, T.; Naito, S.; Ohsugi, T.; Yatagai, C.; Sumi, H.; Kawaguchi, A.; Chiba-Kamosida, K.; Ogawa, M.; Adachi, T.; et al. X-ray structure determination and deuteration of nattokinase. J. Synchrotron Radiat. 2013, 20 Pt 6, 875–879. [Google Scholar] [CrossRef]
- Chen, H.; McGowan, E.M.; Ren, N.; Lal, S.; Nassif, N.; Shad-Kaneez, F.; Qu, X.; Lin, Y. Nattokinase: A Promising Alternative in Prevention and Treatment of Cardiovascular Diseases. Biomark. Insights 2018, 13, 1177271918785130. [Google Scholar] [CrossRef] [Green Version]
- Cancarevic, I.; Malik, B.H. SARS-CoV-2 (COVID 19) Infection in Hypertensive Patients and in Patients with Cardiac Disease. Cureus 2020, 12, e8557. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanikawa, T.; Kiba, Y.; Yu, J.; Hsu, K.; Chen, S.; Ishii, A.; Yokogawa, T.; Suzuki, R.; Inoue, Y.; Kitamura, M. Degradative Effect of Nattokinase on Spike Protein of SARS-CoV-2. Molecules 2022, 27, 5405. https://doi.org/10.3390/molecules27175405
Tanikawa T, Kiba Y, Yu J, Hsu K, Chen S, Ishii A, Yokogawa T, Suzuki R, Inoue Y, Kitamura M. Degradative Effect of Nattokinase on Spike Protein of SARS-CoV-2. Molecules. 2022; 27(17):5405. https://doi.org/10.3390/molecules27175405
Chicago/Turabian StyleTanikawa, Takashi, Yuka Kiba, James Yu, Kate Hsu, Shinder Chen, Ayako Ishii, Takami Yokogawa, Ryuichiro Suzuki, Yutaka Inoue, and Masashi Kitamura. 2022. "Degradative Effect of Nattokinase on Spike Protein of SARS-CoV-2" Molecules 27, no. 17: 5405. https://doi.org/10.3390/molecules27175405
APA StyleTanikawa, T., Kiba, Y., Yu, J., Hsu, K., Chen, S., Ishii, A., Yokogawa, T., Suzuki, R., Inoue, Y., & Kitamura, M. (2022). Degradative Effect of Nattokinase on Spike Protein of SARS-CoV-2. Molecules, 27(17), 5405. https://doi.org/10.3390/molecules27175405