
Biophysical Characterization of the Interaction between a Transport Human Plasma Protein and the 5,10,15,20-Tetra(pyridine-4-yl)porphyrin




Abstract
:1. Introduction
2. Results
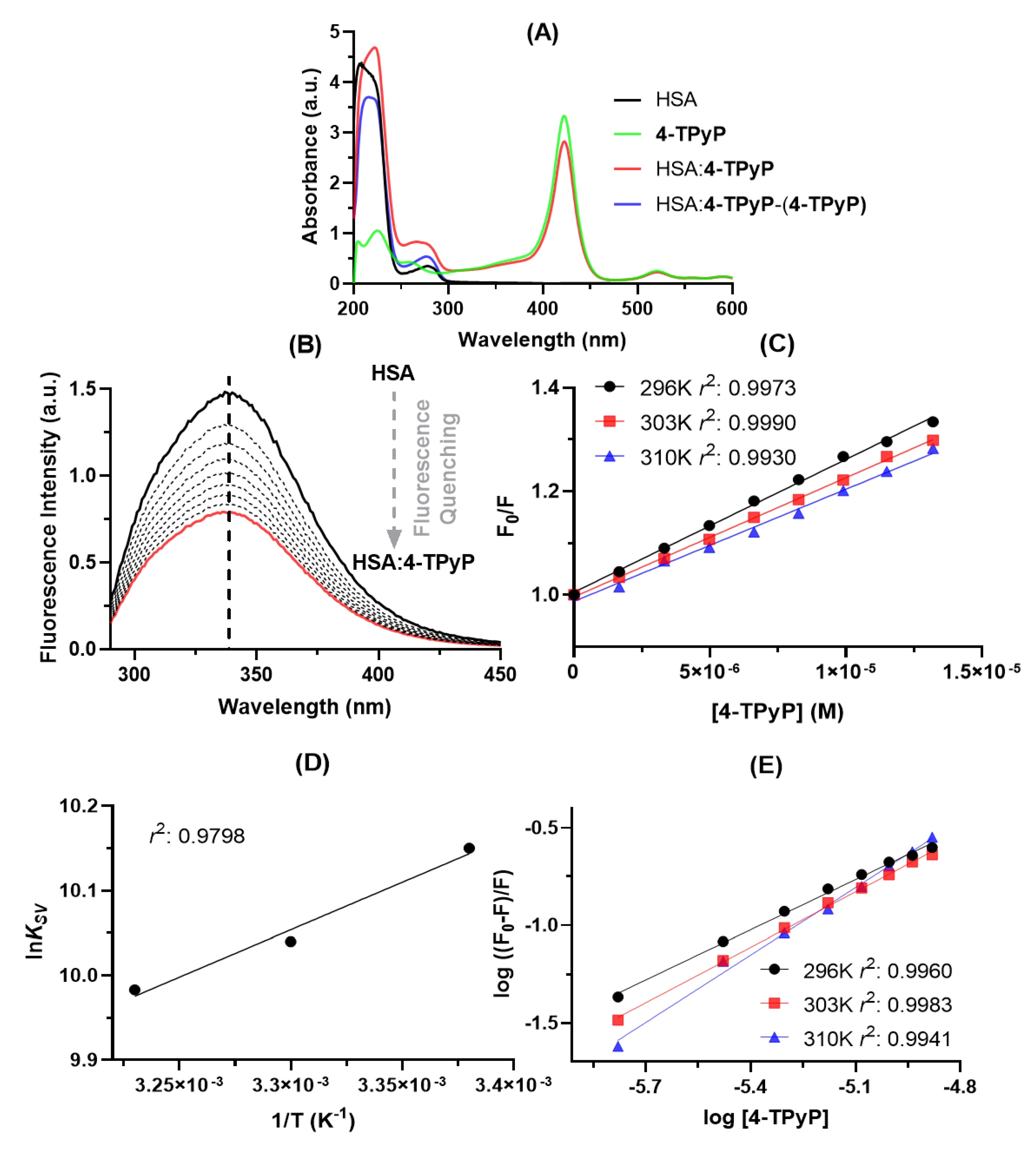
2.1. Experimental Binding Capacity of 4-TPyP to HSA
2.2. Structural and Microenvironment Perturbation of HSA upon 4-TPyP Binding
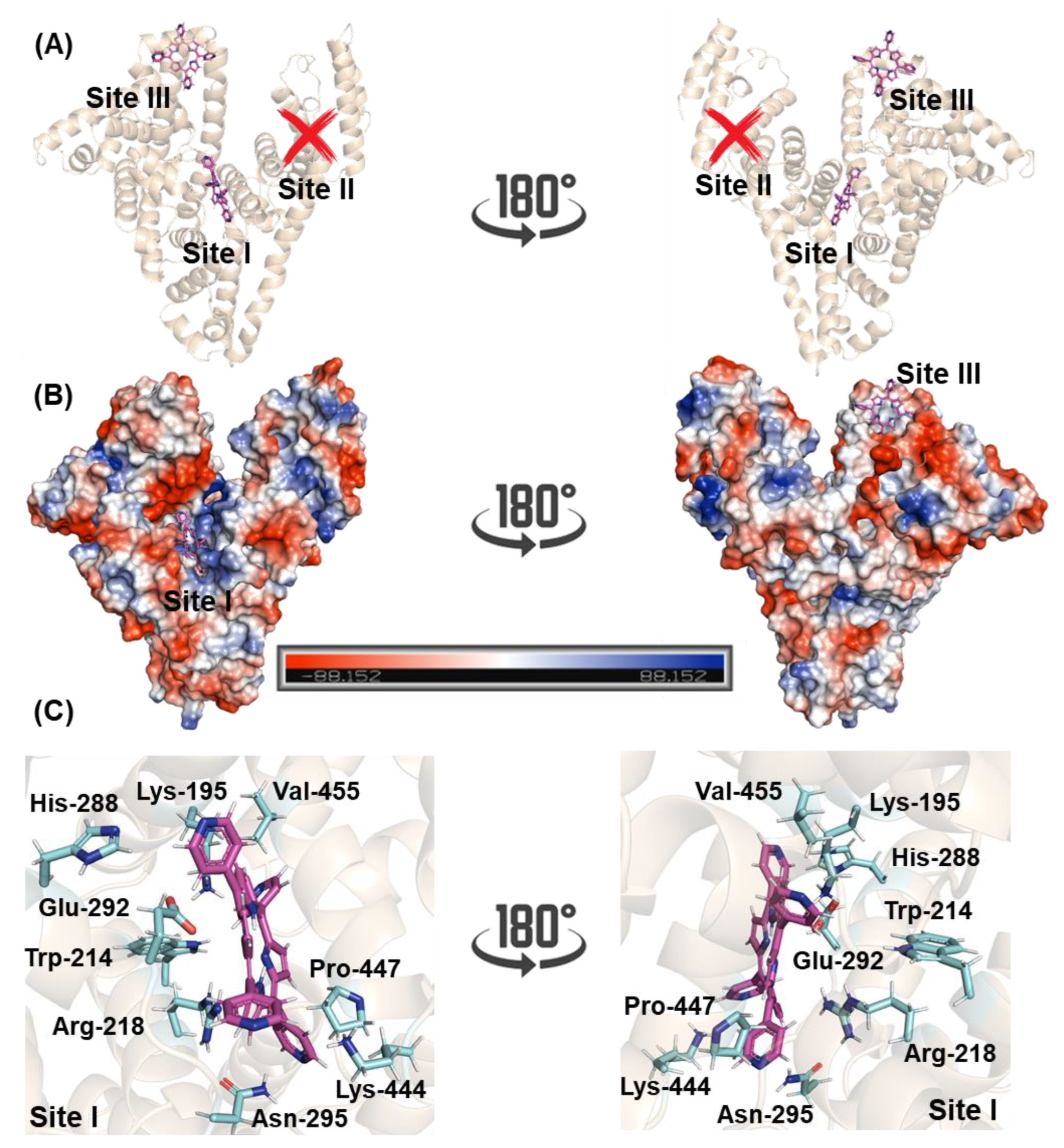
2.3. Molecular-Level Explanation on the Binding Capacity of 4-TPyP to HSA
3. Discussion
4. Materials and Methods
4.1. Chemicals and Instruments
4.2. Spectroscopic Measurements
4.3. Molecular Docking Procedure
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Lee, P.; Wu, X. Review: Modifications of human serum albumin and their binding effect. Curr. Pharm. Des. 2015, 21, 1862–1865. [Google Scholar] [CrossRef] [Green Version]
- Merlot, A.M.; Kalinowski, D.S.; Richardson, D.R. Unraveling the mysteries of serum albumin—More than just a serum protein. Front. Physiol. 2014, 5, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Luo, H.; Chen, W.; Huang, Q.; Zheng, K.; Xu, D.; Li, S.; Liu, A.; Huang, L.; Zheng, Y.; et al. Pharmacokinetics, tissue distribution, and human serum albumin binding properties of delicaflavone, a novel anti-tumor candidate. Front. Pharmacol. 2021, 12, 761884. [Google Scholar] [CrossRef] [PubMed]
- Zhivkova, Z.D. Studies on drug-human serum albumin binding: The current state of the matter. Curr. Pharm. Des. 2015, 21, 1817–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, D.C.; Ho, J.X. Structure of serum albumin. Adv. Protein Chem. 1994, 45, 153–203. [Google Scholar]
- Wardell, M.; Wang, Z.; Ho, J.X.; Robert, J.; Ruker, F.; Ruble, J.; Carter, D.C. The atomic structure of human methemalbumin at 1.9 A. Biochem. Biophys. Res. Commun. 2002, 291, 913–918. [Google Scholar] [CrossRef]
- Sudlow, G.; Birkett, D.J.; Wade, D.N. Further characterization of specific drug binding sites on human serum albumin. Mol. Pharmacol. 1976, 12, 1052–1061. [Google Scholar]
- Kou, J.; Dou, D.; Yang, L. Porphyrin photosensitizers in photodynamic therapy and its applications. Oncotarget 2017, 8, 81591–81603. [Google Scholar] [CrossRef] [Green Version]
- Amos-Tautua, B.M.; Songca, S.P.; Oluwafemi, O.S. Application of porphyrins in antibacterial photodynamic therapy. Molecules 2019, 24, 2456. [Google Scholar] [CrossRef] [Green Version]
- Tasso, T.T.; Tsubone, T.M.; Baptista, M.S.; Mattiazzi, L.M.; Acunha, T.V.; Iglesias, B.A. Isomeric effect on the properties of tetraplatinated porphyrins showing optimized phototoxicity for photodynamic therapy. Dalton Trans. 2017, 46, 11037–11045. [Google Scholar] [CrossRef]
- Chen, X.; Cai, Y.; Zhao, Y.; Ma, H.; Wu, D.; Du, B.; Wei, Q. Quenching and binding mechanism of the intrinsic fluorescence of bovine serum albumin by 5-phenyl-10,15,20-tri-(4-pyridyl)-porphyrin. J. Porphyr. Phthalocyanines 2009, 13, 933–938. [Google Scholar] [CrossRef]
- Jokar, S.; Pourjavadi, A.; Adeli, M. Albumin–graphene oxide conjugates; carriers for anticancer drugs. RSC Adv. 2014, 4, 33001. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, F.; Xiong, X.; Ge, Y.; Liu, Y. Spectroscopic and molecular docking studies on the interaction of dimetridazole with human serum albumin. J. Chil. Chem. Soc. 2013, 58, 1717–1721. [Google Scholar] [CrossRef] [Green Version]
- Bordbar, A.K.; Eslami, A.; Tangestaninejad, S. Spectral investigations of the solution properties of 5,10,15,20-tetrakis(4-N-benzyl-pyridyl)porphyrin (TBzPyP) and its interaction with human serum albumin (HSA). J. Porphyr. Phthalocyanines 2002, 6, 225–232. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer: New York, NY, USA, 2006. [Google Scholar]
- Chaves, O.A.; da Silva, V.A.; Sant’Anna, C.M.R.; Ferreira, A.B.B.; Ribeiro, T.A.N.; de Carvalho, M.G.; Cesarin-Sobrinho, D.; Netto-Ferreira, J.C. Binding studies of lophirone B with bovine serum albumin (BSA): Combination of spectroscopic and molecular docking techniques. J. Mol. Struct. 2017, 1128, 606–611. [Google Scholar] [CrossRef]
- Montalti, M.; Credi, A.; Prodi, L.; Gandolfi, M.T. Handbook of Photochemistry, 3rd ed.; CRC Press; Taylor & Francis: Boca Raton, FL, USA, 2006. [Google Scholar]
- Sun, H.; Liu, Y.; Li, M.; Han, S.; Yang, X.; Liu, R. Toxic effects of chrysoidine on human serum albumin: Isothermal titration calorimetry and spectroscopic investigations. Luminescence 2016, 31, 335–340. [Google Scholar] [CrossRef]
- Chaves, O.A.; Soares, M.A.G.; de Oliveira, M.C.C. Monosaccharides interact weakly with human serum albumin. Insights for the functional perturbations on the binding capacity of albumin. Carbohydr. Res. 2021, 501, 108274. [Google Scholar] [CrossRef]
- Amiri, M.; Jankeje, K.; Albani, J.R. Origin of fluorescence lifetimes in human serum albumin. Studies on native and denatured protein. J. Fluoresc. 2010, 20, 651–656. [Google Scholar] [CrossRef]
- Chaves, O.A.; Cesarin-Sobrinho, D.; Sant’Anna, C.M.R.; de Carvalho, M.G.; Suzart, L.R.; Catunda-Junior, F.E.A.; Netto-Ferreira, J.C.; Ferreira, A.B.B. Probing the interaction between 7-O-β-D-glucopyranosyl-6-(3-methylbut-2-enyl)-5,4′-dihydroxyflavonol with bovine serum albumin (BSA). J. Photochem. Photobiol. A 2017, 336, 32–41. [Google Scholar] [CrossRef]
- Chaves, O.A.; Acunha, T.V.; Iglesias, B.A.; Jesus, C.S.H.; Serpa, C. Effect of peripheral platinum(II) bipyridyl complexes on the interaction of tetra-cationic porphyrins with human serum albumin. J. Mol. Liq. 2020, 301, 112466. [Google Scholar] [CrossRef]
- Safarnejad, A.; Shaghaghi, M.; Dehghan, G.; Soltani, S. Binding of carvedilol to serum albumins investigated by multi-spectroscopic and molecular modeling methods. J. Lumin. 2016, 176, 149–158. [Google Scholar] [CrossRef]
- Caruso, I.P.; Filho, J.M.B.; de Araújo, A.S.; de Souza, F.P.; Fossey, M.A.; Cornélio, M.L. An integrated approach with experimental and computational tools outlining the cooperative binding between 2-phenylchromone and human serum albumin. Food Chem. 2016, 196, 935–942. [Google Scholar] [CrossRef]
- Karthiga, D.; Chandrasekaran, N.; Mukherjee, N. Spectroscopic studies on the interactions of bovine serum albumin in presence of silver nanorod. J. Mol. Liq. 2017, 232, 251–257. [Google Scholar] [CrossRef]
- Hashempour, S.; Shahabadi, N.; Adewoye, A.; Murphy, B.; Rouse, C.; Salvatore, B.A.; Stratton, C.; Mahdavian, E. Binding studies of AICAR and human serum albumin by spectroscopic, theoretical, and computational methodologies. Molecules 2020, 25, 5410. [Google Scholar] [CrossRef] [PubMed]
- Byadagi, K.; Meti, M.; Nandibewoor, S.; Chimatadar, S. Investigation of binding behaviour of procainamide hydrochloride with human serum albumin using synchronous, 3D fluorescence and circular dichroism. J. Pharm. Anal. 2017, 7, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Chaves, O.A.; de Oliveira, C.H.C.S.; Ferreira, R.C.; Pereira, R.P.; de Melos, J.L.R.; Rodrigues-Santos, C.E.; Echevarria, A.; Cesarin-Sobrinho, D. Investigation of interaction between human plasmatic albumin and potential fluorinated anti-trypanosomal drugs. J. Fluor. Chem. 2017, 199, 103–112. [Google Scholar] [CrossRef]
- Bertucci, C.; de Simone, A.; Pistolozzi, M.; Rosini, M. Reversible human serum albumin binding of lipocrine: A circular dichroism study. Chirality 2011, 23, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Buddanavar, A.T.; Nandibewoor, S.T. Multi-spectroscopic characterization of bovine serum albumin upon interaction with atomoxetine. J. Pharm. Sci. 2017, 7, 148–155. [Google Scholar] [CrossRef]
- Nasruddin, A.N.; Feroz, S.R.; Mukarram, A.K.; Mohamad, S.B.; Tayyab, S. Fluorometric and molecular docking investigation on the binding characteristics of SB202190 to human serum albumin. J. Lumin. 2016, 174, 77–84. [Google Scholar] [CrossRef]
- Sharma, A.S.; Anandakumar, S.; Ilanchelian, M. A combined spectroscopic and molecular docking study on site selective binding interaction of Toluidine blue O with Human and Bovine serum albumins. J. Lumin. 2014, 151, 206–218. [Google Scholar] [CrossRef]
- Johansson, E.; Nielsen, A.D.; Demuth, H.; Wiberg, C.; Schjødt, C.B.; Huang, T.; Chen, J.; Jensen, S.; Petersen, J.; Thygesen, P. Identification of binding sites on human serum albumin for somapacitan, a long-acting growth hormone derivative. Biochemistry 2020, 59, 1410–1419. [Google Scholar] [CrossRef] [PubMed]
- Da Silveira, C.H.; Chaves, O.A.; Marques, A.C.; Rosa, N.M.O.; Costa, L.A.S.; Iglesias, B.A. Synthesis, photophysics, computational approaches, and biomolecule interactive studies of metalloporphyrins containing pyrenyl units: Influence of the metal center. Europ. J. Inorg. Chem. 2022, 12, e2022000075. [Google Scholar] [CrossRef]
- Rinco, O.; Brenton, J.; Douglas, A.; Maxwell, A.; Henderson, M.; Indrelie, K.; Wessels, J.; Widin, J. The effect of porphyrin structure on binding to human serum albumin by fluorescence spectroscopy. J. Photochem. Photobiol. A 2009, 208, 91–96. [Google Scholar] [CrossRef]
- Yang, L.; Ji, Z.; Peng, Z.; Cheng, G. A novel low symmetry sulfur containing porphyrazine: Synthesis and its interaction with serum albumin. J. Porphyr. Phthalocyanines 2003, 7, 420–425. [Google Scholar] [CrossRef]
- Cohen, S.; Margalit, R. Binding of porphyrin to human serum albumin. Structure–activity relationships. Biochem. J. 1990, 270, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Chaves, O.A.; Menezes, L.B.; Iglesias, B.A. Multiple spectroscopic and theoretical investigation of meso-tetra-(4-pyridyl)porphyrin ruthenium(II) complexes in HSA binding studies. Effect of Zn(II) in protein binding. J. Mol. Liq. 2019, 294, 111581. [Google Scholar] [CrossRef]
- Ding, Y.; Lin, B.; Huie, C.W. Binding studies of porphyrins to human serum albumin using affinity capillary electrophoresis. Electrophoresis 2001, 22, 2210–2216. [Google Scholar] [CrossRef]
- Kubát, P.; Lang, K.; Anzenbacher, P. Modulation of porphyrin binding to serum albumin by pH. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2004, 1670, 40–48. [Google Scholar] [CrossRef]
- Naveenraj, S.; Anandan, S. Binding of serum albumins with bioactive substances—Nanoparticles to drugs. J. Photochem. Photobiol. C 2013, 14, 53–71. [Google Scholar] [CrossRef]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef]
- Chaves, O.A.; Jesus, C.S.H.; Cruz, P.F.; Sant’Anna, C.M.R.; Brito, R.M.M.; Serpa, C. Evaluation by fluorescence, STD-NMR, docking and semi-empirical calculations of the o-NBA photo-acid interaction with BSA. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2016, 169, 175–181. [Google Scholar] [CrossRef]
- Matei, I.; Hillebrand, M. Interaction of kaempferol with human serum albumin: A fluorescence and circular dichroism study. J. Pharm. Biomed. Anal. 2010, 51, 768–773. [Google Scholar] [CrossRef]
- Chaves, O.A.; Amorim, A.P.O.; Castro, L.H.E.; de Sant’Anna, C.M.R.; de Oliveira, M.C.C.; Cesarin-Sobrinho, D.; Netto-Ferreira, J.C.; Ferreira, A.B.B. Fluorescence and docking studies of the interaction between human serum albumin and pheophytin. Molecules 2015, 20, 19526–19539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, N.; Ajmal, M.R.; Rabbani, G.; Ahmad, E.; Khan, R.H. A comprehensive insight into binding of hippuric acid to human serum albumin: A study to uncover its impaired elimination through hemodialysis. PLoS ONE 2013, 8, e71422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossweiner, L.I. A note on the analysis of ligand binding by the double-logarithmic plot. J. Photochem. Photobiol. B 2000, 58, 175–177. [Google Scholar] [CrossRef]
- Hein, K.L.; Kragh-Hansen, U.; Morth, J.P.; Jeppesen, M.D.; Otzen, D.; Moller, J.V.; Nissen, P. Crystallographic analysis reveals a unique lidocaine binding site on human serum albumin. J. Struct. Biol. 2010, 171, 353–360. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T (K) | KSV × 104 (M−1) [a] | kq × 1012 (M−1s−1) [a] | n [b] | ΔH° (kJ/mol) [c] | ΔS° (J/molK) [c] | ΔG° (kJ/mol) |
|---|---|---|---|---|---|---|
| 296 | 2.57 ± 0.05 | 5.39 ± 0.11 | 0.857 ± 0.022 | −9.31 ± 1.34 | 52.9 ± 4.4 | −25.0 |
| 303 | 2.30 ± 0.03 | 4.84 ± 0.06 | 0.948 ± 0.016 | −25.3 | ||
| 310 | 2.17 ± 0.07 | 4.55 ± 0.14 | 1.15 ± 0.04 | −25.7 |
| System | Peak | Peak Position (λexc/λem nm/nm) | Intensity × 103 (a.u.) |
|---|---|---|---|
| a | 225/225 → 355/355 | 4.00 → 1.89 | |
| HSA | b | 230/460 | 2.01 |
| I | 280/335 | 2.13 | |
| II | 225/330 | 3.56 | |
| a | 225/225 → 355/355 | 3.16 → 1.66 | |
| HSA:4-TPyP | b | 230/460 | 1.66 |
| I | 280/340 | 1.90 | |
| II | 225/335 | 2.97 |
| Amino Acid Residue | Interaction | Distance (Å) |
|---|---|---|
| Lys-195 | Hydrogen bonding | 3.40 |
| Trp-214 | Van der Waals | 3.50 |
| Arg-218 | Hydrogen bonding | 3.40 |
| His-288 | Van der Waals | 3.10 |
| Glu-292 | Hydrogen bonding | 3.40 |
| Asn-295 | Hydrogen bonding | 3.30 |
| Lys-444 | Van der Waals | 2.90 |
| Pro-447 | Van der Waals | 3.70 |
| Val-455 | Van der Waals | 3.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves, O.A.; Iglesias, B.A.; Serpa, C. Biophysical Characterization of the Interaction between a Transport Human Plasma Protein and the 5,10,15,20-Tetra(pyridine-4-yl)porphyrin. Molecules 2022, 27, 5341. https://doi.org/10.3390/molecules27165341
Chaves OA, Iglesias BA, Serpa C. Biophysical Characterization of the Interaction between a Transport Human Plasma Protein and the 5,10,15,20-Tetra(pyridine-4-yl)porphyrin. Molecules. 2022; 27(16):5341. https://doi.org/10.3390/molecules27165341
Chicago/Turabian StyleChaves, Otávio Augusto, Bernardo A. Iglesias, and Carlos Serpa. 2022. "Biophysical Characterization of the Interaction between a Transport Human Plasma Protein and the 5,10,15,20-Tetra(pyridine-4-yl)porphyrin" Molecules 27, no. 16: 5341. https://doi.org/10.3390/molecules27165341
APA StyleChaves, O. A., Iglesias, B. A., & Serpa, C. (2022). Biophysical Characterization of the Interaction between a Transport Human Plasma Protein and the 5,10,15,20-Tetra(pyridine-4-yl)porphyrin. Molecules, 27(16), 5341. https://doi.org/10.3390/molecules27165341