
Autofluorescent Biomolecules in Diptera: From Structure to Metabolism and Behavior



Abstract
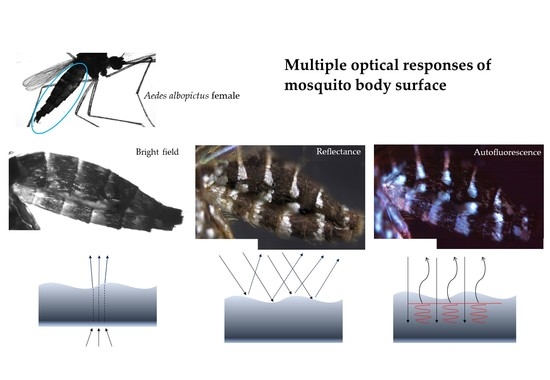
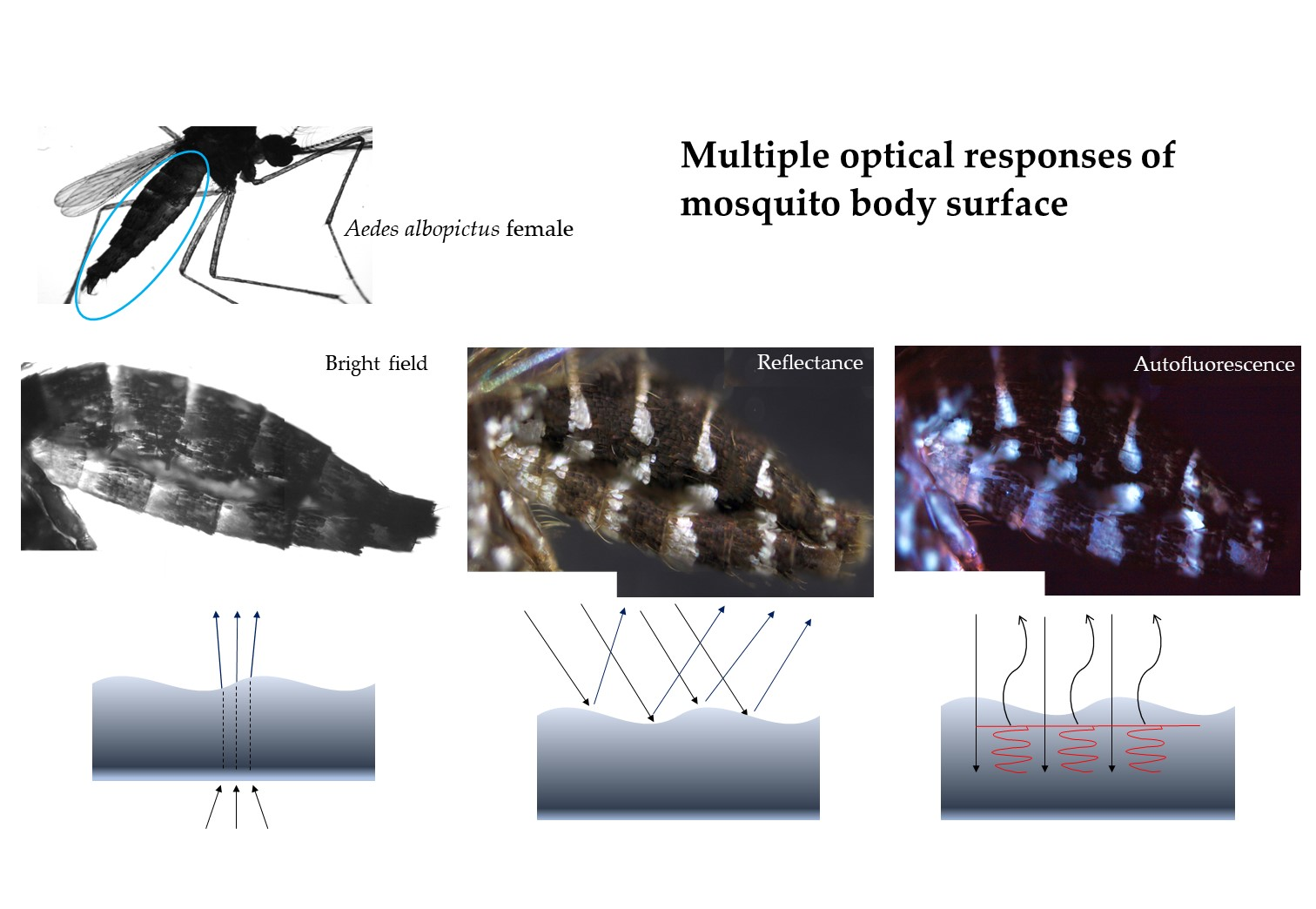
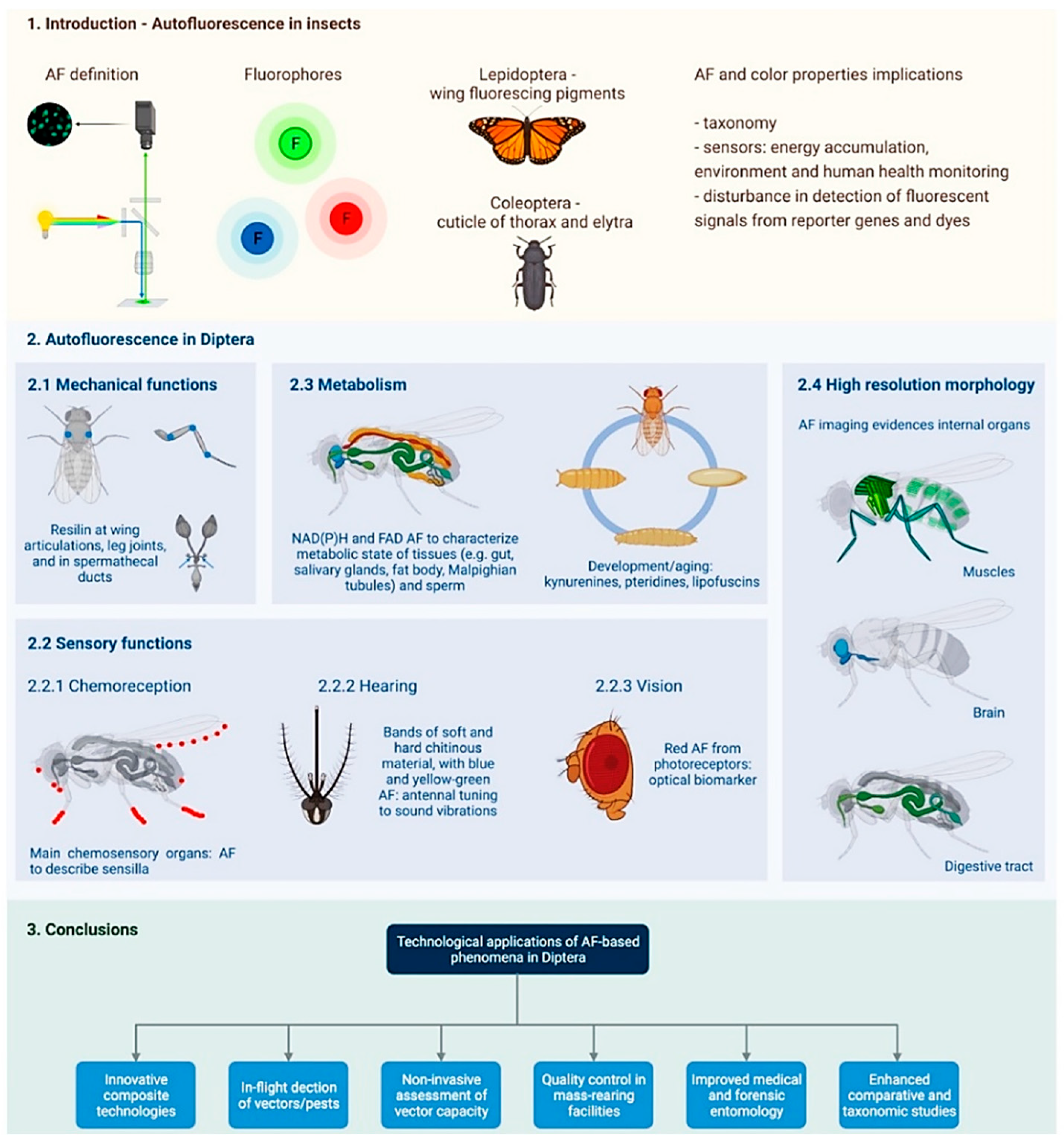
1. Introduction
2. Autofluorescence in Diptera
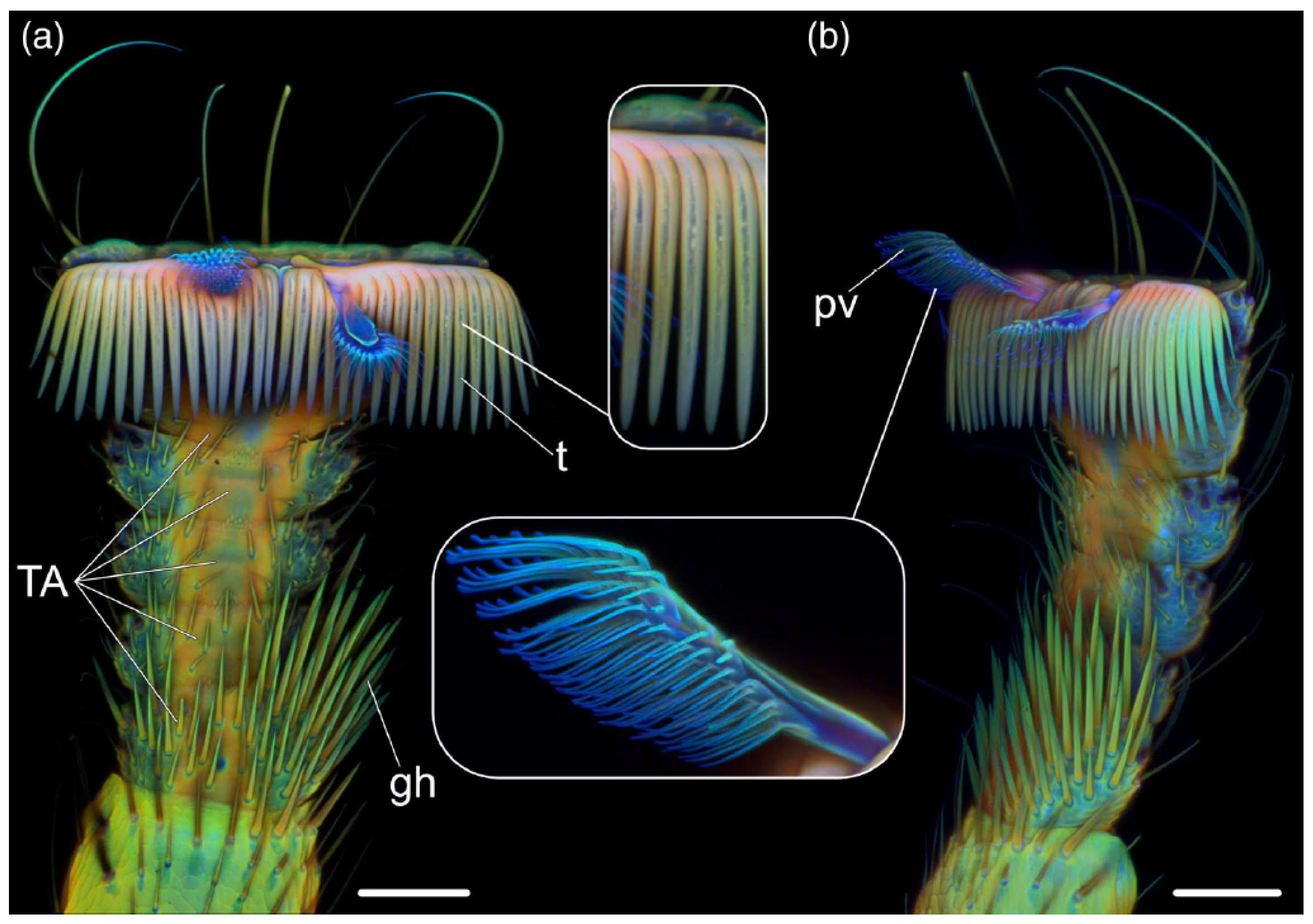
2.1. Mechanical Functions
2.2. Sensory Functions
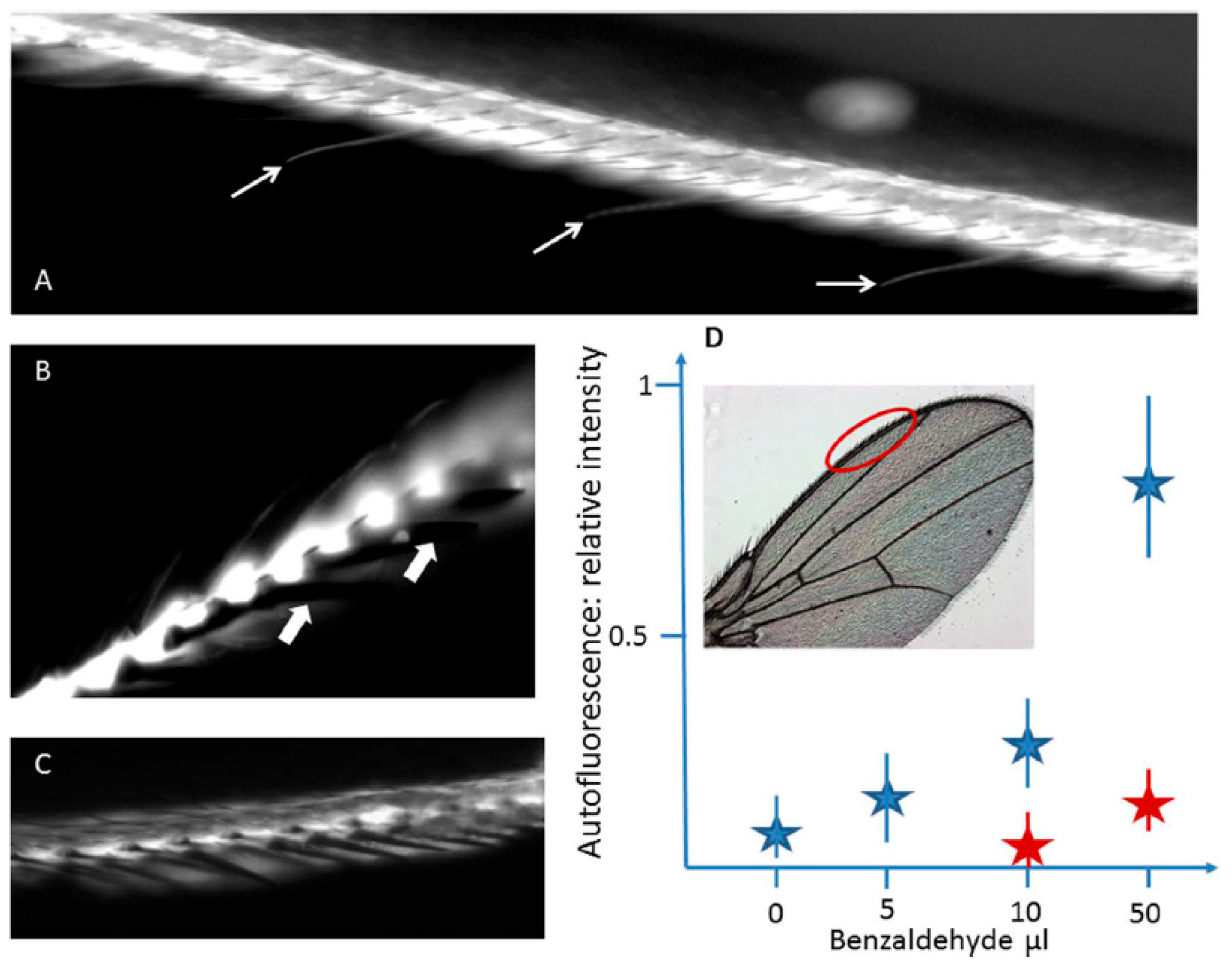
2.2.1. Chemoreception
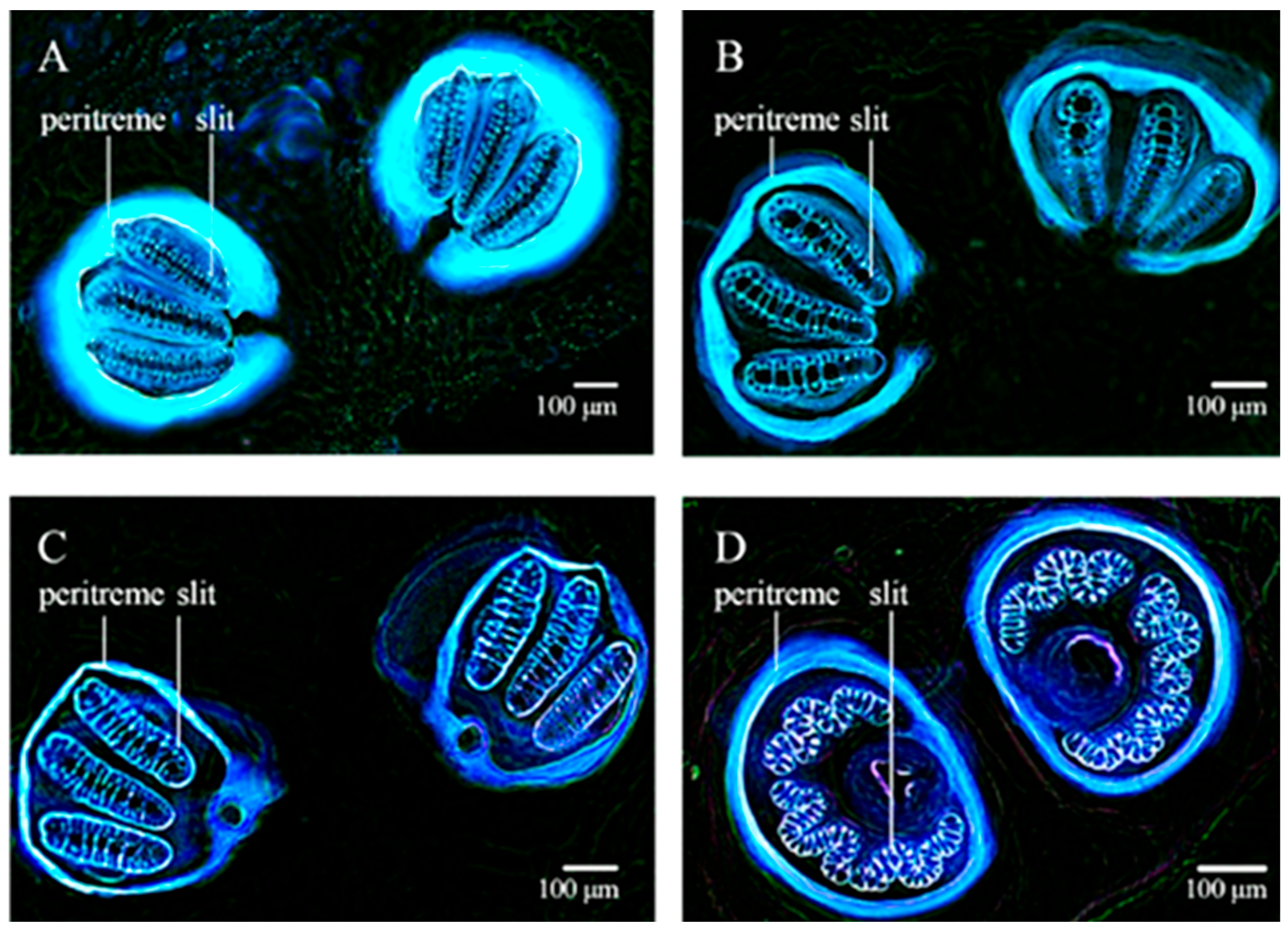
2.2.2. Hearing
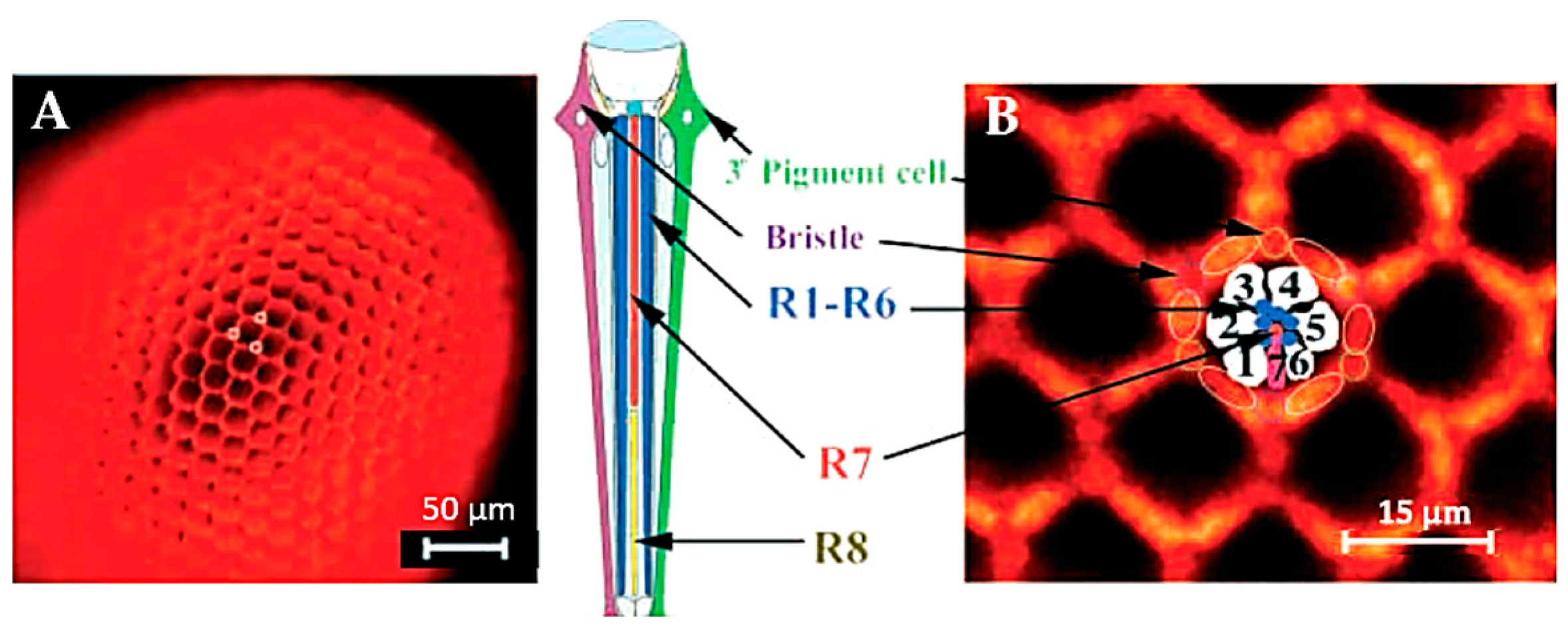
2.2.3. Vision
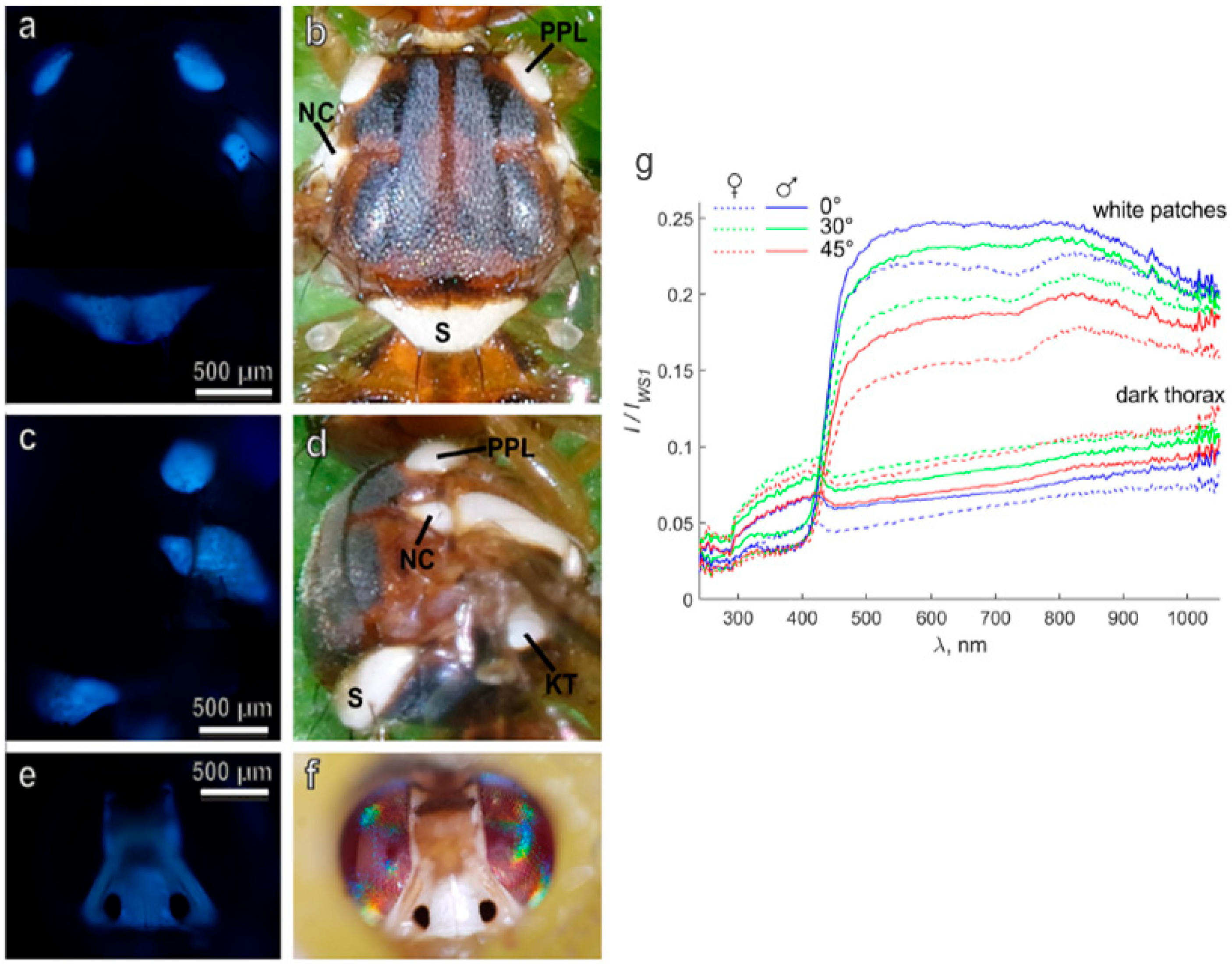
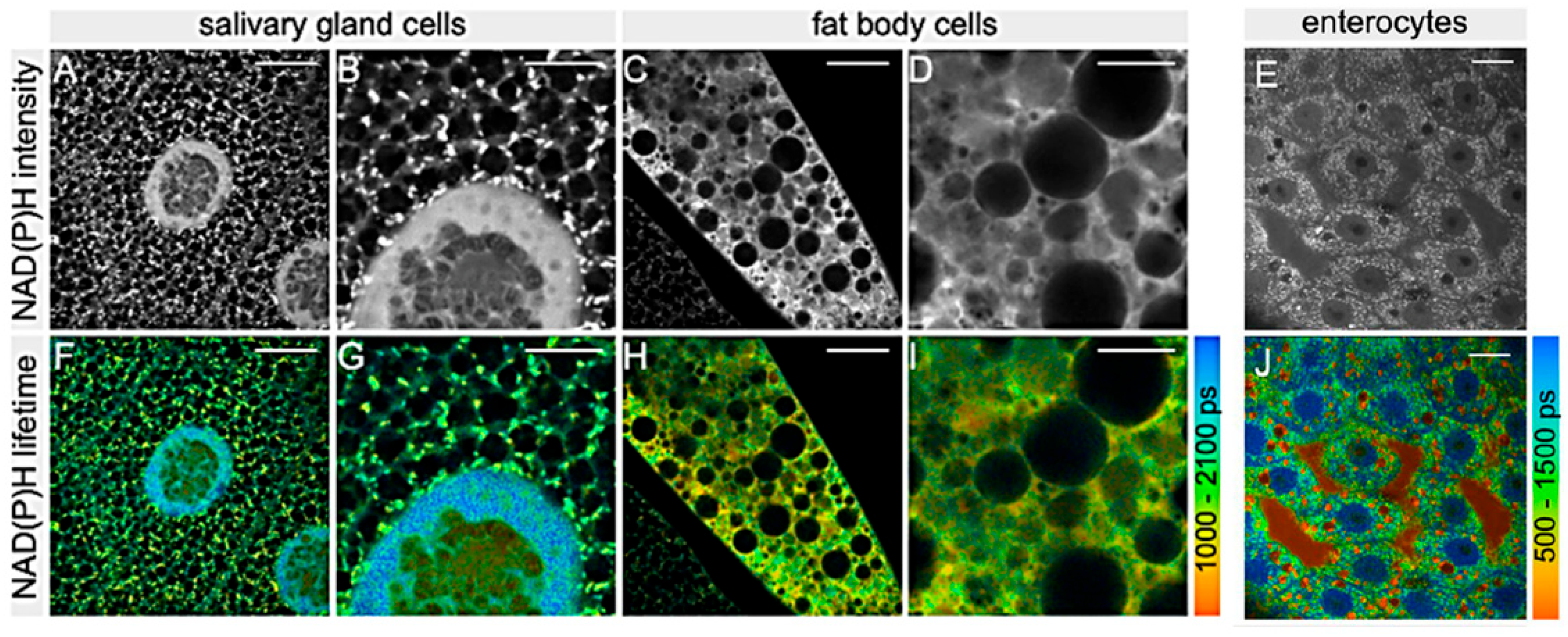
2.3. Metabolism
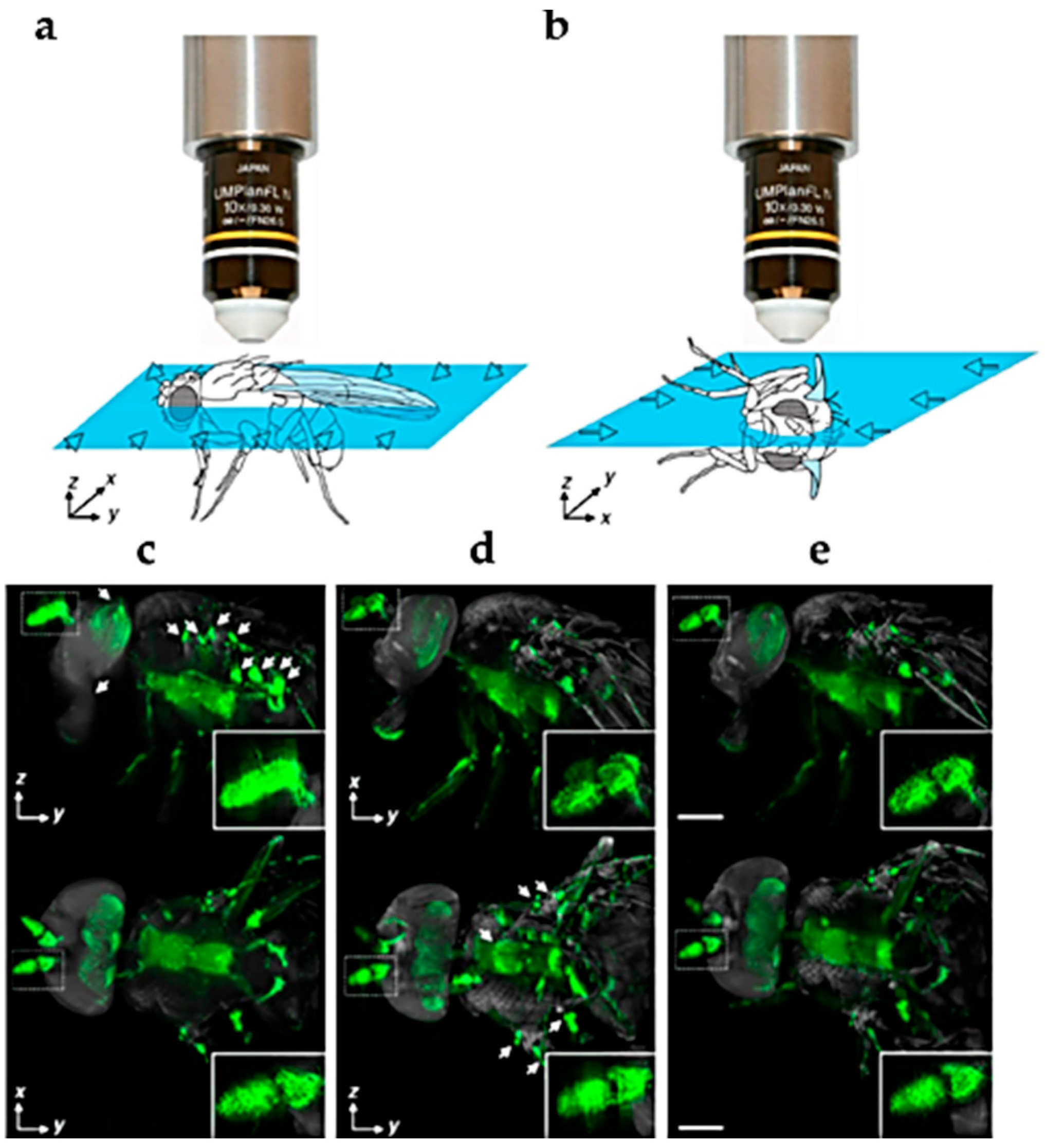
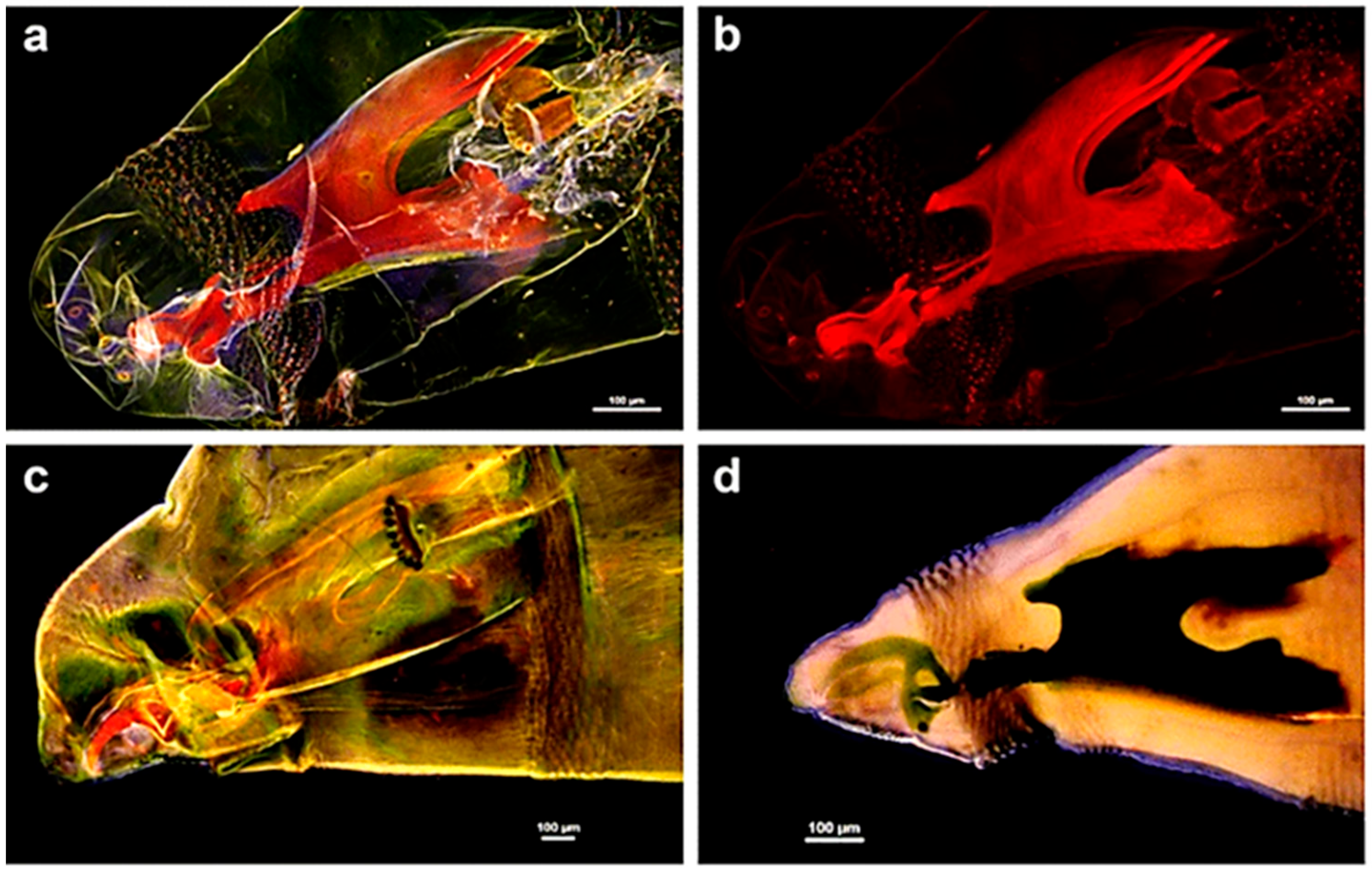
2.4. High Resolution Morphology
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Udenfriend, S. Fluorescence Assay in Biology and Medicine; Academic Press: New York, NY, USA, 1969. [Google Scholar]
- Croce, A.C. Light and autofluorescence, multitasking features in living organisms. Photochem 2021, 1, 67–124. [Google Scholar] [CrossRef]
- Dmitriev, R.I.; Intes, X.; Barroso, M.M. Luminescence lifetime imaging of three-dimensional biological objects. J. Cell Sci. 2021, 134, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Alfonso-Garcia, A.; Bec, J.; Weyers, B.; Marsden, M.; Zhou, X.; Li, C.; Marcu, L. Mesoscopic fluorescence lifetime imaging: Fundamental principles, clinical applications and future directions. J. Biophotonics 2021, 14, e202000472. [Google Scholar] [CrossRef] [PubMed]
- Torrado, B.; Malacrida, L.; Ranjit, S. Linear combination properties of the phasor space in fluorescence imaging. Sensors 2022, 22, 999. [Google Scholar] [CrossRef]
- Umebachi, Y.; Yoshida, K. Some chemical and physical properties of papiliochrome II in the wings of Papilio xuthus. J. Insect Physiol. 1970, 16, 1203–1228. [Google Scholar] [CrossRef]
- Vigneron, J.P.; Kertész, K.; Vértesy, Z.; Rassart, M.; Lousse, V.; Bálint, Z.; Biró, L.P. Correlated diffraction and fluorescence in the backscattering iridescence of the male butterfly Troides magellanus (Papilionidae). Phys. Rev. E 2008, 78, 021903. [Google Scholar] [CrossRef]
- Zobl, S.; Wilts, B.D.; Salvenmoser, W.; Pölt, P.; Gebeshuber, I.C.; Schwerte, T. Orientation-dependent reflection of structurally coloured butterflies. Biomimetics 2020, 5, 5. [Google Scholar] [CrossRef]
- Pirih, P.; Wilts, B.D.; Stavenga, D.G. Spatial reflection patterns of iridescent wings of male pierid butterflies: Curved scales reflect at a wider angle than flat scales. J. Comp. Physiol. A 2011, 197, 987–997. [Google Scholar] [CrossRef]
- Dechkrong, P.; Jiwajinda, S.; Dokchan, P.; Kongtungmon, M.; Chomsaeng, N.; Chairuangsri, T.; Yu, C.-C.; Hsiao, C.-N.; Shiojiri, M. Fine structure of wing scales of butterflies, Euploea mulciber and Troides aeacus. J. Struct. Biol. 2011, 176, 75–82. [Google Scholar] [CrossRef]
- Wilts, B.D.; Trzeciak, T.M.; Vukusic, P.; Stavenga, D.G. Papiliochrome II pigment reduces the angle dependency of structural wing colouration in nireus group papilionids. J. Exp. Biol. 2012, 215, 796–805. [Google Scholar] [CrossRef]
- Wilts, B.D.; Ijbema, N.; Stavenga, D.G. Pigmentary and photonic coloration mechanisms reveal taxonomic relationships of the Cattlehearts (Lepidoptera: Papilionidae: Parides). BMC Evol. Biol. 2014, 14, 160. [Google Scholar] [CrossRef] [PubMed]
- Wilts, B.D.; Matsushita, A.; Arikawa, K.; Stavenga, D.G. Spectrally tuned structural and pigmentary coloration of birdwing butterfly wing scales. J. R. Soc. Interface 2015, 12, 20150717. [Google Scholar] [CrossRef] [PubMed]
- Thayer, R.C.; Allen, F.I.; Patel, N.H. Structural color in Junonia butterflies evolves by tuning scale lamina thickness. Elife 2020, 9, e52187. [Google Scholar] [CrossRef]
- Kumazawa, K.; Tabata, H. A three-dimensional fluorescence analysis of the wings of male Morpho sulkowskyi and Papilio xuthus butterflies. Zool. Sci. 2001, 18, 1073–1079. [Google Scholar] [CrossRef]
- Osotsi, M.I.; Zhang, W.; Zada, I.; Gu, J.; Liu, Q.; Zhang, D. Butterfly wing architectures inspire sensor and energy applications. Natl. Sci. Rev. 2021, 8, 2021. [Google Scholar] [CrossRef]
- Mouchet, S.R.; Van Hooijdonk, E.; Welch, V.L.; Louette, P.; Colomer, J.-F.; Su, B.-L.; Deparis, O. Liquid-induced colour change in a beetle: The concept of a photonic cell. Sci. Rep. 2016, 6, 19322. [Google Scholar] [CrossRef] [PubMed]
- Mouchet, S.R.; Tabarrant, T.; Lucas, S.; Su, B.-L.; Vukusic, P.; Deparis, O.; Seago, A.E.; Brady, P.; Vigneron, J.; Schultz, T.D. Vapor sensing with a natural photonic cell. Opt. Express 2016, 24, 12267–12280. [Google Scholar] [CrossRef]
- Mouchet, S.R.; Verstraete, C.; Mara, D.; Van Cleuvenbergen, S.; Finlayson, E.D.; Van Deun, R.; Deparis, O.; Verbiest, T.; Maes, B.; Vukusic, P.; et al. Nonlinear optical spectroscopy and two-photon excited fluorescence spectroscopy reveal the excited states of fluorophores embedded in a beetle’s elytra. Interface Focus 2019, 9, 20180052. [Google Scholar] [CrossRef]
- Nansen, C.; Ribeiro, L.P.; Dadour, I.; Roberts, J.D. Detection of temporal changes in insect body reflectance in response to killing agents. PLoS ONE 2015, 10, e0124866. [Google Scholar] [CrossRef] [PubMed]
- Michels, J.; Gorb, S.N. Detailed three-dimensional visualization of resilin in the exoskeleton of arthropods using confocal laser scanning microscopy. J. Microsc. 2012, 245, 1–16. [Google Scholar] [CrossRef]
- Michels, J.; Appel, E.; Gorb, S.N. Functional diversity of resilin in Arthropoda. Beilstein J. Nanotechnol. 2016, 7, 1241–1259. [Google Scholar] [CrossRef]
- Bäumler, F.; Büsse, S. Resilin in the flight apparatus of Odonata (Insecta)-Cap tendons and their biomechanical importance for flight. Biol. Lett. 2019, 15, 20190127. [Google Scholar] [CrossRef] [PubMed]
- Hatch, G. Structure and mechanics of the dragonfly pterothorax. Ann. Entomol. Soc. Am. 1966, 59, 702–714. [Google Scholar] [CrossRef]
- Hai, T.A.P.; Sugimoto, R. Fluorescence control of chitin and chitosan fabricated via surface functionalization using direct oxidative polymerization. RSC Adv. 2018, 8, 7005–7013. [Google Scholar]
- Rabasović, M.D.; Pantelić, D.V.; Jelenković, B.M.; Ćurčić, S.B.; Rabasović, M.S.; Vrbica, M.D.; Lazović, V.M.; Ćurčić, B.P.M.; Krmpot, A.J. Nonlinear microscopy of chitin and chitinous structures: A case study of two cave-dwelling insects. J. Biomed. Opt. 2015, 20, 016010. [Google Scholar] [CrossRef]
- Reinhardt, K.; Breunig, H.G.; König, K. Autofluorescence lifetime variation in the cuticle of the bedbug Cimex lectularius. Arthropod Struct. Dev. 2017, 46, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Eshghi, S.H.; Jafarpour, M.; Darvizeh, A.; Gorb, S.N.; Rajabi, H. A simple, high-resolution, non-destructive method for determining the spatial gradient of the elastic modulus of insect cuticle. J. R. Soc. Interface 2018, 15, 20180312. [Google Scholar] [CrossRef]
- Pentzold, S.; Marion-Poll, F.; Grabe, V.; Burse, A. Autofluorescence-based identification and functional validation of antennal gustatory sensilla in a specialist leaf beetle. Front. Physiol. 2019, 10, 343. [Google Scholar] [CrossRef]
- Pende, M.; Becker, K.; Wanis, M.; Saghafi, S.; Kaur, R.; Hahn, C.; Pende, N.; Foroughipour, M.; Hummel, T.; Dodt, H.-U. High-resolution ultramicroscopy of the developing and adult nervous system in optically cleared Drosophila melanogaster. Nat. Commun. 2018, 9, 4731. [Google Scholar] [CrossRef]
- Caroti, F.; Urbansky, S.; Wosch, M.; Lemke, S. Germ line transformation and in vivo labeling of nuclei in Diptera: Report on Megaselia abdita (Phoridae) and Chironomus riparius (Chironomidae). Dev. Genes Evol. 2015, 225, 179–186. [Google Scholar] [CrossRef]
- Fukatsu, T.; Watanabe, K.; Sekiguchi, Y. Specific detection of intracellular symbiotic bacteria of aphids by oligonucleotide-probed in situ hybridization. Appl. Entomol. Zool. 1998, 33, 461–472. [Google Scholar] [CrossRef]
- Thimm, T.; Tebbe, C.C. Protocol for rapid fluorescence in situ hybridization of bacteria in cryosections of microarthropods. Appl. Environ. Microbiol. 2003, 69, 2875–2878. [Google Scholar] [CrossRef] [PubMed]
- Handler, A.M. A current perspective on insect gene transformation. Insect Biochem. Mol. Biol. 2001, 31, 111–128. [Google Scholar] [CrossRef]
- Horn, C.; Schmid, B.G.M.; Pogoda, F.S.; Wimmer, E.A. Fluorescent transformation markers for insect transgenesis. Insect Biochem. Mol. Biol. 2002, 32, 1221–1235. [Google Scholar] [CrossRef]
- De Niz, M.; Kehrer, J.; Brancucci, N.M.B.B.; Moalli, F.; Reynaud, E.G.; Stein, J.V.; Frischknecht, F. 3D imaging of undissected optically cleared Anopheles stephensi mosquitoes and midguts infected with Plasmodium parasites. PLoS ONE 2020, 15, e0238134. [Google Scholar]
- Koga, R.; Tsuchida, T.; Fukatsu, T. Quenching autofluorescence of insect tissues for in situ detection of endosymbionts. Appl. Entomol. Zool. 2009, 44, 281–291. [Google Scholar] [CrossRef]
- Yeh, E.; Gustafson, K.; Bouliannet, G.L. Green fluorescent protein as a vital marker and reporter of gene expression in Drosophila. Proc. Natl. Acad. Sci. USA 1995, 92, 7036–7040. [Google Scholar] [CrossRef] [PubMed]
- Santhosh, P.; Yuhua, D.; Weidong, G.; Xiaoguang, C. Heterologous expression in transgenic mosquitoes. Asian Pac. J. Trop. Med. 2010, 3, 244–250. [Google Scholar] [CrossRef]
- Yoshida, S.; Watanabe, H. Robust salivary gland-specific transgene expression in Anopheles stephensi mosquito. Insect Mol. Biol. 2006, 15, 403–410. [Google Scholar] [CrossRef]
- Scolari, F.; Schetelig, M.F.; Bertin, S.; Malacrida, A.R.; Gasperi, G.; Wimmer, E.A. Fluorescent sperm marking to improve the fight against the pest insect Ceratitis capitata (Wiedemann; Diptera: Tephritidae). N. Biotechnol. 2008, 25, 76–84. [Google Scholar] [CrossRef]
- Barolo, S.; Castro, B.; Posakony, J.W. New Drosophila transgenic reporters: Insulated P-element vectors expressing fast-maturing RFP. Biotechniques 2004, 36, 436–442. [Google Scholar] [CrossRef]
- Warren, I.A.; Fowler, K.; Smith, H. Germline transformation of the stalk-eyed fly, Teleopsis dalmanni. BMC Mol. Biol. 2010, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Mavrakis, M.; Rikhy, R.; Lilly, M.; Lippincott-Schwartz, J. Fluorescence imaging techniques for studying Drosophila embryo development. Curr. Protoc. Cell Biol. 2008, 39, 4–18. [Google Scholar]
- Mangabeira, O.; Sherlock, I.A. Fluorescent tracer and natural fluorescence in Diptera of the genus Phlebotomus. Rev. Bras. Biol. 1963, 23, 181–187. [Google Scholar]
- Schawaroch, V.; Grimaldi, D.; Klaus, A.V. Focusing on morphology: Applications and implications of confocal laser scanning microscopy (Diptera: Campichoetidae, Camillidae, Drosophilidae). Proc. Entomol. Soc. Wash. 2005, 107, 323–335. [Google Scholar]
- Büscher, T.H.; Petersen, D.S.; Bijma, N.N.; Bäumler, F.; Pirk, C.W.W.; Büsse, S.; Heepe, L.; Gorb, S.N. The exceptional attachment ability of the ectoparasitic bee louse Braula coeca (Diptera, Braulidae) on the honeybee. Physiol. Entomol. 2022, 47, 83–95. [Google Scholar] [CrossRef]
- Petersen, D.S.; Kreuter, N.; Heepe, L.; Büsse, S.; Wellbrock, A.H.J.; Witte, K.; Gorb, S.N. Holding tight on feathers—Structural specializations and attachment properties of the avian ectoparasite Crataerina pallida (Diptera, Hippoboscidae). J. Exp. Biol. 2018, 221, jeb179242. [Google Scholar] [CrossRef]
- Lerch, S.; Zuber, R.; Gehring, N.; Wang, Y.; Eckel, B.; Klass, K.-D.; Lehmann, F.-O.; Moussian, B. Resilin matrix distribution, variability and function in Drosophila. BMC Biol. 2020, 18, 195. [Google Scholar] [CrossRef]
- Curtin, T.J.; Jones, J.C. The mechanism of ovulation and oviposition in Aedes aegypti. Ann. Entomol. Soc. Am. 1961, 54, 298–313. [Google Scholar] [CrossRef]
- Clements, A.N.; Potter, S.A. The fine structure of the spermathecae and their ducts in the mosquito Aedes aegypti. J. Insect Physiol. 1967, 13, 1825–1836. [Google Scholar] [CrossRef]
- Giglioli, M.E. The female reproductive system of Anopheles gambiae melas. I. The Structure and Function of the Genital Ducts and Associated Organs. Riv. Malariol. 1963, 42, 149–176. [Google Scholar]
- Pascini, T.V.; Ramalho-Ortigão, M.; Martins, G.F. Morphological and morphometrical assessment of spermathecae of Aedes aegypti females. Mem. Inst. Oswaldo Cruz. 2012, 107, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Lerch, S.; Yang, Y.; Flaven-Pouchon, J.; Gehring, N.; Moussian, B. Resilin is needed for wing posture in Drosophila suzukii. Arch. Insect Biochem. Physiol. 2022, e21913. [Google Scholar] [CrossRef] [PubMed]
- Scolari, F.; Valerio, F.; Benelli, G.; Papadopoulos, N.T.; Vaníčková, L. Tephritid fruit fly semiochemicals: Current knowledge and future perspectives. Insects 2021, 12, 408. [Google Scholar] [CrossRef] [PubMed]
- Coutinho-Abreu, I.V.; Riffell, J.A.; Akbari, O.S. Human attractive cues and mosquito host-seeking behavior. Trends Parasitol. 2022, 38, 246–264. [Google Scholar] [CrossRef]
- Konopka, J.K.; Task, D.; Afify, A.; Raji, J.; Deibel, K.; Maguire, S.; Lawrence, R.; Potter, C.J. Olfaction in Anopheles mosquitoes. Chem. Senses 2021, 46, bjab021. [Google Scholar] [CrossRef]
- Baik, L.S.; Carlson, J.R. The mosquito taste system and disease control. Proc. Natl. Acad. Sci. USA 2020, 117, 32848–32856. [Google Scholar] [CrossRef]
- Hehlert, P.; Zhang, W.; Göpfert, M.C. Drosophila mechanosensory transduction. Trends Neurosci. 2021, 44, 323–335. [Google Scholar] [CrossRef]
- Schnaitmann, C.; Pagni, M.; Reiff, D.F. Color vision in insects: Insights from Drosophila. J. Comp. Physiol. A 2020, 206, 183–198. [Google Scholar] [CrossRef]
- Rings, A.; Goodwin, S.F. To court or not to court-A multimodal sensory decision in Drosophila males. Curr. Opin. Insect Sci. 2019, 35, 48–53. [Google Scholar] [CrossRef]
- Kannan, K.; Galizia, C.G.; Nouvian, M. Olfactory strategies in the defensive behaviour of insects. Insects 2022, 13, 470. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Z.; Anushree; Bilgrami, A.L.; Ahsan, J. Drosophila melanogaster chemosensory pathways as potential targets to curb the insect menace. Insects 2022, 13, 142. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, M.; Hefetz, A. Neural mechanisms and information processing in recognition systems. Insects 2014, 5, 722–741. [Google Scholar] [CrossRef] [PubMed]
- Croce, A.C.; Scolari, F. The bright side of the tiger: Autofluorescence patterns in Aedes albopictus (Diptera, Culicidae) male and female mosquitoes. Molecules 2022, 27, 713. [Google Scholar] [CrossRef] [PubMed]
- Scolari, F.; (IGM-CNR, Pavia, Italy). Personal communication, 2021.
- Valmalette, J.C.; Raad, H.; Qiu, N.; Ohara, S.; Capovilla, M.; Robichon, A. Nano-architecture of gustatory chemosensory bristles and trachea in Drosophila wings. Sci. Rep. 2015, 5, 14198. [Google Scholar] [CrossRef] [PubMed]
- Göpfert, M.C.; Hennig, R.M. Hearing in Insects. Annu. Rev. Entomol. 2016, 61, 257–276. [Google Scholar] [CrossRef]
- Saltin, B.D.; Matsumura, Y.; Reid, A.; Windmill, J.F.; Gorb, S.N.; Jackson, J.C. Resilin distribution and sexual dimorphism in the midge antenna and their influence on frequency sensitivity. Insects 2020, 11, 520. [Google Scholar] [CrossRef]
- Saltin, B.D.; Matsumura, Y.; Reid, A.; Windmill, J.F.; Gorb, S.N.; Jackson, J.C. Material stiffness variation in mosquito antennae. J. R. Soc. Interface 2019, 16, 20190049. [Google Scholar] [CrossRef]
- Figon, F.; Casas, J. Ommochromes in invertebrates: Biochemistry and cell biology. Biol. Rev. 2019, 94, 156–183. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Stavenga, D.G.; Arikawa, K.; Belušič, G.; Kelber, A. Evolution of insect color vision: From spectral sensitivity to visual ecology. Annu. Rev. Entomol. 2021, 66, 435–461. [Google Scholar] [CrossRef]
- Sharkey, C.R.; Blanco, J.; Leibowitz, M.M.; Pinto-Benito, D.; Wardill, T.J. The spectral sensitivity of Drosophila photoreceptors. Sci. Rep. 2020, 10, 18242. [Google Scholar] [CrossRef]
- Peach, D.A.H.; Ko, E.; Blake, A.J.; Gries, G. Ultraviolet inflorescence cues enhance attractiveness of inflorescence odour to Culex pipiens mosquitoes. PLoS ONE 2019, 14, e0217484. [Google Scholar] [CrossRef] [PubMed]
- Ebadi, H.; Perry, M.; Short, K.; Klemm, K.; Desplan, C.; Stadler, P.F.; Mehta, A. Patterning the insect eye: From stochastic to deterministic mechanisms. PLoS Comput. Biol. 2018, 14, e1006363. [Google Scholar] [CrossRef]
- Dontsov, A.E.; Sakina, N.L.; Yakovleva, M.A.; Bastrakov, A.I.; Bastrakova, I.G.; Zagorinsky, A.A.; Ushakova, N.A.; Feldman, T.B.; Ostrovsky, M.A. Ommochromes from the compound eyes of insects: Physicochemical properties and antioxidant activity. Biochemistry 2020, 85, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Feord, R.C.; Wardill, T.J. A novel setup for simultaneous two-photon functional imaging and precise spectral and spatial visual stimulation in Drosophila. Sci. Rep. 2020, 10, 15681. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, N. In vivo fluorescence of the rhabdomeres in an insect eye. In Proceedings of the Int Union Physiol Soc XIII, XXVIIth Int, Congr, Paris, France, 1977; p. 237. [Google Scholar]
- Franceschini, N.; Kirschfeld, K.; Minke, B. Fluorescence of photoreceptor cells observed in vivo. Science 1981, 213, 1264–1267. [Google Scholar] [CrossRef] [PubMed]
- Minke, B.; Hardie, R.C. Chapter 9 Genetic dissection of Drosophila phototransduction. In Handbook of Biological Physics; Agrotech Press: Noord-Holland, The Netherlands, 2000; Volume 3, pp. 449–525. [Google Scholar]
- Stark, W.S.; Frayer, K.L.; Johnson, M.A. Photopigment and receptor properties in Drosophila compound eye and ocellar receptors. Biophys. Struct. Mech. 1979, 5, 197–209. [Google Scholar] [CrossRef]
- Stavenga, D.G.; de Grip, W.J. Progress in phototransduction. Biophys. Struct. Mech. 1983, 9, 225–230. [Google Scholar] [CrossRef]
- Stavenga, D.G.; Hardie, R.C. Metarhodopsin control by arrestin, light-filtering screening pigments, and visual pigment turnover in invertebrate microvillar photoreceptors. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2011, 197, 227–241. [Google Scholar] [CrossRef]
- Stavenga, D.G. Colour in the eyes of insects. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 2002, 188, 337–348. [Google Scholar]
- Stark, W.S.; Thomas, C.F. Microscopy of multiple visual receptor types in Drosophila. Mol. Vis. 2004, 10, 943–955. [Google Scholar] [PubMed]
- Lee, R.D.; Thomas, C.F.; Marietta, R.G.; Stark, W.S. Vitamin A, visual pigments, and visual receptors in Drosophila. Microsc. Res. Tech. 1996, 35, 418–430. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Yoshida, H. Drosophila as a Model Organism. Adv. Exp. Med. Biol. 2018, 1076, 1–10. [Google Scholar] [PubMed]
- Hardie, R.C.; Juusola, M. Phototransduction in Drosophila. Curr. Opin. Neurobiol. 2015, 34, 37–45. [Google Scholar] [CrossRef]
- Ortega, J.T.; Jastrzebska, B. The retinoid and non-retinoid ligands of the rod visual G protein-coupled receptor. Int. J. Mol. Sci. 2019, 20, 6218. [Google Scholar] [CrossRef] [PubMed]
- Stavenga, D.G. On visual pigment templates and the spectral shape of invertebrate rhodopsins and metarhodopsins. J. Comp. Physiol. A 2010, 196, 869–878. [Google Scholar] [CrossRef]
- Pichaud, F.; Desplan, C. A new visualization approach for identifying mutations that affect differentiation and organization of the Drosophila ommatidia. Development 2001, 128, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Wolff, T.; Ready, D.F. Pattern formation in the Drosophila retina. In The Development of Drosophila melanogaster; Bate, M., Ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1993; pp. 1277–1325. [Google Scholar]
- Belušič, G.; Pirih, P.; Stavenga, D.G. Photoreceptor responses of fruitflies with normal and reduced arrestin content studied by simultaneous measurements of visual pigment fluorescence and ERG. J. Comp. Physiol. A 2010, 196, 23. [Google Scholar] [CrossRef]
- Stowasser, A.; Owens, M.; Buschbeck, E.K. Giving invertebrates an eye exam: An ophthalmoscope that utilizes the autofluorescence of photoreceptors. J. Exp. Biol. 2017, 220, 4095–4100. [Google Scholar] [CrossRef]
- Alonso San Alberto, D.; Rusch, C.; Zhan, Y.; Straw, A.D.; Montell, C.; Riffell, J.A. The olfactory gating of visual preferences to human skin and visible spectra in mosquitoes. Nat. Commun. 2022, 13, 555. [Google Scholar] [CrossRef]
- Pimnon, S.; Ngoen-Klan, R.; Sumarnrote, A.; Chareonviriyaphap, T. UV light-emitting-diode traps for collecting nocturnal biting mosquitoes in urban Bangkok. Insects 2022, 13, 526. [Google Scholar] [CrossRef]
- Rebora, M.; Salerno, G.; Piersanti, S.; Kovalev, A.; Gorb, S. Cuticular modified air sacs underlie white coloration in the olive fruit fly, Bactrocera oleae. Commun. Biol. 2021, 4, 881. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, W.G. Sexual behavior and sexual selection in the Mediterranean fruit fly, Ceratitis capitata (Dacinae: Ceratitidini). In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A.L., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 459–489. [Google Scholar]
- Sivinski, J.; Klug, H.; Shapiro, J.; Lane, J.; Mankin, R.W. Ultraviolet reflectance on the heads and wings of Anastrepha suspensa (Loew) and Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Stud. Dipterol. 2004, 11, 313–322. [Google Scholar]
- Croce, A.C.; Bottiroli, G. Autofluorescence spectroscopy and imaging: A tool for biomedical research and diagnosis. Eur. J. Histochem. 2014, 58, 320–337. [Google Scholar] [CrossRef]
- Schneckenburger, H.; Wagner, M.; Weber, P.; Strauss, W.S.L.; Sailer, R. Autofluorescence lifetime imaging of cultivated cells using a UV picosecond laser diode. J. Fluoresc. 2004, 14, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Wetzker, C.; Reinhardt, K. Distinct metabolic profiles in Drosophila sperm and somatic tissues revealed by two-photon NAD(P)H and FAD autofluorescence lifetime imaging. Sci. Rep. 2019, 9, 19534. [Google Scholar] [CrossRef]
- MacLeod, J. The metabolism of human spermatozoa. Am. J. Physiol. Content 1941, 132, 193–201. [Google Scholar] [CrossRef]
- Storey, B.T. Mammalian sperm metabolism: Oxygen and sugar, friend and foe. Int. J. Dev. Biol. 2008, 52, 427–437. [Google Scholar] [CrossRef]
- Perotti, M.E. The mitochondrial derivative of the spermatozoon of Drosophila before and after fertilization. J. Ultrastruct. Res. 1973, 44, 181–198. [Google Scholar] [CrossRef]
- Prokupek, A.M.; Eyun, S.-I.; Ko, L.; Moriyama, E.N.; Harshman, L.G. Molecular evolutionary analysis of seminal receptacle sperm storage organ genes of Drosophila melanogaster. J. Evol. Biol. 2010, 23, 1386–1398. [Google Scholar] [CrossRef]
- Reinhardt, K.; Breunig, H.G.; Uchugonova, A.; König, K. Sperm metabolism is altered during storage by female insects: Evidence from two-photon autofluorescence lifetime measurements in bedbugs. J. R. Soc. Interface 2015, 12, 20150609. [Google Scholar] [CrossRef] [PubMed]
- Massino, C.; Wetzker, C.; Balvin, O.; Bartonicka, T.; Kremenova, J.; Sasinkova, M.; Otti, O.; Reinhardt, K. Seminal fluid and sperm diluent affect sperm metabolism in an insect: Evidence from NAD(P)H and flavin adenine dinucleotide autofluorescence lifetime imaging. Microsc. Res. Tech. 2022, 85, 398–411. [Google Scholar] [CrossRef] [PubMed]
- Turnell, B.R.; Reinhardt, K. Metabolic rate and oxygen radical levels increase but radical generation rate decreases with male age in Drosophila melanogaster sperm. J. Gerontol. Ser. A 2020, 75, 2278–2285. [Google Scholar] [CrossRef]
- Ogilvie, J.P.; Débarre, D.; Solinas, X.; Martin, J.-L.; Beaurepaire, E.; Joffre, M. Use of coherent control for selective two-photon fluorescence microscopy in live organisms. Opt. Express 2006, 14, 759. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Norton, A.; Kissel, M.; Azucena, O.; Kubby, J. Adaptive optical two-photon microscopy using autofluorescent guide stars. Opt. Lett. 2013, 38, 5075. [Google Scholar] [CrossRef]
- Chien, C.-H.; Chen, W.-W.; Wu, J.-T.; Chang, T.-C. Label-free imaging of Drosophila in vivo by coherent anti-Stokes Raman scattering and two-photon excitation autofluorescence microscopy. J. Biomed. Opt. 2011, 16, 016012. [Google Scholar] [CrossRef]
- Rizki, T.M. Genetic control of cytodifferentiation. J. Cell Biol. 1963, 16, 513–520. [Google Scholar] [CrossRef]
- Rizki, M.T.M. Intracellular localization of kynurinine in the fatbody of Drosophila. J. Biophys. Biochem. Cytol. 1961, 9, 567–572. [Google Scholar] [CrossRef]
- Rizki, T.M.; Rizki, R.M. An inducible enzyme system in the larval cells of Drosophila melanogaster. J. Cell Biol. 1963, 17, 87–92. [Google Scholar] [CrossRef]
- Garay, E.; Schuth, N.; Barbanente, A.; Tejeda-Guzmán, C.; Vitone, D.; Osorio, B.; Clark, A.H.; Nachtegaal, M.; Haumann, M.; Dau, H.; et al. Tryptophan regulates Drosophila zinc stores. Proc. Natl. Acad. Sci. USA 2022, 119, e2117807119. [Google Scholar] [CrossRef]
- Young, R.G.; Tappel, A.L. Fluorescent pigment and pentane production by lipid peroxidation in honey bees, Apis mellifera. Exp. Gerontol. 1978, 13, 457–459. [Google Scholar] [CrossRef]
- Münch, D.; Baker, N.; Rasmussen, E.M.K.; Shah, A.K.; Kreibich, C.D.; Heidem, L.E.; Amdam, G.V. Obtaining specimens with slowed, accelerated and reversed aging in the honey bee model. J. Vis. Exp. 2013, 78, 50550. [Google Scholar] [CrossRef] [PubMed]
- Chio, K.S.; Reiss, U.; Fletcher, B.; Tappel, A.L. Peroxidation of subcellular organelles: Formation of lipofuscinlike fluorescent pigments. Science 1969, 166, 1535–1536. [Google Scholar] [CrossRef] [PubMed]
- Wolman, M. Lipid pigments (chromolipids): Their origin, nature, and significance. Pathobiol. Annu. 1980, 10, 253–267. [Google Scholar]
- Jung, T.; Höhn, A.; Grune, T. Lipofuscin: Detection and quantification by microscopic techniques. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2010; Volume 594, pp. 173–219. [Google Scholar]
- Brunk, U.T.; Jones, C.B.; Sohal, R.S. A novel hypothesis of lipofuscinogenesis and cellular aging based on interactions between oxidative stress and autophagocytosis. Mutat. Res. 1992, 275, 395–403. [Google Scholar] [CrossRef]
- Sheldahl, J.A.; Tappel, A.L. Fluorescent products from aging Drosophila melanogaster: An indicator of free radical lipid peroxidation damage. Exp. Gerontol. 1974, 9, 33–41. [Google Scholar] [CrossRef]
- Miquel, J.; Tappel, A.L.; Dillard, C.J.; Herman, M.M.; Bensch, K.G. Fluorescent products and lysosomal components in aging Drosophila melanogaster. J. Gerontol. 1974, 29, 622–637. [Google Scholar] [CrossRef]
- Donato, H.; Sohal, R.S. Age-related changes in lipofuscin-associated fluorescent substances in the adult male housefly, Musca domestica. Exp. Gerontol. 1978, 13, 171–179. [Google Scholar] [CrossRef]
- Sohal, R.S.; Donato, H. Effects of experimentally altered life spans on the accumulation of fluorescent age pigment in the housefly, Musca domestica. Exp. Gerontol. 1978, 13, 335–341. [Google Scholar] [CrossRef]
- Ettershank, G.; Macdonnell, I.; Croft, R. The accumulation of age pigment by the fleshfly, Sarcophaga bullata Parker (Diptera: Sarcophagidae). Aust. J. Zool. 1983, 31, 131. [Google Scholar] [CrossRef]
- Ziegler, I.; Harmsen, R. The biology of pteridines in insects. In Advances in Insect Physiology; Academic Press: New York, NY, USA, 1970; Volume 6, pp. 139–203. [Google Scholar]
- Hopkins, F.G., III. The pigments of the Pieridae. A contribution to the study of excretory substances which function in ornament. Proc. R. Soc. Lond. 1895, 57, 5–6. [Google Scholar]
- Kim, H.; Kim, K.; Yim, J. Biosynthesis of drosopterins, the red eye pigments of Drosophila melanogaster. IUBMB Life 2013, 65, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Robson, S.K.A.; Vickers, M.; Blows, M.W.; Crozier, R.H. Age determination in individual wild-caught Drosophila serrata using pteridine concentration. J. Exp. Biol. 2006, 209, 3155–3163. [Google Scholar] [CrossRef] [PubMed]
- Patat, U. Über das pterinmuster der facettenaugen von Calliphora erythrocephala. Z. Vgl. Physiol. 1965, 51, 103–134. [Google Scholar] [CrossRef]
- Bernhardt, V.; Hannig, L.; Kinast, R.; Verhoff, M.A.; Rothweiler, F.; Zehner, R.; Amendt, J. Quantitative pteridine fluorescence analysis: A possible age-grading technique for the adult stages of the blow fly Calliphora vicina (Diptera: Calliphoridae). J. Insect Physiol. 2017, 98, 356–359. [Google Scholar] [CrossRef]
- Thomas, D.B.; Chen, A.C. Age determination in the adult screwworm (Diptera: Calliphoridae) by pteridine levels. J. Econ. Entomol. 1989, 82, 1140–1144. [Google Scholar] [CrossRef]
- Wall, R.; Langley, P.A.; Stevens, J.; Clarke, G.M. Age-determination in the old-world screw-worm fly Chrysomya bezziana by pteridine fluorescence. J. Insect Physiol. 1990, 36, 213–218. [Google Scholar] [CrossRef]
- Wall, R.; Langley, P.A.; Morgan, K.L. Ovarian development and pteridine accumulation for age determination in the blowfly Lucilia sericata. J. Insect Physiol. 1991, 37, 863–868. [Google Scholar] [CrossRef]
- Zhu, G.; Ye, G.; Hu, C. Determining the adult age of the Oriental latrine fly, Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae) by pteridine fluorescence analysis. Insect Sci. 2003, 10, 245–255. [Google Scholar] [CrossRef]
- Mail, T.S.; Chadwick, J.; Lehane, M.J. Determining the age of adults of Stomoxys calcitrans (L.) (Diptera: Muscidae). Bull. Entomol. Res. 1983, 73, 501–525. [Google Scholar] [CrossRef]
- McIntyre, G.S.; Gooding, R.H. Pteridine accumulation in Musca domestica. J. Insect Physiol. 1995, 41, 357–368. [Google Scholar] [CrossRef]
- Moon, R.D.; Krafsur, E.S. Pterin quantity and gonotrophic stage as indicators of age in Musca autumnalis (Diptera: Muscidae). J. Med. Entomol. 1995, 32, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Krafsur, E.S.; Rosales, A.L.; Robison-Cox, J.F.; Turner, J.P. Age structure of horn fly (Diptera: Muscidae) populations estimated by pterin concentrations. J. Med. Entomol. 1992, 29, 678–686. [Google Scholar] [CrossRef]
- Lehane, M.J.; Mail, T.S. Determining the age of adult male and female Glossina morsitans morsitans using a new technique. Ecol. Entomol. 1985, 10, 219–224. [Google Scholar] [CrossRef]
- Lehane, M.J.; Hargrove, J. Field experiments on a new method for determining age in tsetse flies (Diptera: Glossinidae). Ecol. Entomol. 1988, 13, 319–322. [Google Scholar] [CrossRef]
- Langley, P.A.; Hall, M.J.R.; Felton, T.; Ceesay, M. Determining the age of tsetse flies, Glossina spp. (Diptera: Glossinidae): An appraisal of the pteridine fluorescence technique. Bull. Entomol. Res. 1988, 78, 387–395. [Google Scholar] [CrossRef]
- Zhu, G.H.; Ye, G.Y.; Li, K.; Hu, C.; Xu, X.H. Determining the age of adult flesh flies, Boettcherisca peregrina, using pteridine fluorescence. Med. Vet. Entomol. 2013, 27, 59–63. [Google Scholar] [CrossRef]
- Mochizuki, A.; Shiga, M.; Imura, O. Pteridine accumulation for age determination in the Melon Fly, Bactrocera (Zeugodacus) cucurbitae (Coquillett)(Diptera: Tephritidae). Appl. Entomol. Zool. 1993, 28, 584–586. [Google Scholar] [CrossRef]
- Camin, V.; Baker, P.; Carey, J.; Valenzuela, J.; Arredondo-Peter, R. Biochemical age determination for adult Mediterranean fruit flies (Diptera: Tephritidae). J. Econ. Entomol. 1991, 84, 1283–1288. [Google Scholar] [CrossRef]
- Millest, A.L.; Cheke, R.A.; Howe, M.A.; Lehane, M.J.; Garms, R. Determining the ages of adult females of different members of the Simulium damnosum complex (Diptera: Simuliidae) by the pteridine accumulation method. Bull. Entomol. Res. 1992, 82, 219–226. [Google Scholar] [CrossRef]
- Wu, D.; Lehane, M.J. Pteridine fluorescence for age determination of Anopheles mosquitoes. Med. Vet. Entomol. 1999, 13, 48–52. [Google Scholar] [CrossRef]
- Lardeux, F.; Ung, A.; Chebret, M. Spectrofluorometers are not adequate for aging Aedes and Culex (Diptera: Culicidae) using pteridine fluorescence. J. Med. Entomol. 2000, 37, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Tower, J.; Agrawal, S.; Alagappan, M.P.; Bell, H.S.; Demeter, M.; Havanoor, N.; Hegde, V.S.; Jia, Y.; Kothawade, S.; Lin, X.; et al. Behavioral and molecular markers of death in Drosophila melanogaster. Exp. Gerontol. 2019, 126, 110707. [Google Scholar] [CrossRef]
- Dye, C. The analysis of parasite transmission by bloodsucking insects. Annu. Rev. Entomol. 1992, 37, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.J.; Hargrove, J.W. On the interpretation of age–prevalence curves for trypanosome infections of tsetse flies. Parasitology 1998, 116, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Bala, M.; Sharma, A. Review of some recent techniques of age determination of blow flies having forensic implications. Egypt. J. Forensic Sci. 2016, 6, 203–208. [Google Scholar] [CrossRef]
- Amendt, J.; Bugelli, V.; Bernhardt, V. Time flies—Age grading of adult flies for the estimation of the post-mortem interval. Diagnostics 2021, 11, 152. [Google Scholar] [CrossRef]
- Parodi, V.; Jacchetti, E.; Osellame, R.; Cerullo, G.; Polli, D.; Raimondi, M.T. Nonlinear optical microscopy: From fundamentals to applications in live bioimaging. Front. Bioeng. Biotechnol. 2020, 8, 1174. [Google Scholar] [CrossRef]
- Paoletti, M.E.; Haut, J.M.; Plaza, J.; Plaza, A. Deep learning classifiers for hyperspectral imaging: A review. ISPRS J. Photogramm. Remote Sens. 2019, 158, 279–317. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Hovhannisyan, V.; Wu, J.-T.; Lin, C.-W.; Chen, J.-H.; Lin, S.-J.; Dong, C.-Y. Label-free imaging of Drosophila larva by multiphoton autofluorescence and second harmonic generation microscopy. J. Biomed. Opt. 2008, 13, 050502. [Google Scholar] [CrossRef]
- Grzywacz, A.; Góral, T.; Szpila, K.; Hall, M.J.R. Confocal laser scanning microscopy as a valuable tool in Diptera larval morphology studies. Parasitol. Res. 2014, 113, 4297–4302. [Google Scholar] [CrossRef] [PubMed]
- McGurk, L.; Morrison, H.; Keegan, L.P.; Sharpe, J.; O’Connell, M.A. Three-dimensional Imaging of Drosophila melanogaster. PLoS ONE 2007, 2, e834. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.D.; Kaiser, K.; Müller, A.; Fischbach, K.-F.; Merchant, N.; Strausfeld, N.J. Flybrain, an on-line atlas and database of the Drosophila nervous system. Neuron 1995, 15, 17–20. [Google Scholar] [CrossRef]
- Heisenberg, M.; Böhl, K. Isolation of anatomical brain mutants of Drosophila by histological means. Z. Nat. 1979, 34, 143–147. [Google Scholar] [CrossRef]
- Jährling, N.; Becker, K.; Schönbauer, C.; Schnorrer, F.; Dodt, H.U. Three-dimensional reconstruction and segmentation of intact Drosophila by ultramicroscopy. Front. Syst. Neurosci. 2010, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Becker, K.; Jährling, N.; Kramer, E.R.; Schnorrer, F.; Dodt, H.U. Ultramicroscopy: 3D reconstruction of large microscopical specimens. J. Biophotonics 2008, 1, 36–42. [Google Scholar] [CrossRef]
- Saghafi, S.; Becker, K.; Hahn, C.; Dodt, H.-U. 3D-ultramicroscopy utilizing aspheric optics. J. Biophotonics 2014, 7, 117–125. [Google Scholar] [CrossRef]
- Gualda, E.J. 3D volume rendering of invertebrates using light-sheet fluorescence microscopy. Photonics 2022, 9, 208. [Google Scholar] [CrossRef]
- Mensa, F.S.; Muzzi, M.; Spani, F.; Tromba, G.; Dullin, C.; Di Giulio, A. When the utility of micro-computed tomography collides with insect sample preparation: An entomologist user guide to solve post-processing issues and achieve optimal 3D models. Appl. Sci. 2022, 12, 769. [Google Scholar] [CrossRef]
- Warren, J.-A.; Ratnasekera, T.D.P.; Campbell, D.A.; Anderson, G.S. Spectral signatures of immature Lucilia sericata (Meigen) (Diptera: Calliphoridae). Insects 2017, 8, 34. [Google Scholar] [CrossRef]
- Voss, S.C.; Magni, P.; Dadour, I.; Nansen, C. Reflectance-based determination of age and species of blowfly puparia. Int. J. Leg. Med. 2017, 131, 263–274. [Google Scholar] [CrossRef]
- Pickering, C.L.; Hands, J.R.; Fullwood, L.M.; Smith, J.A.; Baker, M.J. Rapid discrimination of maggots utilising ATR-FTIR spectroscopy. Forensic Sci. Int. 2015, 249, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Apasrawirote, D.; Boonchai, P.; Muneesawang, P.; Nakhonkam, W.; Bunchu, N. Assessment of deep convolutional neural network models for species identification of forensically-important fly maggots based on images of posterior spiracles. Sci. Rep. 2022, 12, 4753. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, T.; Silva, F.F.; Mascarin, G.M.; Silva, C.B. Multispectral imaging for quality control of laboratory-reared Anastrepha fraterculus (Diptera: Tephritidae) pupae. J. Appl. Entomol. 2019, 143, 1072–1079. [Google Scholar] [CrossRef]
- Jahnke, S.M.; Gebbers, R.; Capella Zanotta, D.; Intress, J. Differences in optical properties of males and females pupae of Anastrepha fraterculus (Diptera: Tephritidae). Rev. Colomb. Entomol. 2021, 47. [Google Scholar] [CrossRef]
- Moran, Z.R.; Parker, A.G. Near infrared imaging as a method of studying tsetse fly (Diptera: Glossinidae) pupal development. J. Insect Sci. 2016, 16, 72. [Google Scholar] [CrossRef]
- Vreysen, M.J.B.; Robinson, A.S.; Hendrichs, J. Area-Wide Control of Insect Pests from Research to Field Implementation; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Lutrat, C.; Giesbrecht, D.; Marois, E.; Whyard, S.; Baldet, T.; Bouyer, J. Sex sorting for pest control: It’s raining men! Trends Parasitol. 2019, 35, 649–662. [Google Scholar] [CrossRef]
- Ferguson, H.M.; Killeen, G.F.; Michel, K.; Wirtz, R.A.; Benedict, M.Q.; Dowell, F.E.; Mayagaya, V.S. Non-destructive determination of age and species of Anopheles gambiae s.l. using near-infrared spectroscopy. Am. J. Trop. Med. Hyg. 2009, 81, 622–630. [Google Scholar]
- Ntamatungiro, A.J.; Mayagaya, V.S.; Rieben, S.; Moore, S.J.; Dowell, F.E.; Maia, M.F. The influence of physiological status on age prediction of Anopheles arabiensis using near infra-red spectroscopy. Parasit. Vectors 2013, 6, 298. [Google Scholar] [CrossRef][Green Version]
- Sikulu, M.; Killeen, G.F.; Hugo, L.E.; Ryan, P.A.; Dowell, K.M.; Wirtz, R.A.; Moore, S.J.; Dowell, F.E. Near-infrared spectroscopy as a complementary age grading and species identification tool for African malaria vectors. Parasit. Vectors 2010, 3, 49. [Google Scholar] [CrossRef]
- Mayagaya, V.; Ntamatungiro, A.; Moore, S.; Wirtz, R.; Dowell, F.; Maia, M. Evaluating preservation methods for identifying Anopheles gambiae s.s. and Anopheles arabiensis complex mosquitoes species using near infra-red spectroscopy. Parasit. Vectors 2015, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Sikulu, M.; Dowell, K.M.; Hugo, L.E.; Wirtz, R.A.; Michel, K.; Peiris, K.H.; Moore, S.; Killeen, G.F.; Dowell, F.E. Evaluating RNAlater® as a preservative for using near-infrared spectroscopy to predict Anopheles gambiae age and species. Malar. J. 2011, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Krajacich, B.J.; Meyers, J.I.; Alout, H.; Dabiré, R.K.; Dowell, F.E.; Foy, B.D. Analysis of near infrared spectra for age-grading of wild populations of Anopheles gambiae. Parasit. Vectors 2017, 10, 552. [Google Scholar] [CrossRef]
- Sikulu-Lord, M.T.; Devine, G.J.; Hugo, L.E.; Dowell, F.E. First report on the application of near-infrared spectroscopy to predict the age of Aedes albopictus Skuse. Sci. Rep. 2018, 8, 9590. [Google Scholar] [CrossRef] [PubMed]
- Milali, M.P.; Sikulu-Lord, M.T.; Kiware, S.S.; Dowell, F.E.; Corliss, G.F.; Povinelli, R.J. Age grading An. gambiae and An. arabiensis using near infrared spectra and artificial neural networks. PLoS ONE 2019, 14, e0209451. [Google Scholar] [CrossRef] [PubMed]
- Joy, T.; Chen, M.; Arnbrister, J.; Williamson, D.; Li, S.; Nair, S.; Brophy, M.; Garcia, V.M.; Walker, K.; Ernst, K.; et al. Assessing near-infrared spectroscopy (NIRS) for evaluation of Aedes aegypti population age structure. Insects 2022, 13, 360. [Google Scholar] [CrossRef] [PubMed]
- Westphal-Ferreira, B.; Navarro-Silva, M.A. Near-infrared spectroscopy and microstructure of the scales of Sabethes (Sabethes) albiprivus (Diptera: Culicidae). Rev. Bras. Entomol. 2017, 61, 91–98. [Google Scholar] [CrossRef][Green Version]
- Mwanga, E.P.; Mapua, S.A.; Siria, D.J.; Ngowo, H.S.; Nangacha, F.; Mgando, J.; Baldini, F.; González Jiménez, M.; Ferguson, H.M.; Wynne, K.; et al. Using mid-infrared spectroscopy and supervised machine-learning to identify vertebrate blood meals in the malaria vector, Anopheles arabiensis. Malar. J. 2019, 18, 187. [Google Scholar] [CrossRef]
- Bomfleur, B.; Kerp, H.; Imnek, Z. A composite fluorescence micrograph of a complete pinnule of Autunia conferta. In Fossile Ökosysteme; Elicki, O., Schneider, J.W., Eds.; Wissenschaftliche Mitteilungen des Instituts für Geologie der TU Bergakademie Freiberg: Freiberg, Germany, 2007; p. 17. [Google Scholar]
- Haug, J.T.; Haug, C.; Ehrlich, M. First fossil stomatopod larva (Arthropoda: Crustacea) and a new way of documenting Solnhofen fossils (Upper Jurassic, Southern Germany). Palaeodiversity 2008, 1, 103–109. [Google Scholar]
- Haug, J.T.; Haug, C.; Kutschera, V.; Mayer, G.; Maas, A.; Liebau, S.; Castellani, C.; Wolfram, U.; Clarkson, E.N.K.; Waloszek, D. Autofluorescence imaging, an excellent tool for comparative morphology. J. Microsc. 2011, 244, 259–272. [Google Scholar] [CrossRef]
- Baranov, V.A.; Schädel, M.; Haug, J.T. Fly palaeo-evo-devo: Immature stages of bibionomorphan dipterans in Baltic and Bitterfeld amber. PeerJ 2019, 7, e7843. [Google Scholar] [CrossRef]
- Andersen, S.O. Characterization of a new type of cross-linkage in resilin, a rubber-like protein. Biochim. Biophys. Acta 1963, 69, 249–262. [Google Scholar] [CrossRef]
- Su, R.S.C.; Kim, Y.; Liu, J.C. Resilin: Protein-based elastomeric biomaterials. Acta Biomater. 2014, 10, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Roberts, G.A.F. Chitin Chemistry; Macmillan Education UK: London, UK, 1992; ISBN 978-1-349-11547-1. [Google Scholar]
- Stavenga, D.G. Thin film and multilayer optics cause structural colors of many insects and birds. Mater. Today Proc. 2014, 1, 109–121. [Google Scholar] [CrossRef]
- Gebru, A.; Jansson, S.; Ignell, R.; Kirkeby, C.; Prangsma, J.C.; Brydegaard, M. Multiband modulation spectroscopy for the determination of sex and species of mosquitoes in flight. J. Biophotonics 2018, 11, e201800014. [Google Scholar] [CrossRef] [PubMed]
- Stavenga, D.G. Fluorescence of blowfly metarhodopsin. Biophys. Struct. Mech. 1983, 9, 309–317. [Google Scholar] [CrossRef][Green Version]
- Stavenga, D.G.; Wehling, M.F.; Belušič, G. Functional interplay of visual, sensitizing and screening pigments in the eyes of Drosophila and other red-eyed dipteran flies. J. Physiol. 2017, 595, 5481–5494. [Google Scholar] [CrossRef]
- Salmon, J.-M.; Kohen, E.; Viallet, P.; Hirschberg, J.G.; Wouters, A.W.; Kohen, C.; Thorell, B. Microspectrofluorometric approach to the study of free/bound NAD(P)H ratio as metabolic indicator in various cell types. Photochem. Photobiol. 1982, 36, 585–593. [Google Scholar] [CrossRef]
- Kunz, W.S. Spectral properties of fluorescent flavoproteins of isolated rat liver mitochondria. FEBS Lett. 1986, 195, 92–96. [Google Scholar] [CrossRef]
- Kashio, S.; Miura, M. Kynurenine metabolism in the fat body non-autonomously regulates imaginal disc repair in Drosophila. iScience 2020, 23, 101738. [Google Scholar] [CrossRef] [PubMed]
- Park, H.B.; Lam, Y.C.; Gaffney, J.P.; Weaver, J.C.; Krivoshik, S.R.; Hamchand, R.; Pieribone, V.; Gruber, D.F.; Crawford, J.M. Bright green biofluorescence in sharks derives from bromo-kynurenine metabolism. iScience 2019, 19, 1291–1336. [Google Scholar] [CrossRef] [PubMed]
- Uyeda, K.; Rabinowitz, J.C. Fluorescence properties of tetrahydrofolate and related compounds. Anal. Biochem. 1963, 6, 100–108. [Google Scholar] [CrossRef]
- Penilla, R.P.; Rodríguez, M.H.; López, A.D.; Viader-Salvadó, J.M.; Sánchez, C.N. Pteridine concentrations differ between insectary-reared and field-collected Anopheles albimanus mosquitoes of the same physiological age. Med. Vet. Entomol. 2002, 16, 225–234. [Google Scholar] [CrossRef]
- Armstrong, D.; Wilhelm, J.; Smid, F.; Elleder, M. Chromatography and spectrofluorometry of brain fluorophores in neuronal ceroid lipofuscinosis (NCL). Mech. Ageing Dev. 1992, 64, 293–302. [Google Scholar] [CrossRef]
- Sohal, R.S.; Brunk, U.T. Lipofuscin as an indicator of oxidative stress and aging. Adv. Exp. Med. Biol. 1990, 266, 17–29. [Google Scholar]
- Münch, D.; Kreibich, C.D.; Amdam, G.V. Aging and its modulation in a long-lived worker caste of the honey bee. J. Exp. Biol. 2013, 216, 1638–1649. [Google Scholar] [CrossRef] [PubMed]
- Rigon, L.; De Filippis, C.; Napoli, B.; Tomanin, R.; Orso, G. Exploiting the potential of Drosophila models in lysosomal storage disorders: Pathological mechanisms and drug discovery. Biomedicines 2021, 9, 268. [Google Scholar] [CrossRef]
- Sakalem, M.E.; De Sibio, M.T.; da Silva da Costa, F.A.; de Oliveira, M. Historical evolution of spheroids and organoids, and possibilities of use in life sciences and medicine. Biotechnol. J. 2021, 16, 2000463. [Google Scholar] [CrossRef]
- Rosner, M.; Reithofer, M.; Fink, D.; Hengstschläger, M. Human embryo models and drug discovery. Int. J. Mol. Sci. 2021, 22, 637. [Google Scholar] [CrossRef]
- Franco, N.H.; Sandøe, P.; Olsson, I.A.S. Researchers’ attitudes to the 3Rs—An upturned hierarchy? PLoS ONE 2018, 13, e0200895. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomolecules | Fluorophores | Functions | Absorption/ Excitation (nm) | Emission (nm) | Detection Methods | References |
|---|---|---|---|---|---|---|
| Resilin | Di-/tri-tyrosine residues, crosslinks and bridges | Structural protein/Biomechanical | 320–380 (violet-blue) | 400–500 (blue-green) | Spectroscopy/Wide-field, confocal microscopy imaging | [21,22,49,192,193] |
| Chitin and chitinous-mixed compounds | Undefined components | Polysaccharide semicrystalline biocomposite/Biomechanical | 450–560 (blue-green) | >510–640 (green-red) | Spectroscopy/Wide-field, confocal microscopy imaging | [21,22,25,26,194] |
| Optically active thin films | Scatter/Iridescence | Dual wavelength polarimetric spectroscopy/Reflectance, interference imaging | [195,196] | |||
| Rhodopsins | Metarhodopsin | Activated form of G-protein coupled receptor rhodopsin/Vision | 300–400; 400–650 (violet); (blue-red) | 600–750 (red-deep red) | Spectroscopy/Wide-field microscopy imaging | [90,91,93,197,198] |
| Pyridinic derivatives | NAD(P)H bound/free | Coenzymes in redox reactions in energy metabolism, reductive biosynthesis | 330–380 (violet) | 440bound; 480free (violet-blue); (blue) | Spectroscopy/Wide-field microscopy imaging, multiphoton lifetime microscopy imaging | [101,102,107,108,109,199,200] |
| Riboflavin derivative | FAD | 360/445 (violet)/(blue) | 480/540 (blue)/(green) | |||
| Intermediate metabolites of tryptophan | Kynurenine and derivatives | Nitrogen excretion pathway; light screening in eyes; aging markers | 300–390/380–420 (violet)/(violet-blue) | 430–550 (blue-yellow) | Spectroscopy/Wide-field microscopy imaging | [113,114,115,116,128,131,201,202,203,204] |
| Pterine derivatives | Pteridines | 355 (violet) | 400–550 (blue-yellow) | |||
| Peroxidized lipids, proteins, carotenoids and their mixed compounds | Lipofuscin-like lipopigments | Aging and oxidative stress markers | Near UV—~500 (violet-green) | >460 (blue-red) | Spectroscopy/Wide-field microscopy imaging | [118,120,124,125,205,206,207] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Croce, A.C.; Scolari, F. Autofluorescent Biomolecules in Diptera: From Structure to Metabolism and Behavior. Molecules 2022, 27, 4458. https://doi.org/10.3390/molecules27144458
Croce AC, Scolari F. Autofluorescent Biomolecules in Diptera: From Structure to Metabolism and Behavior. Molecules. 2022; 27(14):4458. https://doi.org/10.3390/molecules27144458
Chicago/Turabian StyleCroce, Anna C., and Francesca Scolari. 2022. "Autofluorescent Biomolecules in Diptera: From Structure to Metabolism and Behavior" Molecules 27, no. 14: 4458. https://doi.org/10.3390/molecules27144458
APA StyleCroce, A. C., & Scolari, F. (2022). Autofluorescent Biomolecules in Diptera: From Structure to Metabolism and Behavior. Molecules, 27(14), 4458. https://doi.org/10.3390/molecules27144458