
Is Glyceryl Trinitrate, a Nitric Oxide Donor Responsible for Ameliorating the Chemical-Induced Tissue Injury In Vivo?


Abstract
:1. Introduction
2. Results
2.1. Effect of Glyceryl Trinitrate (GTN) and NG-Nitroarginine Methyl Ester (l-NAME) on Hepatic GSH-Dependent Enzymes in Rats Treated with Thioacetamide (TAA)
2.2. Effect of Glyceryl Trinitrate (GTN) and NG-Nitroarginine Methyl Ester (l-NAME) on Serum Transaminases in Rats Treated with Thioacetamide (TAA)
2.3. Effect of GTN and l-NAME on Hepatic GSH and Lipid Peroxidation in Rats Treated with Thioacetamide (TAA)
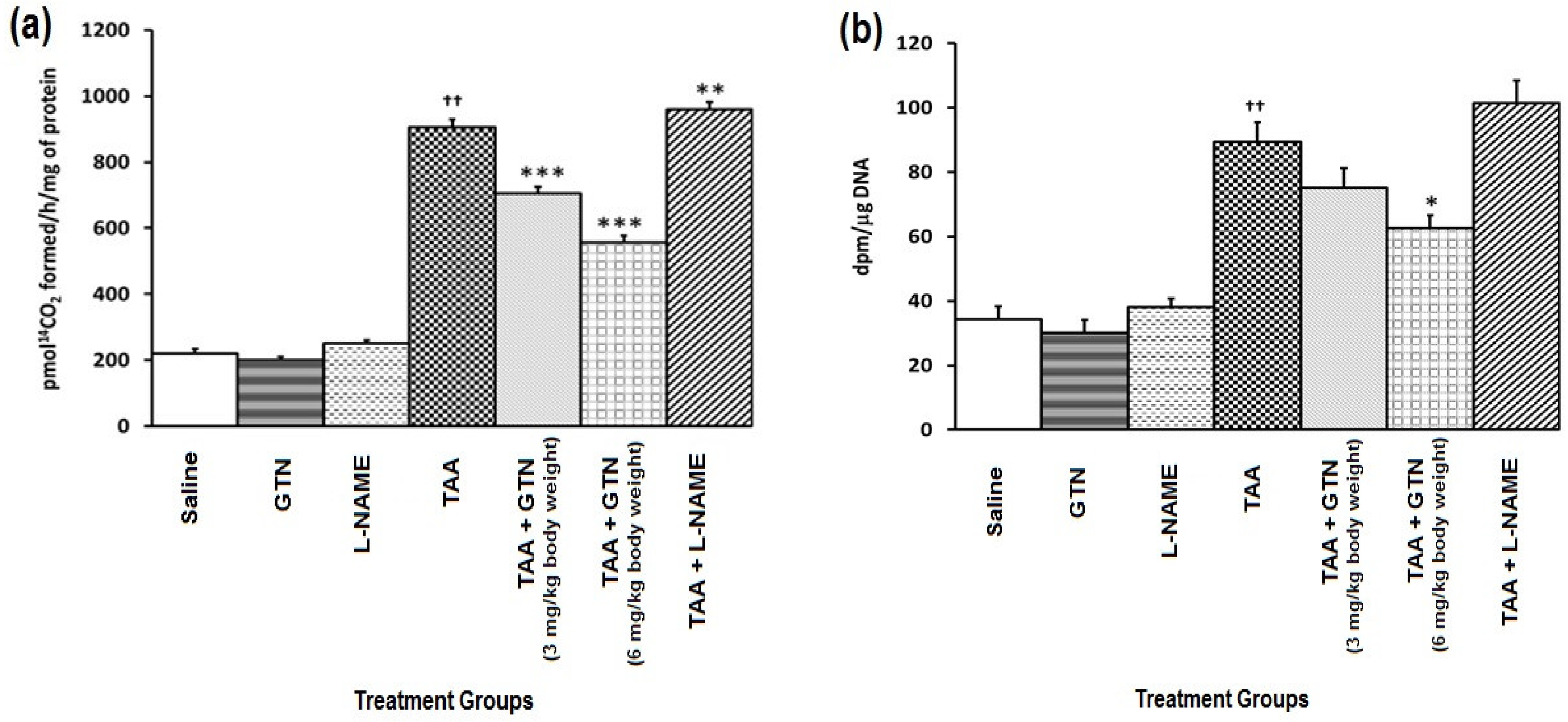
2.4. Effect of GTN and l-NAME on Hepatic Ornithine Decarboxylase (ODC) Activity and [3H]-Thymidine Incorporation in Rats Treated with Thioacetamide (TAA)
2.5. Effect of GTN and l-NAME on Hepatic Histopathology in Rats Treated with Thioacetamide (TAA)
2.6. Amount of Nitrite Generated by GTN
3. Discussion
4. Materials and Methods
4.1. Chemicals ad Reagents
4.2. Thioacetamide (TAA) Preparation
4.3. Scintillation Fluid Preparation
4.4. Animals and Experimental Protocols
4.5. Post-Mitochondrial Supernatant (PMS, Cytosol, and Microsome Preparation)
4.6. Reduced Glutathione (GSH)
4.7. Glutathione S-transferase (GST)
4.8. Glutathione Reductase (GR)
4.9. Glutathione Peroxidase (GPx)
4.10. γ-glutamyl Transpeptidase (GGT)
4.11. Glucose 6-phosphate Dehydrogenase (G6PD)
4.12. Lipid Peroxidation
4.13. Ornithine Decarboxylase (ODC) Activity
4.14. [3H]-thymidine Incorporation Assay
4.15. Serum Transaminases (AST and ALT)
4.16. Histopathological Studies
4.17. Nitrite Determination
4.18. Protein Assay
4.19. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Kucera, O.; Endlicher, R.; Rychtrmoc, D.; Lotkova, H.; Sobotka, O.; Cervinkova, Z. Acetaminophen toxicity in rat and mouse hepatocytes in vitro. Drug Chem. Toxicol. 2017, 40, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Moravcova, A.; Cervinkova, Z.; Kucera, O.; Mezera, V.; Lotkova, H. Antioxidative effect of epigallocatechin gallate against D-galactosamine-induced injury in primary culture of rat hepatocytes. Acta Med. 2014, 57, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, A.; Vasenwala, S.M.; Iqbal, M. Hepatoprotective potential of glyceryl trinitrate against chemically induced oxidative stress and hepatic injury in rats. Hum. Exp. Toxicol. 2017, 36, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Stankova, P.; Kucera, O.; Lotkova, H.; Rousar, T.; Endlicher, R.; Cervinkova, Z. The toxic effect of thioacetamide on rat liver in vitro. Toxicol. Vitr. 2010, 24, 2097–2103. [Google Scholar] [CrossRef]
- Rahman, A.; Ahmed, S.; Khan, N.; Sultana, S.; Athar, M. Glyceryl trinitrate, a nitric oxide donor, suppresses renal oxidant damage caused by potassium bromate. Redox Rep. 1999, 4, 263–269. [Google Scholar] [CrossRef]
- Zhao, S.; Lu, Z.H.; Cui, J.J.; Li, D.; Zhu, Y.L.; Gan, J.H. Nitroglycerin ameliorates liver injury and regulates adaptive immunity in mice. Drug Dev. Res. 2020, 81, 557–563. [Google Scholar] [CrossRef]
- Aprioku, J.S. Pharmacology of free radicals and the impact of reactive oxygen species on the testis. J. Reprod. Infertil. 2013, 14, 158–172. [Google Scholar]
- Knockaert, L.; Berson, A.; Ribault, C.; Prost, P.E.; Fautrel, A.; Pajaud, J.; Lepage, S.; Lucas-Clerc, C.; Bégué, J.M.; Fromenty, B.; et al. Carbon tetrachloride-mediated lipid peroxidation induces early mitochondrial alterations in mouse liver. Lab. Investig. 2012, 92, 396–410. [Google Scholar] [CrossRef] [Green Version]
- Hsu, D.Z.; Chu, P.Y.; Li, Y.H.; Chandrasekaran, V.R.; Liu, M.Y. Role of flavin-containing-monooxygenase-dependent neutrophil activation in thioacetamide-induced hepatic inflammation in rats. Toxicology 2012, 298, 52–58. [Google Scholar] [CrossRef]
- Ansil, P.N.; Nitha, A.; Prabha, S.P.; Wills, P.J.; Jazaira, V.; Latha, M.S. Protective effect of Amorphophallus campanulatus (Roxb.) Blume. tuber against thioacetamide induced oxidative stress in rats. Asian Pac. J. Trop. Med. 2011, 4, 870–877. [Google Scholar] [CrossRef] [Green Version]
- Bautista, M.; Andres, D.; Cascales, M.; Morales-Gonzalez, J.A.; Sanchez-Reus, M.I.; Madrigal-Santillan, E.; Valadez-Vega, C.; Fregoso-Aguilar, T.; Mendoza-Pérez, J.A.; Gutiérrez-Salinas, J.; et al. Role of Kupffer cells in thioacetamide-induced cell cycle dysfunction. Molecules 2011, 16, 8319–8331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, M.; Del Rio, M.A.; Benedi, J.; Sanchez-Reus, M.I.; Morales-Gonzalez, J.A.; Tellez-Lopez, A.M.; López-Orozco, M. Effect of dichloromethylene diphosphonate on liver regeneration following thioacetamide-induced necrosis in rats. World J. Hepatol. 2013, 5, 379–386. [Google Scholar] [CrossRef]
- De David, C.; Rodrigues, G.; Bona, S.; Meurer, L.; Gonzalez-Gallego, J.; Tunon, M.J.; Marroni, N.P. Role of quercetin in preventing thioacetamide-induced liver injury in rats. Toxicol. Pathol. 2011, 39, 949–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Seo, K.S.; Tadi, S.; Ahn, B.H.; Lee, J.U.; Heo, J.Y.; Han, J.; Song, M.S.; Kim, S.H.; Yim, Y.H.; et al. An indole derivative protects against acetaminophen-induced liver injury by directly binding to N-acetyl-p-benzoquinone imine in mice. Antioxid. Redox Signal. 2013, 18, 1713–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel Salam, O.M.; Mohammed, N.A.; Sleem, A.A.; Farrag, A.R. The effect of antidepressant drugs on thioacetamide-induced oxidative stress. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 735–744. [Google Scholar] [PubMed]
- Harris, C.; Shuster, D.Z.; Roman Gomez, R.; Sant, K.E.; Reed, M.S.; Pohl, J.; Hansen, J.M. Inhibition of glutathione biosynthesis alters compartmental redox status and the thiol proteome in organogenesis-stage rat conceptuses. Free Radic. Biol. Med. 2013, 63, 325–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.L.; Zhao, Q.; He, Q. Nitric oxide. Med. Gas. Res. 2019, 9, 170. [Google Scholar] [CrossRef]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Iwakiri, Y. Nitric oxide in liver fibrosis: The role of inducible nitric oxide synthase. Clin. Mol. Hepatol. 2015, 21, 319–325. [Google Scholar] [CrossRef]
- Heinrich, T.A.; da Silva, R.S.; Miranda, K.M.; Switzer, C.H.; Wink, D.A.; Fukuto, J.M. Biological nitric oxide signalling: Chemistry and terminology. Br. J. Pharmacol. 2013, 169, 1417–1429. [Google Scholar] [CrossRef] [Green Version]
- Toledo, J.C., Jr.; Augusto, O. Connecting the chemical and biological properties of nitric oxide. Chem. Res. Toxicol. 2012, 25, 975–989. [Google Scholar] [CrossRef] [PubMed]
- Gross, F.; Durner, J.; Gaupels, F. Nitric oxide, antioxidants and prooxidants in plant defence responses. Front. Plant. Sci. 2013, 4, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, K.; Mehta, S.K.; Shekhawat, G.S. Nitric oxide (NO) counteracts cadmium induced cytotoxic processes mediated by reactive oxygen species (ROS) in Brassica juncea: Cross-talk between ROS, NO and antioxidant responses. Biometals 2013, 26, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.F.; Wang, D.; Leu, F.J.; Chen, H.I. The protective role of nitric oxide and nitric oxide synthases in whole-body hyperthermia-induced hepatic injury in rats. Int. J. Hyperth. 2012, 28, 421–430. [Google Scholar] [CrossRef]
- Diesen, D.L.; Kuo, P.C. Nitric oxide and redox regulation in the liver: Part I. General considerations and redox biology in hepatitis. J. Surg. Res. 2010, 162, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Al-Jawad, F.H.; Al-Attar, Z.; Abbood, M.S. The Protective Effect of Nitroglycerin, N-Acetyl Cysteine and Metoprolol in CCL4 Induced Animal Model of Acute Liver Injury. Open Access Maced. J. Med. Sci. 2019, 7, 1739–1743. [Google Scholar] [CrossRef] [Green Version]
- Diao, T.J.; Chen, X.; Deng, L.H.; Chen, H.X.; Liang, Y.; Zhao, X.D.; Wang, Q.H.; Yuan, W.S.; Gao, B.C.; Ye, Y. Protective effect of nitric oxide on hepatopulmonary syndrome from ischemia-reperfusion injury. World J. Gastroenterol. 2012, 18, 3310–3316. [Google Scholar] [CrossRef]
- Mustafa, H.N.; El Awdan, S.A.; Hegazy, G.A. Protective role of antioxidants on thioacetamide-induced acute hepatic encephalopathy: Biochemical and ultrastructural study. Tissue Cell 2013, 45, 350–362. [Google Scholar] [CrossRef]
- Bahde, R.; Kapoor, S.; Bandi, S.; Bhargava, K.K.; Palestro, C.J.; Gupta, S. Directly acting drugs prostacyclin or nitroglycerine and endothelin receptor blocker bosentan improve cell engraftment in rodent liver. Hepatology 2013, 57, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Bilska-Wilkosz, A.; Kotanska, M.; Gorny, M.; Filipek, B.; Iciek, M. Is the mechanism of nitroglycerin tolerance associated with aldehyde dehydrogenase activity? A contribution to the ongoing discussion. Acta Biochim. Pol. 2019, 66, 627–632. [Google Scholar] [CrossRef]
- Girotti, A.W.; Korytowski, W. Nitric Oxide Inhibition of Chain Lipid Peroxidation Initiated by Photodynamic Action in Membrane Environments. Cell Biochem. Biophys. 2020, 78, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Koshiishi, I.; Yokota, A.; Takajo, T. Nitric oxide converts fatty acid alkoxyl radicals into fatty acid allyl radicals. Arch. Biochem. Biophys. 2011, 516, 154–159. [Google Scholar] [CrossRef] [PubMed]
- El-Beltagi, H.S.; Mohamed, H.I. Reactive Oxygen Species, Lipid Peroxidation and Antioxidative Defense Mechanism. Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Hein, T.W.; Zhang, C.; Zawieja, D.C.; Liao, J.C.; Kuo, L. Oxidized low-density lipoprotein inhibits nitric oxide-mediated coronary arteriolar dilation by up-regulating endothelial arginase I. Microcirculation 2011, 18, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Paolisso, G.; Casamassimi, A.; Al-Omran, M.; Barbieri, M.; Sommese, L.; Infante, T.; Ignarro, L.J. Effects of nitric oxide on cell proliferation: Novel insights. J. Am. Coll. Cardiol. 2013, 62, 89–95. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, C.; Chen, J.; Choi, S.; Zhou, Y.; Zhao, M.; Song, X.; Chen, X.; Maletić-Savatić, M.; Palzkill, T.; et al. Quantitative Real-Time Imaging of Glutathione with Subcellular Resolution. Antioxid. Redox Signal. 2019, 30, 1900–1910. [Google Scholar] [CrossRef]
- Lorincz, T.; Szarka, A. The determination of hepatic glutathione at tissue and subcellular level. J. Pharmacol. Toxicol. Methods 2017, 88, 32–39. [Google Scholar] [CrossRef]
- Djukic, M.M.; Jovanovic, M.D.; Ninkovic, M.; Stevanovic, I.; Ilic, K.; Curcic, M.; Vekic, J. Protective role of glutathione reductase in paraquat induced neurotoxicity. Chem. Biol. Interact. 2012, 199, 74–86. [Google Scholar] [CrossRef]
- Galal, A.M.; Walker, L.A.; Khan, I.A. Induction of GST and related events by dietary phytochemicals: Sources, chemistry, and possible contribution to chemoprevention. Curr. Top. Med. Chem. 2015, 14, 2802–2821. [Google Scholar] [CrossRef]
- Isei, M.O.; Kamunde, C. Effects of copper and temperature on heart mitochondrial hydrogen peroxide production. Free Radic. Biol. Med. 2020, 147, 114–128. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toshihiro, W.; Kurata, S.; Takuro, S.; Ichiro, O.; Yasuaki, S.; Ikeda, T.; Ureshino, H.; Ayuse, T. Protective Effect of Nitric Oxide on Liver Circulation from Ischemia Reperfusion Injury. J. Invest. Surg. 2015, 28, 236–244. [Google Scholar] [CrossRef]
- Schyman, P.; Printz, R.L.; Estes, S.K.; O’Brien, T.P.; Shiota, M.; Wallqvist, A. Assessing Chemical-Induced Liver Injury In Vivo From In Vitro Gene Expression Data in the Rat: The Case of Thioacetamide Toxicity. Front. Genet. 2019, 10, 1233. [Google Scholar] [CrossRef]
- Sui, H.; Matsumoto, H.; Wako, Y.; Kawasako, K. Evaluation of in vivo genotoxicity by thioacetamide in a 28-day repeated-dose liver micronucleus assay using male young adult rats. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2015, 780–781, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Michalopoulos, G.K. Advances in liver regeneration. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Miyajima, A. Liver regeneration and fibrosis after inflammation. Inflamm. Regen. 2016, 36, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, A. Reduced glutathione: A radioprotector or a modulator of DNA-repair activity? Nutrients 2013, 5, 525–542. [Google Scholar] [CrossRef] [Green Version]
- Ince, S.; Kucukkurt, I.; Cigerci, I.H.; Fatih Fidan, A.; Eryavuz, A. The effects of dietary boric acid and borax supplementation on lipid peroxidation, antioxidant activity, and DNA damage in rats. J. Trace Elem. Med. Biol. 2010, 24, 161–164. [Google Scholar] [CrossRef]
- Boutwell, R.K. Evidence that an elevated level of ornithine decarboxylase activity is essential component of tumor promotion. Adv. Polyam. Res. 1982, 4, 127–134. [Google Scholar]
- Chi, D.; Rong, L.; Wang, S.; Wang, X.; Wang, X.; Ma, L.; Shao, L. Role of oxidative/nitrative stress in hepatic encephalopathy induced by thioacetamide. Pharmazie. 2011, 66, 378–381. [Google Scholar]
- Czechowska, G.; Celinski, K.; Korolczuk, A.; Wojcicka, G.; Dudka, J.; Bojarska, A.; Reiter, R.J. Protective effects of melatonin against thioacetamide-induced liver fibrosis in rats. J. Physiol. Pharmacol. 2015, 66, 567–579. [Google Scholar]
- El Awdan, S.A.; Abdel Rahman, R.F.; Ibrahim, H.M.; Hegazy, R.R.; El Marasy, S.A.; Badawi, M.; Arbid, M.S. Regression of fibrosis by cilostazol in a rat model of thioacetamide-induced liver fibrosis: Up regulation of hepatic cAMP, and modulation of inflammatory, oxidative stress and apoptotic biomarkers. PLoS ONE 2019, 14, e0216301. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.C.; Hamesch, K.; Lunova, M.; Kim, Y.; Weiskirchen, R.; Strnad, P.; Friedman, S.L. Standard operating procedures in experimental liver research: Thioacetamide model in mice and rats. Lab. Anim. 2015, 49, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.D.; Gnanaraj, C.; Haque, A.T.; Iqbal, M. Antioxidative and chemopreventive effects of Nephrolepis biserrata against carbon tetrachloride (CCl4)-induced oxidative stress and hepatic dysfunction in rats. Pharm. Biol. 2015, 53, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Gnanaraj, C.; Shah, M.D.; Haque, A.T.; Makki, J.S.; Iqbal, M. Hepatoprotective and Immunosuppressive Effect of Synedrella nodiflora L. on Carbon Tetrachloride (CCl4)-Intoxicated Rats. J. Environ. Pathol. Toxicol. Oncol. 2016, 35, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Hayashi, N.; George, J.; Toshikuni, N.; Arisawa, T.; Hiratake, J.; Tsuchishima, M.; Tsutsumi, M. GGsTop, a novel and specific gamma-glutamyl transpeptidase inhibitor, protects hepatic ischemia-reperfusion injury in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G305–G312. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.D.; D’Souza, U.J.A.; Iqbal, M. The potential protective effect of Commelina nudiflora L. against carbon tetrachloride (CCl4)-induced hepatotoxicity in rats, mediated by suppression of oxidative stress and inflammation. Environ. Health Prev. Med. 2017, 22, 66. [Google Scholar] [CrossRef] [Green Version]
- Gnanaraj, C.; Shah, M.D.; Makki, J.S.; Iqbal, M. Hepatoprotective effects of Flagellaria indica are mediated through the suppression of pro-inflammatory cytokines and oxidative stress markers in rats. Pharm. Biol. 2016, 54, 1420–1433. [Google Scholar] [CrossRef] [Green Version]
- Ray, R.M.; Viar, M.J.; Johnson, L.R. Amino acids regulate expression of antizyme-1 to modulate ornithine decarboxylase activity. J. Biol. Chem. 2012, 287, 3674–3690. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, M.; Sundaram, H. Drug Design and Testing: Profiling of Antiproliferative Agents for Cancer Therapy Using a Cell-Based Methyl-[3H]-Thymidine Incorporation Assay. In Cancer Cell Culture: Methods and Protocols, 2nd ed.; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2011; Volume 731, pp. 451–465. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Treatment Groups | GST (nmol CDNB Conjugate Formed min−1 mg−1 Protein) | GR (nmol NADPH Oxidized min−1 mg−1 Protein) | GPx (nmol NADPH Oxidized min−1 mg−1 Protein) | GGT (nmol p-Nitroaniline Formed min−1 mg−1 Protein) | G6PD (nmol NADPH Formed min−1 mg−1 Protein) |
|---|---|---|---|---|---|
| Saline | 926.73 ± 35.91 | 220.69 ± 8.78 | 248.55 ± 10.96 | 417.76 ± 16.17 | 168.23 ± 7.20 |
| GTN | 908.21 ± 42.68 | 232.05 ± 10.17 | 259.65 ± 12.83 | 422.20 ± 17.96 | 181.02 ± 13.47 |
| l-NAME | 940.39 ± 26.12 | 245.57 ± 12.42 | 219.06 ± 9.71 | 449.54 ± 14.48 | 227.36 ± 11.45 † |
| TAA | 1504.36 ± 66.52 †† | 380.61 ± 14.42 †† | 129.43 ± 8.57 †† | 1041.13 ± 37.76 †† | 349.89 ± 16.27 †† |
| TAA + GTN (3 mg/kg bwt) | 1316.14 ± 59.44 | 306.23 ± 12.47 ** | 171.32 ± 9.11 ** | 870.71 ± 27.97 ** | 292.21 ± 9.10 * |
| TAA + GTN (6 mg/kg bwt) | 1242.39 ± 45.57 ** | 275.72 ± 10.12 *** | 198.43 ± 7.62 *** | 714.39 ± 14.36 *** | 267.88 ± 10.92 ** |
| TAA + l-NAME | 1682.18 ± 79.09 | 408.68 ± 13.45 | 96.22 ± 5.98 ** | 1132.19 ± 23.15 | 391.71 ± 13.74 |
| Treatment Groups | AST (IU/L) | ALT (IU/L) |
|---|---|---|
| Saline | 22.83 ± 1.14 | 16.07 ± 0.54 |
| GTN | 21.69 ± 1.55 | 16.26 ± 0.63 † |
| l-NAME | 28.29 ± 1.45 † | 20.41 ± 0.72 †† |
| TAA | 75.82 ± 2.23 †† | 46.72 ± 1.67 †† |
| TAA + GTN (3 mg/kg bwt) | 56.40 ± 1.30 *** | 38.37 ± 1.25 *** |
| TAA + GTN (6 mg/kg bwt) | 44.45 ± 1.23 *** | 24.97 ± 1.09 *** |
| TAA + l-NAME | 84.57 ± 2.08 * | 56.49 ± 1.38 ** |
| Group Number | Treatment Groups | Dosage Regimen |
|---|---|---|
| Group I, Control | Sodium chloride 0.9% w/v (Saline) | A dose of 10 mL/kg bwt, at the same time as the test agents. Control for test groups, II, III, and IV. |
| Group II | GTN | GTN: 6 mg/kg bwt, 1 h before killing the rats. |
| Group III | l-NAME | l-NAME: 40 mg/kg bwt, at the same times as l-NAME treatment in group VII. |
| Group IV | TAA | TAA: 500 mg/kg bwt, at the same time as TAA treatment in the groups V, VI, and VII. |
| Group V, Experimental Dose 1 | TAA + GTN (3 mg/kg bwt) | TAA: 500 mg/kg bwt + GTN: 3 mg/kg bwt, 1 h after TAA administration. |
| Group VI, Experimental Dose 2 | TAA + GTN (6 mg/kg bwt) | TAA: 500 mg/kg bwt + GTN: 6 mg/kg bwt, 1 h after TAA administration. |
| Group VII, Experimental | TAA + l-NAME | TAA: 500 mg/kg bwt + l-NAME: 40 mg/kg bwt, 1 h after TAA administration. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, A.R.; Ahmed, M.; Vun-Sang, S.; Iqbal, M. Is Glyceryl Trinitrate, a Nitric Oxide Donor Responsible for Ameliorating the Chemical-Induced Tissue Injury In Vivo? Molecules 2022, 27, 4362. https://doi.org/10.3390/molecules27144362
Ahmed AR, Ahmed M, Vun-Sang S, Iqbal M. Is Glyceryl Trinitrate, a Nitric Oxide Donor Responsible for Ameliorating the Chemical-Induced Tissue Injury In Vivo? Molecules. 2022; 27(14):4362. https://doi.org/10.3390/molecules27144362
Chicago/Turabian StyleAhmed, Ayesha Rahman, Mahiba Ahmed, Senty Vun-Sang, and Mohammad Iqbal. 2022. "Is Glyceryl Trinitrate, a Nitric Oxide Donor Responsible for Ameliorating the Chemical-Induced Tissue Injury In Vivo?" Molecules 27, no. 14: 4362. https://doi.org/10.3390/molecules27144362
APA StyleAhmed, A. R., Ahmed, M., Vun-Sang, S., & Iqbal, M. (2022). Is Glyceryl Trinitrate, a Nitric Oxide Donor Responsible for Ameliorating the Chemical-Induced Tissue Injury In Vivo? Molecules, 27(14), 4362. https://doi.org/10.3390/molecules27144362