
Synergistic Anti-Inflammatory Activity of Ginger and Turmeric Extracts in Inhibiting Lipopolysaccharide and Interferon-γ-Induced Proinflammatory Mediators

 ,
,  and
and
Abstract
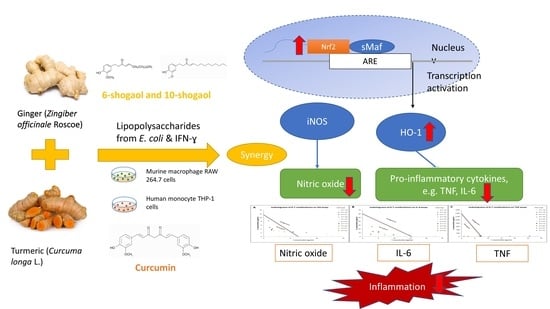
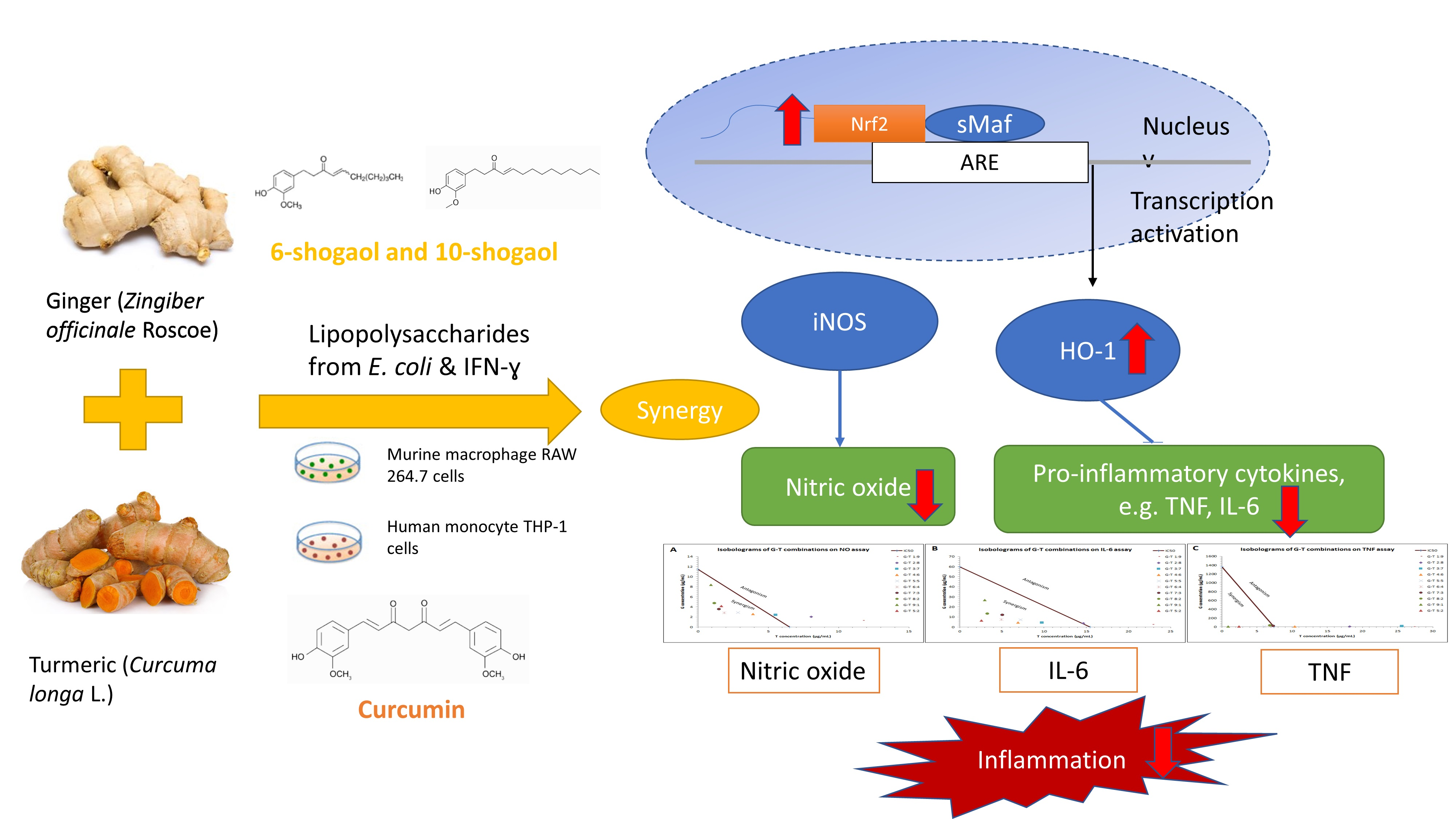
:
1. Introduction
2. Results
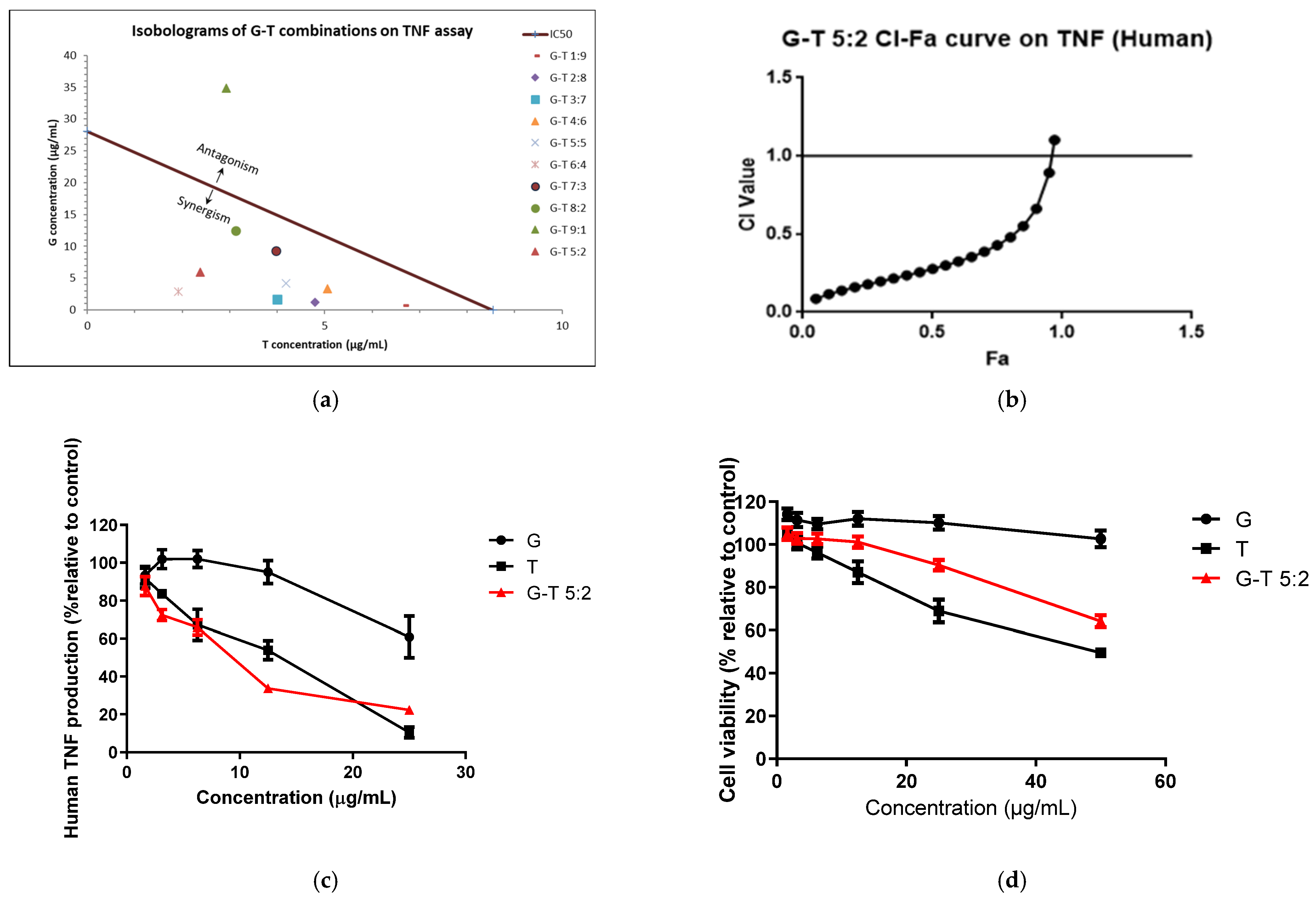
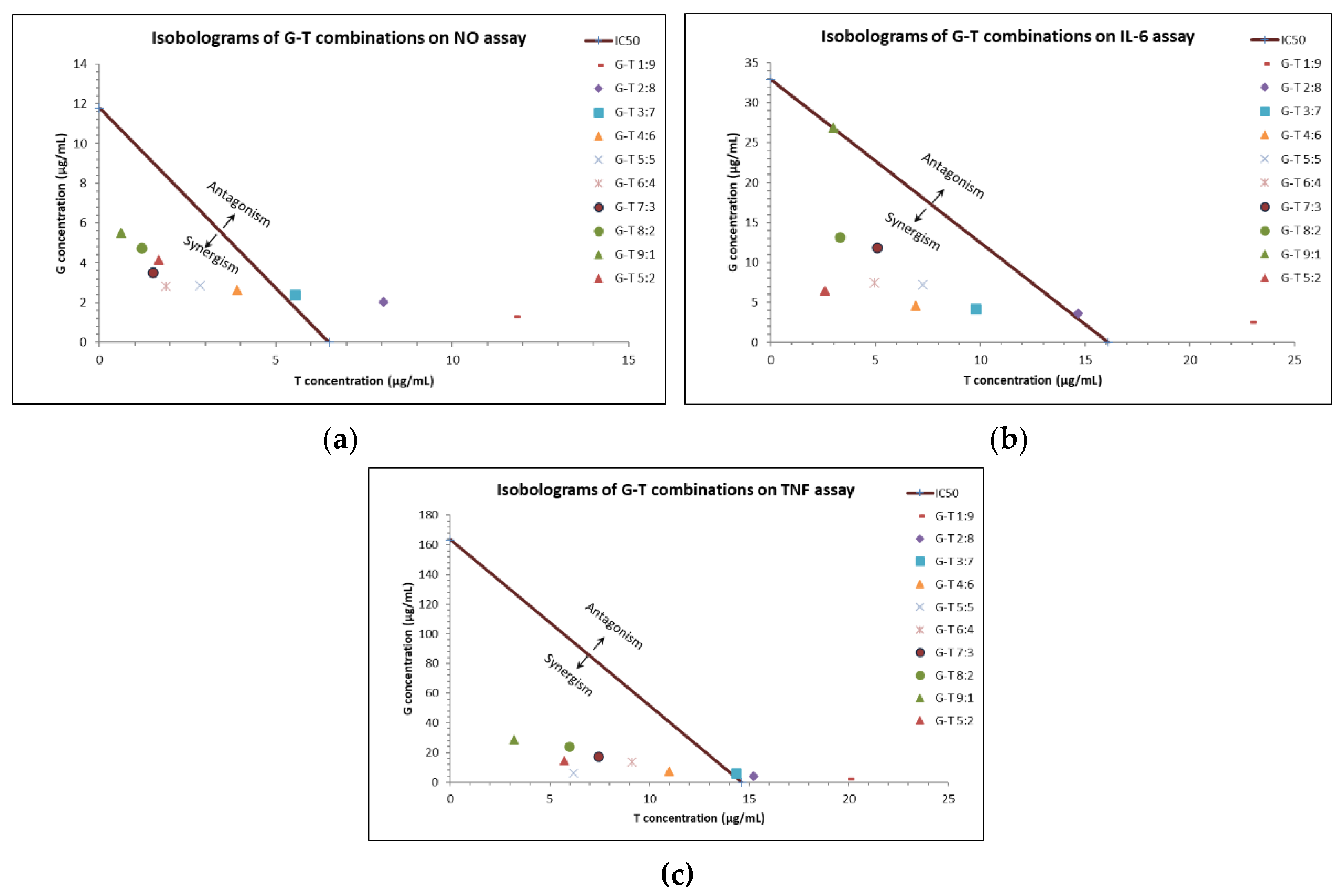
2.1. Synergistic Anti-Inflammatory Effects of G-T Combinations in RAW 264.7 Cells
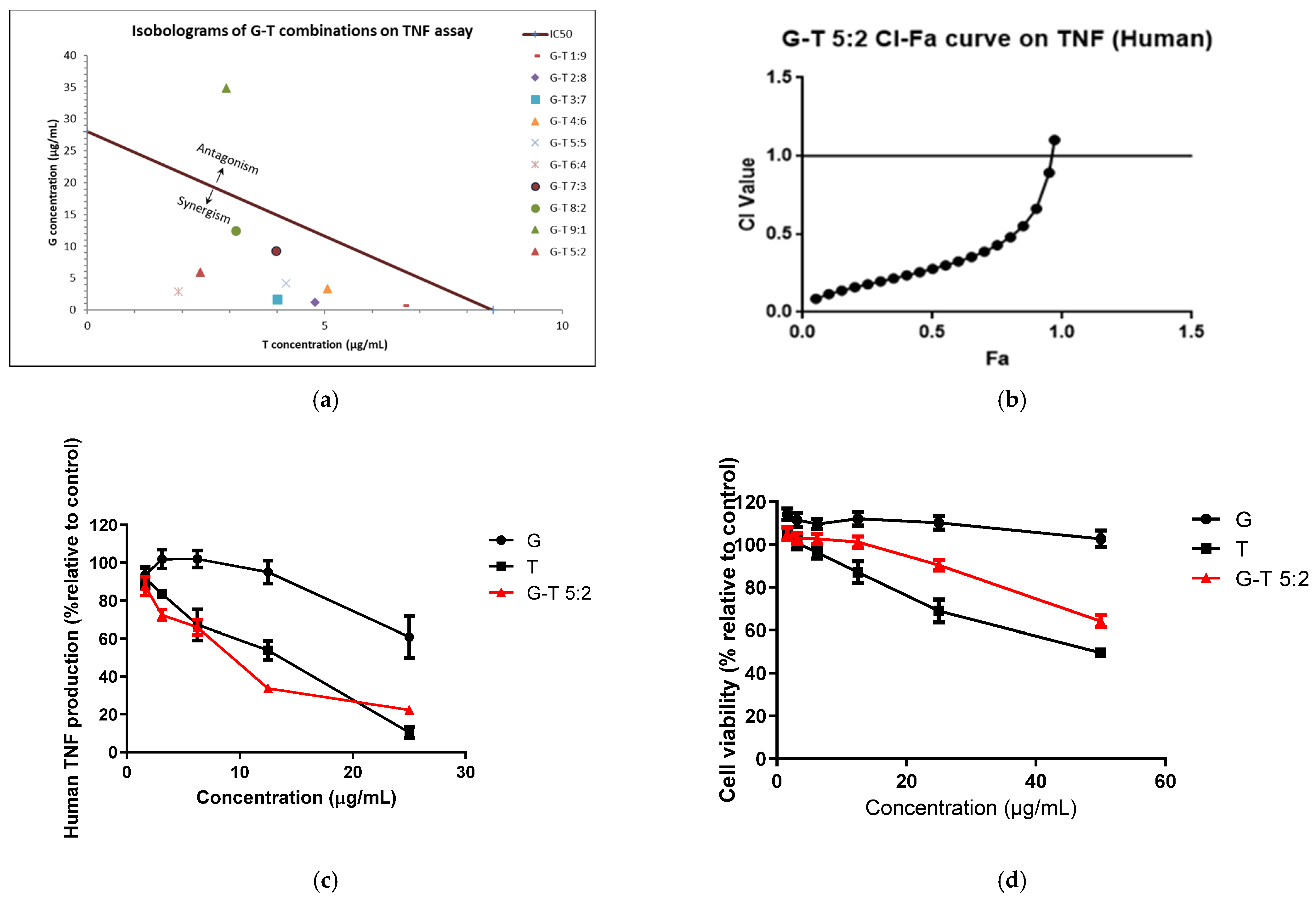
2.2. Synergistic Anti-Inflammatory Effects of G-T Combinations in THP-1 Cells
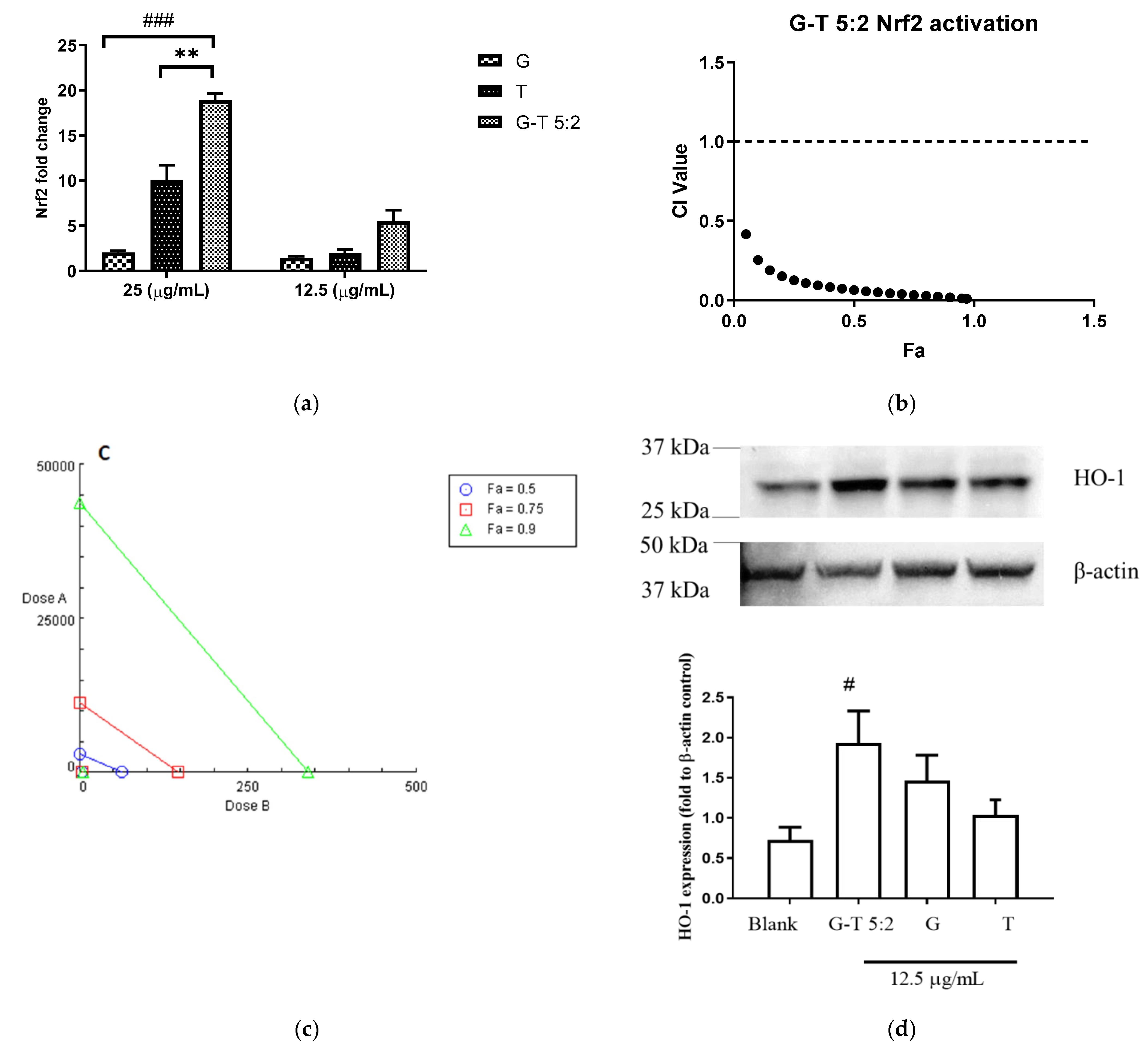
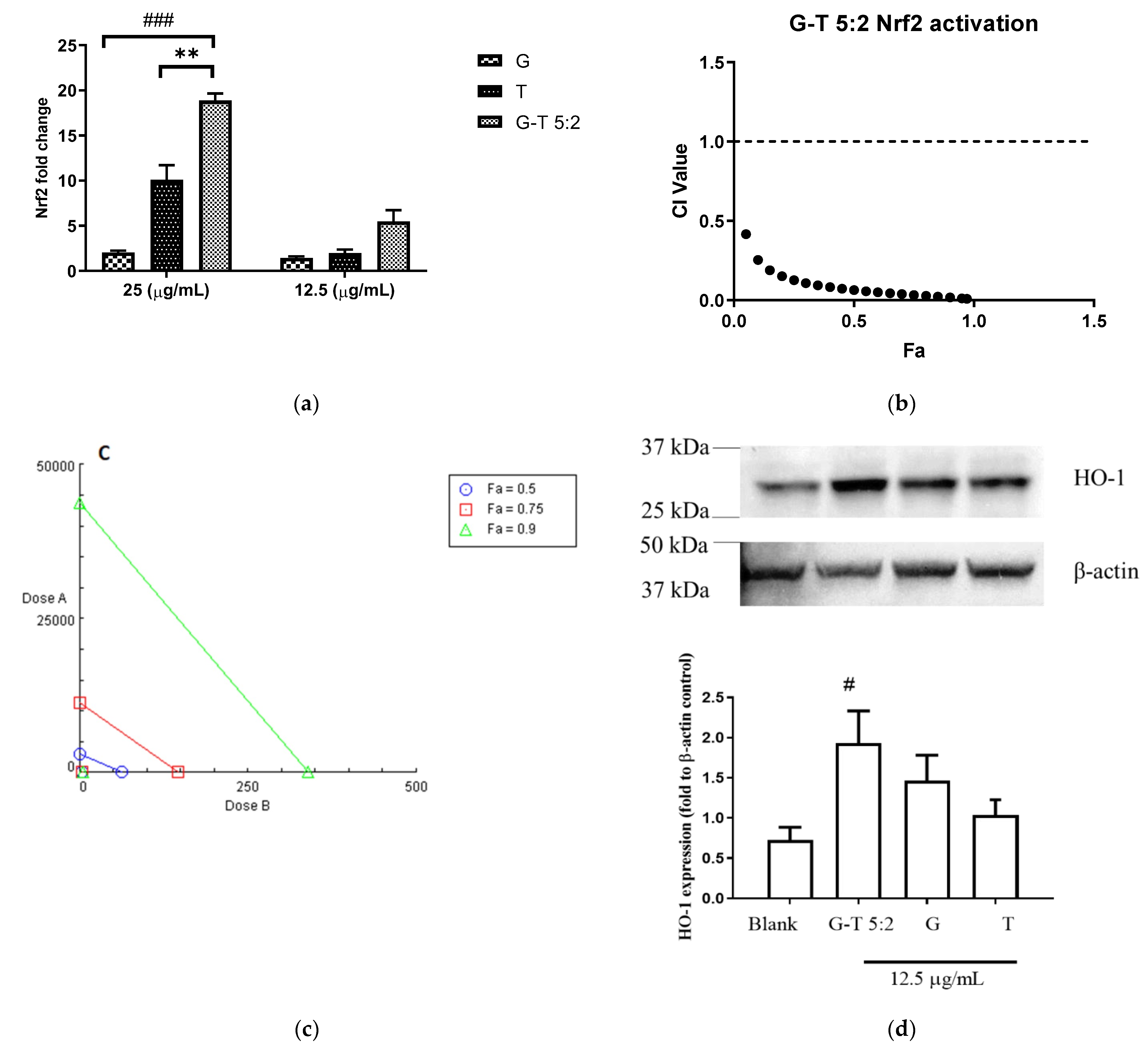
2.3. Effect of G-T 5:2 on the Activation of Nrf2 Luciferase in AREc32 Cells
2.4. Determining the Principal Compounds Contributing to the Synergistic Activity of G-T 5:2 Using NO Assay on RAW 264.7 Cells
3. Discussion
4. Materials and Methods
4.1. Preparation of Herbal Samples and Their Chemical Compounds
4.2. Cell Culture
4.3. Nitric Oxide Assay
4.4. TNF and IL-6 ELISA Assay
4.5. Cell Viability Assays
4.6. Determination of Nrf2 Expression by Luciferase Assay
4.7. Determination of HO-1 Protein Expression by Western Blotting Analysis
4.8. Determination of Synergistic, Additive or Antagonistic Interactions
4.9. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zeng, X.; Li, X.; Mehta, J.L.; Wang, X. Role of NLRP3 inflammasome in the pathogenesis of cardiovascular diseases. Basic Res. Cardiol. 2018, 113, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zingaropoli, M.A.; Nijhawan, P.; Carraro, A.; Pasculli, P.; Zuccalà, P.; Perri, V.; Marocco, R.; Kertusha, B.; Siccardi, G.; Del Borgo, C. Increased sCD163 and sCD14 plasmatic levels and depletion of peripheral blood pro-inflammatory monocytes, myeloid and plasmacytoid dendritic cells in patients with severe COVID-19 pneumonia. Front. Immunol. 2021, 12, 147. [Google Scholar] [CrossRef] [PubMed]
- Tay, M.Z.; Poh, C.M.; Rénia, L.; MacAry, P.A.; Ng, L.F. The trinity of COVID-19: Immunity, inflammation and intervention. Nat. Rev. Immunol. 2020, 20, 363–374. [Google Scholar] [CrossRef] [PubMed]
- García, L.F. Immune response, inflammation, and the clinical spectrum of COVID-19. Front. Immunol. 2020, 11, 1441. [Google Scholar] [CrossRef]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.-A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The role of inflammation in diabetes: Current concepts and future perspectives. Eur. Cardiol. 2019, 14, 50. [Google Scholar] [CrossRef] [Green Version]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63. [Google Scholar]
- Greten, F.R.; Grivennikov, S.I. Inflammation and cancer: Triggers, mechanisms, and consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Murata, M. Inflammation and cancer. Environ. Health Prev. Med. 2018, 23, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Simpson, D.S.; Oliver, P.L. ROS generation in microglia: Understanding oxidative stress and inflammation in neurodegenerative disease. Antioxidants 2020, 9, 743. [Google Scholar] [CrossRef]
- Ilievski, V.; Zuchowska, P.K.; Green, S.J.; Toth, P.T.; Ragozzino, M.E.; Le, K.; Aljewari, H.W.; O’Brien-Simpson, N.M.; Reynolds, E.C.; Watanabe, K. Chronic oral application of a periodontal pathogen results in brain inflammation, neurodegeneration and amyloid beta production in wild type mice. PLoS ONE 2018, 13, e0204941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, J.-F.; Lu, Z.Y.; Massart, C.; Levon, K. Dynamic immune/inflammation precision medicine: The good and the bad inflammation in infection and cancer. Front. Immunol. 2021, 12, 595722. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ying, Y. The inhibitory effect of curcumin on virus-induced cytokine storm and its potential use in the associated severe pneumonia. Front. Cell Dev. Biol. 2020, 8, 479. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, P.P.; Helson, L. Curcumin suppression of cytokine release and cytokine storm. A potential therapy for patients with Ebola and other severe viral infections. in vivo 2015, 29, 1–4. [Google Scholar]
- Noor, H.; Ikram, A.; Rathinavel, T.; Kumarasamy, S.; Nasir Iqbal, M.; Bashir, Z. Immunomodulatory and anti-cytokine therapeutic potential of curcumin and its derivatives for treating COVID-19–A computational modeling. J. Biomol. Struct. Dyn. 2021. [Google Scholar] [CrossRef]
- Xiao, B.; Zhang, Z.; Viennois, E.; Kang, Y.; Zhang, M.; Han, M.K.; Chen, J.; Merlin, D. Combination therapy for ulcerative colitis: Orally targeted nanoparticles prevent mucosal damage and relieve inflammation. Theranostics 2016, 6, 2250. [Google Scholar] [CrossRef]
- Zhou, X.; Seto, S.W.; Chang, D.; Kiat, H.; Razmovski-Naumovski, V.; Chan, K.; Bensoussan, A. Synergistic effects of Chinese herbal medicine: A comprehensive review of methodology and current research. Front. Pharmacol. 2016, 7, 201. [Google Scholar] [CrossRef] [Green Version]
- Geary, N. Understanding synergy. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E237–E253. [Google Scholar] [CrossRef]
- Williamson, E.M. Synergy and other interactions in phytomedicines. Phytomedicine 2001, 8, 401–409. [Google Scholar] [CrossRef]
- Roell, K.R.; Reif, D.M.; Motsinger-Reif, A.A. An introduction to terminology and methodology of chemical synergy—Perspectives from across disciplines. Front. Pharmacol. 2017, 8, 158. [Google Scholar] [CrossRef]
- Arts, E.J.; Hazuda, D.J. HIV-1 antiretroviral drug therapy. CSH Perspect. Med. 2012, 2, a007161. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Yeh, P.J. Suppressive drug combinations and their potential to combat antibiotic resistance. J. Antibiot. 2017, 70, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-C. The combination index (CI <1) as the definition of synergism and of synergy claims. Synergy 2018, 7, 49–50. [Google Scholar]
- Zhou, X.; Razmovski-Naumovski, V.; Chang, D.; Li, C.; Kam, A.; Low, M.; Bensoussan, A.; Chan, K. Synergistic effects of danshen (salvia miltiorrhiza radix et rhizoma) and sanqi (notoginseng radix et rhizoma) combination in inhibiting inflammation mediators in RAW 264. 7 cells. Biomed. Res. Int. 2016, 2016, 575819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Razmovski-Naumovski, V.; Kam, A.; Chang, D.; Li, C.G.; Chan, K.; Bensoussan, A. Synergistic study of a Danshen (Salvia Miltiorrhizae Radix et Rhizoma) and Sanqi (Notoginseng Radix et Rhizoma) combination on cell survival in EA. hy926 cells. BMC Complement. Altern. Med. 2019, 19, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Razmovski-Naumovski, V.; Kam, A.; Chang, D.; Li, C.; Bensoussan, A.; Chan, K. Synergistic effects of Danshen (Salvia Miltiorrhizae Radix et Rhizoma) and Sanqi (Notoginseng Radix et Rhizoma) combination in angiogenesis behavior in EAhy 926 cells. Medicines 2017, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Morvaridzadeh, M.; Fazelian, S.; Agah, S.; Khazdouz, M.; Rahimlou, M.; Agh, F.; Potter, E.; Heshmati, S.; Heshmati, J. Effect of ginger (Zingiber officinale) on inflammatory markers: A systematic review and meta-analysis of randomized controlled trials. Cytokine 2020, 135, 155224. [Google Scholar] [CrossRef]
- Chanda, S.; Ramachandra, T. Phytochemical and pharmacological importance of turmeric (Curcuma longa): A review. RRJoP 2019, 9, 16–23. [Google Scholar]
- Hong, M.-K.; Hu, L.-L.; Zhang, Y.-X.; Xu, Y.-L.; Liu, X.-Y.; He, P.-K.; Jia, Y.-H. 6-Gingerol ameliorates sepsis-induced liver injury through the Nrf2 pathway. Int. Immunopharmacol. 2020, 80, 106196. [Google Scholar] [CrossRef]
- Bischoff-Kont, I.; Fürst, R. Benefits of Ginger and Its Constituent 6-Shogaol in Inhibiting Inflammatory Processes. Pharmaceuticals 2021, 14, 571. [Google Scholar] [CrossRef]
- Boroumand, N.; Samarghandian, S.; Hashemy, S.I. Immunomodulatory, anti-inflammatory, and antioxidant effects of curcumin. J. HerbMed Pharmacol. 2018, 7, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Srirod, S.; Tewtrakul, S. Anti-inflammatory and wound healing effects of cream containing Curcuma mangga extract. J. Ethnopharmacol. 2019, 238, 111828. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.S.; Hussain, M.B.; Sultan, M.T.; Arshad, M.S.; Waheed, M.; Shariati, M.A.; Plygun, S.; Hashempur, M.H. Biochemistry, safety, pharmacological activities, and clinical applications of turmeric: A mechanistic review. Evid. Based Complementary Altern. Med. 2020, 2020, 7656919. [Google Scholar] [CrossRef] [PubMed]
- White, C.M.; Pasupuleti, V.; Roman, Y.M.; Li, Y.; Hernandez, A.V. Oral turmeric/curcumin effects on inflammatory markers in chronic inflammatory diseases: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2019, 146, 104280. [Google Scholar] [CrossRef] [PubMed]
- Kubra, I.R.; Rao, L.J.M. An impression on current developments in the technology, chemistry, and biological activities of ginger (Zingiber officinale Roscoe). Crit. Rev. Food Sci. Nutr. 2012, 52, 651–688. [Google Scholar] [CrossRef]
- Suryawati, N.; Jawi, I.M. Potential development of turmeric extract nanoparticles as a topical anti-inflammatory agent. Bali Med. J. 2020, 9, 573–578. [Google Scholar] [CrossRef]
- Muhammad, I.; Wang, X.; Li, S.; Li, R.; Zhang, X. Curcumin confers hepatoprotection against AFB 1-induced toxicity via activating autophagy and ameliorating inflammation involving Nrf2/HO-1 signaling pathway. Mol. Biol. Rep. 2018, 45, 1775–1785. [Google Scholar] [CrossRef]
- Chen, F.; Tang, Y.; Sun, Y.; Veeraraghavan, V.P.; Mohan, S.K.; Cui, C. 6-shogaol, a active constiuents of ginger prevents UVB radiation mediated inflammation and oxidative stress through modulating NrF2 signaling in human epidermal keratinocytes (HaCaT cells). J. Photochem. Photobiol. B 2019, 197, 111518. [Google Scholar] [CrossRef]
- Jafarzadeh, A.; Nemati, M. Therapeutic potentials of ginger for treatment of Multiple sclerosis: A review with emphasis on its immunomodulatory, anti-inflammatory and anti-oxidative properties. J. Neuroimmunol. 2018, 324, 54–75. [Google Scholar] [CrossRef]
- Jalali, M.; Mahmoodi, M.; Moosavian, S.P.; Jalali, R.; Ferns, G.; Mosallanezhad, A.; Imanieh, M.H.; Mosallanezhad, Z. The effects of ginger supplementation on markers of inflammatory and oxidative stress: A systematic review and meta-analysis of clinical trials. Phytother. Res. 2020, 34, 1723–1733. [Google Scholar] [CrossRef]
- Verma, R.K.; Kumari, P.; Maurya, R.K.; Kumar, V.; Verma, R.; Singh, R.K. Medicinal properties of turmeric (Curcuma longa L.): A review. Int. J. Chem. Stud. 2018, 6, 1354–1357. [Google Scholar]
- Ramadan, G.; El-Menshawy, O. Protective effects of ginger-turmeric rhizomes mixture on joint inflammation, atherogenesis, kidney dysfunction and other complications in a rat model of human rheumatoid arthritis. Int. J. Rheum. Dis. 2013, 16, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Munch, G.; Wohlmuth, H.; Afzal, S.; Kao, M.-H.T.; Al-Khazaleh, A.; Low, M.; Leach, D.; Li, C.G. Synergistic inhibition of pro-inflammatory pathways by ginger and turmeric extracts in RAW 264. 7 cells. Front. Pharmacol. 2022. [Google Scholar] [CrossRef]
- Zhong, Y.; Liu, C.; Feng, J.; Li, J.F.; Fan, Z.C. Curcumin affects ox-LDL-induced IL-6, TNF-α, MCP-1 secretion and cholesterol efflux in THP-1 cells by suppressing the TLR4/NF-κB/miR33a signaling pathway. Exp. Ther. Med. 2020, 20, 1856–1870. [Google Scholar] [CrossRef] [PubMed]
- Kahroba, H.; Davatgaran-Taghipour, Y. Exosomal Nrf2: From anti-oxidant and anti-inflammation response to wound healing and tissue regeneration in aged-related diseases. Biochimie 2020, 171, 103–109. [Google Scholar] [CrossRef]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Surh, Y.-J.; Na, H.-K. NF-κB and Nrf2 as prime molecular targets for chemoprevention and cytoprotection with anti-inflammatory and antioxidant phytochemicals. Genes Nutr. 2008, 2, 313. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Khor, T.O.; Saw, C.L.L.; Lin, W.; Wu, T.; Huang, Y.; Kong, A.-N.T. Role of Nrf2 in suppressing LPS-induced inflammation in mouse peritoneal macrophages by polyunsaturated fatty acids docosahexaenoic acid and eicosapentaenoic acid. Mol. Pharm. 2010, 7, 2185–2193. [Google Scholar] [CrossRef]
- Song, Y.; Hao, D.; Jiang, H.; Huang, M.; Du, Q.; Lin, Y.; Liu, F.; Chen, B. Nrf2 regulates Chi3l1 to suppress inflammation and improve post-traumatic osteoarthritis. J. Inflamm. Res. 2021, 14, 4079. [Google Scholar] [CrossRef]
- Zhou, S.; Shi, J.; Wen, H.; Xie, W.; Han, X.; Li, H. A chondroprotective effect of moracin on IL-1β-induced primary rat chondrocytes and an osteoarthritis rat model through Nrf2/HO-1 and NF-κB axes. Food Funct. 2020, 11, 7935–7945. [Google Scholar] [CrossRef]
- Peng, S.; Yao, J.; Liu, Y.; Duan, D.; Zhang, X.; Fang, J. Activation of Nrf2 target enzymes conferring protection against oxidative stress in PC12 cells by ginger principal constituent 6-shogaol. Food Funct. 2015, 6, 2813–2823. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-L.; Liu, X.-H.; Qiao, Y.; Ning, L.-N.; Li, W.-J.; Sun, Y.-S.; Liu, D.-S.; Gao, W.; Ma, C.-M. Allicin alleviates inflammation of diabetic macroangiopathy via the Nrf2 and NF-kB pathway. Eur. J. Pharmacol. 2020, 876, 173052. [Google Scholar] [CrossRef] [PubMed]
- Hennig, P.; Garstkiewicz, M.; Grossi, S.; Di Filippo, M.; French, L.E.; Beer, H.-D. The crosstalk between Nrf2 and inflammasomes. Int. J. Mol. Sci. 2018, 19, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, A.-M.; Tzieply, N.; Schmidt, M.V.; Von Knethen, A.; Namgaladze, D.; Yamamoto, M.; Brüne, B. Antioxidant signaling via Nrf2 counteracts lipopolysaccharide-mediated inflammatory responses in foam cell macrophages. Free Radic. Biol. Med. 2011, 50, 1382–1391. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-F.; Kuo, H.-P.; Liu, M.; Chou, C.-K.; Xia, W.; Du, Y.; Shen, J.; Chen, C.-T.; Huo, L.; Hsu, M.-C. KEAP1 E3 ligase-mediated downregulation of NF-κB signaling by targeting IKKβ. Mol. Cell 2009, 36, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Luo, P.; Li, X.; Liu, P.; Li, Y.; Xu, J. Nrf2/ARE is a key pathway for curcumin-mediated protection of TMJ chondrocytes from oxidative stress and inflammation. Cell Stress Chaperones 2020, 25, 395–406. [Google Scholar] [CrossRef]
- Dai, C.; Li, B.; Zhou, Y.; Li, D.; Zhang, S.; Li, H.; Xiao, X.; Tang, S. Curcumin attenuates quinocetone induced apoptosis and inflammation via the opposite modulation of Nrf2/HO-1 and NF-kB pathway in human hepatocyte L02 cells. Food Chem. Toxicol. 2016, 95, 52–63. [Google Scholar] [CrossRef]
- Ibrahim, S.G.; El-Emam, S.Z.; Mohamed, E.A.; Abd Ellah, M.F. Dimethyl fumarate and curcumin attenuate hepatic ischemia/reperfusion injury via Nrf2/HO-1 activation and anti-inflammatory properties. Int. Immunopharmacol. 2020, 80, 106131. [Google Scholar] [CrossRef]
- Liu, L.; Shang, Y.; Li, M.; Han, X.; Wang, J.; Wang, J. Curcumin ameliorates asthmatic airway inflammation by activating nuclear factor-E2-related factor 2/haem oxygenase (HO)-1 signalling pathway. Clin. Exp. Pharmacol. Physiol. 2015, 42, 520–529. [Google Scholar] [CrossRef]
- Lee, S.; Khoo, C.; Halstead, C.W.; Huynh, T.; Bensoussan, A. Liquid chromatographic determination of 6-, 8-, 10-gingerol, and 6-shogaol in ginger (Zingiber officinale) as the raw herb and dried aqueous extract. J. AOAC Int. 2007, 90, 1219–1226. [Google Scholar] [CrossRef] [Green Version]
- Upton, R.; Petrone, C. American Herbal Pharmacopoeia and Therapeutic Compendium. Bilberry Fruit Vaccinium myrtillus L: Standards of Analysis, Quality Control and Therapeutics 1999. Available online: https://www.semanticscholar.org/paper/Bilberry-Fruit-Vaccinium-myrtillus-L-.-Standards-of-Herbalist/7afb8b84d4e0a6adac06a5aebba3e151dd723eac (accessed on 23 May 2022).
- The United States Pharmacopeia. Powered Turmeric Extract. 2020. Available online: https://www.uspnf.com/errata/powdered-turmeric-extract-2017-02-01 (accessed on 23 May 2022).
- Wang, X.J.; Hayes, J.D.; Wolf, C.R. Generation of a stable antioxidant response element–driven reporter gene cell line and its use to show redox-dependent activation of Nrf2 by cancer chemotherapeutic agents. Cancer Res. 2006, 66, 10983–10994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Afzal, S.; Zheng, Y.-F.; Münch, G.; Li, C.G. Synergistic Protective Effect of Curcumin and Resveratrol against Oxidative Stress in Endothelial EAhy926 Cells. Evid. Based Complementary Altern. Med. 2021, 2021, 2661025. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts and Combinations | Cell Viability LC50 (µg/mL) (Mean ± STD) | NO | IL-6 | TNF | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| IC50 (µg/mL) (Mean ± STD) | CI Value at IC50 | Therapeutic Index | IC50 (µg/mL) (Mean ± STD) | CI Value at IC50 | Therapeutic Index | IC50 (µg/mL) (Mean ± STD) | CI Value at IC50 | Therapeutic Index | ||
| G | 104.3 ± 5.63 * | 11.78 ± 1.58 | N/A | 8.85 | 32.91 ± 9.06 | N/A | 3.17 | 163.40 ± 3.94 * | N/A | 0.64 |
| T | 83.90 ± 7.19 | 6.51 ± 1.28 | N/A | 12.88 | 16.10 ± 3.09 | N/A | 5.21 | 14.63 ± 2.19 | N/A | 5.73 |
| G-T 1:9 | 72.29 ± 4.77 ∆ | 13.08 ± 0.97 | 1.91 | 5.52 | 25.46 ± 4.28 | 1.50 | 2.84 | 22.20 ± 6.13 ∆ | 1.86 | 3.26 |
| G-T 2:8 | 76.93 ± 6.72 ∆ | 10.08 ± 0.96 | 1.41 | 7.63 | 18.33 ± 3.68 ∆ | 1.01 | 4.20 | 19.02 ± 3.09 ∆ | 1.10 | 4.04 |
| G-T 3:7 | 82.15 ± 3.00 ∆ | 7.93 ± 0.81 ∆ | 1.05 | 10.36 | 13.98 ± 2.00 ∆ | 0.70 | 5.88 | 20.51 ± 5.17 ∆ | 0.40 | 4.01 |
| G-T 4:6 | 81.00 ± 2.86 ∆ | 6.52 ± 0.70 ∆ | 0.83 | 12.42 | 11.52 ± 2.33 ∆ | 0.52 | 7.03 | 18.30 ± 2.64 ∆ | 0.73 | 4.43 |
| G-T 5:5 | 102.70 ± 2.10 | 5.72 ± 0.62 ∆ | 0.69 | 17.95 | 14.47 ± 3.04 ∆ | 0.59 | 7.10 | 12.40 ± 2.16 ∆ | 0.31 | 8.53 |
| G-T 6:4 | 115.80 ± 18.44 & | 4.72 ± 0.48 ∆ | 0.54 | 24.53 | 12.40 ± 2.02 ∆ | 0.45 | 9.34 | 22.81 ± 5.38 ∆ | 0.57 | 5.08 |
| G-T 7:3 | 93.29 ± 5.04 | 5.02 ± 0.23 ∆ | 0.53 | 18.58 | 16.94 ± 2.36 ∆ | 0.53 | 5.51 | 24.79 ± 4.58 ∆ | 0.62 | 3.76 |
| G-T 5:2 | 115.80 ± 11.4 & | 5.83 ± 0.81 ∆ | 0.61 | 19.86 | 9.07 ± 1.47 ∆ | 0.23 | 12.76 | 20.07 ± 3.33 ∆ | 0.28 | 5.77 |
| G-T 8:2 | 104.80 ± 6.23 & | 5.92 ± 1.18 ∆ | 0.59 | 17.70 | 16.53 ± 3.79 ∆ | 0.43 | 6.34 | 29.92 ± 6.24 ∆ | 0.49 | 3.50 |
| G-T 9:1 | 107.40 ± 8.67 & | 6.11 ± 0.96 ∆ | 0.57 | 17.58 | 29.87 ± 7.66 | 1.00 | 3.60 | 31.91 ± 6.16 ∆ | 0.41 | 3.37 |
| Extracts and Combinations | Cell Viability LC50 (µg/mL) | IC50 (µg/mL) Mean ± STD | CI Value at IC50 | Therapeutic Index |
|---|---|---|---|---|
| G | 123.8 ± 9.10 | 28.04 ± 3.99 | N/A | N/A |
| T | 47.24 ± 7.84 | 8.54 ± 1.43 | N/A | 5.53 |
| G-T 1:9 | 40.94 ± 11.08 ∆ | 7.40 ± 1.32 ∆ | 1.52 | 5.53 |
| G-T 2:8 | 41.16 ± 10.90 ∆ | 5.99 ± 0.70 ∆ | 0.60 | 6.87 |
| G-T 3:7 | 45.30 ± 6.74 ∆ | 5.72 ± 0.57 ∆ | 0.11 | 7.92 |
| G-T 4:6 | 52.03 ± 8.97 ∆ | 8.43 ± 1.25 ∆ | 0.71 | 6.17 |
| G-T 5:5 | 53.16 ± 7.79 ∆ | 8.37 ± 1.44 ∆ | 0.64 | 6.35 |
| G-T 6:4 | 51.26 ± 7.62 ∆ | 4.79 ± 1.08 ∆ | 0.33 | 10.70 |
| G-T 7:3 | 52.61 ± 5.41 ∆ | 13.24 ± 3.11 ∆ | 0.80 | 3.97 |
| G-T 5:2 | 73.61 ± 8.05 ∆,& | 8.32 ± 1.85 ∆ | 0.49 | 8.85 |
| G-T 8:2 | 56.35 ± 6.25 ∆ | 15.58 ± 2.93 ∆ | 0.81 | 3.62 |
| G-T 9:1 | 118.90 ± 6.52 & | N/A | N/A | N/A |
| Bioactive Compounds in G and T | Content in G or T (mg/g) | IC50 (µg/mL) of Individual Compounds (µg/mL) | IC50 (µg/mL) of Paired Combinations Equivalent to Their Ratio in G-T 5:2 | IC50 (µg/mL) of Three-Compound Combinations Equivalent to Their Ratio in G-T 5:2 | IC50 (µg/mL) of Six-Compound Combinations Equivalent to Their Ratio in G-T 5:2 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Compounds in G | C | B | D | Compounds in G | C | Compounds in G | C, B, D | |||
| 6-g | 69.57 ± 0.16 | ND * | 6-g | 17.21 ± 2.41 | 43.93 ± 0.67 | 9.44 ± 1.42 | 6-g, 8-g | 6.31 ± 0.83 | 6-g, 8-g, 10-g | 5.52 ± 0.64 |
| 8-g | 10.43 ± 0.23 | ND * | 8-g | 12.31 ± 4.41 | 27.33 ± 3.75 | 4.89 ± 1.07 | 6-g, 10-g | 5.37 ± 0.58 | 6-s, 8-s, 10-s | 5.41 ± 0.70 |
| 10-g | 19.62 ± 0.63 | ND * | 10-g | 10.02 ± 5.28 | 17.34 ± 5.38 | 11.86 ± 2.86 | 8-g, 10-g | 5.12 ± 0.56 | Extracts | IC50 (μg/mL) |
| 6-s | 7.48 ± 0.19 | 2.90 ± 1.22 | 6-s | 16.06 ± 1.12 | 12.70 ± 2.09 | 9.06 ± 1.16 | 6-s, 8-s | 3.34 ± 0.44 | G | 11.78 ± 1.58 |
| 8-s | 1.56 ± 0.03 | 1.96 ± 1.50 | 8-s | 13.19 ± 1.91 | 12.61 ± 1.64 | 6.33 ± 0.81 | 6-s, 10-s | 2.91 ± 0.20 | T | 6.51 ± 1.28 |
| 10-s | 2.30 ± 0.03 | 3.69 ± 0.86 | 10-s | 10.02 ± 1.95 | 40.32 ± 2.50 | 9.68 ± 0.92 | 8-s, 10-s | 3.82 ± 0.25 | G-T 5:2 | 5.83 ± 0.81 |
| C | 751.76 ± 101.45 | 5.87 ± 0.12 | ||||||||
| B | 14.77 ± 3.63 | 16.14 ± 1.68 | ||||||||
| D | 156.15 ± 26.24 | 8.30 ± 1.78 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Afzal, S.; Wohlmuth, H.; Münch, G.; Leach, D.; Low, M.; Li, C.G. Synergistic Anti-Inflammatory Activity of Ginger and Turmeric Extracts in Inhibiting Lipopolysaccharide and Interferon-γ-Induced Proinflammatory Mediators. Molecules 2022, 27, 3877. https://doi.org/10.3390/molecules27123877
Zhou X, Afzal S, Wohlmuth H, Münch G, Leach D, Low M, Li CG. Synergistic Anti-Inflammatory Activity of Ginger and Turmeric Extracts in Inhibiting Lipopolysaccharide and Interferon-γ-Induced Proinflammatory Mediators. Molecules. 2022; 27(12):3877. https://doi.org/10.3390/molecules27123877
Chicago/Turabian StyleZhou, Xian, Sualiha Afzal, Hans Wohlmuth, Gerald Münch, David Leach, Mitchell Low, and Chun Guang Li. 2022. "Synergistic Anti-Inflammatory Activity of Ginger and Turmeric Extracts in Inhibiting Lipopolysaccharide and Interferon-γ-Induced Proinflammatory Mediators" Molecules 27, no. 12: 3877. https://doi.org/10.3390/molecules27123877
APA StyleZhou, X., Afzal, S., Wohlmuth, H., Münch, G., Leach, D., Low, M., & Li, C. G. (2022). Synergistic Anti-Inflammatory Activity of Ginger and Turmeric Extracts in Inhibiting Lipopolysaccharide and Interferon-γ-Induced Proinflammatory Mediators. Molecules, 27(12), 3877. https://doi.org/10.3390/molecules27123877