
RETRACTED: Artemisia Extracts and Artemisinin-Based Antimalarials for COVID-19 Management: Could These Be Effective Antivirals for COVID-19 Treatment? †
{kind=link}
Abstract
:1. Introduction
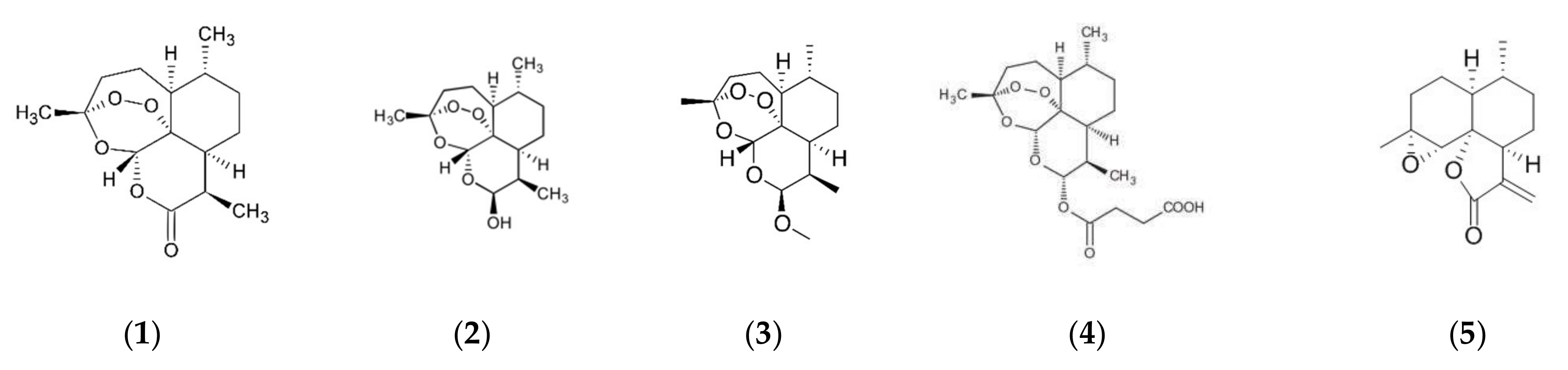
2. Traditional Use and Bioactive Compounds of Artemisia
3. Anti-Viral and Immune-Stimulatory Potential of Artemisia
4. Artemisia Extracts and COVID-19
5. Artemisia Supplement and Formulation
6. Artemisinin and Artemisinins’ Potential in Management of COVID-19
7. Artemisinin-Based Combination Therapies (ACTs)
8. Artemisia and Zinc
9. Brief Description of SARS-CoV-2 Genome and Possible Targets for Inhibition and Mode of Action
10. Inhibition Potential of Artemisia Constituents, including Artemisinin and Its Derivatives “Artemisinins” (such as Artesunate and Artmether) against SARS-CoV-2 Proteases
11. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Billingsley, A. FDA COVID-19 Vaccine Approval: Live Updates on Pfizer, Moderna, and J&J Vaccines. Available online: https://www.goodrx.com/conditions/covid-19/fda-covid-19-vaccine-approval-updates (accessed on 14 February 2022).
- Woolf, S.H.; Chapman, D.A.; Lee, J.H. COVID-19 as the leading cause of death in the United States. JAMA 2021, 325, 123–124. [Google Scholar] [CrossRef] [PubMed]
- Faust, J.S.; Krumholz, H.M.; Du, C.; Mayes, K.D.; Lin, Z.; Gilman, C.; Walensky, R.P. All-cause excess mortality and COVID-19-related mortality among US adults aged 25–44 years, March–July 2020. JAMA 2021, 325, 785–787. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, Y.; Zhang, C.; Huang, F.; Wang, F.; Yuan, J.; Wang, Z.; Li, J.; Feng, C.; Zhang, Z.; et al. Clinical and biochemical indexes from 2019-nCoV infected patients linked to viral loads and lung injury. Sci. China Life Sci. 2020, 63, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Muratov, E.N.; Amaro, R.; Andrade, C.H.; Brown, N.; Ekins, S.; Fourches, D.; Isayev, O.; Kozakov, D.; Medina-Franco, J.L.; Merz, K.M.; et al. A critical overview of computational approaches employed for COVID-19 drug discovery. Chem. Soc. Rev. 2021, 50, 9121–9151. [Google Scholar] [CrossRef] [PubMed]
- Qazi, S.; Das, S.; Khuntia, B.K.; Sharma, V.; Sharma, S.; Sharma, G.; Raza, B.K. In silico molecular docking and molecular dynamic simulation analysis of phytochemicals from Indian foods as potential inhibitors of SARS-CoV-2 RdRp and 3CLpro. Nat. Prod. Commun. 2021, 16, 1–12. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Pharmacological significance of hesperidin and hesperetin, two citrus flavonoids, as promising antiviral compounds for prophylaxis against and combating COVID-19. Nat. Prod. Commun. 2021, 16, 1934578X211042540. [Google Scholar] [CrossRef]
- Doharey, P.K.; Singh, V.; Rao Gedda, M.; Sahoo, A.K.; Varadwaj, P.K.; Sharma, B. In silico study indicates antimalarials as direct inhibitors of SARS-CoV-2-RNA dependent RNA polymerase. J. Biomol. Struct. Dyn. 2021, 1–18. [Google Scholar] [CrossRef]
- Bhuiyan, F.R.; Howlader, S.; Raihan, T.; Hasan, M. Plants metabolites: Possibility of natural therapeutics against the COVID-19 pandemic. Front. Med. 2020, 7, 444. [Google Scholar] [CrossRef]
- Remali, J.; Aizat, W.M. A review on plant bioactive compounds and their modes of action against coronavirus infection. Front. Pharmacol. 2021, 11, 589044. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Naringenin as a possible candidate against SARS-CoV-2 infection and in the pathogenesis of COVID-19. Nat. Prod. Commun. 2021, 16, 1934578X211066723. [Google Scholar] [CrossRef]
- Aherfi, S.; Pradines, B.; Devaux, C.; Honore, S.; Colson, P.; Scola, B.L.; Raoult, D. Drug repurposing against SARS-CoV-1, SARS-CoV-2 and MERS-CoV. Future Microbiol. 2021, 16, 1341–1370. [Google Scholar] [CrossRef]
- Boukhatem, M.N.; Setzer, W.N. Aromatic herbs, medicinal plant-derived essential oils, and phytochemical extracts as potential therapies for coronaviruses: Future perspectives. Plants 2020, 9, 800. [Google Scholar] [CrossRef]
- Ogwang, P.E.; Ogwal, J.O.; Kasasa, S.; Olila, D.; Ejobi, F.; Kabasa, D.; Obua, C. Artemisia annua L. infusion consumed once a week reduces risk of multiple episodes of malaria: A randomized trial in a Ugandan community. Trop. J. Pharm. Res. 2012, 11, 445–453. [Google Scholar] [CrossRef]
- Daddy, N.B.; Kalisya, L.M.; Bagire, P.G.; Watt, R.L.; Towler, M.J.; Weathers, P.J. Artemisia annua dried leaf tablets treated malaria resistant to ACT and i.v. artesunate: Case reports. Phytomedicine 2017, 32, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Munyangi, J.; Cornet-Vernet, L.; Idumbo, M.; Lu, C.; Lutgen, P.; Perronne, C.; Ngombe, N.; Bianga, J.; Mupenda, B.; Lalukala, P.; et al. Artemisia annua and Artemisia afra tea infusions vs. artesunate-amodiaquine (ASAQ) in treating Plasmodium falciparum malaria in a large scale, double blind, randomized clinical trial. Phytomedicine 2019, 57, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Cheong, D.H.J.; Tan, D.W.S.; Wong, F.W.S.; Tran, T. Anti-malarial drug, artemisinin and its derivatives for the treatment of respiratory diseases. Pharmacol. Res. 2020, 158, 104901. [Google Scholar] [CrossRef]
- Sadiq, A.; Hayat, M.Q.; Ashraf, M. Ethnopharmacology of Artemisia annua L.: A review. In Artemisia annua—Pharmacology and Biotechnology; Aftab, T., Ferreira, J.F.S., Khan, M.M.A., Naeem, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 9–25. [Google Scholar]
- Nigam, M.; Atanassova, M.; Mishra, A.P.; Pezzani, R.; Devkota, H.P.; Plygun, S.; Salehi, B.; Setzer, W.N.; Sharifi-Rad, J. Bioactive compounds and health benefits of Artemisia species. Nat. Prod. Commun. 2019, 14, 1934578X19850354. [Google Scholar]
- Kshirsagar, S.G.; Rao, R.V. Antiviral and immunomodulation effects of Artemisia. Medicina 2021, 57, 217. [Google Scholar] [CrossRef]
- De Ridder, S.; van der Kooy, F.; Verpoorte, R. Artemisia annua as a self-reliant treatment for malaria in developing countries. J. Ethnopharmacol. 2008, 120, 302–314. [Google Scholar] [CrossRef]
- Klayman, D.L. Artemisia annua: From weed to respectable antimalarial plant. In Human Medicinal Agents from Plants; Kinghorn, A.D., Balandrin, M.F., Eds.; American Chemical Society: Washington, DC, USA, 1993; pp. 242–255. [Google Scholar]
- Efferth, T. From ancient herb to modern drug: Artemisia annua and artemisinin for cancer therapy. Semin. Cancer Biol. 2017, 46, 65–83. [Google Scholar] [CrossRef]
- Pinheiro, L.C.S.; Feitosa, L.M.; Silveira, F.F.D.A.; Boechat, N. Current antimalarial therapies and advances in the development of semisynthetic artemisinin derivatives. An. Acad. Bras. Ciênc. 2018, 90, 1251–1271. [Google Scholar] [CrossRef]
- An, J.; Minie, M.; Sasaki, T.; Woodward, J.J.; Elkon, K.B. Antimalarial drugs as immune modulators: New mechanisms for old drugs. Annu. Rev. Med. 2017, 68, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Li, H.; Yang, Y.; Hou, L. Anti-inflammatory and immunoregulatory functions of artemisinin and its derivatives. Mediat. Inflamm. 2015, 2015, 435713. [Google Scholar] [CrossRef] [PubMed]
- Alesaeidi, S.; Miraj, S. A systematic review of anti-malarial properties, immunosuppressive properties, anti-inflammatory properties, and anti-cancer properties of Artemisia annua. Electron. Physician 2016, 8, 3150–3155. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.V. Artemisia—Antiviral and Immunomodulation Effects. Available online: https://encyclopedia.pub/8738 (accessed on 16 February 2022).
- Khanal, P. Antimalarial and anticancer properties of artesunate and other artemisinins: Current development. Monatsh. Chem. 2021, 152, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.F.S.; Luthria, D.L.; Sasaki, T.; Heyerick, A. Flavonoids from Artemisia annua L. as antioxidants and their potential synergism with artemisinin against malaria and cancer. Molecules 2010, 15, 3135–3170. [Google Scholar] [CrossRef] [PubMed]
- Mesa, L.E.; Lutgen, P.; Velez, I.D.; Segura, A.M.; Sara, M.; Robledo, S.M. Artemisia annua L., potential source of molecules with pharmacological activity in human diseases. Am. J. Phytomed. Clin. Ther. 2015, 3, 436–445. [Google Scholar]
- Feng, X.; Cao, S.; Qiu, F.; Zhang, B. Traditional application and modern pharmacological research of Artemisia annua L. Pharmacol. Ther. 2020, 216, 107650. [Google Scholar] [CrossRef]
- Ekiert, H.; Świątkowska, J.; Klin, P.; Rzepiela, A.; Szopa, A. Artemisia annua—Importance in traditional medicine and current state of knowledge on the chemistry, biological activity and possible applications. Planta Med. 2021, 87, 584–599. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Vishwakarma, R.A.; Jain, D.C.; Roy, R. High field NMR spectroscopic studies of arteannuin B and a reappraisal of the structure of arteannuin C. Phytochemistry 1991, 30, 3469–3471. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Bishnoi, V. Sterol and taraxastane derivatives from Artemisia annua and a rational approach based upon C-13 NMR for the identification of skeletal type of amorphane sesquiterpenoids. Ind. J. Chem. 1996, 35B, 86–88. [Google Scholar]
- Singh, A.K.; Pathak, V.; Agrawal, P.K. Annphenone, a phenolic acetophenone from Artemisia Annu. Phytochemistry 1997, 44, 555–557. [Google Scholar] [CrossRef]
- Brown, G.D. The biosynthesis of Artemisinin (Qinghaosu) and the phytochemistry of Artemisia annua L. (Qinghao). Molecules 2010, 15, 7603–7698. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.P.; Li, H.B.; Pang, Q.Q.; Huang, Y.X.; Pan, D.B.; Su, Z.Z.; Yao, X.J.; Yao, X.S.; Xiao, W.; Yu, Y. Structurally diverse sesquiterpenoids from the aerial parts of Artemisia annua (Qinghao) and their striking systemically anti-inflammatory activities. Bioorg. Chem. 2020, 103, 104221. [Google Scholar] [CrossRef] [PubMed]
- Septembre-Malaterre, A.; Lalarizo Rakoto, M.; Marodon, C.; Bedoui, Y.; Nakab, J.; Simon, E.; Hoarau, L.; Savriama, S.; Strasberg, D.; Guiraud, P.; et al. Artemisia annua, a traditional plant brought to light. Int. J. Mol. Sci. 2020, 21, 4986. [Google Scholar] [CrossRef] [PubMed]
- Bisht, D.; Kumar, D.; Kumar, D.; Kamal Dua, K.; Chellappan, D.K. Phytochemistry and pharmacological activity of the genus Artemisia. Arch. Pharm. Res. 2021, 44, 439–474. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the Treatment of Malaria, 3rd ed.; WHO: Geneva, Switzerland, 2015; Available online: https://apps.who.int/iris/bitstream/handle/10665/162441/9789241549127_eng.pdf (accessed on 1 May 2022).
- Zeyuan, L. Artemisinin chemical research. In Artemisinin-Based and Other Antimalarials; Guoqiao, L., Ying, L., Zelin, L., Meiyi, Z., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 129–175. [Google Scholar]
- Efferth, T.; Romero, M.R.; Wolf, D.G.; Stamminger, T.; Marin, J.J.G.; Marschall, M. The antiviral activities of artemisinin and artesunate. Clin. Infect. Dis. 2008, 47, 804–811. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, B.; Ashraf, U.; Zhang, H.; Cao, C.; Li, Q.; Chen, Z.; Imran, M.; Chen, H.; Cao, S.; et al. Artemisinin inhibits the replication of flaviviruses by promoting the type I interferon production. Antivir. Res. 2020, 179, 104810. [Google Scholar] [CrossRef]
- Desrosiers, M.R.; Mittelman, A.; Weathers, P.J. Dried leaf Artemisia annua improves bioavailability of artemisinin via cytochrome P450 inhibition and enhances artemisinin efficacy downstream. Biomolecules 2020, 10, 254. [Google Scholar] [CrossRef]
- Lin, L.; Han, Y.; Yang, Z. Clinical observation on 103 patients of severe acute respiratory syndrome treated by integrative traditional Chinese and Western Medicine. Chin. J. Integr. Trad. Western Med. 2003, 23, 409–413. [Google Scholar] [CrossRef]
- Li, S.Y.; Chen, C.; Zhang, H.Q.; Guo, H.Y.; Wang, H.; Wang, L.; Zhang, X.; Hua, S.N.; Yu, J.; Xiao, P.G.; et al. Identification of natural compounds with antiviral activities against SARS-associated coronavirus. Antivir. Res. 2005, 67, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Jana, S.; Iram, S.; Thomas, J.; Hayat, M.Q.; Pannecouque, C.; Dehaen, W. Application of the triazolization reaction to afford dihydroartemisinin derivatives with anti-HIV activity. Molecules 2017, 22, 303. [Google Scholar] [CrossRef] [PubMed]
- Laila, U.; Akram, M.; Shariati, M.A.; Hashmi, A.M.; Akhtar, N.; Tahir, I.M.; Ghauri, A.O.; Munir, N.; Riaz, M.; Akhter, N.; et al. Role of medicinal plants in HIV/AIDS therapy. Clin. Exp. Pharmacol. Physiol. 2019, 46, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Lubbe, A.; Seibert, I.; Klimkait, T.; Van der Kooy, F. Ethnopharmacology in overdrive: The remarkable anti-HIV activity of Artemisia annua. J. Ethnopharmacol. 2012, 141, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Milbradt, J.; Auerochs, S.; Korn, K.; Marschall, M. Sensitivity of human herpesvirus 6 and other human herpesviruses to the broad-spectrum antiinfective drug artesunate. J. Clin. Virol. 2009, 46, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Naesens, L.; Bonnafous, P.; Agut, H.; De Clercq, E. Antiviral activity of diverse classes of broad-acting agents and natural compounds in HHV-6-infected lymphoblasts. J. Clin. Virol. 2006, 37, S69–S75. [Google Scholar] [CrossRef]
- Nagamune, K.; Moreno, S.N.; Sibley, L.D. Artemisinin-resistant mutants of Toxoplasma gondii have altered calcium homeostasis. Antimicrob. Agents Chemother. 2007, 51, 3816–3823. [Google Scholar] [CrossRef]
- Karamoddini, M.; Emami, S.; Ghannad, M.; Sani, E.; Sahebkar, A. Antiviral activities of aerial subsets of Artemisia species against Herpes Simplex virus type 1 (HSV1) in vitro. Asian Biomed. 2017, 5, 63–68. [Google Scholar] [CrossRef]
- Dai, R.; Xiao, X.; Peng, F.; Li, M.; Gong, G. Artesunate, an antimalarial drug, has a potential to inhibit HCV replication. Virus Genes 2016, 52, 22–28. [Google Scholar] [CrossRef]
- Paeshuyse, J.; Coelmont, L.; Vliegen, I.; Van hemel, J.; Vandenkerckhove, J.; Peys, E.; Sas, B.; De Clercq, E.; Neyts, J. Hemin potentiates the anti-hepatitis C virus activity of the antimalarial drug artemisinin. Biochem. Biophys. Res. Commun. 2006, 348, 139–144. [Google Scholar] [CrossRef]
- Qian, R.S.; Li, Z.; Yu, J.; Ma, D.J. The immunologic and antiviral effect of qinghaosu. J. Trad. Chin. Med. 1982, 2, 271–276. [Google Scholar]
- Romero, M.R.; Efferth, T.; Serrano, M.A.; Castano, B.; Macias, R.I.R.; Briz, O.; Marin, J.J.G. Effect of artemisinin/artesunate as inhibitors of hepatitis B virus production in an “In Vitro” replicative system. Antiviral. Res. 2005, 68, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.R.; Serrano, M.A.; Vallejo, M.; Efferth, T.; Alvarez, M.; Marin, J.J. Antiviral effect of artemisinin from Artemisia annua against a model member of the Flaviviridae family, the bovine viral diarrhoea virus (BVDV). Planta Med. 2006, 72, 1169–1174. [Google Scholar] [CrossRef] [PubMed]
- Haq, F.U.; Roman, M.; Ahmad, K.; Rahman, S.U.; Shah, S.M.; Suleman, N.; Ahmad, I.; Ullah, W. Artemisia annua: Trials are needed for COVID-19. Phytother. Res. 2020, 34, 2423–2424. [Google Scholar] [CrossRef] [PubMed]
- Canivet, C.; Menasria, R.; Rhéaume, C.; Piret, J.; Boivin, G. Valacyclovir combined with artesunate or rapamycin improves the outcome of herpes simplex virus encephalitis in mice compared to antiviral therapy alone. Antivir. Res. 2015, 123, 105–113. [Google Scholar] [CrossRef]
- Efferth, T. Beyond malaria: The inhibition of viruses by artemisinin-type compounds. Biotechnol. Adv. 2018, 36, 1730–1737. [Google Scholar] [CrossRef]
- Raffetin, A.; Bruneel, F.; Roussel, C.; Thellier, M.; Buffet, P.; Caumes, E.; Jauréguiberry, S. Use of artesunate in non-malarial indications. Med. Mal. Infect. 2018, 48, 238–249. [Google Scholar] [CrossRef]
- Shapira, M.Y.; Resnick, I.B.; Chou, S.; Neumann, A.U.; Lurain, N.S.; Stamminger, T.; Caplan, O.; Saleh, N.; Efferth, T.; Marschall, M.; et al. Artesunate as a potent antiviral agent in a patient with late drug-resistant cytomegalovirus infection after hematopoietic stem cell transplantation. Clin. Infect. Dis. 2008, 46, 1455–1457. [Google Scholar] [CrossRef]
- Flobinus, A.; Taudon, N.; Desbordes, M.; Labrosse, B.; Simon, F.; Mazeron, M.C.; Schnepf, N. Stability and antiviral activity against human cytomegalovirus of artemisinin derivatives. J. Antimicrob. Chemother. 2014, 69, 34–40. [Google Scholar] [CrossRef]
- Han, Y.; Pham, H.T.; Xu, H.; Quan, Y.; Mesplede, T. Antimalarial drugs and their metabolites are potent Zika virus inhibitors. J. Med. Virol. 2019, 91, 1182–1190. [Google Scholar] [CrossRef]
- Oiknine-Djian, E.; Bar-On, S.; Laskov, I.; Lantsberg, D.; Haynes, R.K.; Panet, A.; Wolf, D.G. Artemisone demonstrates synergistic antiviral activity in combination with approved and experimental drugs active against human cytomegalovirus. Antivir. Res. 2019, 172, 104639. [Google Scholar] [CrossRef] [PubMed]
- Oiknine-Djian, E.; Weisblum, Y.; Panet, A.; Wong, H.N.; Haynes, R.K.; Wolf, D.G. The artemisinin derivative artemisone is a potent inhibitor of human cytomegalovirus replication. Antimicrob. Agents Chemother. 2018, 62, e00288-18. [Google Scholar] [CrossRef] [PubMed]
- Obeid, S.; Alen, J.; Nguyen, V.H.; Pham, V.C.; Meuleman, P.; Pannecouque, C.; Le, T.N.; Neyts, J.; Dehaen, W.; Paeshuyse, J. Artemisinin analogues as potent inhibitors of in vitro hepatitis C virus replication. PLoS ONE 2013, 8, e81783. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, H.; Kim, S.; Shin, D.H.; Kim, M.S. Characteristics of flavonoids as potent MERS-CoV 3C-like protease inhibitors. Chem. Biol. Drug Design 2019, 94, 2023–2030. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzym. Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Solnier, J.; Fladerer, J.P. Flavonoids: A complementary approach to conventional therapy of COVID-19? Phytochem. Rev. 2021, 20, 773–795. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Quercetin: Antiviral significance and possible COVID-19 integrative considerations. Nat. Prod. Commun. 2020, 15, 1934578X20976293. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Agrawal, C.; Blunden, G. Rutin: A potential antiviral for repurposing as a SARS-CoV-2 main protease (Mpro) inhibitor. Nat. Prod. Commun. 2021, 16, 1934578X21991723. [Google Scholar] [CrossRef]
- Honscheid, A.; Rink, L.; Haase, H. T-lymphocytes: A target for stimulatory and inhibitory effects of zinc ions. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 132–144. [Google Scholar] [CrossRef]
- Brisibe, E.A.; Umoren, U.E.; Brisbe, F.; Magalhaes, P.M.; Ferreira, J.F.S.; Luthria, D.; Wu, X.; Prior, R.L. Nutritional characterisation and antioxidant capacity of different tissues of Artemisia annua L. Food Chem. 2009, 115, 1240–1246. [Google Scholar] [CrossRef]
- Liu, X.; Cao, J.; Huang, G.; Zhao, Q.; Shen, J. Biological activities of artemisinin derivatives beyond malaria. Curr. Top. Med. Chem. 2019, 19, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, C.; Gong, M.; Wang, M. Combination of artemisinin-based natural compounds from Artemisia annua L. for the treatment of malaria: Pharmacodynamic and pharmacokinetic studies. Phytother. Res. 2018, 32, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Rath, K.; Taxis, K.; Walz, G.; Gleiter, C.H.; Li, S.M.; Heide, L. Pharmacokinetic study of artemisinin after oral intake of a traditional preparation of Artemisia annua L (annual wormwood). Am. Trop. Med. Hyg. 2004, 70, 128–132. [Google Scholar] [CrossRef]
- Radulovic, N.S.; Randjelovic, P.J.; Stojanovic, N.M.; Blagojevic, P.D.; Stojanovic-Radic, Z.Z.; Ilic, I.R.; Djordjevic, V.B. Toxic essential oils. Part II: Chemical, toxicological, pharmacological and microbiological profiles of Artemisia annua L. volatiles. Food Chem. Toxicol. 2013, 58, 37–49. [Google Scholar] [CrossRef]
- Fuzimoto, A.D. An overview of the anti-SARS-CoV-2 properties of Artemisia annua, its antiviral action, protein-associated mechanisms, and repurposing for COVID-19 treatment. J. Integr. Med. 2021, 19, 375–388. [Google Scholar] [CrossRef]
- Zyad, A.; Tilaoui, M.; Jaafari, A.; Oukerrou, M.A.; Mouse, H.A. More insights into the pharmacological effects of artemisinin. Phytother. Res. 2018, 32, 216–229. [Google Scholar] [CrossRef]
- Law, S.; Leung, A.W.; Xu, C. Is the traditional Chinese herb “Artemisia annua” possible to fight against COVID-19? Integr. Med. Res. 2020, 9, 100474. [Google Scholar] [CrossRef]
- Bolarin, J.A.; Oluwatoyosi, M.A.; Orege, J.I.; Ayeni, E.A.; Ibrahim, Y.A.; Adeyemi, S.B.; Tiamiyu, B.B.; Gbadegesin, L.A.; Akinyemi, T.O.; Odoh, V.K.; et al. Therapeutic drugs for SARS-CoV-2 treatment: Current state and perspective. Int. Immunopharmacol. 2020, 90, 107228. [Google Scholar] [CrossRef]
- Yang, B.; Zhou, S.; Li, C.; Wang, Y. Toxicity and side effects of artemisiae annuae CQ-189. J. Chin. Mater. Med. 2010, 35, 204–207. [Google Scholar]
- Nordling, L. Unproven herbal remedy against COVID-19 could fuel drug-resistant malaria, scientists warn. Science 2020. [Google Scholar] [CrossRef]
- Kapepula, P.M.; Kabengele, J.K.; Kingombe, M.; Bambeke, F.V.; Tulkens, P.M.; Kishabongo, A.S.; Decloedt, E.; Zumla, A.; Tiberi, S.; Suloeman, F.; et al. Artemisia spp. derivatives for COVID-19 treatment: Anecdotal use, political hype, treatment potential, challenges, and road map to randomized clinical trials. Am. J. Trop. Med. Hyg. 2020, 103, 960–964. [Google Scholar] [CrossRef] [PubMed]
- Gendrot, M.; Duflot, I.; Boxberger, M.; Delandre, O.; Jardot, P.; Bideau, M.L.; Andreani, J.; Fonta, I.; Mosnier, J.; Rolland, C.; et al. Antimalarial artemisinin-based combination therapies (ACT) and COVID-19 in Africa: In vitro inhibition of SARS-CoV-2 replication by mefloquine-artesunate. Int. J. Infect. Dis. 2020, 99, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Danis, M. Proposal for treatment of malaria with Artemisia leaves. Bull. Acad. Natl. Med. 2019, 203, 122–123. [Google Scholar] [CrossRef]
- Nie, C.; Trimpert, J.; Moon, S.; Haag, R.; Gilmore, K.; Kaufer, B.B.; Seeberger, P.H. In vitro efficacy of Artemisia extracts against SARS-CoV-2. Virol. J. 2021, 18, 182. [Google Scholar] [CrossRef]
- Gilmore, K.; Zhou, Y.; Ramirez, S.; Pham, L.V.; Fanhoe, U.; Feng, S.; Offersgaard, A.; Trimpert, J.; Bukh, J.; Osterrieder, K.; et al. In vitro efficacy of Artemisinin-based treatments against SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Artemisia Derivative Affects Replication of SARS-CoV-2. Available online: https://www.medindia.net/news/extract-of-artemisia-affects-replication-of-sars-cov-2-199351-1.html (accessed on 10 April 2022).
- Artemisinin Raises Hopes and Fears amid COVID-19. Available online: https://cen.acs.org/biological-chemistry/infectious-disease/Artemisinin-raises-hopes-fears-amid-COVID-19/98/i21 (accessed on 10 April 2022).
- Nair, M.S.; Huang, Y.; Fidock, D.A.; Polyak, S.J.; Wagoner, J.; Towler, M.J.; Weathers, P.J. Artemisia annua L. extracts prevent in vitro replication of SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Nair, M.S.; Huang, Y.; Fidock, D.A.; Polyak, S.J.; Wagoner, J.; Towler, M.J.; Weathers, P.J. Artemisia annua L. extracts inhibit the in vitro replication of SARS-CoV-2 and two of its variants. J. Ethnopharmacol. 2021, 274, 114016. [Google Scholar] [CrossRef]
- Dandara, C.; Dzobo, K.; Chirikure, S. COVID-19 Pandemic and Africa: From the situation in Zimbabwe to a case for precision herbal medicine. Omics 2020, 25, 209–212. [Google Scholar] [CrossRef]
- Tih, F. WHO Holds Meeting with African Traditional Medicine Experts. 2020. Available online: https://www.aa.com.tr/en/africa/who-holds-meeting-with-african-traditional-medicine-experts/1838004 (accessed on 31 January 2021).
- World Health Organization. Regional Office for Africa. WHO Supports Scientifically-Proven Traditional Medicine. 2020. Available online: https://www.afro.who.int/news/who-supports-scientifically-proven-traditional-medicine (accessed on 31 January 2021).
- Zhou, Y.; Gilmore, K.; Ramirez, S.; Settels, E.; Gammeltoft, K.A.; Pham, L.V.; Fahnoe, U.; Feng, S.; Offersgaard, A.; Trimpert, J.; et al. In vitro efficacy of artemisinin-based treatments against SARS-CoV-2. Sci. Rep. 2021, 11, 14571. [Google Scholar] [CrossRef]
- Runestad, T. Available online: https://www.naturalproductsinsider.com/herbs-botanicals/herb-discovered-have-activity-against-sars-cov-2-virus (accessed on 10 November 2021).
- Nair, M.S.; Huang, Y.; Fidock, D.A.; Towler, M.J.; Weathers, P.J. Artemisia annua L. hot-water extracts show potent activity in vitro against Covid-19 variants including delta. J. Ethnopharmacol. 2022, 284, 114797. [Google Scholar] [CrossRef]
- MPIKG. Max Planck Institute for Colloids and Interfaces Press Release. Available online: https://www.mpikg.mpg.de/6288044/news_publicatio n_14663263_transferred?c=132305 (accessed on 10 May 2020).
- Trieu, V.; Saund, S.; Rahate, P.V.; Barge, V.B.; Naik, S.; Windlass, H.; Uckun, F.M. Targeting TGF-β pathway with COVID-19 drug candidate ARTIVeda/PulmoHeal accelerates recovery from mild-moderate COVID-19. Clin. Investig. 2021, 11, 10–18. [Google Scholar]
- Health Care. MGC Pharmaceutical’s (ASX:MXC) ArtemiC Combats COVID-19. Available online: https://themarketherald.com.au/mgc-pharmaceuticals-asxmgc-artemic-combats-covid-19-2020-12-15/ (accessed on 15 December 2020).
- Barnard, D.L.; Day, C.W.; Bailey, K.; Heiner, M.; Montgomery, R.; Lauridsen, L.; Chan, P.K.S.; Sidwell, R.W. Evaluation of immunomodulators, interferons and known in vitro SARS-coV inhibitors for inhibition of SARS-coV replication in BALB/c mice. Antivir. Chem. Chemother. 2006, 17, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Poisson-Benatouil, C. Action of Artemisia annua on Adaptive Immunity in COVID-19 Infections. Concept Note. 2020. Available online: https://inter-culturel.net/infections-au-covid-19-artemisia.html?lang=en&var_mode=calcul (accessed on 4 March 2022).
- De Biasi, S.; Meschiari, M.; Gibellini, L.; Bellinazzi, C.; Borella, R.; Fidanza, L.; Lo Tartaro, D.; Mattioli, M.; Paolini, A.; Menozzi, M.; et al. Marked T cell activation, senescence, exhaustion and skewing towards TH17 in patients with COVID-19 pneumonia. Nat. Commun. 2020, 11, 3434. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine storm in COVID-19: The current evidence and treatment strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Orege, J.I.; Adeyemi, S.B.; Tiamiyu, B.B.; Akinyemi, T.O.; Ibrahim, Y.A.; Orege, O.B. Artemisia and Artemisia-based products for COVID-19 management: Current state and future perspective. Adv. Tradit. Med. (ADTM) 2021, 5, 89. [Google Scholar] [CrossRef]
- Fan, H.H.; Wang, L.Q.; Liu, W.L.; An, X.P.; Liu, Z.D.; He, X.Q.; Song, L.H.; Tong, Y.G. Repurposing of clinically approved drugs for treatment of coronavirus disease 2019 in a 2019-novel coronavirus (2019-nCoV) related coronavirus model. Chin. Med. J. 2020, 133, 1051–1056. [Google Scholar] [CrossRef]
- Izoulet, M. Countries which primarily use antimalarial drugs as COVID-19 treatment see slower dynamic of daily deaths. SSRN Electron. J. 2020. [Google Scholar] [CrossRef]
- Xu, H.; He, Y.; Yang, X.; Liang, L.; Zhan, Z.; Ye, Y.; Yang, X.; Lian, F.; Sun, L. Antimalarial agent artesunate inhibits TNF-alpha-induced production of pro-inflammatory cytokines via inhibition of NF-kappaB and PI3 kinase/Akt signal pathway in human rheumatoid arthritis fibroblast-like synoviocytes. Rheumatology 2007, 46, 920–926. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, J.; Gu, Q.; Du, P.; Liang, H.; Dong, Q. Optimal temperature zone for dispersal of COVID-19. Sci. Total Environ. 2020, 16, 139487. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Wang, W.; Tian, D.S. Dysregulation of immune response in patients with Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef]
- Gendrot, M.; Andreani, J.; Boxberger, M.; Jardot, P.; Fonta, I.; Bideau, M.L.; Duflot, I.; Mosnier, J.; Rolland, C.; Bogreau, H.; et al. Antimalarial drugs inhibit the replication of SARS-CoV-2: An in vitro evaluation. Travel. Med. Infect. Dis. 2020, 37, 101873. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, V.M.; da Rocha, M.N.; Magalhães, E.P.; da Silva Mendes, F.R.; Marinho, M.M.; de Menezes, R.R.P.P.B.; Sampaio, T.L.; Dos Santos, H.S.; Martins, A.M.C.; Marinho, E.S. Computational approach towards the design of artemisinin-thymoquinone hybrids against main protease of SARS-COV-2. Future J. Pharm. Sci. 2021, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Instituto Nacional de Ciencias Medicas y Nutricion Salvador Zubiran. Camostat and Artemisia annua vs. Placebo in COVID-19 Outpatients. 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT04530617 (accessed on 20 March 2022).
- Read, S.A.; Obeid, S.; Ahlenstiel, C.; Ahlenstiel, G. The role of zinc in antiviral immunity. Adv. Nutr. 2019, 10, 696–710. [Google Scholar] [CrossRef] [PubMed]
- Bogan-Brown, K.; Nkrumah-Elie, Y.; Ishtiaq, Y.; Redpath, P.; Andrew Shao, A. Potential efficacy of nutrient supplements for treatment or prevention of COVID-19. J. Diet. Suppl. 2022, 17, 336–365. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T. Antiviral activities of Zn2+ ions for viral prevention, replication, capsid protein in intracellular proliferation of viruses. World Sci. News (WSN) 2018, 97, 28–50. [Google Scholar]
- Abreu, J.L. Artemisia annua + Zinc for the treatment of COVID-19 a potential successful combination therapy with Ivermectin. Daena-Inter. J. Good Conscienc. 2021, 16, 1–41. [Google Scholar]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; Mclellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef]
- Ulrich, H.; Pillat, M.M. CD147 as a target for COVID-19 treatment: Suggested effects of azithromycin and stem cell engagement. Stem Cell Rev. Rep. 2020, 16, 434–440. [Google Scholar] [CrossRef]
- Zumla, A.; Chan, J.F.W.; Azhar, E.I.; Hui, D.S.C.; Yuen, K.Y. Coronaviruses—Drug discovery and therapeutic options. Nat. Rev. Drug Discov. 2016, 15, 327–347. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V.; Hayashi, Y.; Jung, S.H. An overview of severe acute respiratory syndrome-coronavirus (SARS-CoV) 3CL protease inhibitors: Peptidomimetics and small molecule chemotherapy. J. Med. Chem. 2016, 59, 6595–6628. [Google Scholar] [CrossRef] [PubMed]
- Bzowka, M.; Mitusinska, K.; Raczynska, A.; Samol, A.; Tuszynski, J.; Gora, A. Molecular dynamics simulations indicate the SARS-CoV-2 Mpro is not a viable target for small-molecule inhibitors design. bioRxiv 2020. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro 1 from COVID-19 virus and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Yang, M.; Hong, Z.; Zhang, L.; Huang, Z.; Chen, X.; He, S.; Zhou, Z.; Zhou, Z.; Chen, Q.; et al. Crystal structure of SARS-CoV-2 nucleocapsid protein RNA binding domain reveals potential unique drug targeting sites. Acta Pharm. Sin. B 2020, 10, 1228–1238. [Google Scholar] [CrossRef]
- Ionescu, M.I. An overview of the crystallized structures of the SARS-CoV-2. Protein J. 2020, 39, 600–618. [Google Scholar] [CrossRef]
- Augustin, T.L.; Hajbabaie, R.; Harper, M.T.; Rahman, T. Novel small-molecule scaffolds as candidates against the SARS coronavirus 2 main protease: A fragment-guided in silico approach. Molecules 2020, 25, 5501. [Google Scholar] [CrossRef]
- Wambier, C.G.; Goren, A. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection is likely to be androgen mediated. J. Am. Acad. Dermatol. 2020, 83, 308–309. [Google Scholar] [CrossRef]
- Steely, A.M.; Willoughby, J.A.; Sundar, S.N.; Aivaliotis, V.I.; Firestone, G.L. Artemisinin disrupts androgen responsiveness of human prostate cancer cells by stimulating the 26S proteasome-mediated degradation of the androgen receptor protein. Anticancer Drugs 2017, 28, 1018–1031. [Google Scholar] [CrossRef]
- Ahmad, I.; Ali, R.; dos Santos Lopes, M.J.; Dino Steinmetz, C.H.; Ul Haq, F. Artemisia annua L. and its derivatives: Their antiviral effects on COVID-19 and possible mechanisms. J. Explor. Res. Pharmacol. 2022. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Z.Q.; Wang, C.Q.; Wang, L.S.; Meng, S.; Zhang, Y.C.; Chen, T.; Fan, Y.Q. Artemisinin inhibits extracellular matrix metalloproteinase inducer (EMMPRIN) and matrix metalloproteinase-9 expression via a protein kinase Cδ/p38/extracellular signal-regulated kinase pathway in phorbol myristate acetate-induced THP-1 macrophages. Clin. Exp. Pharmacol. Physiol. 2011, 38, 11–18. [Google Scholar] [CrossRef]
- Chen, W.; Li, S.; Li, J.; Zhou, W.; Wu, S.; Xu, S.; Cui, K.; Zhang, D.D.; Liu, B. Artemisitene activates the Nrf2-dependent antioxidant response and protects against bleomycin-induced lung injury. FASEB J. 2016, 30, 2500–2510. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.M.; Cho, H.Y.; Kleeberger, S.R. Oxidative stress and antioxidants in the pathogenesis of pulmonary fibrosis: A potential role for Nrf2. Antioxid. Redox Signal. 2008, 10, 321–332. [Google Scholar] [CrossRef] [PubMed]
- George, P.M.; Wells, A.U.; Jenkins, R.G. Pulmonary fibrosis and COVID-19: The potential role for antifibrotic therapy. Lancet Respir. Med. 2020, 8, 807–815. [Google Scholar] [CrossRef]
- Kim, M.H.; Seo, J.Y.; Kim, J.S. Artemisia annua L. extract ameliorates galactose-induced cognitive impairment in mice. Food Sci. Biotechnol. 2015, 24, 1901–1905. [Google Scholar] [CrossRef]
- Wang, C.; Xuan, X.; Yao, W.; Huang, G.; Jin, J. Anti-profibrotic effects of artesunate on bleomycin-induced pulmonary fibrosis in Sprague Dawley rats. Mol. Med. Rep. 2015, 12, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Uzun, T.; Toptas, O. Artesunate: Could be an alternative drug to chloroquine in COVID-19 treatment? Chin. Med. 2020, 15, 54. [Google Scholar] [CrossRef]
- Elkhodary, M.S.M. Treatment of COVID-19 by controlling the activity of the nuclear factor-kappa B. Cell Biol. 2020, 9, 109–121. [Google Scholar] [CrossRef]
- Moore, J.B.; June, C.H. Cytokine release syndrome in severe COVID-19. Science 2020, 368, 473–474. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Dadkhah, M.; Talei, S.; Gharari, Z.; Asghariazar, V.; Abdolmaleki, A.; Matin, S.; Molaei, S. Can anti-parasitic drugs help control COVID-19? Future Virol. 2022, 17, 315–339. [Google Scholar] [CrossRef]
- Farmanpour-Kalalagh, K.; Beyraghdar Kashkooli, A.; Babaei, A.; Rezaei, A.; van der Krol, A.R. Artemisinins in combating viral infections like SARS-CoV-2, inflammation and cancers and options to meet increased global demand. Front. Plant. Sci. 2022, 13, 780257. [Google Scholar] [CrossRef] [PubMed]
- Bestle, D.; Heindl, M.R.; Limburg, H.; Van Lam, T.; Pilgram, O.; Moulton, H.; Stein, D.A.; Hardes, K.; Eickmann, M.; Dolnik, O.; et al. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 23, e202000786. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cai, H.; Hu, J.; Lian, J.; Gu, J.; Zhang, S.; Ye, C.; Lu, Y.; Jin, C.; Yu, G.; et al. Epidemiological, clinical characteristics of cases of SARS-CoV-2 infection with abnormal imaging findings. Int. J. Infect. Dis. 2020, 94, 81–87. [Google Scholar] [CrossRef]
- Macchiagodena, M.; Pagliai, M.; Procacci, P. Identification of potential binders of the main protease 3CLpro of the COVID-19 via structure-based ligand design and molecular modeling. Chem. Phys. Lett. 2020, 750, 137489. [Google Scholar] [CrossRef]
- Mengist, H.M.; Dilnessa, T.; Jin, T. Structural Basis of Potential Inhibitors Targeting SARS-CoV-2 Main Protease. Front. Chem. 2021, 9, 622898. [Google Scholar] [CrossRef]
- Kumar, B.K.; Faheem; Chandra Sekhar, K.V.G.; Ojha, R.; Prajapati, V.K.; Pai, A.; Murugesan, S. Pharmacophore based virtual screening, molecular docking, molecular dynamics and MM-GBSA approach for identification of prospective SARS-CoV-2 inhibitor from natural product databases. J. Biomol. Struct. Dyn. 2022, 40, 1363–1386. [Google Scholar] [CrossRef]
- D’alessandro, S.; Scaccabarozzi, D.; Signorini, L.; Perego, F.; Ilboudo, D.P.; Ferrante, P.; Delbue, S. The use of antimalarial drugs against viral infection. Microorganisms 2020, 8, 85. [Google Scholar] [CrossRef]
- Uckun, F.M.; Saund, S.; Windlass, H.; Trieu, V. Repurposing anti-malaria phytomedicine artemisinin as a COVID-19 drug. Front. Pharmacol. 2021, 12, 649532. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [PubMed]
- Sehailia, M.; Chemat, S. In-silico studies of antimalarial-agent artemisinin and derivatives portray more potent binding to Lys353 and Lys31-binding hotspots of SARS-CoV-2 spike protein than hydroxychloroquine: Potential repurposing of artenimol for COVID-19. ChemRxiv 2020. preprint. [Google Scholar] [CrossRef]
- Cao, R.; Hu, H.; Li, Y.; Wang, X.; Xu, M.; Liu, J.; Zhang, H.; Yan, Y.; Zhao, L.; Li, W.; et al. Anti-SARS-CoV-2 potential of artemisinins In Vitro. ACS Infect. Dis. 2020, 6, 2524–2531. [Google Scholar] [CrossRef] [PubMed]
- Kangbai, J.B.; Babawo, L.S.; Kaitibi, D.; Sandi, A.A.; George, A.M.; Sahr, F. Re-reading ACT, BCG, and low COVID-19 in Africa. SN Compr. Clin. Med. 2021, 3, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yuan, M.; Li, H.; Deng, C.; Wang, Q.; Tang, Y.; Zhang, H.; Yu, W.; Xu, Q.; Zou, Y.; et al. Safety and efficacy of artemisinin-piperaquine for treatment of COVID-19: An open-label, non-randomised and controlled trial. Int. J. Antimicrob. Agents 2021, 57, 106216. [Google Scholar] [CrossRef]
- Sehailia, M.; Chemat, S. Antimalarial-agent artemisinin and derivatives portray more potent binding to Lys353 and Lys31-binding hotspots of SARS-CoV-2 spike protein than hydroxychloroquine: Potential repurposing of artenimol for COVID-19. J. Biomol. Struct. Dyn. 2021, 39, 6184–6194. [Google Scholar] [CrossRef]
- Gurung, A.B.; Ali, M.A.; Lee, J.; Farah, M.A.; Al-Anazi, K.M.; Al-Hemaid, F. Artesunate induces substantial topological alterations in the SARS-CoV-2 Nsp1 protein structure. J. King Saud. Univ. Sci. 2022, 3, 101810. [Google Scholar] [CrossRef]
- Rolta, R.; Kumar, V.; Sourirajan, A.; Dev, K. Phytocompounds of Rheum emodi, Thymus serpyllum and Artemisia annua inhibit COVID-19 binding to ACE2 receptor: In silico approach. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Das, S.; Sarmah, S.; Lyndem, S.; Roy, A.S. An investigation into the identification of potential inhibitors of SARS-CoV-2 main protease using molecular docking study. J. Biomol. Struct. Dyn. 2021, 39, 3347–3357. [Google Scholar] [CrossRef]
- Alsaffar, D.; Yaseen, A.; Jabal, G. In silico molecular docking studies of medicinal Arabic plant-based bioactive compounds as a promising drug candidate against COVID-19. Int. J. Innov. Sci. Res. Technol. 2020, 5, 876–896. [Google Scholar]
- Peele, K.A.; Potla Durthi, C.; Srihansa, T.; Krupanidhi, S.; Ayyagari, V.S.; Babu, D.J.; Indira, M.; Reddy, A.R.; Venkateswarulu, T.C. Molecular docking and dynamic simulations for antiviral compounds against SARS-CoV-2: A computational study. Inform. Med. Unlocked. 2020, 19, 100345. [Google Scholar] [CrossRef] [PubMed]
- Sudeep, H.V.; Gouthamchandra, K.; Shyamprasad, K. Molecular docking analysis of withaferin A from Withania somnifera with the glucose regulated protein 78 (GRP78) receptor and the SARS-CoV-2 main protease. Bioinformation 2020, 16, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Badraoui, R.; Saoudi, M.; Hamadou, W.S.; Elkahoui, S.; Siddiqui, A.J.; Alam, J.M.; Jamal, A.; Adnan, M.; Suliemen, A.M.E.; Alreshidi, M.M.; et al. Antiviral effects of artemisinin and its derivatives against SARS-CoV-2 main protease: Computational evidences and interactions with ACE2 allelic variants. Pharmaceuticals 2022, 15, 129. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.S.; Vanzara, A.G.; Patel, N.R.; Vasava, A.; Patil, S.; Rajput, K. Discovery of fungal metabolites Bergenin, Quercitrin and Dihydroartemisinin as potential inhibitors against main protease of SARS-CoV-2. ChemRxiv 2020. [Google Scholar] [CrossRef]
- Sharma, S.; Deep, S. In-silico drug repurposing for targeting SARS-CoV-2 main protease (Mpro). J. Biomol. Struct. Dyn. 2022, 40, 3003–3010. [Google Scholar] [CrossRef]
- Prashantha, C.N.; Gouthami, K.; Lavanya, L.; Bhavanam, S.; Jakhar, A.; Shakthiraju, R.G.; Suraj, V.; Sahana, K.V.; Sujana, H.S.; Guruprasad, N.M.; et al. Molecular screening of antimalarial, antiviral, anti-inflammatory and HIV protease inhibitors against spike glycoprotein of coronavirus. J. Mol. Graph. Model. 2021, 102, 107769. [Google Scholar] [CrossRef]
- Roy Chattopadhyay, N.; Chatterjee, K.; Banerjee, A.; Choudhuri, T. Combinatorial therapeutic trial plans for COVID-19 treatment armed up with antiviral, antiparasitic, cell-entry inhibitor, and immune-boosters. Virus Dis. 2020, 31, 479–489. [Google Scholar] [CrossRef]
- Alazmi, M.; Motwalli, O. Molecular basis for drug repurposing to study the interface of the S protein in SARS-CoV-2 and human ACE2 through docking, characterization, and molecular dynamics for natural drug candidates. J. Mol. Model. 2020, 26, 338. [Google Scholar] [CrossRef]
- Khan, R.J.; Jha, R.K.; Singh, E.; Jain, M.; Amera, G.M.; Singh, R.P.; Muthukumaran, J.; Singh, A.K. Identification of promising antiviral drug candidates against non-structural protein 15 (NSP15) from SARS-CoV-2: An in silico assisted drug-repurposing study. J. Biomol. Struct. Dyn. 2022, 40, 438–448. [Google Scholar] [CrossRef]
- Marak, B.N.; Dowarah, J.; Khiangte, L.; Singh, V.P. Step toward repurposing drug discovery for COVID-19 therapeutics through in silico approach. Drug Dev. Res. 2021, 82, 374–392. [Google Scholar] [CrossRef]
- Gupta, S.; Singh, V.; Varadwaj, P.K.; Chakravartty, N.; Katta, A.V.S.K.M.; Lekkala, S.P.; Thomas, G.; Narasimhan, S.; Reddy, A.R.; Reddy Lachagari, V.B. Secondary metabolites from spice and herbs as potential multitarget inhibitors of SARS-CoV-2 proteins. J. Biomol. Struct. Dyn. 2022, 40, 2264–2283. [Google Scholar] [CrossRef] [PubMed]
- Ribaudo, G.; Coghi, P.; Yang, L.J.; Ng, J.P.L.; Mastinu, A.; Memo, M.; Wong, V.K.W.; Gianoncelli, A. Computational and experimental insights on the interaction of artemisinin, dihydroartemisinin and chloroquine with SARS-CoV-2 spike protein receptor binding domain (RBD). Nat. Prod. Res. 2021, 33, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, C.; Wadhwa, A.; Kumari, A.; Hussain, F.; Jha, P.; Kaushik, N.K. In silico potential of approved antimalarial drugs for repurposing against COVID-19. OMICS 2020, 24, 568–580. [Google Scholar] [CrossRef]
- Dey, D.; Borkotoky, S.; Banerjee, M. In silico identification of tretinoin as a SARS-CoV-2 envelope (E) protein ion channel inhibitor. Comput. Biol. Med. 2020, 127, 104063. [Google Scholar] [CrossRef]
- Rai, K.K.; Apoorva; Sharma, L.; Pandey, N.; Meena, R.P.; Rai, S.P. Repurposing Artemisia annua L. flavonoids, artemisinin and its derivatives as potential drugs against novel coronavirus (SARS –nCoV) as revealed by in-silico studies. Int. J. Appl. Sci. Biotechnol. 2020, 8, 374–393. [Google Scholar] [CrossRef]
- Tang, Y.; Li, X.; Yuan, Y.; Zhang, H.; Zou, Y.; Xu, Z.; Xu, Q.; Song, J.; Deng, C.; Wang, Q. Network pharmacology-based predictions of active components and pharmacological mechanisms of Artemisia annua L. for the treatment of the novel Corona virus disease 2019 (COVID-19). BMC Complement. Med. Ther. 2022, 22, 56. [Google Scholar] [CrossRef]
- Zhan, Y.; Ta, W.; Tang, W.; Hua, R.; Wang, J.; Wang, C.; Lu, W. Potential antiviral activity of isorhamnetin against SARS-CoV-2 spike pseudotyped virus in vitro. Drug Dev. Res. 2021, 82, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Emirik, M. In silico inhibition potential of artemisinin derivatives against SARS-CoV-2 main protease. El-Cezerî J. Sci. Eng. 2021, 8, 809–816. [Google Scholar] [CrossRef]
- You, X.; Jiang, X.; Zhang, C.; Jiang, K.; Zhao, X.; Guo, T.; Zhu, X.; Bao, J.; Dou, H. Dihydroartemisinin attenuates pulmonary inflammation and fibrosis in rats by suppressing JAK2/STAT3 signaling. Aging (Albany NY) 2022, 14, 1110–1127. [Google Scholar] [CrossRef]
- Herrmann, L.; Yaremenko, I.; Çapcı, A.; Struwe, J.; Hodek, J.; Belyakova, Y.; Radulov, P.; Stepanov, G.; Weber, J.; Terentev, A.; et al. Artemisinin and quinoline hybrid compounds inhibit replication of SARS-CoV-2 In Vitro. ChemRxiv 2021. preprint. [Google Scholar] [CrossRef]
- Firestone, T.M.; Oyewole, O.O.; Reid, S.P.; Ng, C.L. Repurposing quinoline and artemisinin antimalarials as therapeutics for SARS-CoV-2: Rationale and implications. ACS Pharm. Transl. Sci. 2021, 4, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wu, F.; Xie, Z.; Song, X.; Song, X.; Zhu, Q.; Wei, J.; Tan, S.; Liang, L.; Gong, B. Clinical study of artesunate in the treatment of coronavirus disease 2019. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2020, 32, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, B.; Nanda, B.C.; Pradhan, G. Artesunate: An artemisinin derivative having antiviral properties with multiple pleotropic effects is a perfect potential agent for the treatment of symptomatic COVID-19 infection and related hyper inflammation states. J. Med. Sci. Clin. Res. 2020, 8, 215–225. [Google Scholar] [CrossRef]
- Bae, J.Y.; Lee, G.E.; Park, H.; Cho, J.; Kim, Y.E.; Lee, J.Y.; Ju, C.; Kim, W.K.; II Kim, J.; Park, M.-S. Pyronaridine and artesunate are potential antiviral drugs against COVID-19 and influenza. bioRxiv 2020. [Google Scholar] [CrossRef]
- Krishna, S.; Augustin, Y.; Wang, J.; Xu, C.; Staines, H.M.; Platteeuw, H.; Kamarulzaman, A.; Sall, A.; Kremsner, P. Repurposing antimalarials to tackle the COVID-19 pandemic. Trends Parasitol. 2021, 37, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Lyu, M.; Fan, G.; Xiao, G.; Wang, T.; Xu, D.; Gao, J.; Ge, S.; Li, Q.; Ma, Y.; Zhang, H.; et al. Traditional Chinese medicine in COVID-19. Acta Pharm. Sin. B 2021, 11, 3337–3363. [Google Scholar] [CrossRef]
- Rein, T. Harnessing autophagy to fight SARS-CoV-2: An update in view of recent drug development efforts. J. Cell Biochem. 2022, 123, 155–160. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, M.; Qin, H.; Lin, H.; An, X.; Shi, Z.; Song, L.; Yang, X.; Fan, H.; Tong, Y. Artemether, artesunate, arteannuin B, echinatin, licochalcone B and andrographolide effectively inhibit SARS-CoV-2 and related viruses In Vitro. Front. Cell Infect. Microbiol. 2021, 11, 680127. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agrawal, P.K.; Agrawal, C.; Blunden, G. RETRACTED: Artemisia Extracts and Artemisinin-Based Antimalarials for COVID-19 Management: Could These Be Effective Antivirals for COVID-19 Treatment? Molecules 2022, 27, 3828. https://doi.org/10.3390/molecules27123828
Agrawal PK, Agrawal C, Blunden G. RETRACTED: Artemisia Extracts and Artemisinin-Based Antimalarials for COVID-19 Management: Could These Be Effective Antivirals for COVID-19 Treatment? Molecules. 2022; 27(12):3828. https://doi.org/10.3390/molecules27123828
Chicago/Turabian StyleAgrawal, Pawan K., Chandan Agrawal, and Gerald Blunden. 2022. "RETRACTED: Artemisia Extracts and Artemisinin-Based Antimalarials for COVID-19 Management: Could These Be Effective Antivirals for COVID-19 Treatment?" Molecules 27, no. 12: 3828. https://doi.org/10.3390/molecules27123828
APA StyleAgrawal, P. K., Agrawal, C., & Blunden, G. (2022). RETRACTED: Artemisia Extracts and Artemisinin-Based Antimalarials for COVID-19 Management: Could These Be Effective Antivirals for COVID-19 Treatment? Molecules, 27(12), 3828. https://doi.org/10.3390/molecules27123828