
Review on the Pharmacological Properties of Phillyrin

 , and
, and
Abstract
:1. Introduction
2. Biology
3. Pharmacological Properties of Phillyrin
3.1. Effects on Metabolic Disorders
3.1.1. Obesity
3.1.2. Diabetic Nephropathy (DN)
3.2. Anti-Inflammatory Effects
3.2.1. NF-κB Signaling Pathway
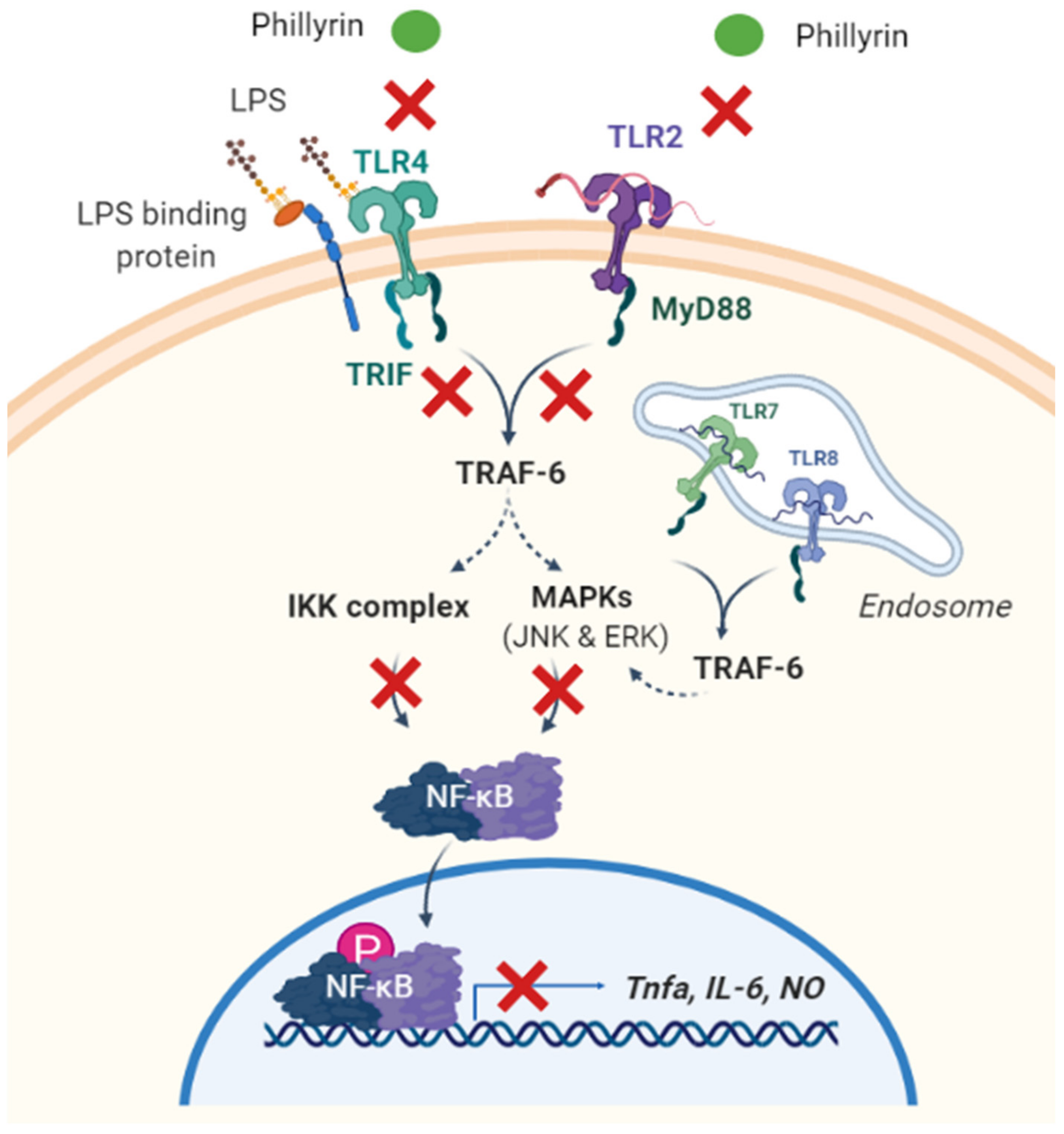
3.2.2. Toll-Like Receptors 4 (TLR4)
3.2.3. MAPK Signaling Pathway
3.3. Anti-Aging Effect
3.4. Antiviral Effects
3.5. Antibacterial Effects
3.6. Hepatoprotective Effects
3.7. Anti-Cancer Effects
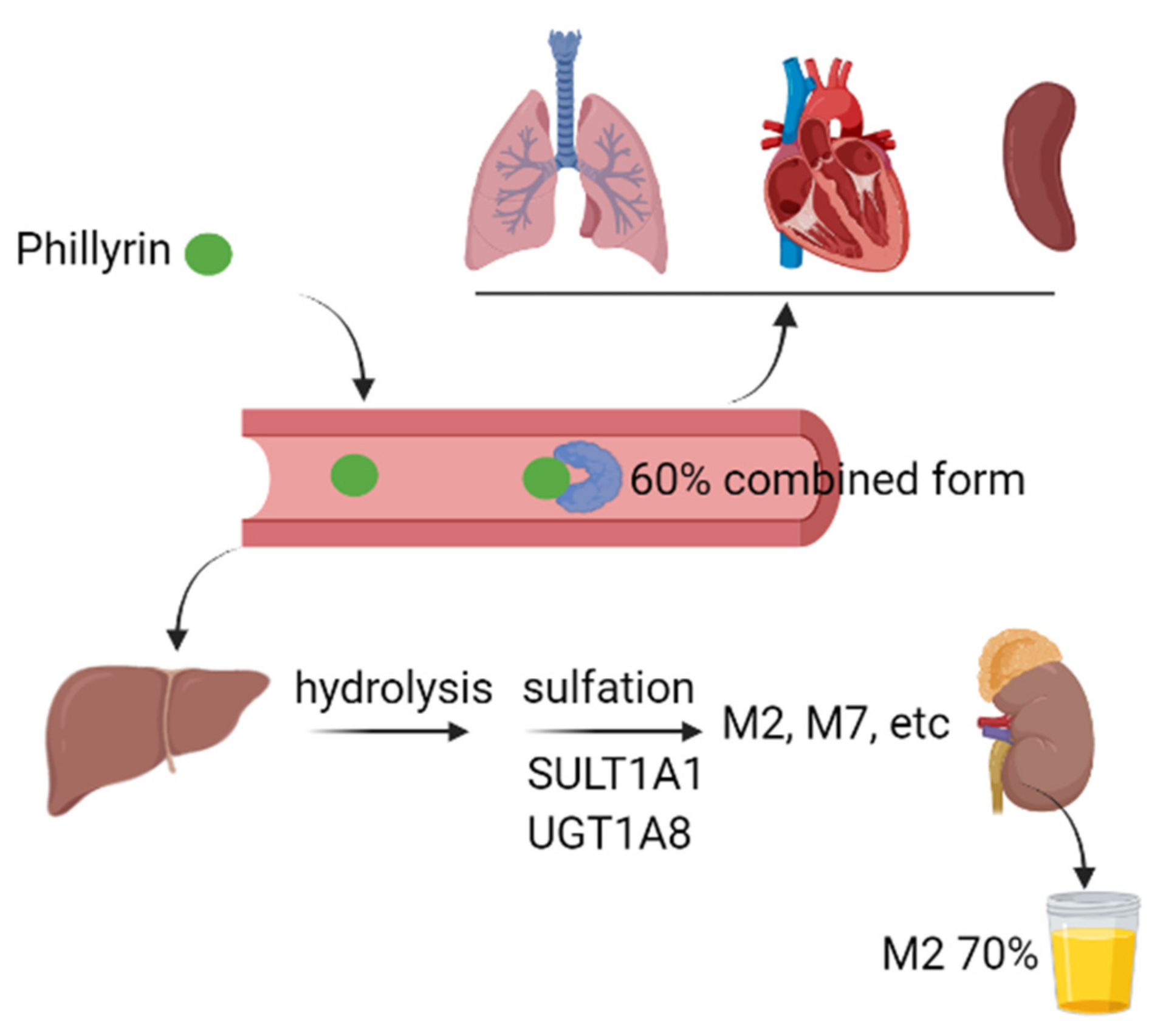
4. Metabolism and Tissue Distribution
5. Toxicology
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feng, Z.P.; Gao, X.Q.; Han, Y.C.; Wang, F.F.; Zhou, S.M.; Jiang, Y.X.; Wang, B.Q.; Tian, Q.C.; Cui, X.S. Study progress of Forsythia suspensa. Mod. Agric. Sci. Technol. 2018, 12, 60–62+64. [Google Scholar]
- Xia, W.; Dong, C.M.; Yang, C.F.; Chen, H. Advances in chemical constituents and pharmacology of Forsythia suspensa. Mod. Chin. Med. 2016, 18, 1670–1674. [Google Scholar]
- Bai, Y.; Li, J.; Liu, W.; Jiao, X.C.; He, J.; Liu, J.; Ma, L.; Gao, X.M.; Chang, Y.X. Pharmacokinetic of 5 components after oral administration of Fructus Forsythiae by HPLC-MS/MS and the effects of harvest time and administration times. J. Chromatogr. B 2015, 993, 36–46. [Google Scholar] [CrossRef]
- Fang, X.S.; Wang, Y.Z.; Wang, J.H.; Zhang, J.; Wang, X. Microwave-assisted extraction followed by RP-HPLC for the simultaneous extraction and determination of forsythiaside A, rutin, and phillyrin in the fruits of Forsythia suspensa. J. Sep. Sci. 2014, 36, 2672–2679. [Google Scholar] [CrossRef] [PubMed]
- Commission of Chinese Pharmacopoeia. Pharmacopoeia of the People’s Republic of China I; China Medical Science Press: Beijing, China, 2020; p. 178. [Google Scholar]
- Liao, X.; Hong, Y.; Chen, Z. Identification and quantification of the bioactive components in Osmanthus fragrans roots by HPLC-MS/MS. J. Pharm. Anal. 2020, 11, 299–307. [Google Scholar] [CrossRef]
- Sakamoto, S.; Machida, K.; Kikuchi, M. Lignan glycosides from the leaves of Osmanthus heterophyllus. J. Nat. Med. 2008, 62, 362–363. [Google Scholar] [CrossRef]
- Song, H.Y.; Jeong, D.E.; Lee, M. Bioactivity-guided extract optimization of Osmanthus fragrans var. aurantiacus leaves and anti-Inflammatory Activities of phillyrin. Plants 2021, 10, 1545. [Google Scholar] [CrossRef] [PubMed]
- Boyer, L.; Baghdikian, B.; Bun, S.S.; Taoubi, K.; Diaz-Lanza, A.; Elias, R.; Ollivier, E. Chionanthus virginicus L.: Phytochemical analysis and quality control of herbal drug and herbal preparations. Nat. Prod. Commun. 2011, 6, 753–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, J.H.; Kang, M.W.; Roh, J.H.; Choi, S.U.; Zee, O.P. Cytotoxic phenolic compounds from Chionanthus retusus. Arch. Pharmacal Res. 2009, 32, 1681–1687. [Google Scholar] [CrossRef]
- Song, Z.H.; Wang, Y.H.; Qian, Z.Z.; Smillie, T.J.; Khan, I.A. Quantitative determination of 10 phenylpropanoid and lignan compounds in Lancea tibetica by high-performance liquid chromatography with UV detection. Planta Med. 2011, 77, 1562–1566. [Google Scholar] [CrossRef]
- Hu, Q.H.; Yu, J.; Yang, W.J.; Kimatu, B.M.; Fang, Y.; Ma, N.; Pei, F. Identification of flavonoids from Flammulina velutipes and its neuroprotective effect on pheochromocytoma-12 cells. Food Chem. 2016, 204, 274–282. [Google Scholar] [CrossRef]
- Zhang, Q.M.; Wei, X.Y.; Wang, J.W. Phillyrin produced by Colletotrichum gloeosporioides, an endophytic fungus isolated from Forsythia suspensa. Fitoterapia 2012, 83, 1500–1505. [Google Scholar] [CrossRef]
- Editorial Board of Flora of China. Flora of China; Science Press: Beijing, China, 1978; p. 163. [Google Scholar]
- Wang, L.; He, X.L.; Zhang, H.X.; Li, Y.C.; Li, H.T.; Ji, H. Effects of producing area, harvesting period and drying method on the content of medicinal components in Forsythia suspensa. Hunan Agric. Sci. 2020, 2, 76–78. [Google Scholar]
- Hu, J.W. Evaluation of Whole Plant of Forsythia suspensa from Different Producing Areas Based on Differences of Active Ingredients; Henan University of Science and Technology: Luoyang, China, 2020. [Google Scholar]
- Ma, L.S.; Jia, J.P.; Zhang, Y.B.; Li, S.F.; Zhang, L.W. Comparative study on biological activity of different parts of Forsythia suspensa. Chem. Res. Appl. 2018, 30, 6. [Google Scholar]
- Li, R.Y.; Han, X.; Cheng, L.Z.; Tian, J. Comparative study on the content of phillyrin in the leaves of Green Fructus forsythiae and Grown Fructus forsythiae. J. Chang. Med. Coll. 2016, 30, 99–101. [Google Scholar]
- Wang, Z.Y.; Xia, Q.; Liu, X.; Liu, W.X.; Huang, W.Z.; Mei, X.; Luo, J.; Shan, M.X.; Ma, Z.Q.; Lin, R.C. Phytochemistry, pharmacology, quality control and future research of Forsythia suspensa (Thunb.) Vahl: A review. J. Ethnopharmacol. 2018, 210, 318–339. [Google Scholar] [CrossRef]
- Xiao, H.B.; Sui, G.G.; Lu, X.Y. Phillyrin lowers body weight in obese mice via the modulation of PPAR/-ANGPTL 4 pathway. Obes. Res. Clin. Pract. 2018, 12 (Suppl. S2), 71–79. [Google Scholar] [CrossRef]
- Xu, X.Q.; Saadeldeen, F.S.A.; Xu, L.T.; Zhao, Y.Y.; Wei, J.F.; Wang, H.D.; Liu, Z.H.; Kang, W.Y. The mechanism of phillyrin from the leaves of Forsythia suspensa for improving insulin resistance. BioMed Res. Int. 2019, 2019, 3176483. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, Q.C.; Gao, M.Y.; Zhu, X.P.; Chen, Z.T.; Wang, S.K.; Sun, G.J. Preventive effect of phillyrin extracted from Forsythia suspense leaves on obesity induced by high fat diet in rats. Food Sci. 2021, 42, 85–90. [Google Scholar]
- Jiang, Q.; Chen, J.; Long, X.B.; Yao, X.L.; Zou, X.; Yang, Y.P.; Huang, G.Y.; Zhang, H.Q. Phillyrin protects mice from traumatic brain injury by inhibiting the inflammation of microglia via PPARγ signaling pathway. Int. Immunopharmacol. 2020, 79, 106083. [Google Scholar] [CrossRef]
- Zhang, D.; Qi, B.Y.; Li, D.X.; Feng, J.L.; Huang, X.; Ma, X.H.; Huang, L.N.; Wang, X.Z.; Liu, X.Y. Phillyrin relieves lipopolysaccharide-induced AKI by protecting against glycocalyx damage and inhibiting inflammatory responses. Inflammation 2019, 43, 540–551. [Google Scholar] [CrossRef]
- Yang, L.L.; Zhou, X.J.; Huang, W.J.; Fang, Q.; Hu, J.L.; Yu, L.Z.; Ma, N.; Zhang, W.Q. Protective effect of phillyrin on lethal LPS-induced neutrophil inflammation in zebrafish. Cell. Physiol. Biochem. 2017, 43, 2074–2087. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.J.; Wu, M.F.; Xu, T. Study on the inhibitory effect of phillyrin on inflammation and its mechanism. Acta Univ. Med. Anhui 2020, 55, 1093–1097. [Google Scholar]
- Wang, Y.; Zhao, H.F.; Lin, C.X.; Ren, J.; Ye, Y.Y.; Ji, Z.H.; Zhang, S.Z. Inhibitory effect of phillyrin on inflammatory response of LPS-stimulated BV2 microglia. J. Apoplexy Nerv. Dis. 2016, 33, 338–341. [Google Scholar]
- Su, J.J.; Zhu, Y.C.; Zhang, W.L. Effects of phillyrin on p38 MAPK/C-FOS signaling pathway and osteoclast activation in periodontitis rats. J. Oral Sci. Res. 2021, 37, 33–38. [Google Scholar]
- Xie, X.Y.; Wei, X.H.; Li, T.Z.; Wang, W.L.; Gao, Y.; Yi, Q.; Wang, L. Effects of phillyrin on secretion of inflammatory factors by lipopolysaccharide mediated mammary epithelial cells. Cent. South Pharm. 2018, 16, 1379–1383. [Google Scholar]
- Li, J.H.; Yang, S.Q.; Liu, J.J.; Luo, M.M.; Chen, J.; Li, X.H. Inhibitory effect of phillyrin on LPS-induced activation of hepatic stellate cells. Acta Acad. Med. Mil. Tertiae 2020, 42, 342–349. [Google Scholar]
- Wang, J.H.; Wan, X.X.; Liu, D. Inhibition of phillyrin on the inflammatory response of human mononuclear macrophages stimulated by Staphylococcus aureus. J. Xinxiang Med. Coll. 2016, 33, 466–468. [Google Scholar]
- Zhou, N.X.; Li, P.; Shi, W.D.; Bai, S.P. Study on the therapeutic effect and mechanism of phillyrin on atherosclerosis model rats. Pharmacol. Clin. Chin. Mater. Med. 2016, 32, 28–33. [Google Scholar]
- Zhang, Z.; Zhang, Q.F.; Li, W.; Yang, C.Q.; Liu, B.L.; Li, D.M. Study on the effect of phillyrin on bone healing in traumatic fracture rat model. J. Immunol. 2019, 35, 71–76. [Google Scholar]
- Yan, Y.L.; Liu, M.J.; Yan, H.R.; Li, X.; Xu, J.H.; Yang, J.X. Study on antiaging effect of phillyrin in mice. Chin. Pharm. 2015, 26, 37–39. [Google Scholar]
- Qu, X.Y.; Li, Q.J.; Zhang, H.M.; Zhang, X.J.; Shi, P.H.; Zhang, X.J.; Yang, J.; Zhou, Z.; Wang, S.Q. Protective effects of phillyrin against influenza A virus in vivo. Arch. Pharmacal Res. 2016, 39, 998–1005. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, A.; Chu, W. Phillyrin is an effective inhibitor of quorum sensing with potential as an anti-Pseudomonas aeruginosa infection therapy. J. Vet. Med. Sci. 2019, 81, 473–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, S.; Fu, Q.D.; Fu, Y.X. Effects of phillyrin on Klebsiella pneumoniae infected mice. Chin. J. Clin. Pharmacol. 2021, 37, 2463–2467. [Google Scholar]
- Wu, C.; Wang, C.X.; Han, Y.J. Promoting effect of phillyrin on wound angiogenesis of perianal abscess in rats. Chin. J. Derm. 2020, 34, 1435–1442. [Google Scholar]
- Li, J.H. Study on Ameliorative Effect and Mechanism of Phillyrin on Liver Fibrosis; PLA Army Military Medical University: Shanghai, China, 2020. [Google Scholar]
- Liu, Y.H.; Qi, Z.L.; Xu, G.X.; He, L.; Yang, J.H. Protective effect of phillyrin on alcoholic liver injury. Chin. J. Clin. Pharmacol. Ther. 2016, 21, 6–9+15. [Google Scholar]
- Leng, W.; Liu, C.Y.; Shang, C.; Chen, M.X. Study on protective effect and mechanism of phillyrin on diabetic nephropathy rats. Chin. J. Immunol. 2019, 35, 2604–2608. [Google Scholar]
- Wang, T.Y.; Wen, X.J.; Zhang, Z.W.; Xie, M.J.; Zhou, J. Phillyrin ameliorates diabetic nephropathy through the PI3K/Akt/GSK-3β signalling pathway in streptozotocin-induced diabetic mice. Hum. Exp. Toxicol. 2021, 40 (Suppl. S12), S487–S496. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Jiang, Z.M. Effect of phillyrin on expression of VEGF and endostatin in Lewis lung cancer. Chin. J. Pathophysiol. 2016, 32, 167–171. [Google Scholar]
- Wang, D.H.; He, X.; He, Q. Combining use of phillyrin and autophagy blocker alleviates laryngeal squamous cell carcinoma via AMPK/mTOR/p70S6K signaling. Biosci. Rep. 2019, 39, BSR20190459. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.L.; Zhu, L.; Jiang, J.G. Active ingredients from natural botanicals in the treatment of obesity. Obes. Rev. 2014, 15, 957–967. [Google Scholar] [CrossRef]
- Li, L.Y. Identification and Validation of Weight Loss Targets of Phillyrin; Shanxi University: Taiyuan, China, 2019. [Google Scholar]
- Degerman, E.; Ahmad, F.; Chung, Y.W.; Guirguis, E.; Omar, B.; Stenson, L.; Manganiello, V. From PDE3B to the regulation of energy homeostasis. Curr. Opin. Pharmacol. 2011, 11, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.Q.; Quan, Y.Y.; Yu, L.Y.; Zheng, L.; Li, Y.X. Effects of forsythiaside on LPS-induced inflammatory response of RAW264.7 cells. Nat. Prod. Res. Dev. 2019, 31, 1117–1123. [Google Scholar]
- Zhong, W.T.; Wu, Y.C.; Xie, X.X.; Zhou, X.; Wei, M.M.; Soromou, L.W.; Ci, X.X.; Wang, D.C. Phillyrin attenuates LPS-induced pulmonary inflammation via suppression of MAPK and NF-κB activation in acute lung injury mice. Fitoterapia 2013, 90, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Guo, Y.H. Research progress on NF-κB mediated inflammatory pathway and gastrointestinal tumor. Smart Healthc. 2020, 6, 28–29. [Google Scholar]
- Mcguire, V.A.; Arthur, J.S.C. Subverting Toll-Like Receptor Signaling by Bacterial Pathogens. Front. Immunol. 2015, 6, 607. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.L. TLR4/NF-κB signaling pathway, autophagy and inflammatory pain. J. Gannan Med. Coll. 2019, 39, 984–988. [Google Scholar]
- Yue, S.J.; Zhang, P.X.; Zhu, Y.; Li, N.G.; Chen, Y.Y.; Li, J.J.; Zhang, S.; Jin, R.Y.; Yan, H.; Shi, X.Q.; et al. A ferulic acid derivative FXS-3 inhibits proliferation and metastasis of human lung cancer A549 cells via positive JNK signaling pathway and negative ERK/p38, AKT/mTOR and MEK/ERK signaling pathways. Molecules 2019, 24, 2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, J.M.; Nho, K.J.; Kim, H.S.; Lee, A.Y.; Moon, B.C.; Kim, H.K. An ethyl acetate fraction derived from Houttuynia cordata extract inhibits the production of inflammatory markers by suppressing NF-kB and MAPK activation in lipopolysaccharide-stimulated RAW 264.7 macrophages. BMC Complement. Altern. Med. 2014, 14, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, P.; Zhang, L.L.; Guo, Y.Y.; Lu, Y.L.; Li, D.P. Phillyrin, a natural lignan, attenuates tumor necrosis factor α-mediated insulin resistance and lipolytic acceleration in 3T3-L1 adipocytes. Planta Med. 2014, 80, 880–886. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, G.; Zhang, Q.Q.; Zhao, F.L.; Yu, X.L.; Ma, X.M.; Liu, M. Phillyrin attenuates osteoclast formation and function and prevents LPS-induced osteolysis in mice. Front. Pharmacol. 2019, 10, 1188. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.X.; Shi, L.; Liu, W.; Zhang, M.L.; Yang, J.X.; Li, F. Modern study on chemical constituents and pharmacological activity of Forsythia suspensa. J. Liaoning Univ. Tradit. Chin. Med. 2016, 18, 222–224. [Google Scholar]
- Hensel, A.; Bauer, R.; Heinrich, M.; Spiegler, V.; Kayser, O.; Hempel, G.; Kraft, K. Challenges at the time of COVID-19: Opportunities and innovations in antivirals from nature. Planta Med. 2020, 86, 659–664. [Google Scholar]
- Dmitrieva-Posocco, O.; Dzutsev, A.; Posocco, D.F.; Hou, V.; Yuan, W.; Thovarai, V.; Mufazalov, I.A.; Gunzer, M.; Shilovskiy, I.P.; Khaitov, M.R.; et al. Cell-type-specific responses to Interleukin-1 control microbial invasion and tumor-elicited inflammation in colorectal cancer. Immunity 2019, 50, 166–180. [Google Scholar] [CrossRef] [Green Version]
- Du, C.J.; Zhu, Q.Z.; Hao, J.; Wang, W. Determination of plasma protein binding rate of phillyrin by ultrafiltration combined with HPLC. Guangzhou Chem. 2022, 50, 3. [Google Scholar]
- Wang, H.; Zhang, X.; Jia, P.; Zhang, Y.; Tang, S.; Wang, H.; Li, S.; Yu, X.; Li, Y.; Zhang, L. Metabolic profile of phillyrin in rats obtained by UPLC-Q-TOF-MS. Biomed. Chromatogr. Int. J. Devoted Res. Chromatogr. Methodol. Appl. Biosci. 2016, 30, 913–922. [Google Scholar] [CrossRef]
- Pan, L.L.; Yang, Y.; Hui, M.; Wang, S.; Li, C.Y.; Zhang, H.; Ding, Y.H.; Fu, L.; Diao, X.X.; Zhong, D.F. Sulfation predominates the pharmacokinetics, metabolism, and excretion of forsythin in humans: Major enzymes and transporters identified. Acta Pharmacol. Sin. 2020, 42, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, Z.Q.; Zhang, M.B.; Sun, Y.T. Distributional difference of effective components of Flos lonicerae and Fructus Forsythiae Drug Pairs in fever SD rats organs. Liaoning J. Tradit. Chin. Med. 2018, 45, 1698–1700. [Google Scholar]
- Huang, Y.Q.; Lei, X.L.; Xiao, B.Q.; Sun, H.; Yang, W. Long-term toxicity test of phillyrin in beagle dogs. In Proceedings of the Sixth Annual Conference of Drug Toxicology, Chongqing, China, 27–30 October 2016. [Google Scholar]
- Lei, X.L.; Sun, H.; Lin, J.; Li, W.Q.; Yang, W. Study on acute toxicity of phillyrin to mice. In Proceedings of the Sixth Annual Conference of Drug Toxicology, Chongqing, China, 27–30 October 2016. [Google Scholar]
- Han, Z.; Lei, X.L.; Zhang, H.; Liu, L.; Chen, Z.S.; Yang, W.; Lun, Z.R. Evaluating the safety of forsythin from Forsythia suspensa leaves by acute and sub-chronic oral administration in rodent models. Asian Pac. J. Trop. Med. 2017, 10, 47–51. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Region | Reference |
|---|---|---|
| Oleaceae | ||
| Forsythia suspensa (Thunb.) Vahl | Japan, Korea, China | [1] |
| Osmanthus fragrans (Thunb.) Lour. | China | [6] |
| Osmanthus heterophyllus (G. Don) P. S. Green | Japan, China | [7] |
| Osmanthus fragrans var. aurantiacus Makino | China | [8] |
| Chionanthus virginicus L. | Japan, Korea | [9] |
| Chionanthus retusus Lindl. et Paxt. | Japan, Korea, China | [10] |
| Other sources | ||
| Lancea tibetica Hook. f. et Thoms | India, China | [11] |
| Flammulina velutipes (Curt. ex Fr.) Sing | Cultivated globally | [12] |
| Colletotrichum gloeosporioides (Penz.) Penz. et Sacc. | Distributed worldwide | [13] |
| Models | Mechanism | Reference |
|---|---|---|
| Effects on metabolic disorders | ||
| Mice fed with HFD | Phillyrin lowered body weight via the modulation of PPARβ/δ–ANGPTL 4 signaling pathway. | [20] |
| 3T3-L1 adipocytes | Phillyrin promoted glucose uptake in insulin resistance 3T3-L1 adipocyte through activation of PI3K/Akt signaling pathway. | [21] |
| Rats fed with HFD | Not available | [22] |
| Anti-inflammatory effects | ||
| Mouse model of traumatic brain injury | Phillyrin activated PPARγ signaling pathway to inhibit phosphorylation of NF-κB and its downstream pro-inflammatory action in microglia. | [23] |
| Mouse model of acute kidney injury induced by LPS | Phillyrin inhibited the activation of the NF-κB and MAPK signaling pathway, decreasing the levels of inflammatory cytokines (TNF-α, IL-1β, IL-6). | [24] |
| Lethal LPS-induced neutrophil Inflammation in zebrafish | Phillyrin reduced neutrophil infiltration, necrosis and inflammation via suppression of MyD88–NF-κB signaling pathway. | [25] |
| LPS-treated RAW264.7 cells; LPS-induced acute lung injury | Phillyrin inhibited the secretion of IL-6 and NO in RAW264.7 cells via TLR4 signaling pathway. | [26] |
| LPS-treated BV2 microglia cells | Phillyrin downregulated the expression of TLR4. | [27] |
| Periodontitis rats | Phillyrin reduced the phosphorylation of p38 MAPK and the expression of c-Fos. | [28] |
| LPS-treated mouse mammary epithelial cells | Phillyrin may decrease the secretion of inflammatory cytokines via inhibition of TLR4/MyD88/Traf-6/NIK or the TLR4/MyD88/Traf-6/IκB pathway. | [29] |
| LPS-treated rat hepatic stellate cells | Phillyrin restrained the expression of phosphorylated NF-κB p65 protein to inhibit HSC-T6 activation. | [30] |
| THP-1 cells stimulated with Staphylococcus aureus in vitro | Phillyrin inhibited the expression of TLR2 and TRL4. | [31] |
| Atherosclerosis in SD rats | Phillyrin reduced oxidative stress via decreasing gene and protein expression of NHE-1. | [32] |
| Traumatic fracture in SD rats | Phillyrin reduced the serum levels of inflammatory factors such as iNOS, TNF-α and IL-6. | [33] |
| Anti-aging effect | ||
| Mouse model of aging induced by D-galactose | Phillyrin enhanced the activity of SOD and decreased the activity of MAO-B to improve the ability of scavenging free radicals in mice to inhibit aging. | [34] |
| Antiviral effect | ||
| Mice infected with influenza A virus | Phillyrin may reduce inflammation induced by influenza A virus and inhibit viral replication. | [35] |
| Antibacterial effect | ||
| Caenorhabditis elegans–Pseudomonas aeruginosa infection model | Phillyrin possibly suppress pathogen virulence factors to protect Caenorhabditis elegans from Pseudomonas aeruginosa. | [36] |
| Klebsiella pneumonia (Kp) infected mice | Phillyrin activated STAT5/Foxp3 pathway in Kp infected mice to promote the balance of Th17/Treg cells and relieving the disease. | [37] |
| Wound surface of rats with perianal abscess | Phillyrin reduced the number of Escherichia coli in the wound of perianal abscess rats by activating JAK2/STAT3 pathway. | [38] |
| Hepatoprotective effects | ||
| Mouse model of liver fibrosis | Phillyrin inhibited NF-κB and TGF-β1/Smad2/3 signal pathway to repress the inflammatory response of macrophages and the activation of hepatic stellate. | [39] |
| Human liver cell line LO2 treated with alcohol | Phillyrin inhibited the expression of apoptosis related proteins PARP and Caspase 3. | [40] |
| Nephroprotective effects | ||
| Diabetic nephropathy in rats induced by HFD and streptozotocin | Phillyrin inhibited inflammation and alleviated renal injury associated with depressed TGF-β1 expression. | [41] |
| Diabetic nephropathy in mice induced by streptozotocin | Phillyrin suppressed renal cell apoptosis via activation of PI3K/Akt/GSK-3β signaling pathway in kidney. | [42] |
| Anti-cancer effects | ||
| Lewis lung carcinoma mice | Phillyrin inhibited lung tumor development by downregulating VEGF expression and upregulating endostatin expression, respectively. | [43] |
| HEp-2 cells | Phillyrin-induced autophagy may be through the AMPK/mTOR/p70S6K signaling pathway. | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, C.; Lu, M.; Cheng, J.; Rohani, E.R.; Hamezah, H.S.; Han, R.; Tong, X. Review on the Pharmacological Properties of Phillyrin. Molecules 2022, 27, 3670. https://doi.org/10.3390/molecules27123670
Zhou C, Lu M, Cheng J, Rohani ER, Hamezah HS, Han R, Tong X. Review on the Pharmacological Properties of Phillyrin. Molecules. 2022; 27(12):3670. https://doi.org/10.3390/molecules27123670
Chicago/Turabian StyleZhou, Chenyu, Mengya Lu, Jialei Cheng, Emelda Rosseleena Rohani, Hamizah Shahirah Hamezah, Rongchun Han, and Xiaohui Tong. 2022. "Review on the Pharmacological Properties of Phillyrin" Molecules 27, no. 12: 3670. https://doi.org/10.3390/molecules27123670
APA StyleZhou, C., Lu, M., Cheng, J., Rohani, E. R., Hamezah, H. S., Han, R., & Tong, X. (2022). Review on the Pharmacological Properties of Phillyrin. Molecules, 27(12), 3670. https://doi.org/10.3390/molecules27123670