
Production of 4-Deoxy-L-erythro-5-Hexoseulose Uronic Acid Using Two Free and Immobilized Alginate Lyases from Falsirhodobacter sp. Alg1

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Preparation of Crude and Recombinant AlyFRA and AlyFRB
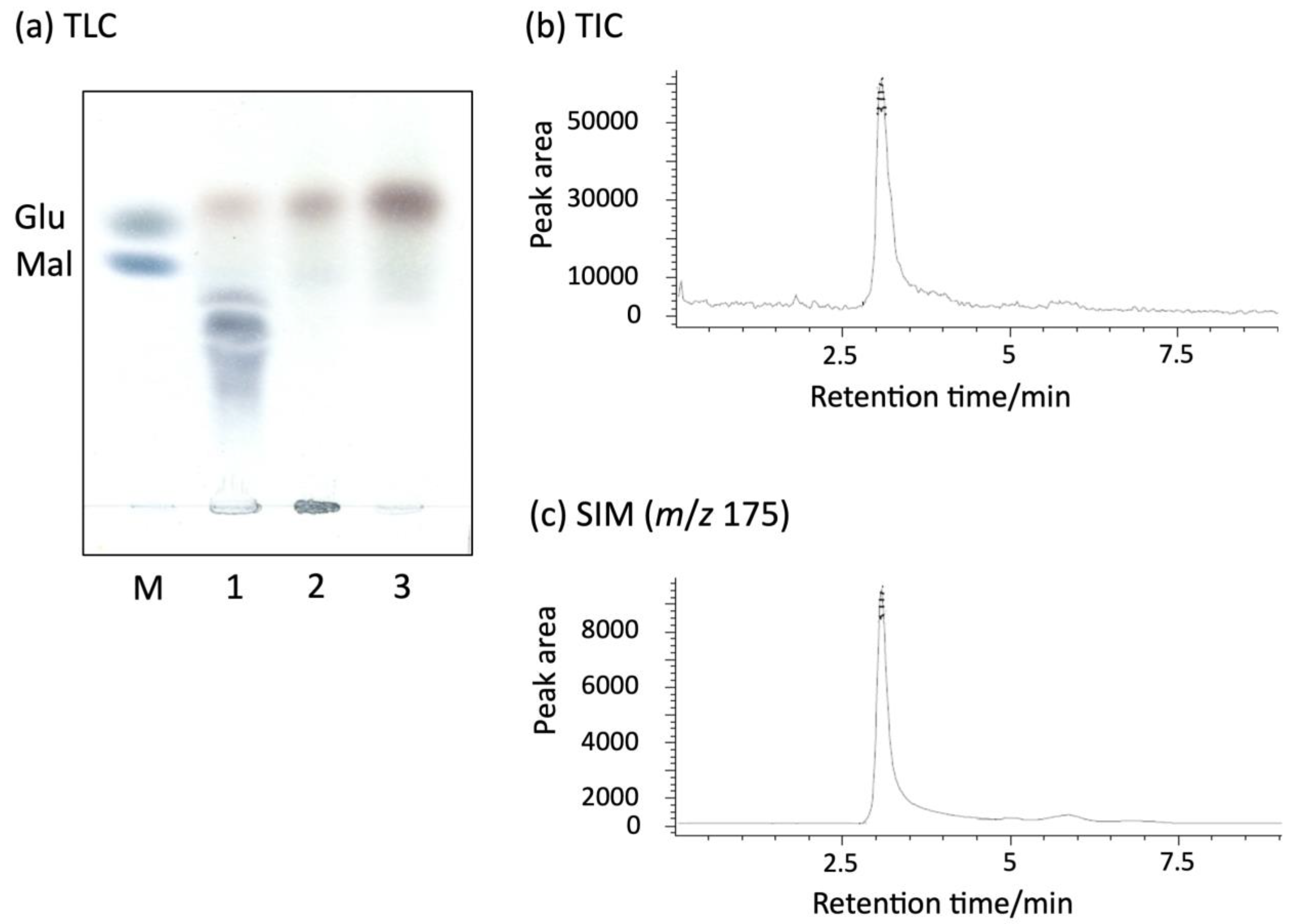
2.2. Enzyme Reaction Products
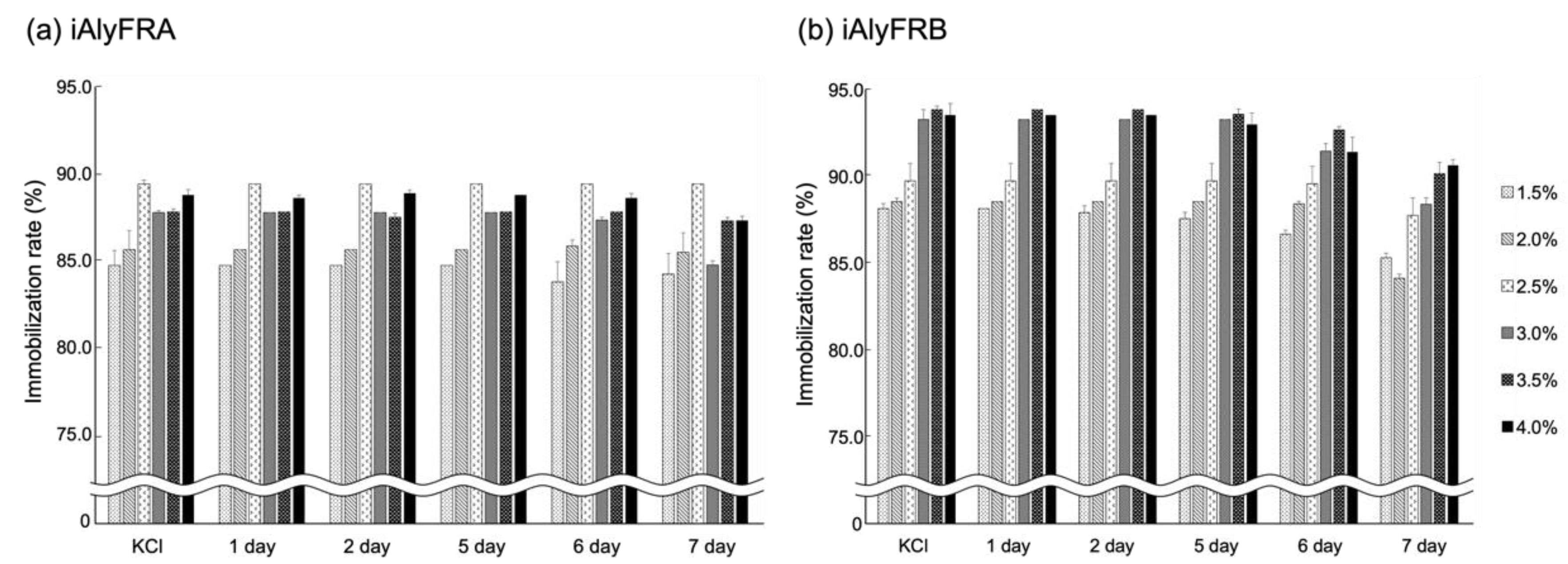
2.3. Immobilization Rate of AlyFRA and AlyFRB
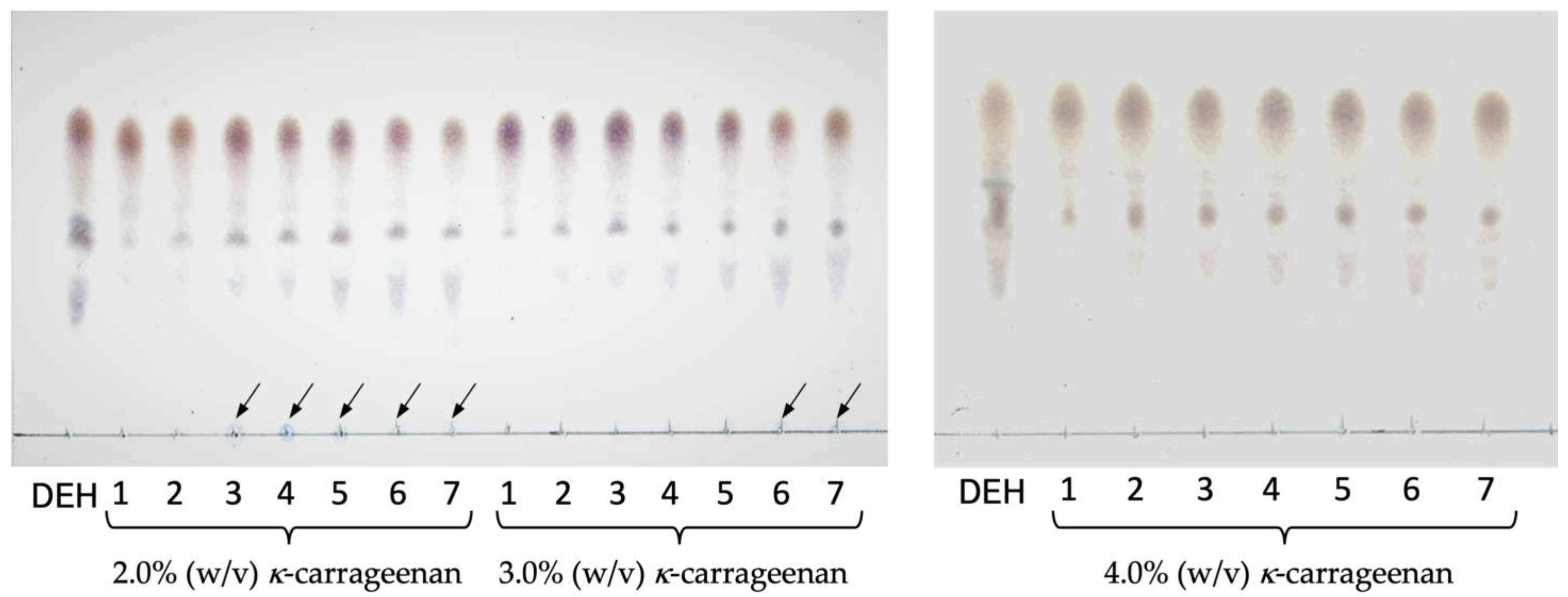
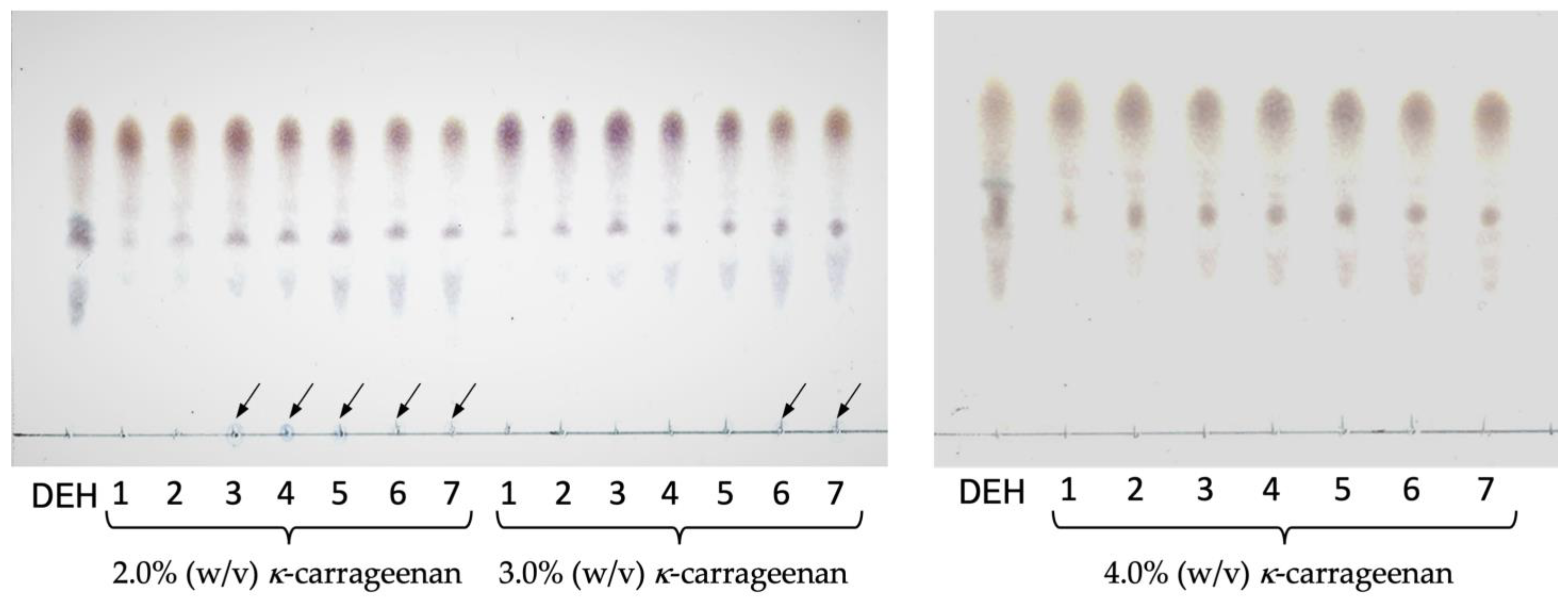
2.4. Batch Reaction of Immobilized Enzymes
3. Discussion
4. Materials and Methods
4.1. Overexpression and Purification of Recombinant AlyFRA and AlyFRB
4.2. TLC Analysis of Products
4.3. Identification of Products by LC/MS Analysis
4.4. Product Yield
4.5. Preparation of Immobilized Enzyme
4.6. Immobilization Rate of AlyFRA and AlyFRB
4.7. Batch Reaction of Immobilized Enzymes
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- FAO. Fishery and Aquaculture. Available online: https://www.fao.org/fishery/en/statistics/software/fishstatj/en (accessed on 1 March 2022).
- Kloareg, B.; Quatrano, R.S. Structure of the cell walls of marine algae and ecophysiological functions of the matrix polysaccharides. Oceanogr. Mar. Biol. Annu. Rev. 1988, 26, 259–315. [Google Scholar]
- Preiss, J.; Ashwell, G. Alginic acid metabolism in bacteria: I. Enzymatic formation of unsaturated oligosaccharides and 4-deoxy-L-erythro-5-hexoseulose uronic acid. J. Biol. Chem. 1962, 237, 309–316. [Google Scholar] [CrossRef]
- Hobbs, J.K.; Lee, S.M.; Robb, M.; Hof, F.; Barr, C.; Abe, K.T.; Hehemann, J.H.; McLean, R.; Abbott, D.W.; Boraston, A.B. KdgF, the missing link in the microbial metabolism of uronate sugars from pectin and alginate. Proc. Natl. Acad. Sci. USA 2016, 113, 6188–6193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, S.; Hashimoto, W. 4-Deoxy-L-erythro-5-hexoseulose uronate (DEH) and DEH reductase: Key molecule and enzyme for the metabolism and utilization of alginate. Molecules 2022, 27, 338. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Takahashi, M.; Tanaka, R.; Shibata, T.; Kuroda, K.; Ueda, M.; Takeyama, H. Draft genome sequence of Falsirhodobacter sp. strain alg1, an alginate-degrading bacterium isolated from fermented brown algae. Genome Announc. 2014, 2, e00826-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, T.; Takahashi, M.; Tanaka, R.; Miyake, H.; Shibata, T.; Chow, S.; Kuroda, K.; Ueda, M.; Takeyama, H. Falsirhodobacter sp. alg1 harbors single homologs of endo and exo-type alginate lyases efficient for alginate depolymerization. PLoS ONE 2016, 11, e0155537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wargacki, A.J.; Leonard, E.; Win, M.N.; Regitsky, D.D.; Santos, C.N.; Kim, P.B.; Cooper, S.R.; Raisner, R.M.; Herman, A.; Sivitz, A.B.; et al. An engineered microbial platform for direct biofuel production from brown macroalgae. Science 2012, 335, 308–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, C.N.S.; Regitsky, D.D.; Yoshikuni, Y. Implementation of stable and complex biological systems through recombinase-assisted genome engineering. Nat. Commun. 2013, 4, 2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enquist-Newman, M.; Faust, A.M.E.; Bravo, D.D.; Santos, C.N.S.; Raisner, R.M.; Hanel, A.; Sarvabhowman, P.; Le, C.; Regitsky, D.D.; Cooper, S.R.; et al. Efficient ethanol production from brown macroalgae sugars by a synthetic yeast platform. Nature 2014, 505, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Yokoi, T.; Shibata, T.; Morisaka, H.; Kuroda, K.; Ueda, M. Engineered yeast whole-cell biocatalyst for direct degradation of alginate from macroalgae and production of non-commercialized useful monosaccharide from alginate. Appl. Microbiol. Biotechnol. 2016, 100, 1723–1732. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Sasaki, Y.; Motone, K.; Shibata, T.; Tanaka, R.; Miyake, H.; Mori, T.; Kuroda, K.; Ueda, M. Construction of bioengineered yeast platform for direct bioethanol production from alginate and mannitol. Appl. Microbiol. Biotechnol. 2017, 101, 6627–6636. [Google Scholar] [CrossRef] [PubMed]
- Takeda, H.; Yoneyama, F.; Kawai, S.; Hashimoto, W.; Murata, K. Bioethanol production from marine biomass alginate by metabolically engineered bacteria. Energy Environ. Sci. 2011, 4, 2575–2581. [Google Scholar] [CrossRef]
- Wang, D.M.; Kim, H.T.; Yun, E.J.; Kim, D.H.; Park, Y.C.; Woo, H.C.; Kim, K.H. Optimal production of 4-deoxy-L-erythro-5-hexoseulose uronic acid from alginate for brown macro algae saccharification by combining endo- and exo-type alginate lyases. Bioprocess Biosyst. Eng. 2014, 37, 2105–2111. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takata, I.; Tosa, T.; Chibata, I. Screening of matrix suitable for immobilization of microbial cells. J. Solid-Phase Biochem. 1977, 2, 225–236. [Google Scholar] [CrossRef]
- Pace, C.N.; Vajdos, F.; Fee, L.; Grimsley, G.; Gray, T. How to measure and predict the molar absorption coefficient of a protein. Protein Sci. 1995, 4, 2411–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, R.W.; Bourne, E.J. Colour reactions given by sugars and diphenylamine-aniline spray reagents on paper chromatograms. J. Chromatogr. 1960, 4, 206–213. [Google Scholar] [CrossRef]
- Shibata, T.; Fujii, R.; Miyake, H.; Tanaka, R.; Mori, T.; Takahashi, M.; Takagi, T.; Yoshikawa, H.; Kuroda, K.; Ueda, M. Development of an analysis method for 4-deoxy-L-erythro-5-hexoseulose uronic acid by LC/ESI/MS with selected ion monitoring. Nat. Prod. Commun. 2017, 12, 1934578X1701200627. [Google Scholar] [CrossRef] [Green Version]
- Borges, D.G.; Junior, A.B.; Farinas, C.S.; Giordano, R.D.L.C.; Tardioli, P.W. Enhanced saccharification of sugarcane bagasse using soluble cellulase supplemented with immobilized β-glucosidase. Bioresour. Technol. 2014, 167, 206–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| rAlyFRA | rAlyFRB | Yield (%) |
|---|---|---|
| + 2 | + | 84.8 ± 5.2 |
| + | ++ | 96.8 ± 1.2 |
| + | +++ | 81.8 ± 3.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, Y.; Murase, Y.; Shibata, T.; Tanaka, R.; Mori, T.; Miyake, H. Production of 4-Deoxy-L-erythro-5-Hexoseulose Uronic Acid Using Two Free and Immobilized Alginate Lyases from Falsirhodobacter sp. Alg1. Molecules 2022, 27, 3308. https://doi.org/10.3390/molecules27103308
Tanaka Y, Murase Y, Shibata T, Tanaka R, Mori T, Miyake H. Production of 4-Deoxy-L-erythro-5-Hexoseulose Uronic Acid Using Two Free and Immobilized Alginate Lyases from Falsirhodobacter sp. Alg1. Molecules. 2022; 27(10):3308. https://doi.org/10.3390/molecules27103308
Chicago/Turabian StyleTanaka, Yuzuki, Yoshihiro Murase, Toshiyuki Shibata, Reiji Tanaka, Tetsushi Mori, and Hideo Miyake. 2022. "Production of 4-Deoxy-L-erythro-5-Hexoseulose Uronic Acid Using Two Free and Immobilized Alginate Lyases from Falsirhodobacter sp. Alg1" Molecules 27, no. 10: 3308. https://doi.org/10.3390/molecules27103308
APA StyleTanaka, Y., Murase, Y., Shibata, T., Tanaka, R., Mori, T., & Miyake, H. (2022). Production of 4-Deoxy-L-erythro-5-Hexoseulose Uronic Acid Using Two Free and Immobilized Alginate Lyases from Falsirhodobacter sp. Alg1. Molecules, 27(10), 3308. https://doi.org/10.3390/molecules27103308