
Antiproliferative Properties of Triterpenoids by ECIS Method—A New Promising Approach in Anticancer Studies?

 , , ,
, , , 
Abstract
:
1. Introduction
φ = arctan (X/R)
2. Results
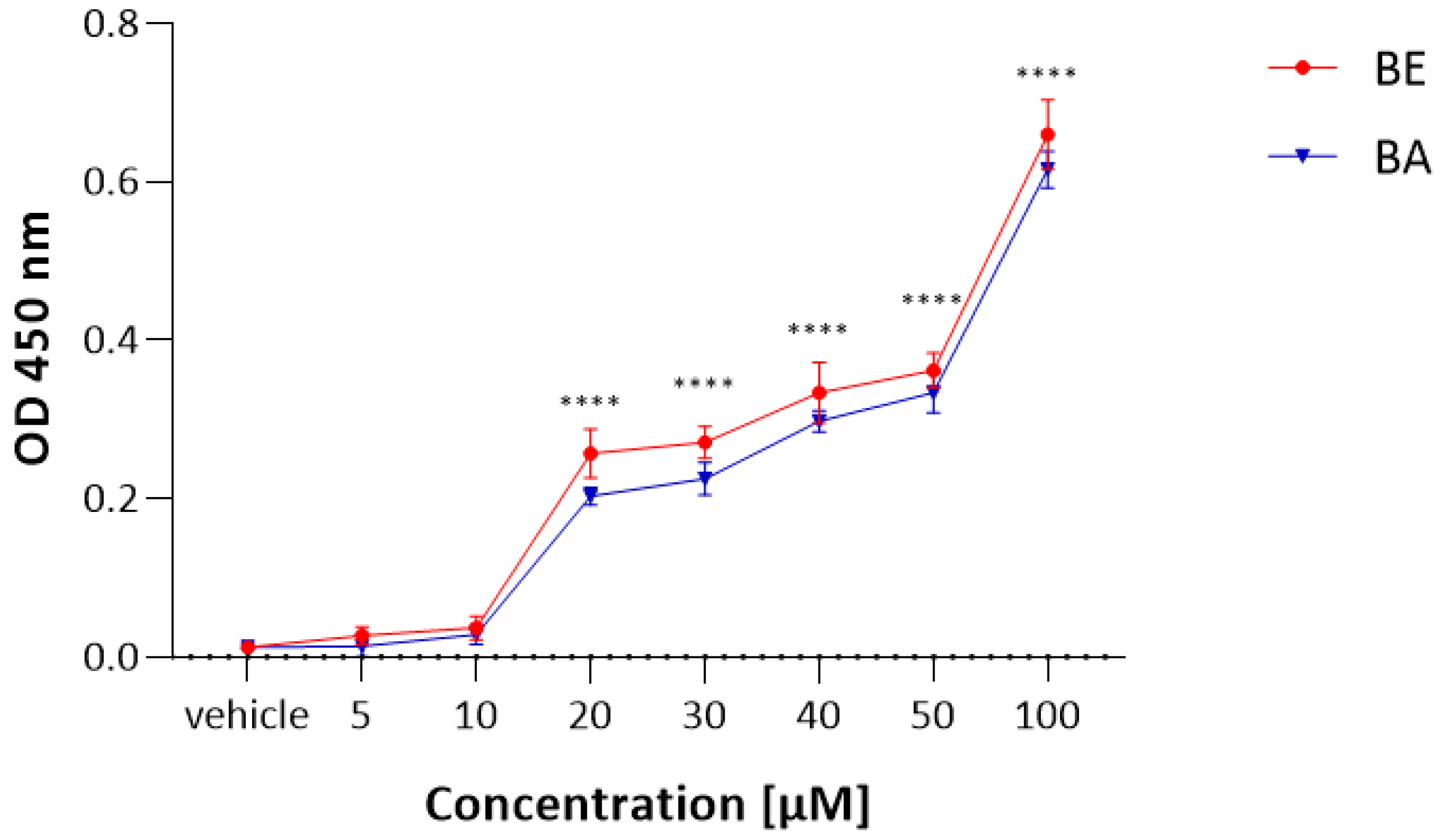
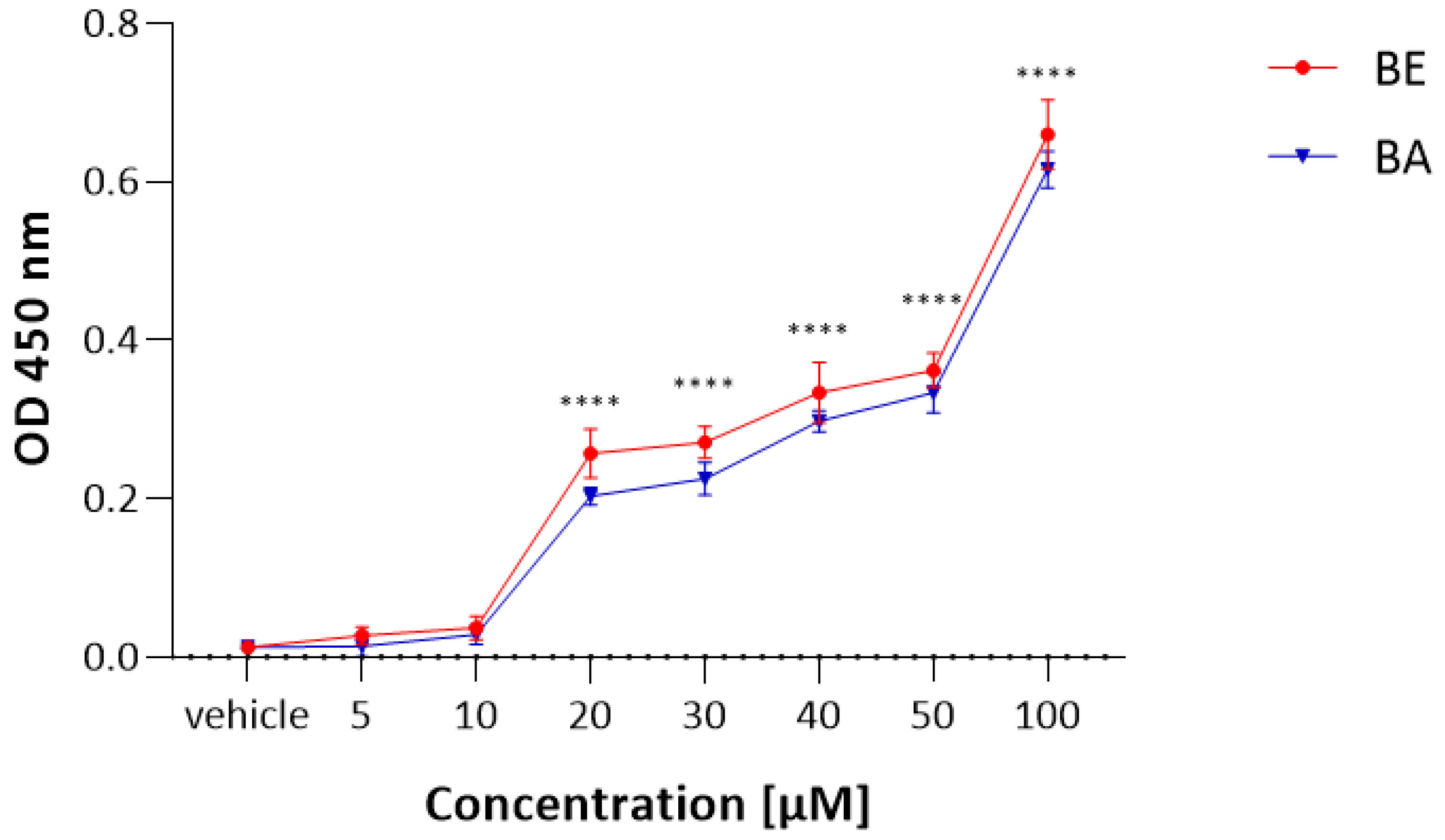
2.1. The MTT Results
2.2. The LDH Results
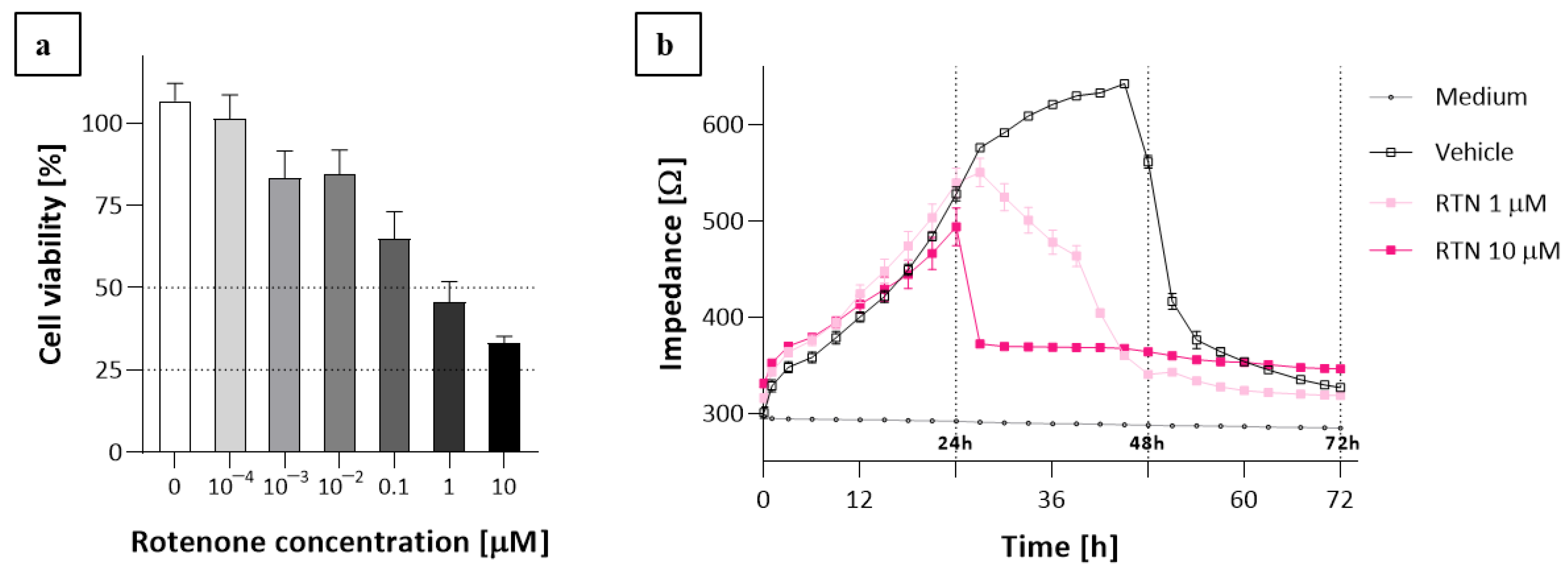
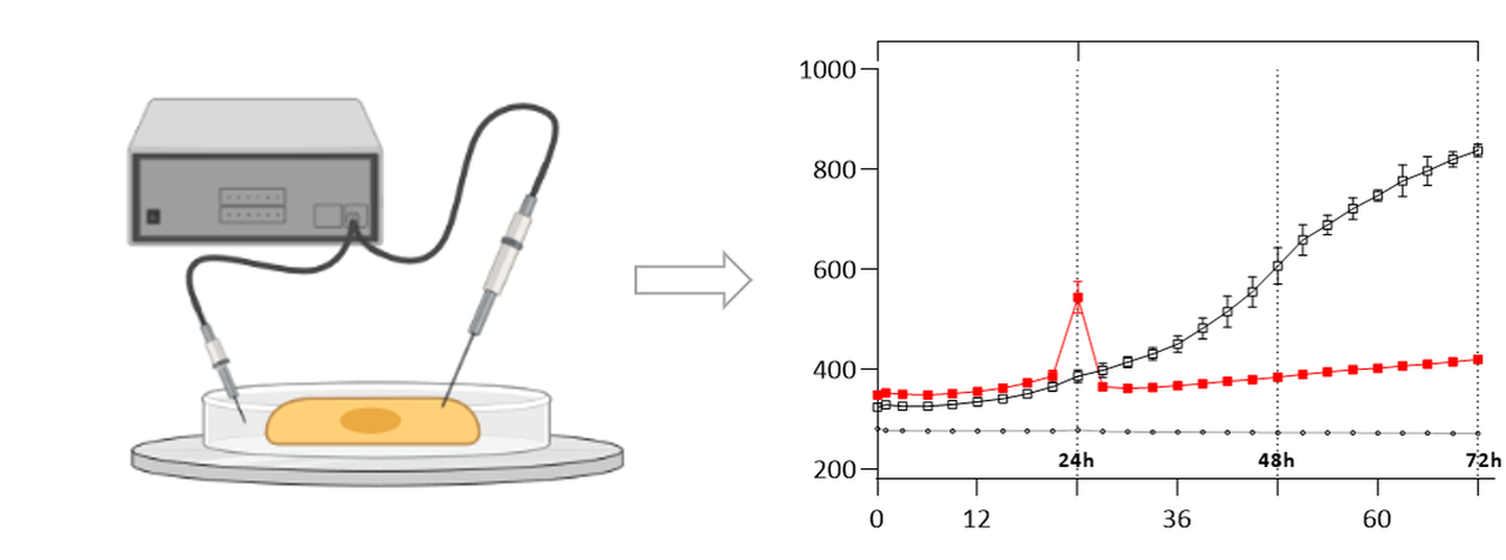
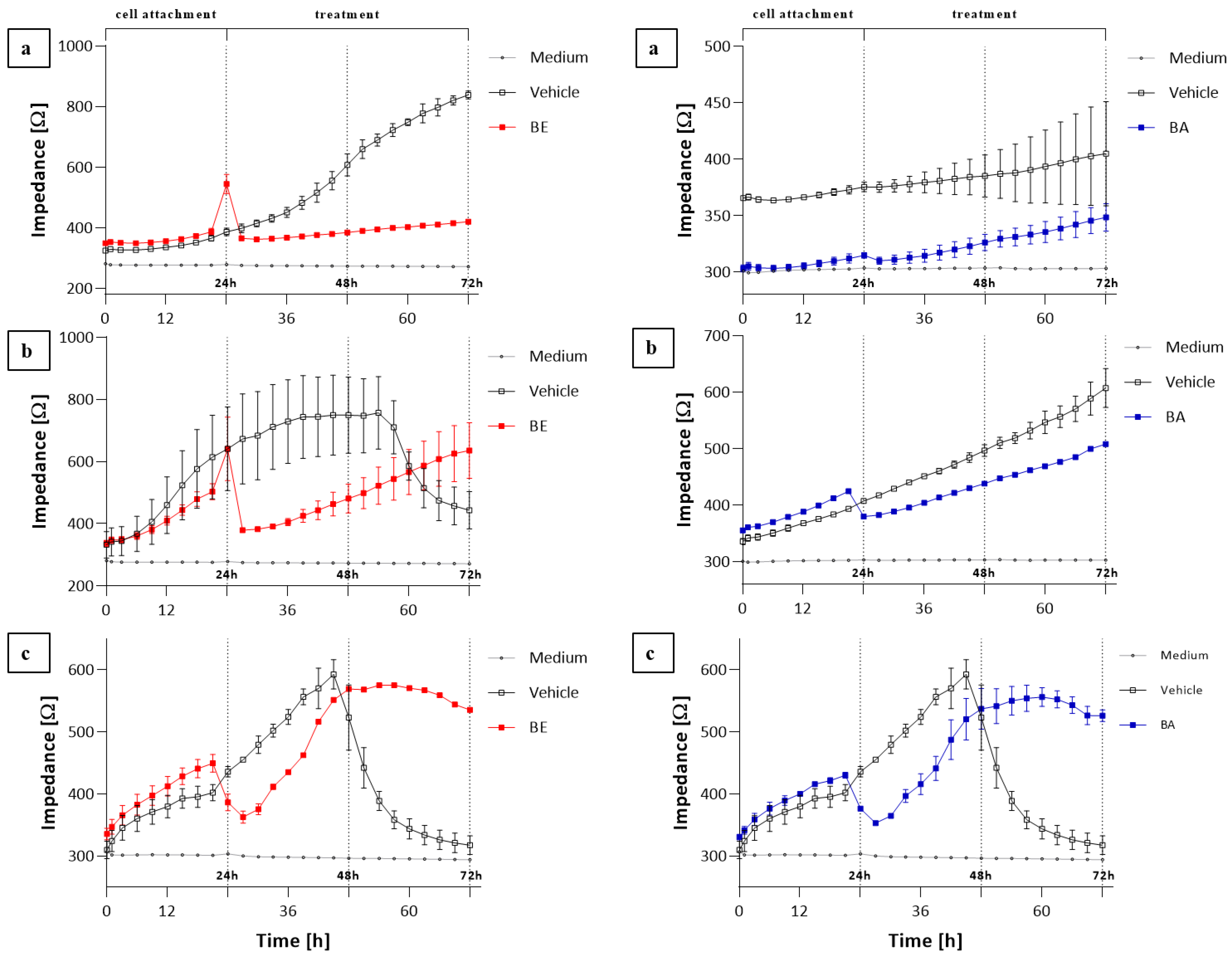
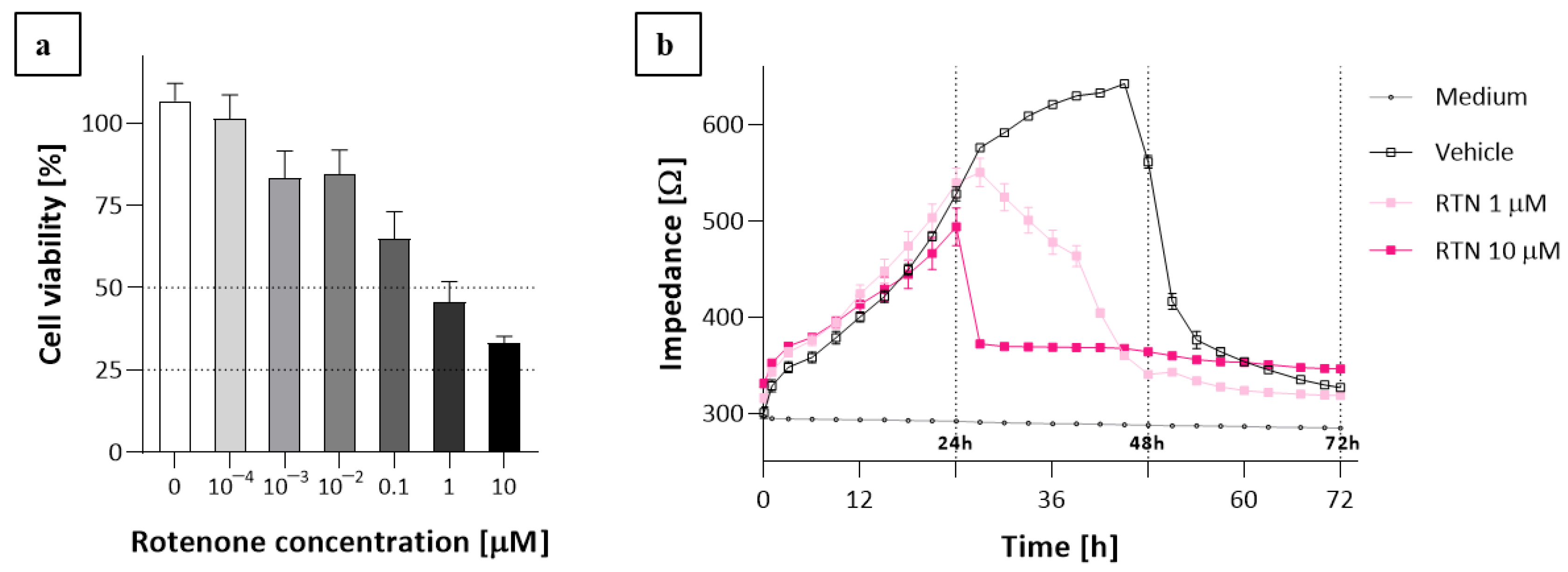
2.3. The ECIS Results
3. Discussion
4. Material and Methods
4.1. Preparation of Betulin and Betulinic Acid
4.2. Cell Lines Cultures
4.3. The Cell Proliferation Assay—MTT Assay
4.4. Cytotoxicity Assay—LDH Assay
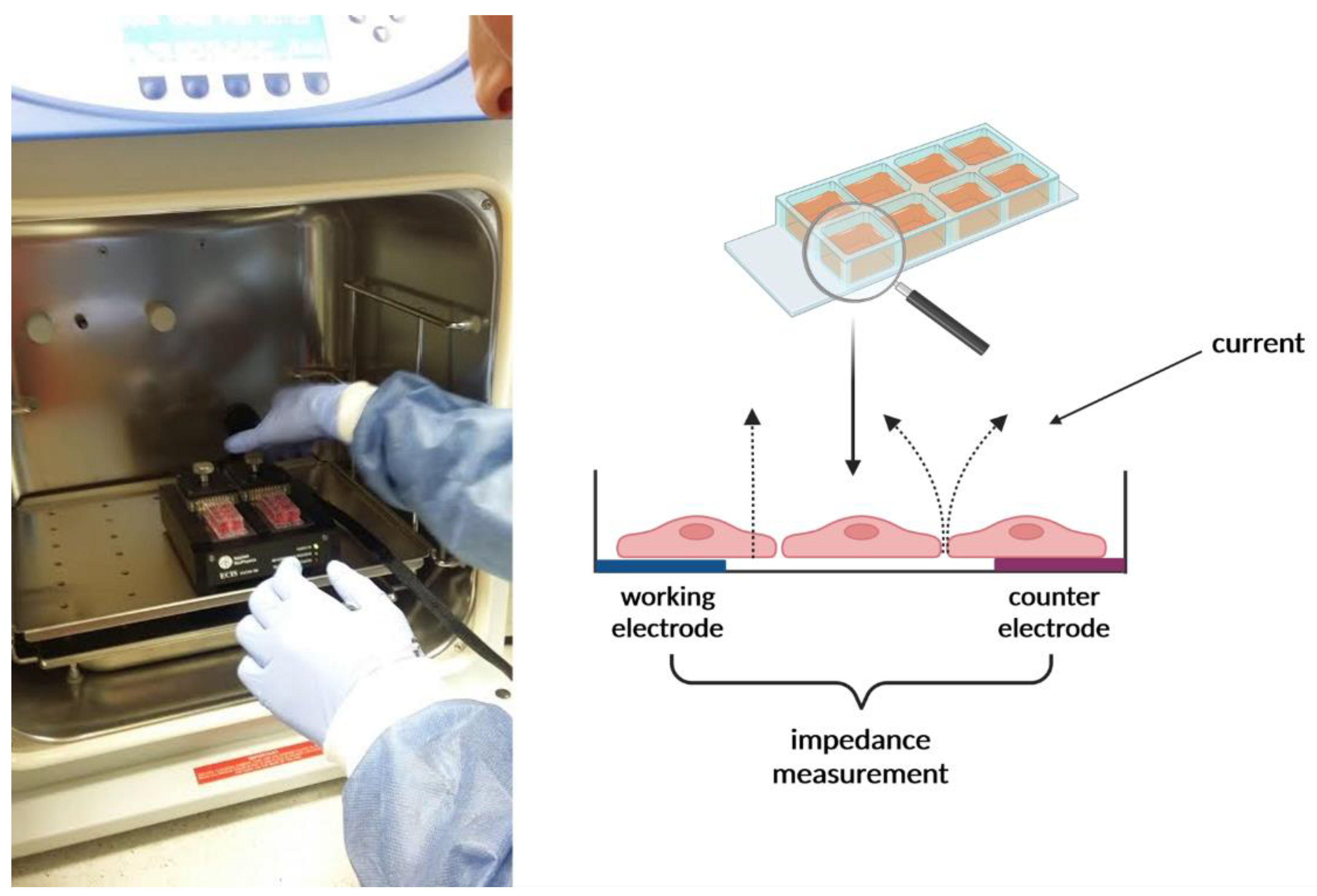
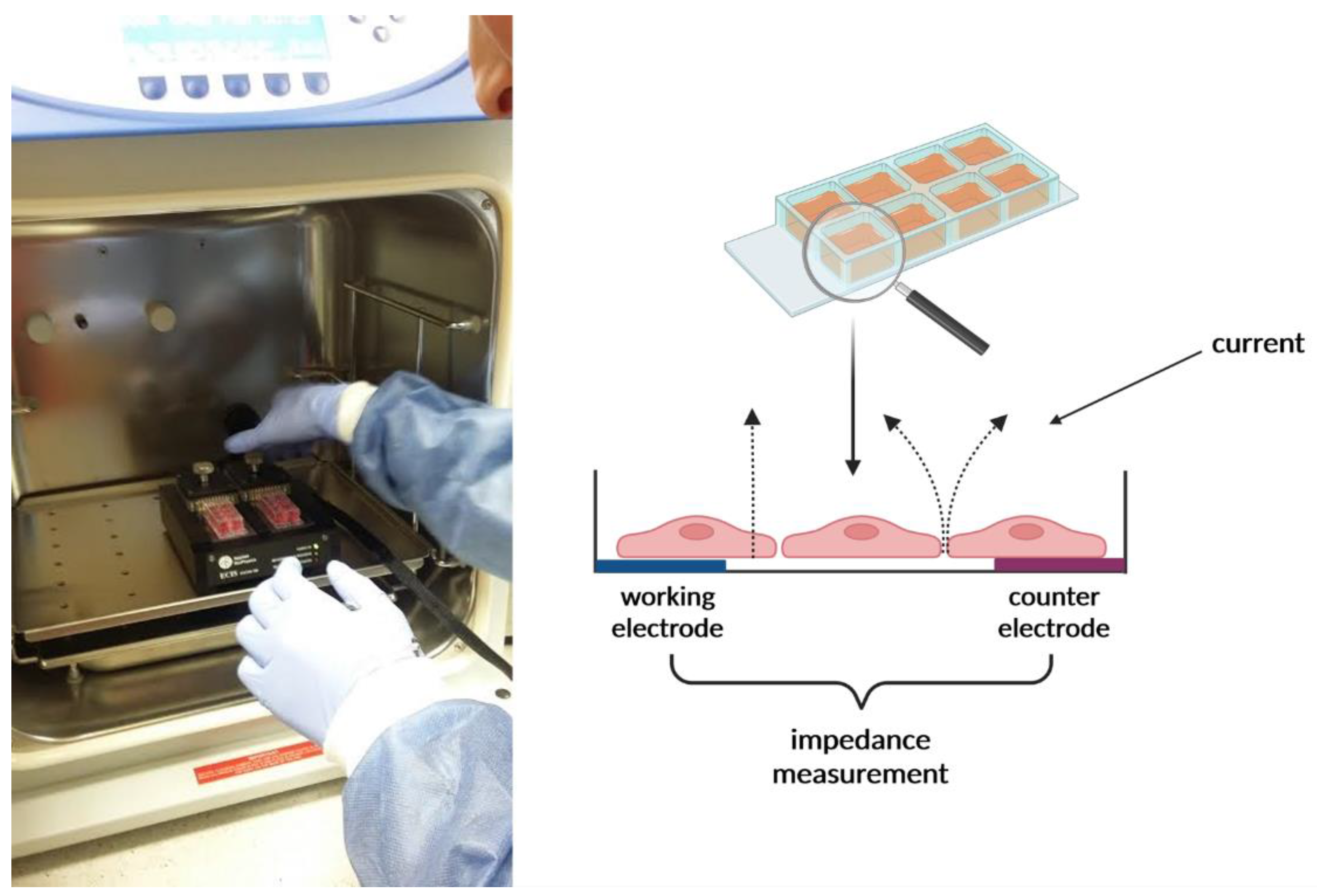
4.5. Impedance Sensing Assay—ECIS Assay
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Giaever, I.; Keese, C.R. A morphological biosensor for mammalian cells. Nature 1993, 366, 591–592. [Google Scholar] [CrossRef] [PubMed]
- Giaever, I.; Keese, C.R. Monitoring fibroblast behavior in tissue culture with an applied electric field. Proc. Natl. Acad. Sci. USA 1984, 81, 3761–3764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pänke, O.; Balkenhohl, T.; Kafka, J.; Schäfer, D.; Lisdat, F. Impedance spectroscopy and biosensing. Adv. Biochem. Eng. Biotechnol. 2007, 109, 195–237. [Google Scholar]
- Serrano, J.A.; Huertas, G.; Maldonado-Jacobi, A.; Olmo, A.; Pérez, P.; Martín, M.E.; Daza, P.; Yúfera, A. An Empirical-Mathematical Approach for Calibration and Fitting Cell-Electrode Electrical Models in Bioimpedance Tests. Sensors 2018, 18, 2354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, C.M.; Keese, C.R.; Giaever, I. Impedance analysis of MDCK cells measured by electric cell-substrate impedance sensing. Biophys. J. 1995, 69, 2800–2807. [Google Scholar] [CrossRef] [Green Version]
- Opp, D.; Wafula, B.; Lim, J.; Huang, E.; Lo, J.-C.; Lo, C.-M. Use of electric cell-substrate impedance sensing to assess in vitro cytotoxicity. Biosens. Bioelectron. 2009, 24, 2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robilliard, L.D.; Kho, D.T.; Johnson, R.H.; Anchan, A.; O’Carroll, S.J.; Graham, E.S. The importance of multifrequency impedance sensing of endothelial barrier formation using ECIS technology for the generation of a strong and durable paracellular barrier. Biosensors 2018, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Szulcek, R.; Bogaard, H.J.; van Nieuw Amerongen, G.P. Electric cell-substrate impedance sensing for the quantification of endothelial proliferation, barrier function, and motility. J. Vis. Exp. 2014, 28, e51300. [Google Scholar] [CrossRef] [Green Version]
- Bennet, D.; Kim, S. Impedance-based cell culture platform to assess light-induced stress changes with antagonist drugs using retinal cells. Anal. Chem. 2013, 85, 4902–4911. [Google Scholar] [CrossRef]
- Dehelean, C.A.; Feflea, S.; Molnár, J.; Zupko, I.; Soica, C. Betulin as an Antitumor Agent Tested in vitro on A431, HeLa and MCF7, and as an Angiogenic Inhibitor in vivo in the CAM Assay. Nat. Prod. Commun. 2012, 7, 1934578X1200700. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Xu, Y.; Wang, L.; Mitchelson, K.; Xing, W.; Cheng, J. Use of cellular electrical impedance sensing to assess in vitro cytotoxicity of anticancer drugs in a human kidney cell nephrotoxicity model. Analyst 2012, 137, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xie, X.; Duan, Y.; Wang, L.; Cheng, Z.; Cheng, J. A review of impedance measurements of whole cells. Biosens. Bioelectron. 2016, 77, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Ostapiuk, A.; Kurach, Ł.; Strzemski, M.; Kurzepa, J.; Hordyjewska, A. Evaluation of Antioxidative Mechanisms In Vitro and Triterpenes Composition of Extracts from Silver Birch (Betula pendula Roth) and Black Birch (Betula obscura Kotula) Barks by FT-IR and HPLC-PDA. Molecules 2021, 26, 4633. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. Betulinic Acid for Cancer Treatment and Prevention. Int. J. Mol. Sci. 2008, 9, 1096. [Google Scholar] [CrossRef] [Green Version]
- Kvasnica, M.; Sarek, J.; Klinotova, E.; Dzubak, P.; Hajduch, M. Synthesis of phthalates of betulinic acid and betulin with cytotoxic activity. Bioorganic Med. Chem. 2005, 13, 3447–3454. [Google Scholar] [CrossRef]
- Oh, S.H.; Choi, J.E.; Lim, S.C. Protection of betulin against cadmium-induced apoptosis in hepatoma cells. Toxicology 2006, 220, 1–12. [Google Scholar] [CrossRef]
- Rzeski, W.; Stepulak, A.; Szymański, M.; Juszczak, M.; Grabarska, A.; Sifringer, M.; Kaczor, J.; Kandefer-Szerszeń, M. Betulin elicits anti-cancer effects in tumour primary cultures and cell lines in vitro. Basic Clin. Pharmacol. Toxicol. 2009, 105, 425–432. [Google Scholar] [CrossRef]
- Zuco, V.; Supino, R.; Righetti, S.C.; Cleris, L.; Marchesi, E.; Gambacorti-Passerini, C.; Formelli, F. Selective cytotoxicity of betulinic acid on tumor cell lines, but not on normal cells. Cancer Lett. 2002, 175, 17–25. [Google Scholar] [CrossRef]
- Fulda, S.; Kroemer, G. Targeting mitochondrial apoptosis by betulinic acid in human cancers. Drug Discov. Today 2009, 14, 885–890. [Google Scholar] [CrossRef]
- Hordyjewska, A.; Ostapiuk, A.; Horecka, A. Betulin and betulinic acid in cancer research. J. Pre-Clin. Clin. Res. 2018, 12, 72–75. [Google Scholar] [CrossRef]
- Bhattarai, B.; Schmidt, M.F.; Ghosh, M.; Sinha Ray, A.; Manhas, S.; Oke, V.; Agu, C.C.; Basunia, M.R.; Enriquez, D.; Quist, J.; et al. Lung Cancer with Skin and Breast Metastasis: A Case Report and Literature Review. Case Rep. Pulmonol. 2015, 2015, 136970. [Google Scholar] [CrossRef] [PubMed]
- McCoy, M.H.; Wang, E. Use of electric cell-substrate impedance sensing as a tool for quantifying cytopathic effect in influenza A virus infected MDCK cells in real-time. J. Virol. Methods 2005, 130, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.R.; Van de Walle, G.R. Electric Cell-Substrate Impedance Sensing To Monitor Viral Growth and Study Cellular Responses to Infection with Alphaherpesviruses in Real Time. mSphere 2017, 2, e00039-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegener, J.; Keese, C.R.; Giaever, I. Electric cell-substrate impedance sensing (ECIS) as a noninvasive means to monitor the kinetics of cell spreading to artificial surfaces. Exp. Cell Res. 2000, 259, 158–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahid, M.; Campbell, C.; Ong, K.; Barnhill, J.; Washington, M. An evaluation of the impact of clinical bacterial isolates on epithelial cell monolayer integrity by the electric Cell-Substrate Impedance Sensing (ECIS) method. J. Microbiol. Methods 2020, 169, 105833. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Jin, T.; Zhang, F.; He, P. Electric cell-substrate impedance sensing (ECIS) for profiling cytotoxicity of cigarette smoke. J. Electroanal. Chem. 2019, 834, 180–186. [Google Scholar] [CrossRef]
- Kutkowska, J.; Strzadala, L.; Rapak, A. Synergistic activity of sorafenib and betulinic acid against clonogenic activity of non-small cell lung cancer cells. Cancer Sci. 2017, 108, 2265–2272. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, R.; Puthli, A.; Balakrishnan, S.; Sapra, B.K.; Mishra, K.P. Betulinic Acid-Induced Cytotoxicity in Human Breast Tumor Cell Lines MCF-7 and T47D and its Modification by Tocopherol. Case Rep. Pulmonol. 2014, 32, 402–408. [Google Scholar] [CrossRef]
- Amiri, S.; Dastghaib, S.; Ahmadi, M.; Mehrbod, P.; Khadem, F.; Behrouj, H.; Aghanoori, M.R.; Machaj, F.; Ghamsari, M.; Rosik, J.; et al. Betulin and its derivatives as novel compounds with different pharmacological effects. Biotechnol. Adv. 2020, 38, 107409. [Google Scholar] [CrossRef]
- Fulda, S.; Scaffidi, G.; Susin, S.A.; Krammer, P.H.; Kroemer, G.; Peter, M.E.; Debatin, K.M. Activation of mitochondria and release of mitochondrial apoptogenic factors by betulinic acid. J. Biol. Chem. 1998, 273, 33942–33948. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, S.; Garda, H.A.; Heinzen, H.; Moyna, P. Effect of plant monofunctional pentacyclic triterpenes on the dynamic and structural properties of dipalmitoylphosphatidylcholine bilayers. Chem. Phys. Lipids 1997, 89, 119–130. [Google Scholar] [CrossRef]
- Schroeder, F.; Atshaves, B.P.; McIntosh, A.L.; Gallegos, A.M.; Storey, S.M.; Parr, R.D.; Jefferson, J.R.; Ball, J.M.; Kier, A.B. Sterol carrier protein-2: New roles in regulating lipid rafts and signaling. Biochim. Biophys. Acta 2007, 1771, 700–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flasiński, M.; Ha¸c-Wydro, K.; Broniatowski, M. Incorporation of Pentacyclic Triterpenes into Mitochondrial Membrane—Studies on the Interactions in Model 2D Lipid Systems. J. Phys. Chem. B 2014, 118, 12927–12937. [Google Scholar] [CrossRef] [PubMed]
- Broniatowski, M.; Flasiński, M.; Ziȩba, K.; Miśkowiec, P. Interactions of pentacyclic triterpene acids with cardiolipins and related phosphatidylglycerols in model systems. Biochim. Biophys. Acta Biomembr. 2014, 1838, 2530–2538. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Kioka, N.; Kato, H.; Matsuo, M.; Ueda, K. Modulation of drug-stimulated ATPase activity of human MDR1/P-glycoprotein by cholesterol. Biochem. J. 2007, 401, 597–605. [Google Scholar] [CrossRef]
- Ohvo-Rekilä, H.; Ramstedt, B.; Leppimäki, P.; Peter Slotte, J. Cholesterol interactions with phospholipids in membranes. Prog. Lipid Res. 2002, 41, 66–97. [Google Scholar] [CrossRef]
- Prinz, W.A. Non-vesicular sterol transport in cells. Prog. Lipid Res. 2007, 46, 297–314. [Google Scholar] [CrossRef] [Green Version]
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Mikheeva, I.B.; Yashin, V.A.; Penkov, N.V.; Vydrina, V.A.; Ishmuratov, G.Y.; Sharapov, V.A.; Khoroshavina, E.I.; et al. Effect of betulin and betulonic acid on isolated rat liver mitochondria and liposomes. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183383. [Google Scholar] [CrossRef]
- Carvalho, F.S.; Morais, C.M.; Holy, J.; Krasutsky, D.; Yemets, S.V.; Krasutsky, P.A.; Jurado, A.S.; Oliveira, P.J.; Serafim, T.L. Toxicity of lupane derivatives on anionic membrane models, isolated rat mitochondria and selected human cell lines: Role of terminal alkyl chains. Chem. Biol. Interact. 2018, 296, 198–210. [Google Scholar] [CrossRef]
- Natarajan, M.; Mohan, S.; Martinez, B.; Meltz, M.; Herman, T. Antioxidant compounds interfere with the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide cytotoxicity assay. Cancer Detect. Prev. 2000, 24, 405–414. [Google Scholar]
- Karakaş, D.; Ari, F.; Ulukaya, E. The MTT viability assay yields strikingly false-positive viabilities although the cells are killed by some plant extracts. Turk. J. Biol. 2017, 41, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Bruggisser, R.; von Daeniken, K.; Jundt, G.; Schaffner, W.; Tullberg-Reinert, H. Interference of plant extracts, phytoestrogens and antioxidants with the MTT tetrazolium assay. Planta Med. 2002, 68, 445–448. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (µM) | |||
|---|---|---|---|
| Compound/Cell Line | T47D | A549 | Vero |
| BE | 7.8 | 9.5 | 21.3 |
| BA | 5.4 | 6.9 | 18.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hordyjewska, A.; Prendecka-Wróbel, M.; Kurach, Ł.; Horecka, A.; Olszewska, A.; Pigoń-Zając, D.; Małecka-Massalska, T.; Kurzepa, J. Antiproliferative Properties of Triterpenoids by ECIS Method—A New Promising Approach in Anticancer Studies? Molecules 2022, 27, 3150. https://doi.org/10.3390/molecules27103150
Hordyjewska A, Prendecka-Wróbel M, Kurach Ł, Horecka A, Olszewska A, Pigoń-Zając D, Małecka-Massalska T, Kurzepa J. Antiproliferative Properties of Triterpenoids by ECIS Method—A New Promising Approach in Anticancer Studies? Molecules. 2022; 27(10):3150. https://doi.org/10.3390/molecules27103150
Chicago/Turabian StyleHordyjewska, Anna, Monika Prendecka-Wróbel, Łukasz Kurach, Anna Horecka, Anna Olszewska, Dominika Pigoń-Zając, Teresa Małecka-Massalska, and Jacek Kurzepa. 2022. "Antiproliferative Properties of Triterpenoids by ECIS Method—A New Promising Approach in Anticancer Studies?" Molecules 27, no. 10: 3150. https://doi.org/10.3390/molecules27103150
APA StyleHordyjewska, A., Prendecka-Wróbel, M., Kurach, Ł., Horecka, A., Olszewska, A., Pigoń-Zając, D., Małecka-Massalska, T., & Kurzepa, J. (2022). Antiproliferative Properties of Triterpenoids by ECIS Method—A New Promising Approach in Anticancer Studies? Molecules, 27(10), 3150. https://doi.org/10.3390/molecules27103150