
Site-Specific Conversion of Cysteine in a Protein to Dehydroalanine Using 2-Nitro-5-thiocyanatobenzoic Acid


Abstract

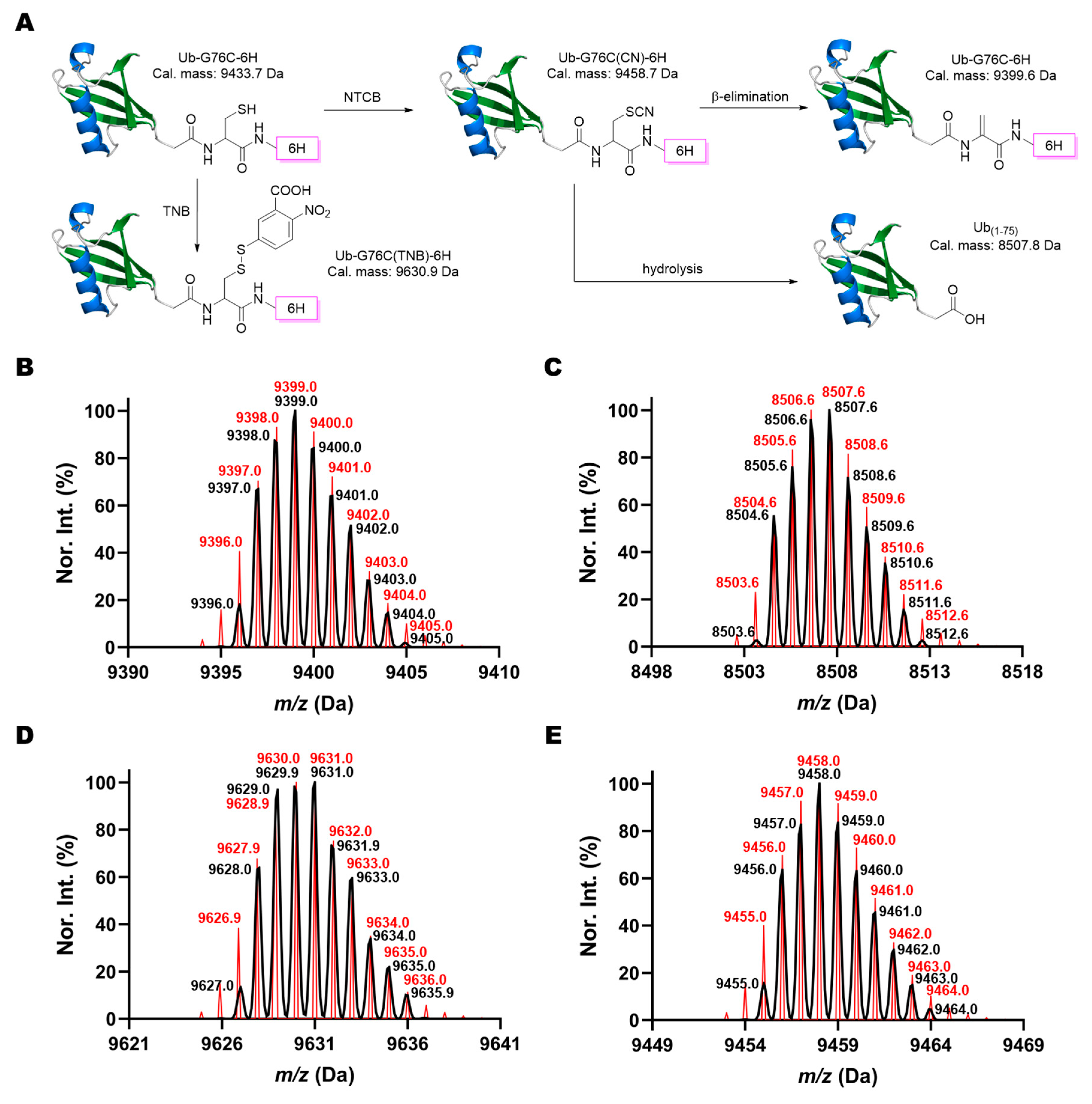
1. Introduction
2. Results
3. Materials and Methods
3.1. General Materials
3.2. Plasmid Construction
3.3. Recombinant Protein Expression and Purification
3.4. NTCB-Triggered Dha Formation
3.5. LC-ESI-MS Analysis
3.6. Mass Spectra Data Processing
3.7. Product Yield Quantitation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Klemm, T.; Ebert, G.; Calleja, D.J.; Allison, C.C.; Richardson, L.W.; Bernardini, J.P.; Lu, B.G.; Kuchel, N.W.; Grohmann, C.; Shibata, Y.; et al. Mechanism and inhibition of the papain-like protease, PLpro, of SARS-CoV-2. EMBO J. 2020, 39, e106275. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.A.; McLellan, L.; Stockley, M.; Gibson, K.R.; Whitlock, G.A.; Knights, C.; Harrigan, J.A.; Jacq, X.; Tate, E.W. Quantitative chemical proteomic profiling of ubiquitin specific proteases in intact cancer cells. ACS Chem. Biol. 2016, 11, 3268–3272. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Wang, Y.; Du, Y.-X.; Liang, L.-J.; Zheng, Q.; Li, Y.-M.; Liu, L. Development and application of ubiquitin-based chemical probes. Chem. Sci. 2020, 11, 12633–12646. [Google Scholar] [CrossRef]
- Chuh, K.N.; Batt, A.R.; Pratt, M.R. Chemical methods for encoding and decoding of posttranslational modifications. Cell Chem. Biol. 2016, 23, 86–107. [Google Scholar] [CrossRef] [PubMed]
- Dawson, P.E.; Muir, T.W.; Clark-Lewis, I.; Kent, S.B. Synthesis of proteins by native chemical ligation. Science 1994, 266, 776–779. [Google Scholar] [CrossRef]
- Muir, T.W.; Sondhi, D.; Cole, P.A. Expressed protein ligation: A general method for protein engineering. Proc. Natl. Acad. Sci. USA 1998, 95, 6705–6710. [Google Scholar] [CrossRef]
- Neumann, H.; Peak-Chew, S.Y.; Chin, J.W. Genetically encoding N(epsilon)-acetyllysine in recombinant proteins. Nat. Chem. Biol. 2008, 4, 232–234. [Google Scholar] [CrossRef]
- Nguyen, D.P.; Mahesh, M.; Elsässer, S.J.; Hancock, S.M.; Uttamapinant, C.; Chin, J.W. Genetic encoding of photocaged cysteine allows photoactivation of TEV protease in live mammalian cells. J. Am. Chem. Soc. 2014, 136, 2240–2243. [Google Scholar] [CrossRef]
- Lee, Y.J.; Wu, B.; Raymond, J.E.; Zeng, Y.; Fang, X.; Wooley, K.L.; Liu, W.R. A genetically encoded acrylamide functionality. ACS Chem. Biol. 2013, 8, 1664–1670. [Google Scholar] [CrossRef]
- Wang, Z.A.; Kurra, Y.; Wang, X.; Zeng, Y.; Lee, Y.J.; Sharma, V.; Lin, H.; Dai, S.Y.; Liu, W.R. A Versatile Approach for Site-Specific Lysine Acylation in Proteins. Angew. Chem. Int. Ed. 2017, 56, 1643–1647. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.A.; Zeng, Y.; Kurra, Y.; Wang, X.; Tharp, J.M.; Vatansever, E.C.; Hsu, W.W.; Dai, S.; Fang, X.; Liu, W.R. A Genetically Encoded Allysine for the Synthesis of Proteins with Site-Specific Lysine Dimethylation. Angew. Chem. Int. Ed. 2017, 56, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.D.; Chu, F.; Racki, L.R.; de la Cruz, C.C.; Burlingame, A.L.; Panning, B.; Narlikar, G.J.; Shokat, K.M. The site-specific installation of methyl-lysine analogs into recombinant histones. Cell 2007, 128, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, G.J.L.; Chalker, J.M.; Errey, J.C.; Davis, B.G. Facile Conversion of Cysteine and Alkyl Cysteines to Dehydroalanine on Protein Surfaces: Versatile and Switchable Access to Functionalized Proteins. J. Am. Chem. Soc. 2008, 130, 5052–5053. [Google Scholar] [CrossRef] [PubMed]
- Chalker, J.M.; Gunnoo, S.B.; Boutureira, O.; Gerstberger, S.C.; Fernández-González, M.; Bernardes, G.J.; Griffin, L.; Hailu, H.; Schofield, C.J.; Davis, B.G. Methods for converting cysteine to dehydroalanine on peptides and proteins. Chem. Sci. 2011, 2, 1666–1676. [Google Scholar] [CrossRef]
- Dadová, J.; Galan, S.R.; Davis, B.G. Synthesis of modified proteins via functionalization of dehydroalanine. Curr. Opin. Chem. Biol. 2018, 46, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.H.; Bower, B.J.; Chalker, J.M.; Bernardes, G.J.; Wiewiora, R.; Ng, W.L.; Raj, R.; Faulkner, S.; Vallee, M.R.; Phanumartwiwath, A.; et al. Posttranslational mutagenesis: A chemical strategy for exploring protein side-chain diversity. Science 2016, 354, 6312. [Google Scholar] [CrossRef]
- Haj-Yahya, N.; Hemantha, H.P.; Meledin, R.; Bondalapati, S.; Seenaiah, M.; Brik, A. Dehydroalanine-based diubiquitin activity probes. Org. Lett. 2014, 16, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Elliott, P.R.; Pinto-Fernandez, A.; Bonham, S.; Kessler, B.M.; Komander, D.; El Oualid, F.; Krappmann, D. A linear diubiquitin-based probe for efficient and selective detection of the deubiquitinating enzyme OTULIN. Cell Chem. Biol. 2017, 24, 1299–1313.e7. [Google Scholar] [CrossRef]
- Xu, L.; Fan, J.; Wang, Y.; Zhang, Z.; Fu, Y.; Li, Y.-M.; Shi, J. An activity-based probe developed by a sequential dehydroalanine formation strategy targets HECT E3 ubiquitin ligases. Chem. Commun. 2019, 55, 7109–7112. [Google Scholar] [CrossRef]
- An, H.; Statsyuk, A.V. Facile synthesis of covalent probes to capture enzymatic intermediates during E1 enzyme catalysis. Chem. Commun. 2016, 52, 2477–2480. [Google Scholar] [CrossRef]
- Meledin, R.; Mali, S.M.; Singh, S.K.; Brik, A. Protein ubiquitination via dehydroalanine: Development and insights into the diastereoselective 1, 4-addition step. Org. Biomol. Chem. 2016, 14, 4817–4823. [Google Scholar] [CrossRef] [PubMed]
- Miseta, A.; Csutora, P. Relationship between the occurrence of cysteine in proteins and the complexity of organisms. Mol. Biol. Evol. 2000, 17, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Krall, N.; Da Cruz, F.P.; Boutureira, O.; Bernardes, G.J. Site-selective protein-modification chemistry for basic biology and drug development. Nat. Chem. 2016, 8, 103–113. [Google Scholar] [CrossRef]
- Dadová, J.; Wu, K.-J.; Isenegger, P.G.; Errey, J.C.; Bernardes, G.A.J.; Chalker, J.M.; Raich, L.; Rovira, C.; Davis, B.G. Precise probing of residue roles by post-translational β, γ-C, N aza-Michael mutagenesis in enzyme active sites. ACS Cent. Sci. 2017, 3, 1168–1173. [Google Scholar] [CrossRef] [PubMed]
- Freedy, A.M.; Matos, M.J.; Boutureira, O.; Corzana, F.; Guerreiro, A.; Akkapeddi, P.; Somovilla, V.J.; Rodrigues, T.; Nicholls, K.; Xie, B. Chemoselective installation of amine bonds on proteins through aza-Michael ligation. J. Am. Chem. Soc. 2017, 139, 18365–18375. [Google Scholar] [CrossRef]
- Wang, J.; Schiller, S.M.; Schultz, P.G. A Biosynthetic Route to Dehydroalanine-Containing Proteins. Angew. Chem. Int. Ed. 2007, 46, 6849–6851. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.U.; Wang, Y.S.; Pai, P.J.; Russell, W.K.; Russell, D.H.; Liu, W.R. A facile method to synthesize histones with posttranslational modification mimics. Biochemistry 2012, 51, 5232–5234. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Ha, S.; Ahn, J.; Kim, R.; Kim, S.; Lee, Y.; Kim, J.; Soll, D.; Lee, H.Y.; Park, H.S. A chemical biology route to site-specific authentic protein modifications. Science 2016, 354, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; He, D.; Long, T.; Zhang, S.; Meng, R.; Chen, P.R. Genetically encoded cleavable protein photo-cross-linker. J. Am. Chem. Soc. 2014, 136, 11860–11863. [Google Scholar] [CrossRef]
- Yang, B.; Wang, N.; Schnier, P.D.; Zheng, F.; Zhu, H.; Polizzi, N.F.; Ittuveetil, A.; Saikam, V.; DeGrado, W.F.; Wang, Q.; et al. Genetically Introducing Biochemically Reactive Amino Acids Dehydroalanine and Dehydrobutyrine in Proteins. J. Am. Chem. Soc. 2019, 141, 7698–7703. [Google Scholar] [CrossRef]
- Qiao, Y.; Yu, G.; Kratch, K.C.; Wang, X.A.; Wang, W.W.; Leeuwon, S.Z.; Xu, S.; Morse, J.S.; Liu, W.R. Expressed Protein Ligation Without Intein. J. Am. Chem. Soc. 2020, 142, 7047–7054. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, G.R.; Schaffer, M.H.; Stark, G.R.; Vanaman, T.C. Specific chemical cleavage in high yield at the amino peptide bonds of cysteine and cystine residues. J. Biol. Chem. 1973, 248, 6583–6591. [Google Scholar] [CrossRef]
- Degani, Y.; Patchornik, A. Cyanylation of sulfhydryl groups by 2-nitro-5-thiocyanobenzoic acid. High-yield modification and cleavage of peptides at cysteine residues. Biochemistry 1974, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.-Y.; Speicher, D.W. Identification of alternative products and optimization of 2-nitro-5-thiocyanatobenzoic acid cyanylation and cleavage at cysteine residues. Anal. Biochem. 2004, 334, 48–61. [Google Scholar] [CrossRef]
- Pickart, C.M.; Eddins, M.J. Ubiquitin: Structures, functions, mechanisms. Biochim. Biophys. Acta Mol. Cell Res. 2004, 1695, 55–72. [Google Scholar] [CrossRef]
- Hu, H.; Sun, S.-C. Ubiquitin signaling in immune responses. Cell Res. 2016, 26, 457–483. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Meng, T.; Chen, L.; Wei, W.; Wang, P. The role of ubiquitination in tumorigenesis and targeted drug discovery. Signal Transduct. Target. Ther. 2020, 5, 1–28. [Google Scholar] [CrossRef]
- Hochstrasser, M. Origin and function of ubiquitin-like proteins. Nature 2009, 458, 422–429. [Google Scholar] [CrossRef]
- Cappadocia, L.; Lima, C.D. Ubiquitin-like protein conjugation: Structures, chemistry, and mechanism. Chem. Rev. 2018, 118, 889–918. [Google Scholar] [CrossRef]
- Mali, S.M.; Singh, S.K.; Eid, E.; Brik, A. Ubiquitin signaling: Chemistry comes to the rescue. J. Am. Chem. Soc. 2017, 139, 4971–4986. [Google Scholar] [CrossRef]
- Love, K.R.; Catic, A.; Schlieker, C.; Ploegh, H.L. Mechanisms, biology and inhibitors of deubiquitinating enzymes. Nat. Chem. Biol. 2007, 3, 697. [Google Scholar] [CrossRef] [PubMed]
- Hewings, D.S.; Flygare, J.A.; Bogyo, M.; Wertz, I.E. Activity-based probes for the ubiquitin conjugation–deconjugation machinery: New chemistries, new tools, and new insights. FEBS J. 2017, 284, 1555–1576. [Google Scholar] [CrossRef]
- Meledin, R.; Mali, S.M.; Kleifeld, O.; Brik, A. Activity-Based Probes Developed by Applying a Sequential Dehydroalanine Formation Strategy to Expressed Proteins Reveal a Potential α-Globin-Modulating Deubiquitinase. Angew. Chem. Int. Ed. 2018, 57, 5645–5649. [Google Scholar] [CrossRef]
- Jiang, H.-K.; Kurkute, P.; Li, C.-L.; Wang, Y.-H.; Chen, P.-J.; Lin, S.-Y.; Wang, Y.-S. Revealing USP7 Deubiquitinase Substrate Specificity by Unbiased Synthesis of Ubiquitin Tagged SUMO2. Biochemistry 2020, 59, 3796–3801. [Google Scholar] [CrossRef] [PubMed]
- Witting, K.F.; Mulder, M.P.; Ovaa, H. Advancing our understanding of ubiquitination using the Ub-toolkit. J. Mol. Biol. 2017, 429, 3388–3394. [Google Scholar] [CrossRef] [PubMed]
- van Wijk, S.J.; Fulda, S.; Dikic, I.; Heilemann, M. Visualizing ubiquitination in mammalian cells. EMBO Rep. 2019, 20, e46520. [Google Scholar] [CrossRef]
- Mulder, M.P.; Witting, K.; Berlin, I.; Pruneda, J.N.; Wu, K.-P.; Chang, J.-G.; Merkx, R.; Bialas, J.; Groettrup, M.; Vertegaal, A.C. A cascading activity-based probe sequentially targets E1–E2–E3 ubiquitin enzymes. Nat. Chem. Biol. 2016, 12, 523. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction Condition | % Yield | ||||
|---|---|---|---|---|---|
| Buffer | pH | Temp. (°C) | Dha | Hydro | TNB |
| 1× PBS | 9 | 37 | 79.9 | 20.1 | 0 |
| 1× PBS | 8 | 37 | 84.4 | 15.6 | 0 |
| 1× PBS | 7 | 37 | 89.3 | 10.7 | 0.9 |
| 1× PBS | 6.5 | 37 | 89.5 | 10.5 | 0 |
| 1× PBS | 7 | 23 | 61.6 | 4.8 | 1.5 |
| 1× PBS | 7 | 5 | 12.2 | 0.6 | 10.2 |
| 20 mM HEPES, 10 mM Py, 6 M GndCl | 7 | 37 | 97.8 | 2.2 | 0 |
| 20 mM HEPES, 10 mM Py | 7 | 37 | 90.2 | 9.8 | 0 |
| 20 mM HEPES, 10 mM Py, 20% DMSO | 7 | 37 | 82.9 | 7.1 | 2.3 |
| 20 mM HEPES, 10 mM Py, 40% DMSO | 7 | 37 | 78.3 | 4.7 | 3.5 |
| 20 mM HEPES, 10 mM Py, 60% DMSO | 7 | 37 | 76.9 | 1.8 | 5.0 |
| 20 mM HEPES, 10 mM Py, 80% DMSO | 7 | 37 | 73.1 | 0.4 | 3.5 |
| 20 mM HEPES, 10 mM Py, 100% DMSO | 7 | 37 | 70.5 | 0 | 1.9 |
| Mass Spectrometer Parameter | Value |
|---|---|
| Spray Voltage (+) | 3.75 kV |
| Capillary Temperature | 320 °C |
| Sheath Gas Flow Rate | 45 |
| Auxiliary Gas Flow Rate | 30 |
| Sweep Gas Flow Rate | 0 |
| S-Lens RF level | 70 |
| Auxiliary Gas Heat Temperature | 30 °C |
| Ion Source | HESI |
| Resolution | 70,000× |
| AGC target | 3 × 106 |
| Maximum Injection Time | 150 ms |
| Protein | Method Duration (min) | Mass Spec Scan Range (m/z) | Gradient (%B) |
|---|---|---|---|
| Ub mutants | 55 | 700–1600 | 5.1–40 min, 15–35%; 40.1 min, 100% |
| Ub mutants | 40 | 700–1600 | 3–20 min, 20–30%; 25.1 min, 100% |
| Ub mutants | 30 | 700–1600 | 3.1–20 min, 25–32%; 20.1 min, 100% |
| SUMO1–4 | 30 | 700–1600 | 3.1–20 min, 25–32%; 20.1 min, 100% |
| NEDD8 | 30 | 700–1600 | 3.1–20 min, 25–35%; 20.1 min, 100% |
| MNSFβ | 30 | 700–1600 | 3.1–20 min, 30–40%; 20.1 min, 100% |
| UFM1 | 55 | 700–1600 | 5.1–40 min, 25–45%; 40.1 min, 100% |
| URM1 | 55 | 700–1800 | 7.1–32 min, 30–45%; 37.1 min, 60%; 40 min, 98% |
| ISG15 | 30 | 700–2000 | 3.1–20 min, 32–42%; 20.1 min, 100% |
| GABARAP | 30 | 700–1800 | 3.1–20 min, 27–37%; 20.1 min, 100% |
| GABARAPL2 | 30 | 700–1800 | 3.1–20 min, 27–37%; 20.1 min, 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, Y.; Yu, G.; Leeuwon, S.Z.; Liu, W.R. Site-Specific Conversion of Cysteine in a Protein to Dehydroalanine Using 2-Nitro-5-thiocyanatobenzoic Acid. Molecules 2021, 26, 2619. https://doi.org/10.3390/molecules26092619
Qiao Y, Yu G, Leeuwon SZ, Liu WR. Site-Specific Conversion of Cysteine in a Protein to Dehydroalanine Using 2-Nitro-5-thiocyanatobenzoic Acid. Molecules. 2021; 26(9):2619. https://doi.org/10.3390/molecules26092619
Chicago/Turabian StyleQiao, Yuchen, Ge Yu, Sunshine Z. Leeuwon, and Wenshe Ray Liu. 2021. "Site-Specific Conversion of Cysteine in a Protein to Dehydroalanine Using 2-Nitro-5-thiocyanatobenzoic Acid" Molecules 26, no. 9: 2619. https://doi.org/10.3390/molecules26092619
APA StyleQiao, Y., Yu, G., Leeuwon, S. Z., & Liu, W. R. (2021). Site-Specific Conversion of Cysteine in a Protein to Dehydroalanine Using 2-Nitro-5-thiocyanatobenzoic Acid. Molecules, 26(9), 2619. https://doi.org/10.3390/molecules26092619