
Natural Product Inhibition and Enzyme Kinetics Related to Phylogenetic Characterization for Bacterial Peptidyl-tRNA Hydrolase 1

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
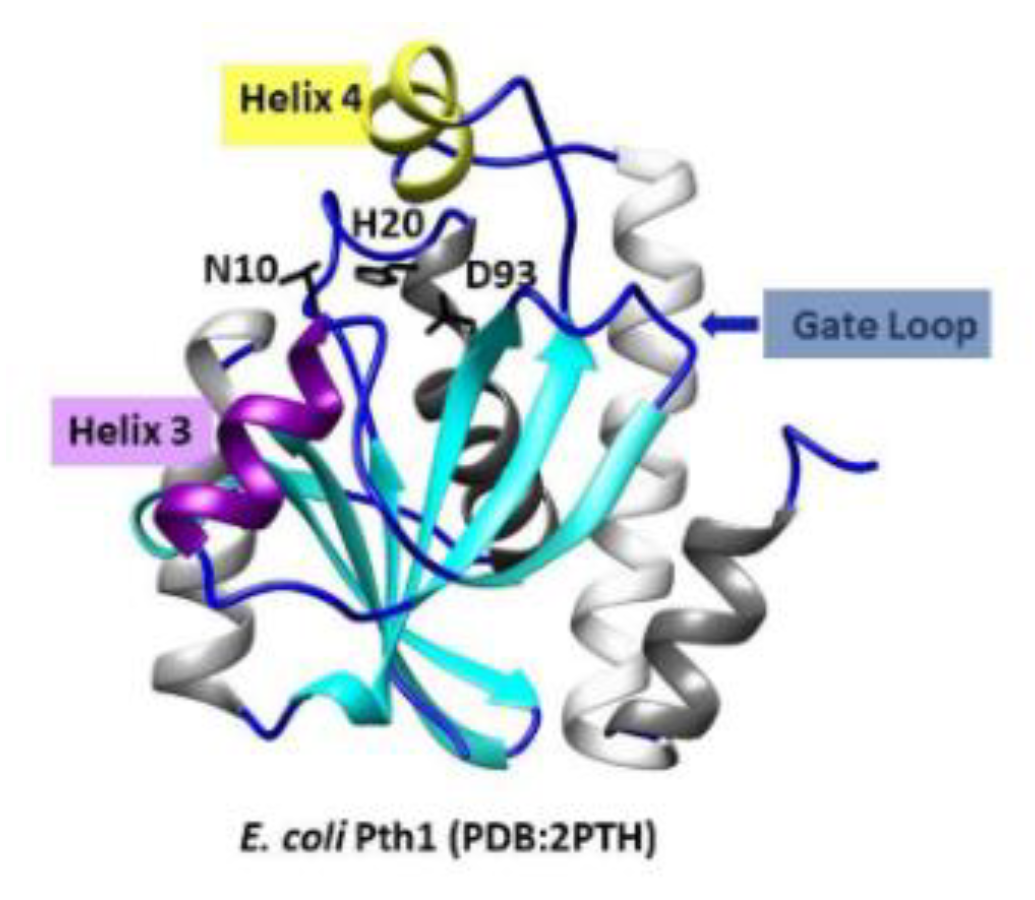
1. Introduction
2. Results
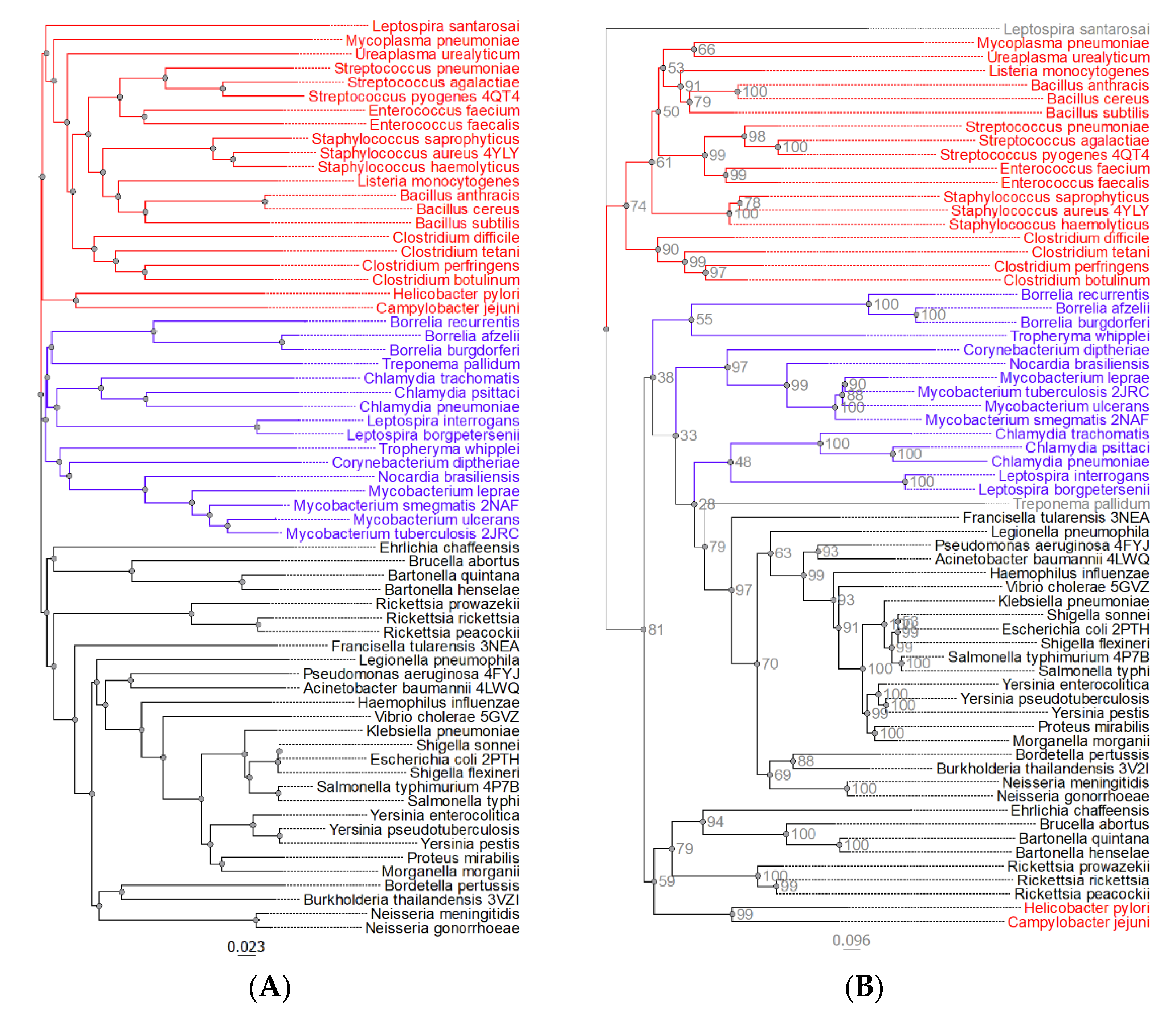
2.1. Phylogenetic Tree
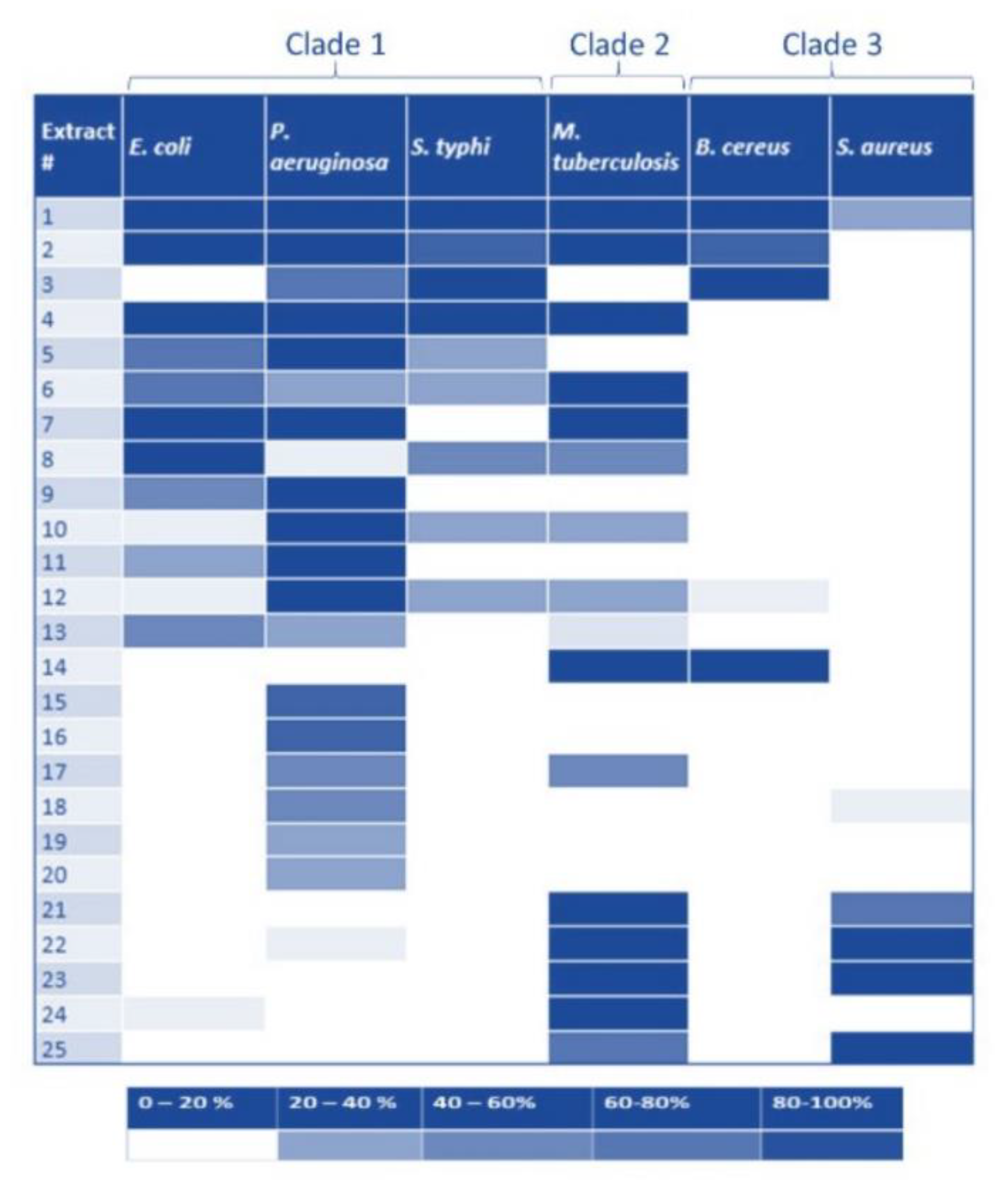
2.2. Pth1 Natural Product Inhibition
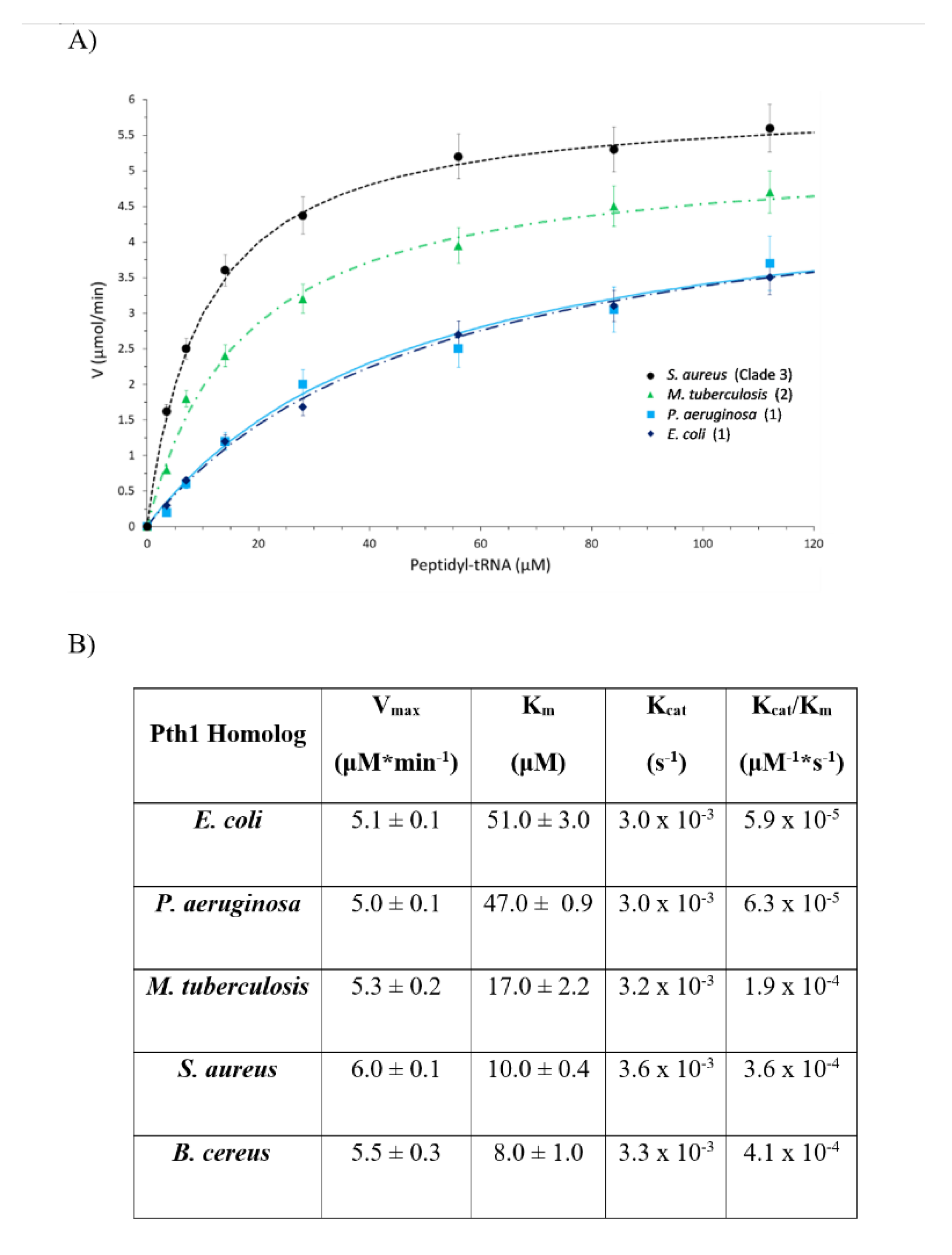
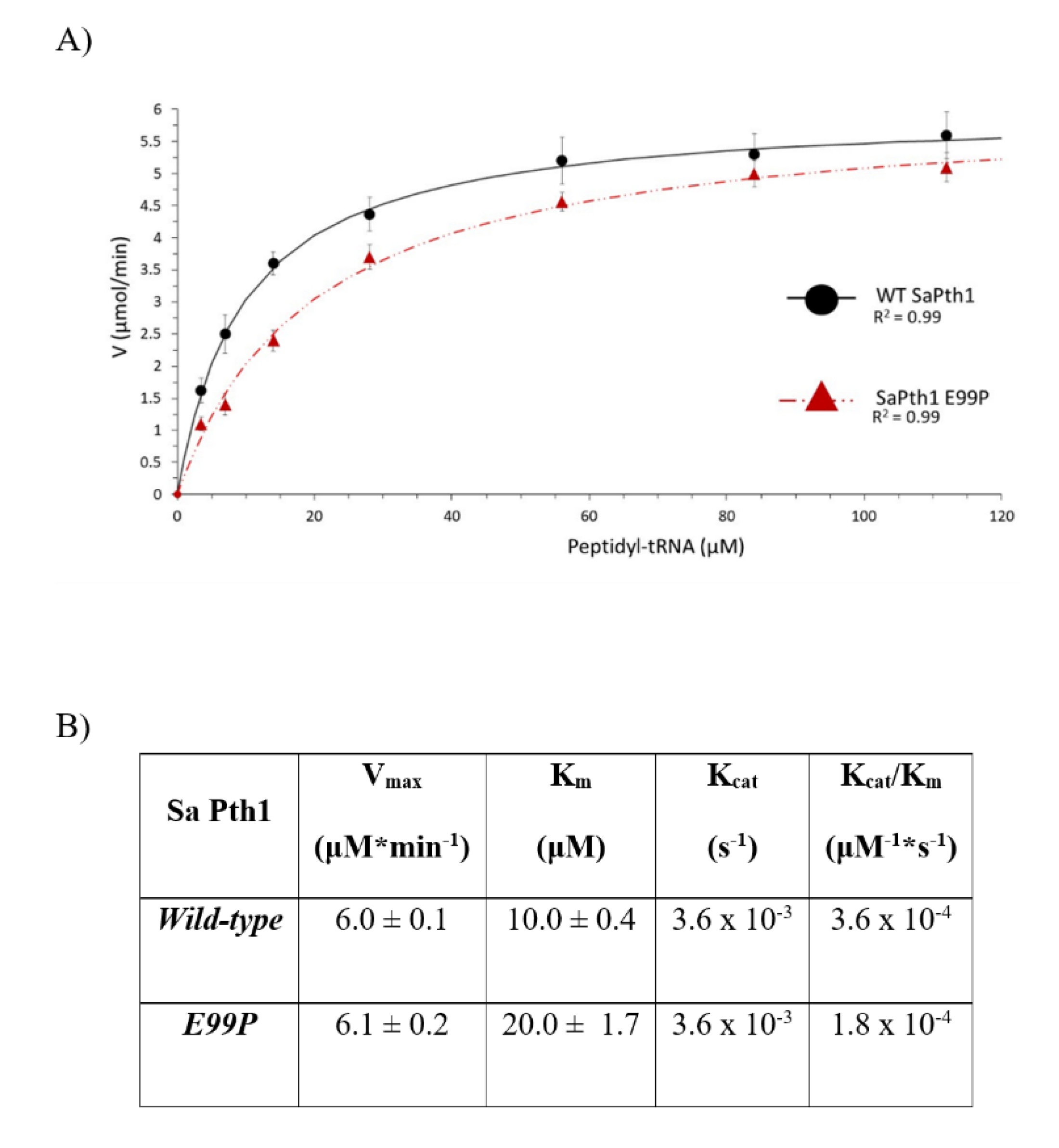
2.3. Enzyme Kinetic Parametrization
3. Discussion
4. Materials and Methods
4.1. Phylogenetic Characterization
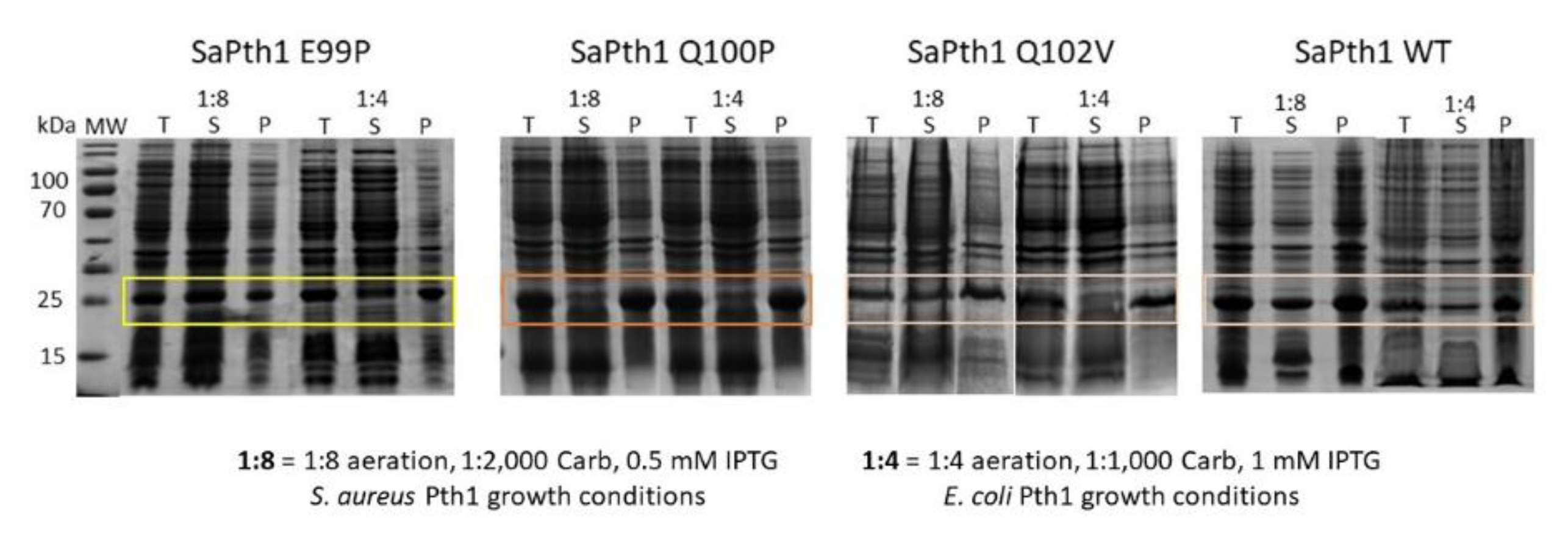
4.2. Pth1 Expression and Purification
4.3. Site-Directed Mutagenesis of the S. aureus Pth1 Gate Loop
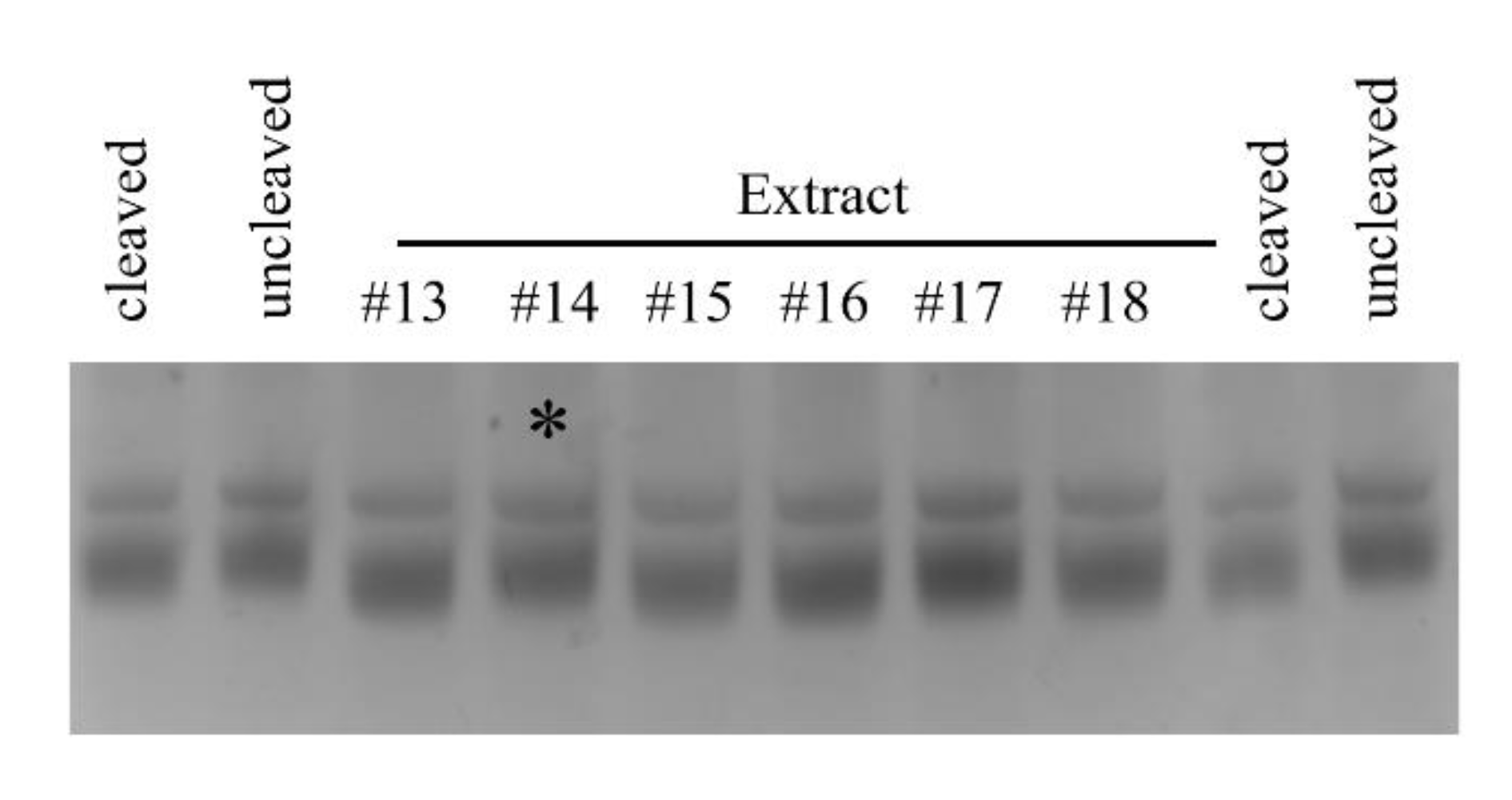
4.4. Pth1 Enzyme Activity
4.5. Natural Product Inhibition
4.6. Pth1 Kinetic Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Jørgensen, F.; Kurland, C.G. Processivity errors of gene expression in Escherichia coli. J. Mol. Biol. 1990, 215, 511–521. [Google Scholar] [CrossRef]
- Kurland, C.G.; Ehrenberg, M. Constraints on the Accuracy of Messenger Rna Movement. Q. Rev. Biophys. 1985, 18, 423–450. [Google Scholar] [CrossRef] [PubMed]
- Manley, J.L. Synthesis and degradation of termination and premature-termination fragments of β-galactosidase in vitro and in vivo. J. Mol. Biol. 1978, 125, 407–432. [Google Scholar] [CrossRef]
- Cruz-Vera, L.R.; Hernández-Ramón, E.; Pérez-Zamorano, B.; Guarneros, G. The rate of peptidyl-tRNA dissociation from the ribosome during minigene expression depends on the nature of the last decoding interaction. J. Biol. Chem. 2003, 278, 26065–26070. [Google Scholar] [CrossRef]
- Hernández-Sánchez, J.; Valadez, J.G.; Herrera, J.V.; Ontiveros, C.; Guarneros, G. λ bar minigene-mediated inhibition of protein synthesis involves accumulation of peptidyl-tRNA and starvation for tRNA. EMBO J. 1998, 17, 3758–3765. [Google Scholar] [CrossRef]
- Tenson, T.; Vega Herrera, J.; Kloss, P.; Guarneros, G.; Mankin, A.S. Inhibition of translation and cell growth by minigene expression. J. Bacteriol. 1999, 181, 1617–1622. [Google Scholar] [CrossRef]
- Heurgué-Hamard, V.; Mora, L.; Guarneros, G.; Buckingham, R.H. The growth defect in Escherichia coli deficient in peptidyl-tRNA hydrolase is due to starvation for Lys-tRNA(Lys). EMBO J. 1996, 15, 2826–2833. [Google Scholar] [CrossRef]
- Menninger, J.R. Accumulation of peptidyl tRNA is lethal to Escherichia coli. J. Bacteriol. 1979, 137, 694–696. [Google Scholar] [CrossRef]
- Menez, J.; Buckingham, R.H.; De Zamaroczy, M.; Campelli, C.K. Peptidyl-tRNA hydrolase in Bacillus subtilis, encoded by spoVC, is essential to vegetative growth, whereas the homologous enzyme in Saccharomyces cerevisiae is dispensable. Mol. Microbiol. 2002, 45, 123–129. [Google Scholar] [CrossRef]
- Rosas-Sandoval, G.; Ambrogelly, A.; Rinehart, J.; Wei, D.; Cruz-Vera, L.R.; Graham, D.E.; Stetter, K.O.; Guarneros, G.; Söll, D. Orthologs of a novel archaeal and of the bacterial peptidyl-tRNA hydrolase are nonessential in yeast. Proc. Natl. Acad. Sci. USA 2002, 99, 16707–16712. [Google Scholar] [CrossRef]
- Goodall, J.J.; Chen, G.J.; Page, M.G.P. Essential Role of Histidine 20 in the Catalytic Mechanism of Escherichia coli Peptidyl-tRNA Hydrolase. Biochemistry 2004, 43, 4583–4591. [Google Scholar] [CrossRef]
- Pulavarti, S.V.S.R.K.; Jain, A.; Prakash Pathak, P.; Mahmood, A.; Arora, A. Solution Structure and Dynamics of Peptidyl-tRNA Hydrolase from Mycobacterium tuberculosis H37Rv. J. Mol. Biol. 2008, 378, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Kabra, A.; Shahid, S.; Pal, R.K.; Yadav, R.; Rama Krishna Pulavarti, S.V.S.; Jain, A.; Tripathi, S.; Arora, A. Unraveling the stereochemical and dynamic aspects of the catalytic site of bacterial peptidyl-tRNA hydrolase. RNA 2017, 23. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, L.; Plateau, P.; O’Mahony, G.; Aubard, C.; Fromant, M.; Thureau, A.; Grøtli, M.; Blanquet, S.; Bontems, F. NMR-based substrate analog docking to escherichia coli peptidyl-tRNA hydrolase. J. Mol. Biol. 2011, 412, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, L.; Bontems, F.; Fromant, M.; Aubard, C.; Blanquet, S.; Plateau, P. RNA-binding site of escherichia coli peptidyl-tRNA hydrolase. J. Biol. Chem. 2011, 286, 39585–39594. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Murakami, R.; Mochizuki, M.; Qi, H.; Shimizu, Y.; Miura, K.I.; Ueda, T.; Uchiumi, T. Structural basis for the substrate recognition and catalysis of peptidyl-tRNA hydrolase. Nucleic Acids Res. 2012, 40. [Google Scholar] [CrossRef] [PubMed]
- Hames, M.C.; McFeeters, H.; Blake Holloway, W.; Stanley, C.B.; Urban, V.S.; McFeeters, R.L. Small molecule binding, docking, and characterization of the interaction between Pth1 and peptidyl-tRNA. Int. J. Mol. Sci. 2013, 14, 22741–22752. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, P.P.; Holloway, W.B.; Setzer, W.N.; McFeeters, H.; McFeeters, R.L. Small molecule docking supports broad and narrow spectrum potential for the inhibition of the novel antibiotic target bacterial Pth1. Antibiotics 2016, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Shiloach, J.; Bauer, S.; De Groot, N.; Lapidot, Y. The influence of the peptide chain length on the activity of peptidyl-tRNA hydrolase from E. coli. Nucleic Acids Res. 1975, 2. [Google Scholar] [CrossRef]
- McFeeters, R.L. Recent Antimicrobial Developments Targeting Peptidyl-tRNA Hydrolases. JSM Biotechnol. Biomed. Eng. 2013, 1, 1006–1008. [Google Scholar]
- McFeeters, H.; McFeeters, R.L. Current Methods for Analysis of Enzymatic Peptidyl-tRNA Hydrolase. J. Anal. Bioanal. Tech. 2014, 5. [Google Scholar] [CrossRef]
- Holloway, W.B.; McFeeters, H.; Powell, A.; Nidadavolu, G.S.; McFeeters, R.L. A Highly Adaptable Method for Quantification of Peptidyl-tRNA Hydrolase Activity. J. Anal. Bioanal. Tech. 2015, 6. [Google Scholar] [CrossRef]
- Bonin, P.D.; Erickson, L.A. Development of a fluorescence polarization assay for peptidyl-tRNA hydrolase. Anal. Biochem. 2002, 306. [Google Scholar] [CrossRef]
- Harris, S.M.; McFeeters, H.; Ogungbe, I.V.; Cruz-Vera, L.R.; Setzer, W.N.; Jackes, B.R.; McFeeters, R.L. Peptidyl-tRNA hydrolase screening combined with molecular docking reveals the antibiotic potential of Syzygium johnsonii bark extract. Nat. Prod. Commun. 2011, 6. [Google Scholar] [CrossRef]
- McFeeters, H.; Gilbert, M.J.; Thompson, R.M.; Setzer, W.N.; Cruz-Vera, L.R.; McFeeters, R.L. Inhibition of essential bacterial peptidyl-tRNA hydrolase activity by tropical plant extracts. Nat. Prod. Commun. 2012, 7. [Google Scholar] [CrossRef]
- Wu, S.; Yang, L.; Gao, Y.; Liu, X.; Liu, F. Multi-channel counter-current chromatography for high-throughput fractionation of natural products for drug discovery. J. Chromatogr. A 2008, 1180. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Vera, L.R.; Toledo, I.; Hernández-Sánchez, J.; Guarneros, G. Molecular basis for the temperature sensitivity of Escherichia coli pth(Ts). J. Bacteriol. 2000, 182. [Google Scholar] [CrossRef] [PubMed]
- Sethi, H.S.; Osier, J.L.; Burks, G.L.; Lamar, J.F.; McFeeters, H.; McFeeters, R.L. Expedited isolation of natural product peptidyl-tRNA hydrolase inhibitors from a Pth1 affinity column. AIMS Mol. Sci. 2017, 4, 175–184. [Google Scholar] [CrossRef]
- Bal, N.C.; Agrawal, H.; Meher, A.K.; Arora, A. Characterization of peptidyl-tRNA hydrolase encoded by open reading frame Rv1014c of Mycobacterium tuberculosis H37Rv. Biol. Chem. 2007, 388. [Google Scholar] [CrossRef]
- Klagsbrun, M. An evolutionary study of the methylation of transfer and ribosomal ribonucleic acid in prokaryote and eukaryote organisms. J. Biol. Chem. 1973, 248, 2612–2620. [Google Scholar] [CrossRef]
- Vani, B.R.; Ramakrishnan, T.; Taya, Y.; Noguchi, S.; Yamaizumi, Z.; Nishimura, S. Occurrence of 1-methyladenosine and absence of ribothymidine in transfer ribonucleic acid of Mycobacterium smegmatis. J. Bacteriol. 1979, 137. [Google Scholar] [CrossRef] [PubMed]
- Bocchetta, M.; Xiong, L.; Mankin, A.S. 23S rRNA positions essential for tRNA binding in ribosomal functional sites. Proc. Natl. Acad. Sci. USA 1998, 95. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Douthwaite, S. Methylation at nucleotide G745 or G748 in 23S rRNA distinguishes Gram-negative from Gram-positive bacteria. Mol. Microbiol. 2002, 44. [Google Scholar] [CrossRef] [PubMed]
- Boccaletto, P.; MacHnicka, M.A.; Purta, E.; Pitkowski, P.; Baginski, B.; Wirecki, T.K.; De Crécy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018, 46. [Google Scholar] [CrossRef] [PubMed]
- Chojnacki, S.; Cowley, A.; Lee, J.; Foix, A.; Lopez, R. Programmatic access to bioinformatics tools from EMBL-EBI update: 2017. Nucleic Acids Res. 2017, 45. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32. [Google Scholar] [CrossRef]
- Nei, M.; Saitou, N. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; Varlamov, A.; Vaskin, Y.; Efremov, I.; German Grehov, O.G.; Kandrov, D.; Rasputin, K.; Syabro, M.; et al. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Hughes, R.C.; McFeeters, H.; Coates, L.; McFeeters, R.L. Recombinant production, crystallization and X-ray crystallographic structure determination of the peptidyl-tRNA hydrolase of Pseudomonas aeruginosa. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2012, 68. [Google Scholar] [CrossRef]
- Taylor-Creel, K.; Hames, M.C.; Holloway, W.B.; McFeeters, H.; Mcfeeters, R.L. Expression, purification, and solubility optimization of peptidyl-tRNA hydrolase 1 from Bacillus cereus. Protein Expr. Purif. 2014, 95. [Google Scholar] [CrossRef]
- Vandavasi, V.; Taylor-Creel, K.; McFeeters, R.L.; Coates, L.; McFeeters, H. Recombinant production, crystallization and X-ray crystallographic structure determination of peptidyl-tRNA hydrolase from Salmonella typhimurium. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2014, 70. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, M.; Singh, N.S.; Roy, S.; Sangeetha, R.; Varshney, U.; Vijayan, M. Cloning, expression, purification, crystallization and preliminary X-ray analysis of peptidyl-tRNA hydrolase from Mycobacterium tuberculosis. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2006, 62, 913–915. [Google Scholar] [CrossRef] [PubMed]
- Varshney, U.; Lee, C.P.; RajBhandary, U.L. Direct analysis of aminoacylation levels of tRNAs in vivo: Application to studying recognition of Escherichia coli initiator tRNA mutants by glutaminyl-tRNA synthetase. J. Biol. Chem. 1991, 266, 24712–24718. [Google Scholar] [CrossRef]
- Setzer, M.C.; Setzer, W.N.; Jackes, B.R.; Gentry, G.A.; Moriarity, D.M. The medicinal value of tropical rainforest plants from Paluma, North Queensland, Australia. Pharm. Biol. 2001, 39. [Google Scholar] [CrossRef]
- Setzer, M.C.; Schmidt, J.M.; Irvine, A.K.; Jackes, B.R.; Setzer, W.N. Biological activity of rainforest plant extracts from Far North Queensland, Australia. Aust. J. Med. Herbal. 2006, 18, 21–46. [Google Scholar]
- Setzer, M.C.; Moriarity, D.M.; Lawton, R.O.; Setzer, W.N.; Gentry, G.A.; Haber, W.A. Phytomedicinal potential of tropical cloudforest plants from Monteverde, Costa Rica. Rev. Biol. Trop. 2003, 51, 647–673. [Google Scholar]
- Schmidt, J.M.; Noletto, J.A.; Vogler, B.; Setzer, W.N. Abaco bush medicine: Chemical composition of the essential oils of four aromatic medicinal plants from Abaco Island, Bahamas. J. Herbs Spices Med. Plants 2006, 12. [Google Scholar] [CrossRef]
- Wilkinson, G.N. Statistical estimations in enzyme kinetics. Biochem. J. 1961, 80. [Google Scholar] [CrossRef] [PubMed]
- Maxson, T.; Mitchell, D.A. Targeted treatment for bacterial infections: Prospects for pathogen-specific antibiotics coupled with rapid diagnostics. Tetrahedron 2016, 72, 3609–3624. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strange, D.S.; Gaffin, S.S.; Holloway, W.B.; Kinsella, M.D.; Wisotsky, J.N.; McFeeters, H.; McFeeters, R.L. Natural Product Inhibition and Enzyme Kinetics Related to Phylogenetic Characterization for Bacterial Peptidyl-tRNA Hydrolase 1. Molecules 2021, 26, 2281. https://doi.org/10.3390/molecules26082281
Strange DS, Gaffin SS, Holloway WB, Kinsella MD, Wisotsky JN, McFeeters H, McFeeters RL. Natural Product Inhibition and Enzyme Kinetics Related to Phylogenetic Characterization for Bacterial Peptidyl-tRNA Hydrolase 1. Molecules. 2021; 26(8):2281. https://doi.org/10.3390/molecules26082281
Chicago/Turabian StyleStrange, D. Scott, Steven S. Gaffin, W. Blake Holloway, Meredyth D. Kinsella, Jacob N. Wisotsky, Hana McFeeters, and Robert L. McFeeters. 2021. "Natural Product Inhibition and Enzyme Kinetics Related to Phylogenetic Characterization for Bacterial Peptidyl-tRNA Hydrolase 1" Molecules 26, no. 8: 2281. https://doi.org/10.3390/molecules26082281
APA StyleStrange, D. S., Gaffin, S. S., Holloway, W. B., Kinsella, M. D., Wisotsky, J. N., McFeeters, H., & McFeeters, R. L. (2021). Natural Product Inhibition and Enzyme Kinetics Related to Phylogenetic Characterization for Bacterial Peptidyl-tRNA Hydrolase 1. Molecules, 26(8), 2281. https://doi.org/10.3390/molecules26082281