
Biocatalysis in the Chemistry of Lupane Triterpenoids


Abstract
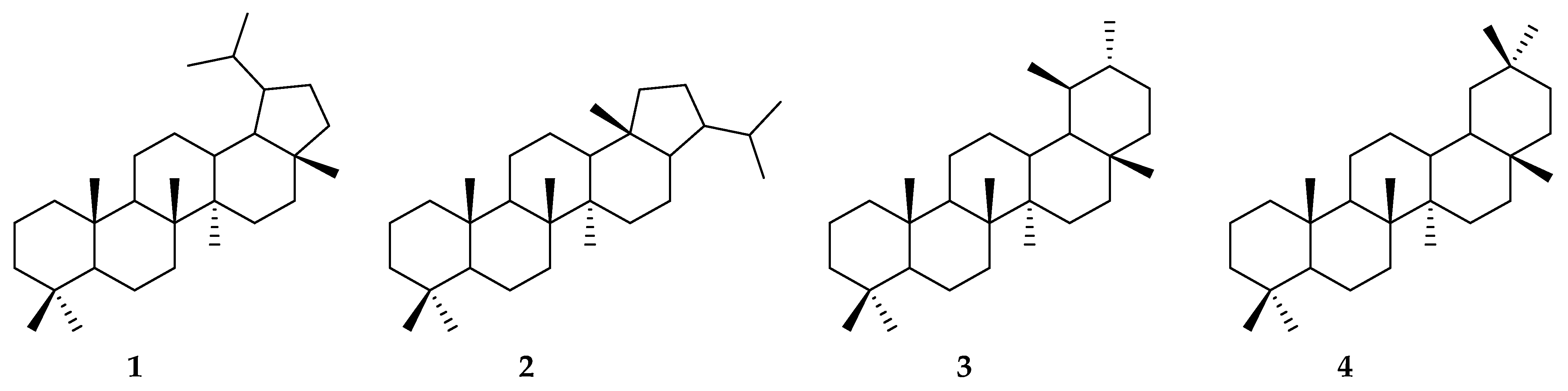
1. Introduction
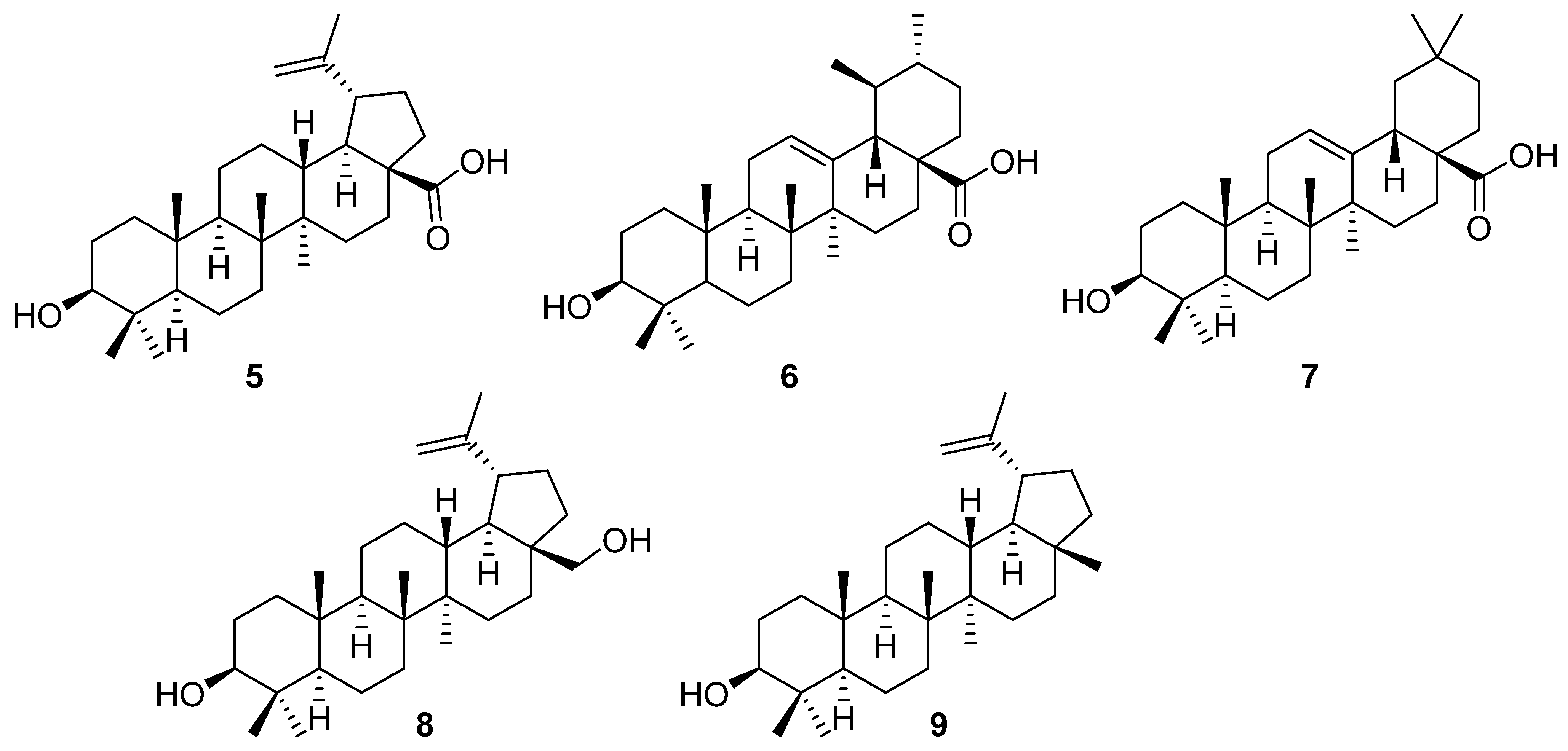
2. Discovery of Betulinic Acid (5) and Its Biological Activity
3. Natural Sources of Betulinic Acid (5), Betulin (8), and Lupeol (9)
4. Semi-Synthetic Procedures for the Preparation of Betulinic Acid (5)
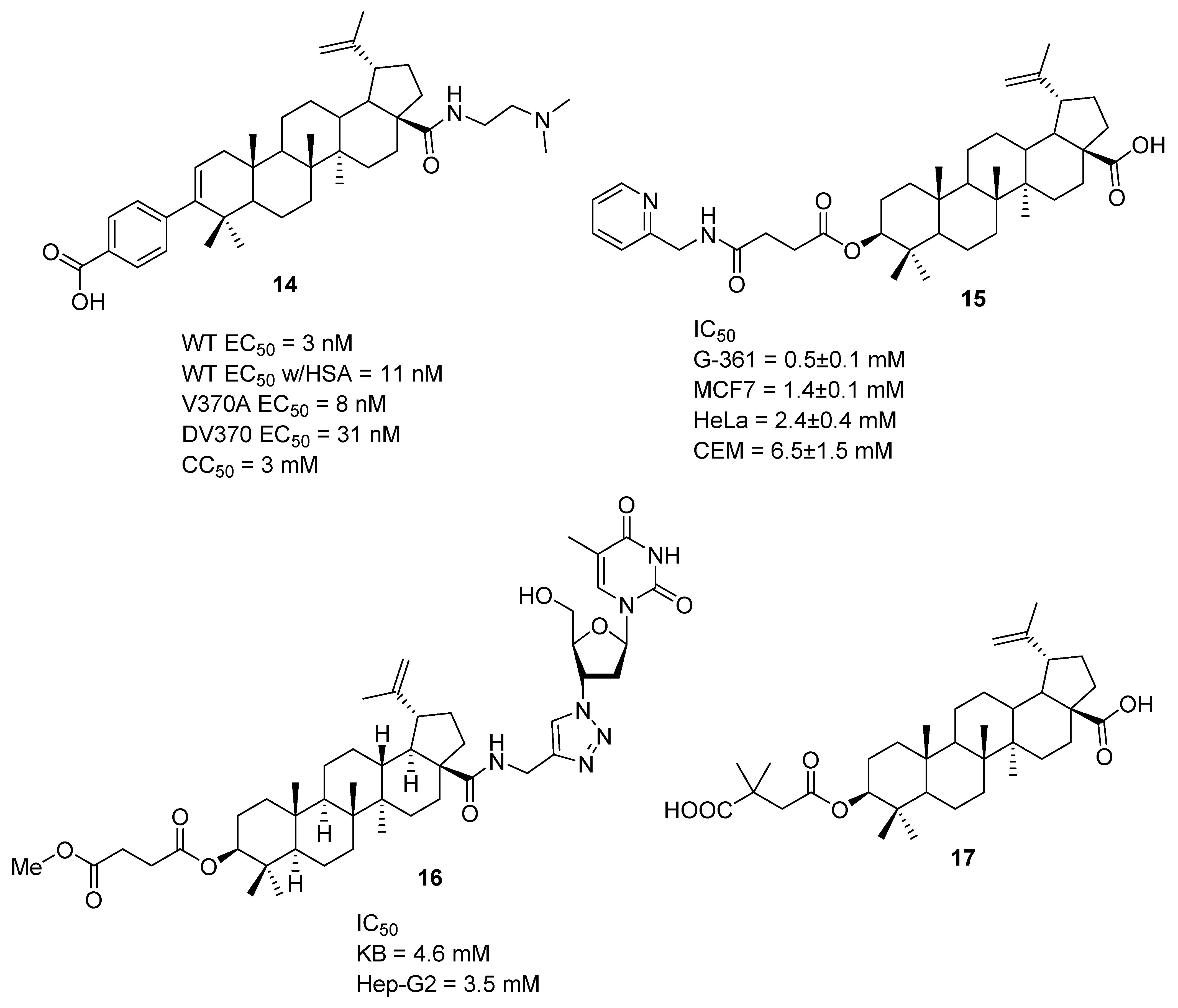
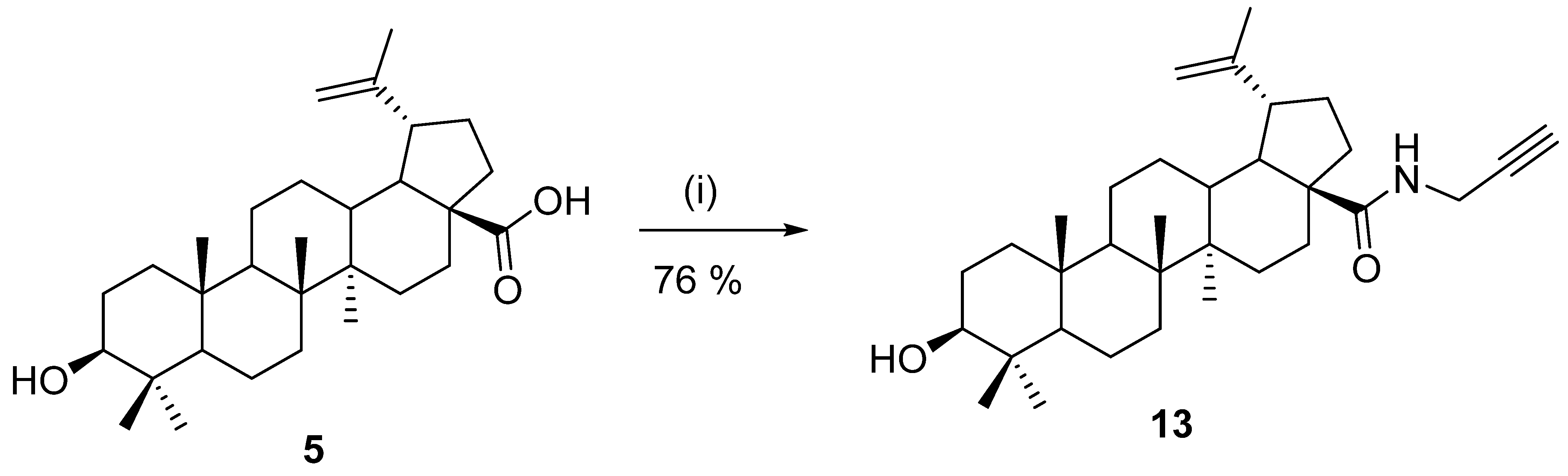
5. Chemical Modification of Betulinic Acid (5)
6. Biocatalysis
7. Biosynthesis of Pentacyclic Triterpenes
8. Biosynthesis of Betulinic Acid (5), Betulin (8), and Lupeol (9)
9. Biocatalyzed Production of Lupane Triterpenoids—Betulinic Acid (5), Betulin (8), and Lupeol (9)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Year | Modification of Biosynthesis | Production | Reference |
|---|---|---|---|---|
| Armillaria luteovirens Sacc | 2011 | optimization of various conditions | production of 5 | [108] |
| Inonotus obliquus | 2012 | optimization of various conditions | 69.37 mg/L of 8 | [109] |
| Inonotus obliquus | 2014 | yield increase with aqueous extract and methanol extract from birch bark | increased production of 8 | [110] |
| Saccharomyces cerevisiae | 2014 | insertion and expression regulation of genes | 5 from 0.01 to 1.92 mg L−1 OD−1 | [112] |
| Saccharomyces cerevisiae | 2015 | changes in intracellular supply of NADPH/oxygen | 1.5 and 3.2 times higher production of 5 | [113] |
| Gal80p mutant of Saccharomyces cerevisiae | 2016 | up-regulation of the expressed genes | 0.16 mg/L/OD600 of 5 | [104] |
| Saccharomyces cerevisiae | 2016 | increase in the supply of squalene | 8.23 mg/L of 9 | [114] |
| Saccharomyces cerevisiae CEN.PK BA4 | 2017 | usage of excess of ethanol | 182 mg/L of 5 | [115] |
| 2017 | [116] | |||
| Phaeodactylum tricornutum | 2019 | introduction of Lotus japonicus oxidosqualene cyclase and Medicago truncatula cytochrome P450 with native reductase | production of 8 and 9 | [117] |
| Escherichia coli and Saccharomyces cerevisiae | 2019 | usage of optimized lupeol pathway genes | production of 9 | [118] |
| Yarrowia lipolytica | 2019 | usage of glycerol as a starting material | 26.53 mg/L of 5 | [120] |
| Yarrowia lipolytica | 2019 | systematic metabolic engineering | 204.89±11.56 mg/L of triterpenoids (23.71% of 5) | [121] |
| Saccharomyces cerevisiae | 2019 | usage of RoCYP01 (CYP716A155) | yields of 5 higher than 1 g/L | [106] |
| various yeast strains | 2020 | SCRaMbLE technique | production of 5 | [122] |
10. Enzymatic Modification of Betulinic Acid (5) and Betulin (8)
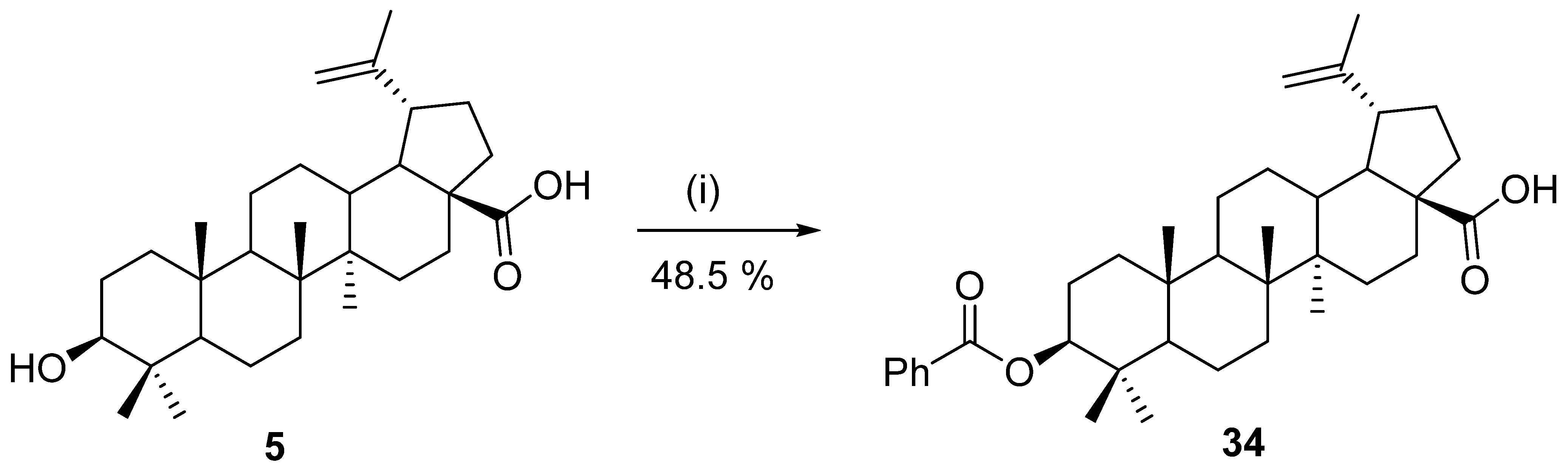
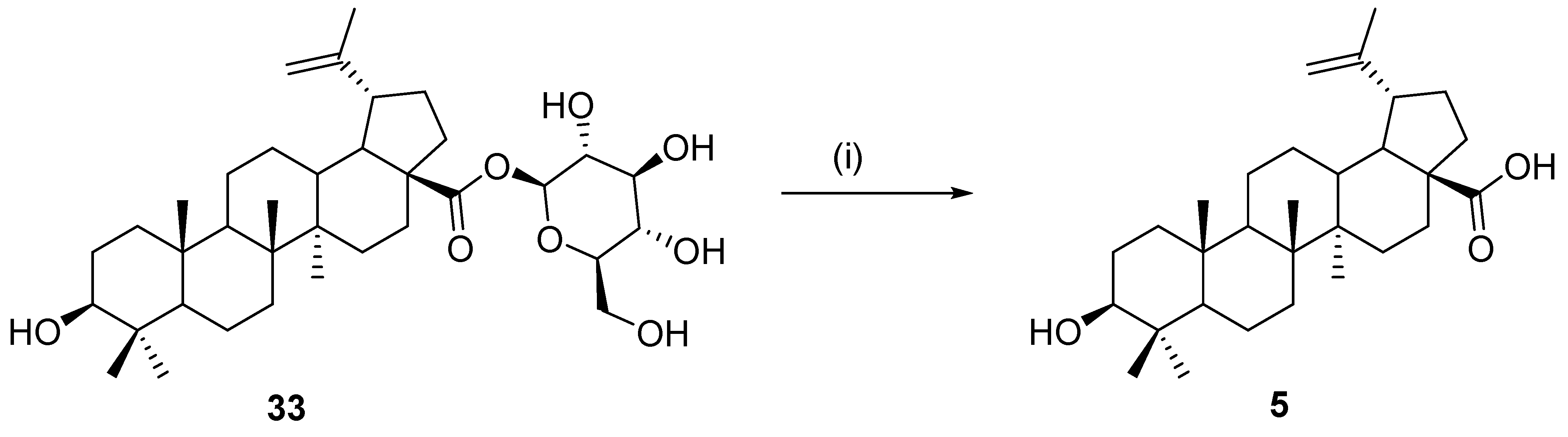
11. Hydrolysis of Sugar Esters of Betulinic Acid
12. Microbial Oxidation of Betulinic Acid (5)
13. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yao, C.-L.; Zhang, J.-Q.; Li, J.-Y.; Wei, W.-L.; Wu, S.-F.; Guo, D.-A. Traditional Chinese Medicine (TCM) as a Source of New Anticancer Drugs. Nat. Prod. Rep. 2021. [Google Scholar] [CrossRef]
- Devi, J.; Kumar, R.; Singh, K.; Gehlot, A.; Bhushan, S.; Kumar, S. In Vitro Adventitious Roots: A Non-Disruptive Technology for the Production of Phytoconstituents on the Industrial Scale. Crit. Rev. Biotechnol. 2021, 1–25. [Google Scholar] [CrossRef]
- Lu, Y.; An, T.; Tian, H.; Gao, X.; Wang, F.; Wang, S.; Ma, K. Depression with Comorbid Diabetes: What Evidence Exists for Treatments Using Traditional Chinese Medicine and Natural Products? Front. Pharm. Ther. 2020, 11, 596362. [Google Scholar] [CrossRef]
- Sun, C.-P.; Jia, Z.-L.; Huo, X.-K.; Tian, X.-G.; Feng, L.; Wang, C.; Zhang, B.-J.; Zhao, W.-Y.; Ma, X.-C. Medicinal Inula Species: Phytochemistry, Biosynthesis, and Bioactivities. Am. J. Chin. Med. 2021, 1–44. [Google Scholar] [CrossRef]
- Zhang, L.; Song, J.; Kong, L.; Yuan, T.; Li, W.; Zhang, W.; Hou, B.; Lu, Y.; Du, G. The Strategies and Techniques of Drug Discovery from Natural Products. Pharm. Ther. 2020, 216, 107686. [Google Scholar] [CrossRef]
- Wohnsland, F.; Faller, B. High-Throughput Permeability PH Profile and High-Throughput Alkane/Water Log P with Artificial Membranes. J. Med. Chem. 2001, 44, 923–930. [Google Scholar] [CrossRef]
- Hiemstra, S.; Ramaiahgari, S.C.; Wink, S.; Callegaro, G.; Coonen, M.; Meerman, J.; Jennen, D.; van den Nieuwendijk, K.; Dankers, A.; Snoeys, J.; et al. High-Throughput Confocal Imaging of Differentiated 3D Liver-like Spheroid Cellular Stress Response Reporters for Identification of Drug-Induced Liver Injury Liability. Arch. Toxicol. 2019, 93, 2895–2911. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Xie, Z.; Kuvelkar, R.; Shah, V.; Bateman, K.; McLaren, D.G.; Cooks, R.G. High-Throughput Bioassays Using “Dip-and-Go” Multiplexed Electrospray Mass Spectrometry. Angew. Chem. Int. Ed. Engl. 2019, 58, 17594–17598. [Google Scholar] [CrossRef]
- Aldewachi, H.; Al-Zidan, R.N.; Conner, M.T.; Salman, M.M. High-Throughput Screening Platforms in the Discovery of Novel Drugs for Neurodegenerative Diseases. Bioengineering (Basel) 2021, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- David, B.; Wolfender, J.-L.; Dias, D.A. The Pharmaceutical Industry and Natural Products: Historical Status and New Trends. Phytochem. Rev. 2015, 14, 299–315. [Google Scholar] [CrossRef]
- Davison, E.K.; Brimble, M.A. Natural Product Derived Privileged Scaffolds in Drug Discovery. Curr. Opin. Chem. Biol. 2019, 52, 1–8. [Google Scholar] [CrossRef]
- Hill, R.A.; Connolly, J.D. Triterpenoids. Nat. Prod. Rep. 2020, 37, 962–998. [Google Scholar] [CrossRef]
- Hill, R.; Connolly, J. Triterpenoids. Nat. Prod. Rep. 2018, 35, 1294–1329. [Google Scholar] [CrossRef]
- Gershenzon, J.; Dudareva, N. The Function of Terpene Natural Products in the Natural World. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Keeling, C.I.; Bohlmann, J. Genes, Enzymes and Chemicals of Terpenoid Diversity in the Constitutive and Induced Defence of Conifers against Insects and Pathogens. New Phytol. 2006, 170, 657–675. [Google Scholar] [CrossRef]
- Tetali, S.D. Terpenes and Isoprenoids: A Wealth of Compounds for Global Use. Planta 2019, 249, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Patočka, J. Biologically Active Pentacyclic Triterpenes and Their Current Medicine Signification. J. Appl. Biomed. 2003, 1, 7–12. [Google Scholar] [CrossRef]
- Luchnikova, N.A.; Grishko, V.V.; Ivshina, I.B. Biotransformation of Oleanane and Ursane Triterpenic Acids. Molecules 2020, 25, 5526. [Google Scholar] [CrossRef] [PubMed]
- Cichewicz, R.H.; Kouzi, S.A. Chemistry, Biological Activity, and Chemotherapeutic Potential of Betulinic Acid for the Prevention and Treatment of Cancer and HIV Infection. Med. Res. Rev. 2004, 24, 90–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-M.; Xu, H.-G.; Wang, L.; Li, Y.-J.; Sun, P.-H.; Wu, X.-M.; Wang, G.-J.; Chen, W.-M.; Ye, W.-C. Betulinic Acid and Its Derivatives as Potential Antitumor Agents. Med. Res. Rev. 2015, 35, 1127–1155. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, J.; Chen, Y. Betulinic Acid and the Pharmacological Effects of Tumor Suppression (Review). Mol. Med. Rep. 2016, 14, 4489–4495. [Google Scholar] [CrossRef]
- Hussain, H.; Green, I.R.; Ali, I.; Khan, I.A.; Ali, Z.; Al-Sadi, A.M.; Ahmed, I. Ursolic Acid Derivatives for Pharmaceutical Use: A Patent Review (2012-2016). Expert. Opin. Ther. Pat. 2017, 27, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Fontanay, S.; Grare, M.; Mayer, J.; Finance, C.; Duval, R.E. Ursolic, Oleanolic and Betulinic Acids: Antibacterial Spectra and Selectivity Indexes. J. Ethnopharmacol. 2008, 120, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Tuli, H.S.; Sharma, A.K. Ursolic Acid (UA): A Metabolite with Promising Therapeutic Potential. Life Sci. 2016, 146, 201–213. [Google Scholar] [CrossRef]
- Lin, C.; Wen, X.; Sun, H. Oleanolic Acid Derivatives for Pharmaceutical Use: A Patent Review. Expert Opin. Ther. Pat. 2016, 26, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Pollier, J.; Goossens, A. Oleanolic Acid. Phytochemistry 2012, 77, 10–15. [Google Scholar] [CrossRef]
- Rodríguez, J.A.; Astudillo, L.; Schmeda-Hirschmann, G. Oleanolic Acid Promotes Healing of Acetic Acid-Induced Chronic Gastric Lesions in Rats. Pharmacol. Res. 2003, 48, 291–294. [Google Scholar] [CrossRef]
- Urban, M.; Kvasnica, M.; Dickinson, N.J.; Sarek, J. Biologically Active Triterpenoids Usable As Prodrugs. In Terpenoids and Squalene: Biosynthesis, Functions and Health Implications; Bates, A.R., Ed.; Nova Science Publishers: New York, NY, USA, 2015; Volume 2015, ISBN 978-1-63463-656-8. [Google Scholar]
- Zhou, M.; Zhang, R.-H.; Wang, M.; Xu, G.-B.; Liao, S.-G. Prodrugs of Triterpenoids and Their Derivatives. Eur. J. Med. Chem. 2017, 131, 222–236. [Google Scholar] [CrossRef]
- Ríos, J.L.; Máñez, S. New Pharmacological Opportunities for Betulinic Acid. Planta Med. 2018, 84, 8–19. [Google Scholar] [CrossRef]
- Żwawiak, J.; Pawełczyk, A.; Olender, D.; Zaprutko, L. Structure and Activity of Pentacyclic Triterpenes Codrugs. A Review. Mini Rev. Med. Chem. 2021. [Google Scholar] [CrossRef]
- Retzlaff, F. Ueber Herba Gratiolae. Arch. Der Pharm. 1902, 240, 561–568. [Google Scholar] [CrossRef]
- Trumbull, E.R.; Bianchi, E.; Eckert, D.J.; Wiedhopf, R.M.; Cole, J.R. Tumor Inhibitory Agents from Vauquelinia Corymbosa (Rosaceae). J. Pharm. Sci. 1976, 65, 1407–1408. [Google Scholar] [CrossRef]
- Fujioka, T.; Kashiwada, Y.; Kilkuskie, R.E.; Cosentino, L.M.; Ballas, L.M.; Jiang, J.B.; Janzen, W.P.; Chen, I.-S.; Lee, K.-H. Anti-AIDS Agents, 11. Betulinic Acid and Platanic Acid as Anti-HIV Principles from Syzigium Claviflorum, and the Anti-HIV Activity of Structurally Related Triterpenoids. J. Nat. Prod. 1994, 57, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Pisha, E.; Chai, H.; Lee, I.S.; Chagwedera, T.E.; Farnsworth, N.R.; Cordell, G.A.; Beecher, C.W.; Fong, H.H.; Kinghorn, A.D.; Brown, D.M. Discovery of Betulinic Acid as a Selective Inhibitor of Human Melanoma That Functions by Induction of Apoptosis. Nat. Med. 1995, 1, 1046–1051. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.L.; Kuzmanoff, K.L.; Ling-Indeck, L.; Pezzuto, J.M. Betulinic Acid Induces Apoptosis in Human Neuroblastoma Cell Lines. Eur. J. Cancer 1997, 33, 2007–2010. [Google Scholar] [CrossRef]
- Freire, C.S.R.; Silvestre, A.J.D.; Neto, C.P.; Cavaleiro, J.S. Lipophilic Extractives of the Inner and Outer Barks of Eucalyptus Globulus. Holzforschung (HF) 2002, 56, 372–379. [Google Scholar] [CrossRef]
- Abe, F.; Yamauchi, T.; Nagao, T.; Kinjo, J.; Okabe, H.; Higo, H.; Akahane, H. Ursolic Acid as a Trypanocidal Constituent in Rosemary. Biol. Pharm. Bull. 2002, 25, 1485–1487. [Google Scholar] [CrossRef]
- Zhao, F.; Mai, Q.; Ma, J.; Xu, M.; Wang, X.; Cui, T.; Qiu, F.; Han, G. Triterpenoids from Inonotus Obliquus and Their Antitumor Activities. Fitoterapia 2015, 101, 34–40. [Google Scholar] [CrossRef]
- Cunha, A.B.; Batista, R.; Castro, M.Á.; David, J.M. Chemical Strategies towards the Synthesis of Betulinic Acid and Its More Potent Antiprotozoal Analogues. Molecules 2021, 26, 1081. [Google Scholar] [CrossRef]
- Liu, J.; Chen, P.; Yao, W.; Wang, J.; Wang, L.; Deng, L.; He, J.; Zhang, G.; Lei, J. Subcritical Water Extraction of Betulinic Acid from Birch Bark. Ind. Crop. Prod. 2015, 74, 557–565. [Google Scholar] [CrossRef]
- Zhao, G.; Yan, W.; Cao, D. Simultaneous Determination of Betulin and Betulinic Acid in White Birch Bark Using RP-HPLC. J. Pharm. Biomed. Anal. 2007, 43, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Omori, S. A Simple Preparation of Betulinic Acid from Sycamore Bark. J. Wood Sci. 2012, 58, 169–173. [Google Scholar] [CrossRef]
- Urban, M.; Sarek, J.; Klinot, J.; Korinkova, G.; Hajduch, M. Synthesis of A-Seco Derivatives of Betulinic Acid with Cytotoxic Activity. J. Nat. Prod. 2004, 67, 1100–1105. [Google Scholar] [CrossRef]
- Mullally, M.; Kramp, K.; Cayer, C.; Saleem, A.; Ahmed, F.; McRae, C.; Baker, J.; Goulah, A.; Otorola, M.; Sanchez, P.; et al. Anxiolytic Activity of a Supercritical Carbon Dioxide Extract of Souroubea Sympetala (Marcgraviaceae). Phytother. Res. 2011, 25, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Patinha, D.J.S.; Domingues, R.M.A.; Villaverde, J.J.; Silva, A.M.S.; Silva, C.M.; Freire, C.S.R.; Neto, C.P.; Silvestre, A.J.D. Lipophilic Extractives from the Bark of Eucalyptus Grandis x Globulus, a Rich Source of Methyl Morolate: Selective Extraction with Supercritical CO2. Ind. Crop. Prod. 2013, 43, 340–348. [Google Scholar] [CrossRef]
- Silva, N.H.C.S.; Morais, E.S.; Freire, C.S.R.; Freire, M.G.; Silvestre, A.J.D. Extraction of High Value Triterpenic Acids from Eucalyptus Globulus Biomass Using Hydrophobic Deep Eutectic Solvents. Molecules 2020, 25, 210. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, S.A.; Skvortsova, G.P.; Maliar, I.N.; Skurydina, E.S.; Veselova, O.F. Extraction of Betulin from Birch Bark and Study of Its Physico-Chemical and Pharmacological Properties. Russ. J. Bioorg. Chem. 2014, 40, 742–747. [Google Scholar] [CrossRef]
- Mazumder, K.; Biswas, B.; Raja, I.M.; Fukase, K. A Review of Cytotoxic Plants of the Indian Subcontinent and a Broad-Spectrum Analysis of Their Bioactive Compounds. Molecules 2020, 25, 1904. [Google Scholar] [CrossRef] [PubMed]
- Räsänen, R.-M.; Hieta, J.-P.; Immanen, J.; Nieminen, K.; Haavikko, R.; Yli-Kauhaluoma, J.; Kauppila, T.J. Chemical Profiles of Birch and Alder Bark by Ambient Mass Spectrometry. Anal. Bioanal. Chem. 2019, 411, 7573–7583. [Google Scholar] [CrossRef] [PubMed]
- Cîntă-Pînzaru, S.; Dehelean, C.A.; Soica, C.; Culea, M.; Borcan, F. Evaluation and Differentiation of the Betulaceae Birch Bark Species and Their Bioactive Triterpene Content Using Analytical FT-Vibrational Spectroscopy and GC-MS. Chem. Cent. J. 2012, 6, 67. [Google Scholar] [CrossRef]
- Baltina, L.A.; Flekhter, O.B.; Nigmatullina, L.R.; Boreko, E.I.; Pavlova, N.I.; Nikolaeva, S.N.; Savinova, O.V.; Tolstikov, G.A. Lupane Triterpenes and Derivatives with Antiviral Activity. Bioorganic Med. Chem. Lett. 2003, 13, 3549–3552. [Google Scholar] [CrossRef]
- Csuk, R.; Schmuck, K.; Schäfer, R. A Practical Synthesis of Betulinic Acid. Tetrahedron Lett. 2006, 47, 8769–8770. [Google Scholar] [CrossRef]
- Barthel, A.; Stark, S.; Csuk, R. Oxidative Transformations of Betulinol. Tetrahedron 2008, 64, 9225–9229. [Google Scholar] [CrossRef]
- Ressmann, A.K.; Kremsmayr, T.; Gaertner, P.; Zirbs, R.; Bica, K. Toward a Benign Strategy for the Manufacturing of Betulinic Acid. Green Chem. 2017, 19, 1014–1022. [Google Scholar] [CrossRef]
- Kim, D.S.H.L.; Chen, Z.; Nguyen, T.; Pezzuto, J.M.; Qiu, S.; Lu, Z.-Z. A Concise Semi-Synthetic Approach to Betulinic Acid from Betulin. Synth. Commun. 1997, 27, 1607–1612. [Google Scholar] [CrossRef]
- Li, T.-S.; Wang, J.-X.; Zheng, X.-J. Simple Synthesis of Allobetulin, 28-Oxyallobetulin and Related Biomarkers from Betulin and Betulinic Acid Catalysed by Solid Acids. J. Chem. Soc. Perkin Trans. 1998, 3957–3966. [Google Scholar] [CrossRef]
- Kvasnica, M.; Urban, M.; Dickinson, N.J.; Sarek, J. Pentacyclic Triterpenoids with Nitrogen- and Sulfur-Containing Heterocycles: Synthesis and Medicinal Significance. Nat. Prod. Rep. 2015, 32, 1303–1330. [Google Scholar] [CrossRef]
- Borkova, L.; Hodon, J.; Urban, M. Synthesis of Betulinic Acid Derivatives with Modified A-Rings and Their Application as Potential Drug Candidates. Asian J. Org. Chem. 2018, 7, 1542–1560. [Google Scholar] [CrossRef]
- Sousa, J.L.C.; Freire, C.S.R.; Silvestre, A.J.D.; Silva, A.M.S. Recent Developments in the Functionalization of Betulinic Acid and Its Natural Analogues: A Route to New Bioactive Compounds. Molecules 2019, 24, 355. [Google Scholar] [CrossRef]
- Swidorski, J.J.; Liu, Z.; Sit, S.-Y.; Chen, J.; Chen, Y.; Sin, N.; Venables, B.L.; Parker, D.D.; Nowicka-Sans, B.; Terry, B.J.; et al. Inhibitors of HIV-1 Maturation: Development of Structure–Activity Relationship for C-28 Amides Based on C-3 Benzoic Acid-Modified Triterpenoids. Bioorganic Med. Chem. Lett. 2016, 26, 1925–1930. [Google Scholar] [CrossRef]
- Tolmacheva, I.A.; Igosheva, E.V.; Vikharev, I.B.; Grishko, V.V.; Savinova, O.V.; Boreko, E.I.; Eremin, V.F. Synthesis and biological activity of 2,3-secotriterpene acid mono- and diamides. Bioorg. Khim. 2013, 39, 212–220. [Google Scholar] [CrossRef]
- Bildziukevich, U.; Rárová, L.; Šaman, D.; Wimmer, Z. Picolyl Amides of Betulinic Acid as Antitumor Agents Causing Tumor Cell Apoptosis. Eur. J. Med. Chem. 2018, 145, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Dang Thi, T.A.; Kim Tuyet, N.T.; Pham The, C.; Thanh Nguyen, H.; Ba Thi, C.; Thi Phuong, H.; Van Boi, L.; Van Nguyen, T.; D’hooghe, M. Synthesis and Cytotoxic Evaluation of Novel Amide–Triazole-Linked Triterpenoid–AZT Conjugates. Tetrahedron Lett. 2015, 56, 218–224. [Google Scholar] [CrossRef]
- Sorokina, I.V.; Baev, D.S.; Zhukova, N.A.; Tolstikova, T.G.; Antimonova, A.N.; Petrenko, N.I.; Shul’ts, E.E.; Grigor’ev, I.A. Hepatoprotective activity of betulonic acid amides containing a piperidine or pyrrolidine nitroxide moiety. Bioorg. Khim. 2013, 39, 749–752. [Google Scholar] [CrossRef] [PubMed]
- Bildziukevich, U.; Kaletová, E.; Šaman, D.; Sievänen, E.; Kolehmainen, E.T.; Šlouf, M.; Wimmer, Z. Spectral and Microscopic Study of Self-Assembly of Novel Cationic Spermine Amides of Betulinic Acid. Steroids 2017, 117, 90–96. [Google Scholar] [CrossRef]
- Xiao, S.; Wang, Q.; Si, L.; Shi, Y.; Wang, H.; Yu, F.; Zhang, Y.; Li, Y.; Zheng, Y.; Zhang, C.; et al. Synthesis and Anti-HCV Entry Activity Studies of β-Cyclodextrin–Pentacyclic Triterpene Conjugates. ChemMedChem 2014, 9, 1060–1070. [Google Scholar] [CrossRef]
- Li, M.; Yuan, L.; Chen, Y.; Ma, W.; Ran, F.; Zhang, L.; Zhou, D.; Xiao, S. Rhodamine B-Based Fluorescent Probes for Molecular Mechanism Study of the Anti-Influenza Activity of Pentacyclic Triterpenes. Eur. J. Med. Chem. 2020, 205, 112664. [Google Scholar] [CrossRef]
- Wiemann, J.; Heller, L.; Perl, V.; Kluge, R.; Ströhl, D.; Csuk, R. Betulinic Acid Derived Hydroxamates and Betulin Derived Carbamates Are Interesting Scaffolds for the Synthesis of Novel Cytotoxic Compounds. Eur. J. Med. Chem. 2015, 106, 194–210. [Google Scholar] [CrossRef]
- Cui, H.-W.; He, Y.; Wang, J.; Gao, W.; Liu, T.; Qin, M.; Wang, X.; Gao, C.; Wang, Y.; Liu, M.-Y.; et al. Synthesis of Heterocycle-Modified Betulinic Acid Derivatives as Antitumor Agents. Eur. J. Med. Chem. 2015, 95, 240–248. [Google Scholar] [CrossRef]
- Smith, P.F.; Ogundele, A.; Forrest, A.; Wilton, J.; Salzwedel, K.; Doto, J.; Allaway, G.P.; Martin, D.E. Phase I and II Study of the Safety, Virologic Effect, and Pharmacokinetics/Pharmacodynamics of Single-Dose 3-O-(3′,3′-Dimethylsuccinyl)Betulinic Acid (Bevirimat) against Human Immunodeficiency Virus Infection. Antimicrob. Agents Chemother. 2007, 51, 3574–3581. [Google Scholar] [CrossRef]
- Hashimoto, F.; Kashiwada, Y.; Cosentino, L.M.; Chen, C.-H.; Garrett, P.E.; Lee, K.-H. Anti-AIDS Agents—XXVII. Synthesis and Anti-HIV Activity of Betulinic Acid and Dihydrobetulinic Acid Derivatives. Bioorganic Med. Chem. 1997, 5, 2133–2143. [Google Scholar] [CrossRef]
- Khan, I.; Guru, S.K.; Rath, S.K.; Chinthakindi, P.K.; Singh, B.; Koul, S.; Bhushan, S.; Sangwan, P.L. A Novel Triazole Derivative of Betulinic Acid Induces Extrinsic and Intrinsic Apoptosis in Human Leukemia HL-60 Cells. Eur. J. Med. Chem. 2016, 108, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Urban, M.; Sarek, J.; Tislerova, I.; Dzubak, P.; Hajduch, M. Influence of Esterification and Modification of A-Ring in a Group of Lupane Acids on Their Cytotoxicity. Bioorganic Med. Chem. 2005, 13, 5527–5535. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.C.; Salvador, J.A.R.; Marín, S.; Cascante, M.; Moreira, J.N.; Dinis, T.C.P. Synthesis and Structure–Activity Relationship Study of Novel Cytotoxic Carbamate and N-Acylheterocyclic Bearing Derivatives of Betulin and Betulinic Acid. Bioorganic Med. Chem. 2010, 18, 4385–4396. [Google Scholar] [CrossRef]
- Ma, C.; Nakamura, N.; Miyashiro, H.; Hattori, M.; Shimotohno, K. Inhibitory Effects of Constituents from Cynomorium Songaricum and Related Triterpene Derivatives on HIV-1 Protease. Chem. Pharm. Bull. 1999, 47, 141–145. [Google Scholar] [CrossRef]
- Perlikova, P.; Kvasnica, M.; Urban, M.; Hajduch, M.; Sarek, J. 2-Deoxyglycoside Conjugates of Lupane Triterpenoids with High Cytotoxic Activity—Synthesis, Activity, and Pharmacokinetic Profile. Bioconjugate Chem. 2019, 30, 2844–2858. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, C.; Legault, J.; Piochon-Gauthier, M.; Pichette, A. Advances in the Synthesis and Pharmacological Activity of Lupane-Type Triterpenoid Saponins. Phytochem. Rev. 2011, 10, 521–544. [Google Scholar] [CrossRef]
- Flekhter, O.B.; Baltina, L.A.; Tolstikov, G.A. Direct Stereospecific Synthesis of Triterpene and Steroid 2-Deoxy-α-Glycosides. Russ. Chem. Bull. 1997, 46, 1335–1338. [Google Scholar] [CrossRef]
- Flekhter, O.B.; Baltina, L.A.; Tolstikov, G.A. Glycals in the Stereoselective Synthesis of Triterpene 2-Deoxy-α-l-Glycosides under Conditions of Acidic Catalysis. J. Nat. Prod. 2000, 63, 992–994. [Google Scholar] [CrossRef]
- Samoshina, N.F.; Denisenko, M.V.; Denisenko, V.A.; Uvarova, N.I. Synthesis of Glycosides of Lupane-Type Triterpene Acids. Chem. Nat. Compd. 2003, 39, 575–582. [Google Scholar] [CrossRef]
- Eignerova, B.; Tichy, M.; Krasulova, J.; Kvasnica, M.; Rarova, L.; Christova, R.; Urban, M.; Bednarczyk-Cwynar, B.; Hajduch, M.; Sarek, J. Synthesis and Antiproliferative Properties of New Hydrophilic Esters of Triterpenic Acids. Eur. J. Med. Chem. 2017, 140, 403–420. [Google Scholar] [CrossRef] [PubMed]
- Milner, S.E.; Maguire, A.R. Recent Trends in Whole Cell and Isolated Enzymes in Enantioselective Synthesis. Arkivoc 2012, 2012, 321–382. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, H.; Ang, E.L.; Zhao, H. Biocatalysis for the Synthesis of Pharmaceuticals and Pharmaceutical Intermediates. Bioorganic Med. Chem. 2018, 26, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Christianson, D.W. Structural and Chemical Biology of Terpenoid Cyclases. Chem. Rev. 2017, 117, 11570–11648. [Google Scholar] [CrossRef]
- Abe, I. Enzymatic Synthesis of Cyclic Triterpenes. Nat. Prod. Rep. 2007, 24, 1311–1331. [Google Scholar] [CrossRef]
- An, T.; Zha, W.; Zi, J. Biotechnological Production of Betulinic Acid and Derivatives and Their Applications. Appl. Microbiol. Biotechnol. 2020, 104, 3339–3348. [Google Scholar] [CrossRef]
- Herrera, J.B.R.; Bartel, B.; Wilson, W.K.; Matsuda, S.P.T. Cloning and Characterization of the Arabidopsis Thaliana Lupeol Synthase Gene. Phytochemistry 1998, 49, 1905–1911. [Google Scholar] [CrossRef]
- Shibuya, M.; Zhang, H.; Endo, A.; Shishikura, K.; Kushiro, T.; Ebizuka, Y. Two Branches of the Lupeol Synthase Gene in the Molecular Evolution of Plant Oxidosqualene Cyclases. Eur. J. Biochem. 1999, 266, 302–307. [Google Scholar] [CrossRef]
- Kushiro, T.; Shibuya, M.; Masuda, K.; Ebizuka, Y. A Novel Multifunctional Triterpene Synthase from Arabidopsis Thaliana. Tetrahedron Lett. 2000, 41, 7705–7710. [Google Scholar] [CrossRef]
- Zhang, H.; Shibuya, M.; Yokota, S.; Ebizuka, Y. Oxidosqualene Cyclases from Cell Suspension Cultures of Betula Platyphylla Var. Japonica: Molecular Evolution of Oxidosqualene Cyclases in Higher Plants. Biol. Pharm. Bull. 2003, 26, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Ebizuka, Y.; Katsube, Y.; Tsutsumi, T.; Kushiro, T.; Shibuya, M. Functional genomics approach to the study of triterpene biosynthesis. Pure Appl. Chem. 2003, 75, 369–374. [Google Scholar] [CrossRef]
- Iturbe-Ormaetxe, I.; Haralampidis, K.; Papadopoulou, K.; Osbourn, A.E. Molecular Cloning and Characterization of Triterpene Synthases from Medicago Truncatula and Lotus Japonicus. Plant Mol. Biol. 2003, 51, 731–743. [Google Scholar] [CrossRef]
- Hayashi, H.; Huang, P.; Takada, S.; Obinata, M.; Inoue, K.; Shibuya, M.; Ebizuka, Y. Differential Expression of Three Oxidosqualene Cyclase MRNAs in Glycyrrhiza Glabra. Biol. Pharm. Bull. 2004, 27, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Guhling, O.; Hobl, B.; Yeats, T.; Jetter, R. Cloning and Characterization of a Lupeol Synthase Involved in the Synthesis of Epicuticular Wax Crystals on Stem and Hypocotyl Surfaces of Ricinus Communis. Arch. Biochem. Biophys. 2006, 448, 60–72. [Google Scholar] [CrossRef]
- Sawai, S.; Shindo, T.; Sato, S.; Kaneko, T.; Tabata, S.; Ayabe, S.; Aoki, T. Functional and Structural Analysis of Genes Encoding Oxidosqualene Cyclases of Lotus Japonicus. Plant Sci. 2006, 170, 247–257. [Google Scholar] [CrossRef]
- Basyuni, M.; Oku, H.; Inafuku, M.; Baba, S.; Iwasaki, H.; Oshiro, K.; Okabe, T.; Shibuya, M.; Ebizuka, Y. Molecular Cloning and Functional Expression of a Multifunctional Triterpene Synthase CDNA from a Mangrove Species Kandelia candel (L.) Druce. Phytochemistry 2006, 67, 2517–2524. [Google Scholar] [CrossRef] [PubMed]
- Basyuni, M.; Oku, H.; Tsujimoto, E.; Kinjo, K.; Baba, S.; Takara, K. Triterpene Synthases from the Okinawan Mangrove Tribe, Rhizophoraceae. FEBS J. 2007, 274, 5028–5042. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yeats, T.; Han, H.; Jetter, R. Cloning and Characterization of Oxidosqualene Cyclases from Kalanchoe Daigremontiana: Enzymes Catalyzing Up to 10 Rearrangement Steps Yielding Friedelin and Other Triterpenoids. J. Biol. Chem. 2010, 285, 29703–29712. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Ren, C.-L.; Zhan, Y.-G.; Li, C.-X.; Xiao, J.-L.; Qiu, W.; Li, X.-Y.; Peng, H.-M. Distribution and Expression Characteristics of Triterpenoids and OSC Genes in White Birch (Betula Platyphylla Suk.). Mol. Biol. Rep. 2012, 39, 2321–2328. [Google Scholar] [CrossRef] [PubMed]
- Khakimov, B.; Kuzina, V.; Erthmann, P.Ø.; Fukushima, E.O.; Augustin, J.M.; Olsen, C.E.; Scholtalbers, J.; Volpin, H.; Andersen, S.B.; Hauser, T.P.; et al. Identification and Genome Organization of Saponin Pathway Genes from a Wild Crucifer, and Their Use for Transient Production of Saponins in Nicotiana Benthamiana. Plant J. 2015, 84, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, E.O.; Seki, H.; Ohyama, K.; Ono, E.; Umemoto, N.; Mizutani, M.; Saito, K.; Muranaka, T. CYP716A Subfamily Members Are Multifunctional Oxidases in Triterpenoid Biosynthesis. Plant Cell Physiol. 2011, 52, 2050–2061. [Google Scholar] [CrossRef]
- Huang, L.; Li, J.; Ye, H.; Li, C.; Wang, H.; Liu, B.; Zhang, Y. Molecular Characterization of the Pentacyclic Triterpenoid Biosynthetic Pathway in Catharanthus Roseus. Planta 2012, 236, 1571–1581. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Li, J.; Li, C.; Zhang, Y. Improvement of Betulinic Acid Biosynthesis in Yeast Employing Multiple Strategies. BMC Biotechnol. 2016, 16, 59. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Seki, H.; Suzuki, H.; Kojoma, M.; Saito, K.; Muranaka, T. CYP716A179 Functions as a Triterpene C-28 Oxidase in Tissue-Cultured Stolons of Glycyrrhiza Uralensis. Plant Cell Rep. 2017, 36, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zha, W.; An, T.; Dong, H.; Huang, Y.; Wang, D.; Yu, R.; Duan, L.; Zhang, X.; Peters, R.J.; et al. Identification of RoCYP01 (CYP716A155) Enables Construction of Engineered Yeast for High-Yield Production of Betulinic Acid. Appl. Microbiol. Biotechnol. 2019, 103, 7029–7039. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Fukushima, E.O.; Shimizu, Y.; Seki, H.; Fujisawa, Y.; Ishimoto, M.; Osakabe, K.; Osakabe, Y.; Muranaka, T. Lotus Japonicus Triterpenoid Profile and Characterization of the CYP716A51 and LjCYP93E1 Genes Involved in Their Biosynthesis In Planta. Plant Cell Physiol. 2019, 60, 2496–2509. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fu, M.L.; Chen, Q.H. Biotransformation Optimization of Betulin into Betulinic Acid Production Catalysed by Cultured Armillaria Luteo-Virens Sacc ZJUQH100-6 Cells. J. Appl. Microbiol. 2011, 110, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.-H.; Feng, Y.-Q.; Mao, D.-B.; Xu, C.-P. Optimization for Betulin Production from Mycelial Culture of Inonotus Obliquus by Orthogonal Design and Evaluation of Its Antioxidant Activity. J. Taiwan Inst. Chem. Eng. 2012, 43, 663–669. [Google Scholar] [CrossRef]
- Wang, L.-X.; Lu, Z.-M.; Geng, Y.; Zhang, X.-M.; Xu, G.-H.; Shi, J.-S.; Xu, Z.-H. Stimulated Production of Steroids in Inonotus Obliquus by Host Factors from Birch. J. Biosci. Bioeng. 2014, 118, 728–731. [Google Scholar] [CrossRef]
- Bertolo, A.P.; Biz, A.P.; Kempka, A.P.; Rigo, E.; Cavalheiro, D. Yeast (Saccharomyces Cerevisiae): Evaluation of Cellular Disruption Processes, Chemical Composition, Functional Properties and Digestibility. J. Food Sci. Technol. 2019, 56, 3697–3706. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y. Increase of Betulinic Acid Production in Saccharomyces Cerevisiae by Balancing Fatty Acids and Betulinic Acid Forming Pathways. Appl. Microbiol. Biotechnol. 2014, 98, 3081–3089. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y. Modulating Betulinic Acid Production in Saccharomyces Cerevisiae by Managing the Intracellular Supplies of the Co-Factor NADPH and Oxygen. J. Biosci. Bioeng. 2015, 119, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-T.; Wang, D.; Dai, Z.-B.; Zhang, X.-L.; Huang, L.-Q. Construction of cell factories for production of lupeol in Saccharomyces cerevisiae. Zhongguo Zhong Yao Za Zhi 2016, 41, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Czarnotta, E.; Dianat, M.; Korf, M.; Granica, F.; Merz, J.; Maury, J.; Jacobsen, S.A.B.; Förster, J.; Ebert, B.E.; Blank, L.M. Fermentation and Purification Strategies for the Production of Betulinic Acid and Its Lupane-Type Precursors in Saccharomyces Cerevisiae. Biotechnol. Bioeng. 2017, 114, 2528–2538. [Google Scholar] [CrossRef]
- Arendt, P.; Miettinen, K.; Pollier, J.; De Rycke, R.; Callewaert, N.; Goossens, A. An Endoplasmic Reticulum-Engineered Yeast Platform for Overproduction of Triterpenoids. Metab. Eng. 2017, 40, 165–175. [Google Scholar] [CrossRef]
- D’Adamo, S.; Schiano di Visconte, G.; Lowe, G.; Szaub-Newton, J.; Beacham, T.; Landels, A.; Allen, M.J.; Spicer, A.; Matthijs, M. Engineering the Unicellular Alga Phaeodactylum Tricornutum for High-Value Plant Triterpenoid Production. Plant Biotechnol. J. 2019, 17, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Qiao, W.; Zhou, Z.; Liang, Q.; Mosongo, I.; Li, C.; Zhang, Y. Improving Lupeol Production in Yeast by Recruiting Pathway Genes from Different Organisms. Sci. Rep. 2019, 9, 2992. [Google Scholar] [CrossRef]
- Zieniuk, B.; Fabiszewska, A. Yarrowia Lipolytica: A Beneficious Yeast in Biotechnology as a Rare Opportunistic Fungal Pathogen: A Minireview. World J. Microbiol. Biotechnol. 2019, 35. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, C.; Nan, W.; Li, D.; Ke, D.; Lu, W. Glycerol Improves Heterologous Biosynthesis of Betulinic Acid in Engineered Yarrowia Lipolytica. Chem. Eng. Sci. 2019, 196, 82–90. [Google Scholar] [CrossRef]
- Jin, C.-C.; Zhang, J.-L.; Song, H.; Cao, Y.-X. Boosting the Biosynthesis of Betulinic Acid and Related Triterpenoids in Yarrowia Lipolytica via Multimodular Metabolic Engineering. Microb. Cell Factories 2019, 18, 77. [Google Scholar] [CrossRef]
- Gowers, G.-O.F.; Chee, S.M.; Bell, D.; Suckling, L.; Kern, M.; Tew, D.; McClymont, D.W.; Ellis, T. Improved Betulinic Acid Biosynthesis Using Synthetic Yeast Chromosome Recombination and Semi-Automated Rapid LC-MS Screening. Nat. Commun. 2020, 11, 868. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.; Pezzuto, J.M.; Kouzi, S.A. Glucosidation of Betulinic Acid by Cunninghamella Species. J. Nat. Prod. 1999, 62, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Yasin, Y.; Basri, M.; Ahmad, F.; Salleh, A.B. Response Surface Methodology as a Tool to Study the Lipase-Catalyzed Synthesis of Betulinic Acid Ester. J. Chem. Technol. Biotechnol. 2008, 83, 694–698. [Google Scholar] [CrossRef]
- Ahmad, F.B.H.; Moghaddam, M.G.; Basri, M.; Rahman, M.B.A. Enzymatic Synthesis of Betulinic Acid Ester as an Anticancer Agent: Optimization Study. Biocatal. Biotransform. 2010, 28, 192–200. [Google Scholar] [CrossRef]
- Ahmad, F.B.H.; Moghaddam, M.G.; Basri, M.; Rahman, M.B.A. Anticancer Activity of 3-O-Acylated Betulinic Acid Derivatives Obtained by Enzymatic Synthesis. Biosci. Biotechnol. Biochem. 2010, 74, 1025–1029. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.-B.; Feng, Y.-Q.; Bai, Y.-H.; Xu, C.-P. Novel Biotransformation of Betulin to Produce Betulone by Rhodotorula Mucilaginosa. J. Taiwan Inst. Chem. Eng. 2012, 43, 825–829. [Google Scholar] [CrossRef]
- Atikah Binti Amin Yusof, N.; Mat Hadzir, N.; Efliza Ashari, S. Identification and Optimisation of Lipase-Catalysed Synthesis of Betulinic Acid Amide in a Solvent System. J. Appl. Chem. 2016, 2016, e5149326. [Google Scholar] [CrossRef]
- Guo, B.; Xu, D.; Liu, X.; Yi, J. Enzymatic Synthesis and in Vitro Evaluation of Folate-Functionalized Liposomes. Drug Des Devel. 2017, 11, 1839–1847. [Google Scholar] [CrossRef]
- Dai, Z.; Liu, Y.; Sun, Z.; Wang, D.; Qu, G.; Ma, X.; Fan, F.; Zhang, L.; Li, S.; Zhang, X. Identification of a Novel Cytochrome P450 Enzyme That Catalyzes the C-2α Hydroxylation of Pentacyclic Triterpenoids and Its Application in Yeast Cell Factories. Metab. Eng. 2019, 51, 70–78. [Google Scholar] [CrossRef]
- Gauthier, C.; Legault, J.; Rondeau, S.; Pichette, A. Synthesis of Betulinic Acid Acyl Glucuronide for Application in Anticancer Prodrug Monotherapy. Tetrahedron Lett. 2009, 50, 988–991. [Google Scholar] [CrossRef]
- Chatterjee, P.; Kouzi, S.A.; Pezzuto, J.M.; Hamann, M.T. Biotransformation of the Antimelanoma Agent Betulinic Acid by Bacillus Megaterium ATCC 13368. Appl. Environ. Microbiol. 2000, 66, 3850–3855. [Google Scholar] [CrossRef] [PubMed]
- Kouzi, S.A.; Chatterjee, P.; Pezzuto, J.M.; Hamann, M.T. Microbial Transformations of the Antimelanoma Agent Betulinic Acid. J. Nat. Prod. 2000, 63, 1653–1657. [Google Scholar] [CrossRef] [PubMed]
- Bastos, D.Z.L.; Pimentel, I.C.; de Jesus, D.A.; de Oliveira, B.H. Biotransformation of Betulinic and Betulonic Acids by Fungi. Phytochemistry 2007, 68, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Goswami, A.; Guo, Z.; Tully, T.P.; Rinaldi, F.A.; Huang, X.S.; Swidorski, J.J.; Regueiro-Ren, A. Microbial Transformations of Betulinic and Betulonic Acids. J. Mol. Catal. B Enzym. 2015, 117, 45–53. [Google Scholar] [CrossRef]
- Chen, C.; Song, K.; Zhang, Y.; Chu, C.; Fan, B.; Song, Y.; Huang, H.; Chen, G. Biotransformation of Betulinic Acid by Circinella Muscae and Cunninghamella Echinulata to Discover Anti-Inflammatory Derivatives. Phytochemistry 2021, 182, 112608. [Google Scholar] [CrossRef]
- Okamoto, W.; Sato, T. Enzymatic Syntheses of Unnatural Head-to-Tail Pentacyclic Triterpenes by Tetraprenyl-β-Curcumene Cyclase. Tetrahedron Lett. 2013, 54, 6747–6750. [Google Scholar] [CrossRef]


















| Plant Source | Year | Extraction Medium | Compounds | Reference |
|---|---|---|---|---|
| Gratiola officinalis | 1902 | methanol | 5 | [32] |
| Vauquelinia corymbosa | 1976 | chloroform | 5 | [33] |
| Syzigium claviflorum | 1994 | methanol | 5 and platanic acid | [34] |
| Eucalyptus globulus | 2002 | dichloromethane | 5 and other terpenoids | [37] |
| Rosmarinus officinalis | 2002 | methanol | 5 | [38] |
| Betula platyphylla | 2006 | various organic solvents | 5; 8 | [42] |
| Betula—various species | 2014 | ethanol | 8 | [48] |
| Platanus occidentalis | 2011 | water and organic solvents | 8 | [43] |
| Platanus hispanica | 2004 | methanol | 5 | [44] |
| Souroubea sympetala | 2011 | carbon dioxide | 5 | [45] |
| Eucalyptus grandis x globulus | 2013 | supercritical carbon dioxide | 5 and other terpenic acids | [46] |
| purchased birch bark | 2015 | subcritical water | 5 | [41] |
| Eucalyptus globulus | 2020 | hydrophobic deep eutectic solvents | 5 and other terpenic acids | [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bachořík, J.; Urban, M. Biocatalysis in the Chemistry of Lupane Triterpenoids. Molecules 2021, 26, 2271. https://doi.org/10.3390/molecules26082271
Bachořík J, Urban M. Biocatalysis in the Chemistry of Lupane Triterpenoids. Molecules. 2021; 26(8):2271. https://doi.org/10.3390/molecules26082271
Chicago/Turabian StyleBachořík, Jan, and Milan Urban. 2021. "Biocatalysis in the Chemistry of Lupane Triterpenoids" Molecules 26, no. 8: 2271. https://doi.org/10.3390/molecules26082271
APA StyleBachořík, J., & Urban, M. (2021). Biocatalysis in the Chemistry of Lupane Triterpenoids. Molecules, 26(8), 2271. https://doi.org/10.3390/molecules26082271