
Genomic and Metabolomic Investigation of a Rhizosphere Isolate Streptomyces netropsis WLXQSS-4 Associated with a Traditional Chinese Medicine

 and
and
Abstract
1. Introduction
2. Results
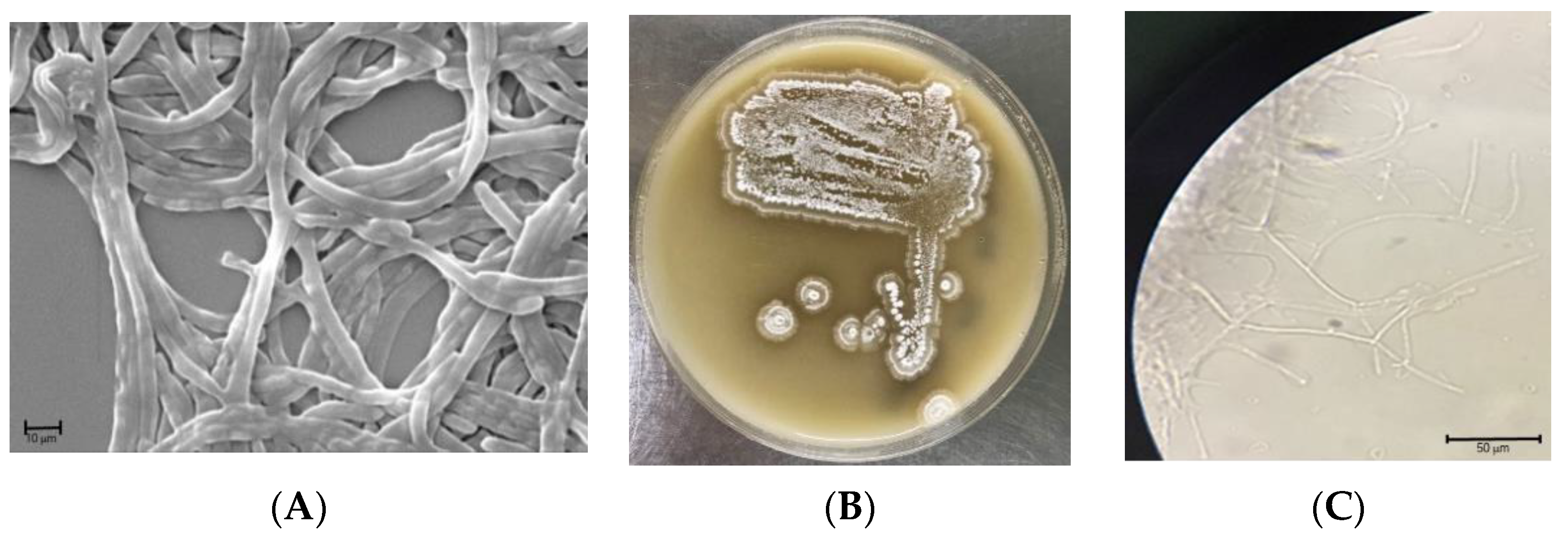
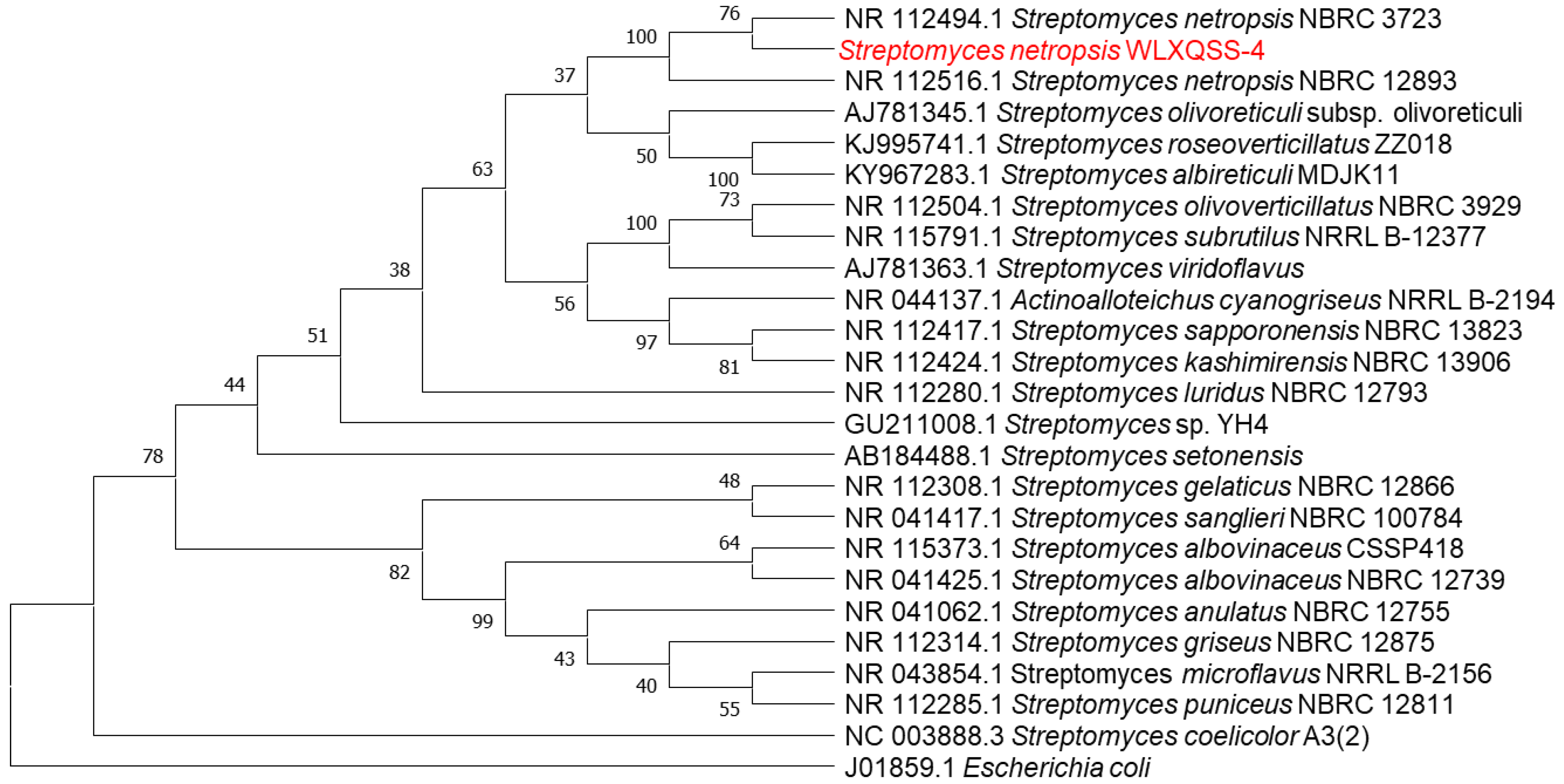
2.1. General Characterization of S. netropsis WLXQSS-4
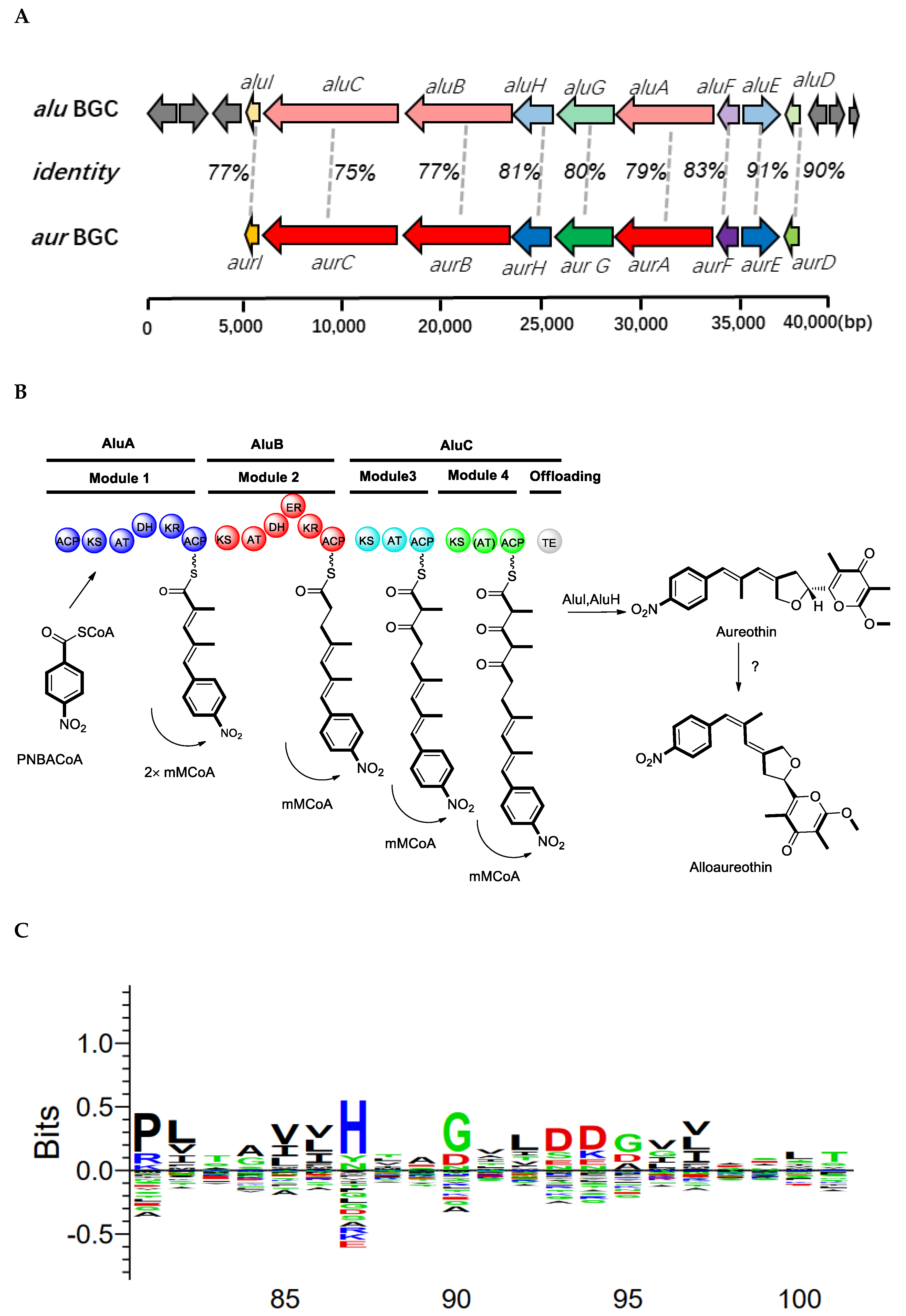
2.2. Genome Sequencing and Annotation of S. netropsis WLXQSS-4
2.3. Metabolome Investigation on Strain S. netropsis WLXQSS-4
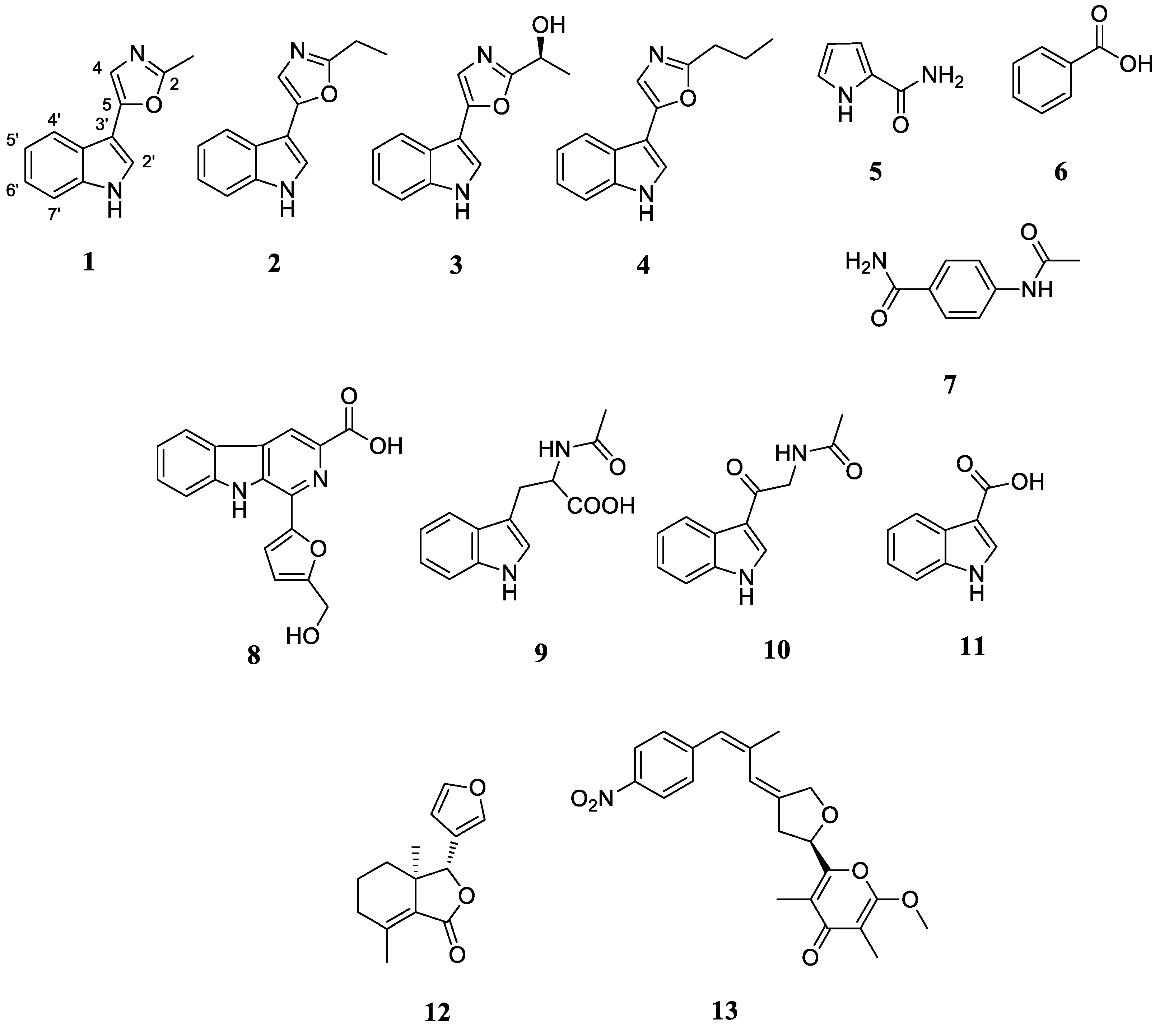
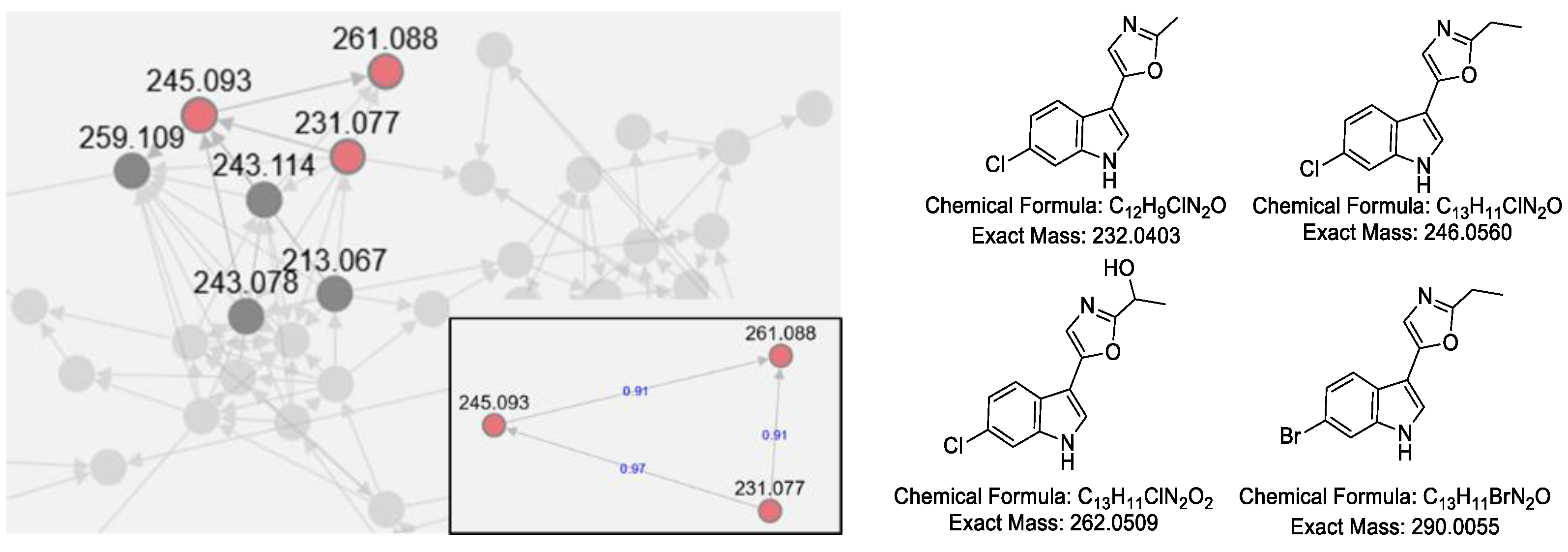
2.3.1. Analysis of the Secondary Metabolites of S. netropsis WLXQSS-4
2.3.2. Activation of a Cryptic Gene Cluster of S. netropsis WLXQSS-4 by Medium Screening
2.3.3. Structural Elucidation
2.4. Biological Activities
3. Discussion
4. Materials and Methods
4.1. Isolation of Strain
4.2. Morphological Identification
4.3. Fermentation in a Medium
4.4. Isolation of Secondary Metabolites (SM) from This Strain Fermentation Broth in a Medium
4.5. Fermentation in NL-19 Medium
4.6. Isolation of SM from This Strain Fermentation Broth in NL-19 Medium
4.7. HR-LCMS Analysis and Generation of the Molecular Networking
4.8. Scanning Electron Microscopy
4.9. Genome Sequencing and Annotation
4.10. Cytotoxicity Assay
4.11. Antibacterial Activity Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Vishwakarma, K.; Kumar, N.; Shandilya, C.; Mohapatra, S.; Bhayana, S.; Varma, A. Revisiting plant–microbe interactions and microbial consortia application for enhancing sustainable agriculture: A review. Front. Microbiol. 2020, 11, 560406. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Babalola, O.O. Streptomyces: Implications and interactions in plant growth promotion. Appl. Microbiol. Biotechnol. 2019, 103, 1179–1188. [Google Scholar] [CrossRef]
- Qi, J.J.; Yao, H.Y.; Ma, X.J.; Zhou, L.L.; Li, X.N. Soil microbial community composition and diversity in the rhizosphere of a Chinese medicinal plant. Commun. Soil. Sci. Plan. 2009, 40, 1462–1482. [Google Scholar]
- Zhang, Y.H.; Zheng, L.L.; Zheng, Y.; Xue, S.; Zhang, J.X.; Huang, P.; Zhao, Y.H.; Hao, X.C.; He, Z.K.; Hu, Z.B.; et al. Insight into the assembly of root-associated microbiome in the medicinal plant Polygonum cuspidatum. Ind. Crop. Prod. 2020, 145, 112163. [Google Scholar] [CrossRef]
- Nakaew, N.; Lumyong, S.; Sloan, W.T.; Sungthong, R. Bioactivities and genome insights of a thermotolerant antibiotics-producing Streptomyces sp. TM32 reveal its potentials for novel drug discovery. MicrobiologyOpen 2019, 8, e842. [Google Scholar] [CrossRef]
- Zhao, K.; Li, J.; Zhang, X.; Chen, Q.; Liu, M.; Ao, X.; Gu, Y.; Liao, D.; Xu, K.; Ma, M.; et al. Actinobacteria associated with Glycyrrhiza inflata Bat. are diverse and have plant growth promoting and antimicrobial activity. Sci. Rep. 2018, 8, 13661. [Google Scholar]
- Shi, S.P.; Tu, P.F.; Dong, C.X.; Jiang, D. Alkaloids from Clematis manshurica Rupr. J. Asian. Nat. Prod. Res. 2006, 8, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic. Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Yu, Z.; Jiang, H.; Wang, L.; Yang, F.X.; Huang, J.P.; Liu, C.; Guo, X.; Xiang, W.; Huang, S.X. Dimeric pimprinine alkaloids from soil-derived Streptomyces sp. NEAU-C99. Front. Chem. 2020, 8, 95. [Google Scholar] [CrossRef]
- Grundmann, F.; Dill, V.; Dowling, A.; Thanwisai, A.; Bode, E.; Chantratita, N.; Ffrench-Constant, R.; Bode, H.B. Identification and isolation of insecticidal oxazoles from Pseudomonas spp. Beilstein. J. Org. Chem. 2012, 8, 749–752. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Zhang, Q.B.; Fang, C.Y.; Zhang, Y.L.; Ma, L.; Liu, Z.W.; Zhai, S.L.; Peng, J.; Zhang, L.P.; Zhu, W.M. Refactoring the concise biosynthetic pathway of cyanogramide unveils spirooxindole formation catalyzed by a P450 enzyme. Angew. Chem. Int. Edit. 2020, 59, 14065–14069. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring structural diversity of microbe secondary metabolites using OSMAC strategy: A literature review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef]
- Ueda, J.Y.; Hashimoto, J.; Nagai, A.; Nakashima, T.; Komaki, H.; Anzai, K.; Harayama, S.; Doi, T.; Takahashi, T.; Nagasawa, K.; et al. New aureothin derivative, alloaureothin, from Streptomyces sp. MM23. J. antibiot. (Tokyo) 2007, 60, 321–324. [Google Scholar] [CrossRef][Green Version]
- Peng, H.; Ishida, K.; Hertweck, C. Loss of single-domain function in a modular assembly line alters the size and shape of a complex polyketide. Angew. Chem. Int. Ed. Engl. 2019, 58, 18252–18256. [Google Scholar] [CrossRef] [PubMed]
- Baerga-Ortiz, A.; Popovic, B.; Siskos, A.P.; O’Hare, H.M.; Spiteller, D.; Williams, M.G.; Campillo, N.; Spencer, J.B.; Leadlay, P.F. Directed mutagenesis alters the stereochemistry of catalysis by isolated ketoreductase domains from the erythromycin polyketide synthase. Chem. Biol. 2006, 13, 277–285. [Google Scholar] [CrossRef]
- Keatinge-Clay, A.T. A tylosin ketoreductase reveals how chirality is determined in polyketides. Chem. Biol. 2007, 14, 898–908. [Google Scholar] [CrossRef]
- Palaniappan, N.; Alhamadsheh, M.M.; Reynolds, K.A. cis-Delta(2,3)-double bond of phoslactomycins is generated by a post-PKS tailoring enzyme. J. Am. Chem. Soc. 2008, 130, 12236–12237. [Google Scholar] [CrossRef]
- Kumar, D.; Sundaree, S.; Patel, G.; Kumar, A. A novel and short synthesis of naturally occurring 5-(3′-indolyl)oxazoles. J. Heterocyclic Chem. 2010, 47, 1425–1428. [Google Scholar] [CrossRef]
- Raju, R.; Gromyko, O.; Fedorenko, V.; Luzhetskyy, A.; Müller, R. Pimprinols A–C, from the terrestrial actinomycete, Streptomyces sp. Tetrahedron Lett. 2012, 53, 3009–3011. [Google Scholar] [CrossRef]
- Miyake, F.; Hashimoto, M.; Tonsiengsom, S.; Yakushijin, K.; Horne, D.A. Synthesis of 5-(3-indolyl)oxazole natural products. Structure revision of Almazole D. Tetrahedron 2010, 66, 4888–4893. [Google Scholar] [CrossRef]
- Noltemeyer, M.; Sheldrick, G.M.; Hoppe, H.U.; Zeeck, A. 2-Ethyl-5-(3-indolyl)oxazole from Streptomyces cinnamomeus discovered by chemical screening. Characterization and structure elucidation by X-ray analysis. J. Antibiot. 1982, 35, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Knight, J.C.; Herald, D.L.; Davenport, R.; Pettit, R.K.; Tucker, B.E.; Schmidt, J.M. Isolation of labradorins 1 and 2 from Pseudomonas syringae pv. coronafaciens. J. Nat. Prod. 2002, 65, 1793–1797. [Google Scholar] [CrossRef] [PubMed]
- Shaaban, K.A.; Saunders, M.A.; Zhang, Y.; Tran, T.; Elshahawi, S.I.; Ponomareva, L.V.; Wang, X.; Zhang, J.; Copley, G.C.; Sunkara, M.; et al. Spoxazomicin D and oxachelin C, potent neuroprotective carboxamides from the Appalachian coal fire-associated isolate Streptomyces sp. RM-14-6. J. Nat. Prod. 2017, 80, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Q.; Bao, T.; Bao, H.Y. Research on chemical constituents of fruiting body of Helvella lacunose. J. Fungal. Res. 2016, 14, 239–244. [Google Scholar]
- Syahrani, A.; Ratnasari, E.; Indrayanto, G.; Wilkins, A.L. Biotransformation of o- and p-aminobenzoic acids and N-acetyl p-aminobenzoic acid by cell suspension cultures of Solanum. Phytochemistry 1999, 51, 615–620. [Google Scholar] [CrossRef]
- Zhou, S.; Junge, K.; Addis, D.; Das, S.; Beller, M. A general and convenient catalytic synthesis of nitriles from amides and silanes. Org. Lett. 2009, 11, 2461–2464. [Google Scholar] [CrossRef]
- Zhou, H.; Jian, R.; Kang, J.; Huang, X.; Li, Y.; Zhuang, C.; Yang, F.; Zhang, L.; Fan, X.; Wu, T.; et al. Anti-inflammatory effects of caper (Capparis spinosa L.) fruit aqueous extract and the isolation of main phytochemicals. J. Agric. Food. Chem. 2010, 58, 12717–12721. [Google Scholar]
- Kytzia, A.; Korth, H.G.; de Groot, H.; Kirsch, M. Catecholamine-induced release of nitric oxide from N-nitrosotryptophan derivatives: A non-enzymatic method for catecholamine oxidation. Org. Biomol. Chem. 2006, 4, 257–267. [Google Scholar] [CrossRef]
- Martínez-Luis, S.; Gómez, J.F.; Spadafora, C.; Guzmán, H.M.; Gutiérrez, M. Antitrypanosomal alkaloids from the marine bacterium Bacillus pumilus. Molecules 2012, 17, 11146–11155. [Google Scholar] [CrossRef]
- Wang, R.P.; Lin, H.W.; Li, L.Z.; Gao, P.Y.; Xu, Y.; Song, S.J. Monoindole alkaloids from a marine sponge Mycale fibrexilis. Biochem. Syst. Ecol. 2012, 43, 210–213. [Google Scholar] [CrossRef]
- Nakatani, M.; Huang, R.C.; Okamura, H.; Iwagawa, T.; Tadera, K. Degraded limonoids from Melia azedarach. Phytochemistry 1998, 49, 1773–1776. [Google Scholar] [CrossRef]
- Yang, R.L.; Jia, T.L.; Zhang, R.Q. Microbial transformation of fraxinellone by Aspergillus niger. J. Asian. Nat. Prod. Res. 2005, 7, 843–845. [Google Scholar] [CrossRef]
- De Souza, R.; Beneduzi, A.; Ambrosini, A.; da Costa, P.B.; Meyer, J.; Vargas, L.K.; Schoenfeld, R.; Passaglia, L.M.P. The effect of plant growth-promoting rhizobacteria on the growth of rice (Oryza sativa L.) cropped in southern Brazilian fields. Plant Soil. 2012, 366, 585–603. [Google Scholar] [CrossRef]
- Fu, S.F.; Wei, J.Y.; Chen, H.W.; Liu, Y.Y.; Lu, H.Y.; Chou, J.Y. Indole-3-acetic acid: A widespread physiological code in interactions of fungi with other organisms. Plant Signal. Behav. 2015, 10, e1048052. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.d.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef]
- Joshi, B.S.; Taylor, W.I.; Bhate, D.S.; Karmarkar, S.S. The structure and synthesis of pimprinine. Tetrahedron 1963, 19, 1437–1439. [Google Scholar] [CrossRef]
- Roy, S.; Haque, S.; Gribble, G.W. Synthesis of novel oxazolyl-indoles. Synthesis 2006, 23, 3948–3954. [Google Scholar] [CrossRef]
- Miao, Y.P.; Wen, R.; Hitoshi, A.; Zhou, P.G. Synthesis and antioxidative activity of 2-substituted phenyl-5-(3′-indolyl) -oxazole derivatives. Acta. Pharmaceut. Sin. 2004, 39, 37–40. [Google Scholar]
- Zhang, M.Z.; Chen, Q.; Mulholland, N.; Beattie, D.; Irwin, D.; Gu, Y.C.; Yang, G.F.; Clough, J. Synthesis and fungicidal activity of novel pimprinine analogues. Eur. J. Med. Chem. 2012, 53, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, R.; Li, Y.; Li, S.; Yu, J.; Zhao, B.; Liao, A.; Wang, Y.; Wang, Z.; Lu, A.; et al. Discovery of pimprinine alkaloids as novel agents against a plant virus. J. Agric. Food. Chem. 2019, 67, 1795–1806. [Google Scholar] [CrossRef]
- Shirling, E.B.; Gottlieb, D. Methods for characterization of Streptomyces species. Int. J. Syst. Bacteriol. 1966, 16, 313–340. [Google Scholar] [CrossRef]
- Ishaque, N.M.; Burgsdorf, I.; Limlingan Malit, J.J.; Saha, S.; Teta, R.; Ewe, D.; Kannabiran, K.; Hrouzek, P.; Steindler, L.; Costantino, V.; et al. Isolation, genomic and metabolomic characterization of Streptomyces tendae VITAKN with quorum sensing inhibitory activity from southern India. Microorganisms 2020, 8, 121. [Google Scholar] [CrossRef] [PubMed]
- Supaphon, P.; Phongpaichit, S.; Rukachaisirikul, V.; Sakayaroj, J. Antimicrobial potential of endophytic fungi derived from three seagrass species: Cymodocea serrulata, Halophila ovalis and Thalassia hemprichii. PLoS ONE 2013, 8, e72520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Klementz, D.; Zhu, J.; Makitrynskyy, R.; Ola Pasternak, A.R.; Günther, S.; Zechel, D.L.; Bechthold, A. Genome mining reveals the origin of a bald phenotype and a cryptic nucleocidin gene cluster in Streptomyces asterosporus DSM 41452. J. Biotechnol. 2019, 292, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhu, J.; Zechel, D.L.; Jessen-Trefzer, C.; Eastman, R.T.; Paululat, T.; Bechthold, A. New WS9326A derivatives and one new annimycin derivative with antimalarial activity are produced by Streptomyces asterosporus DSM 41452 and its mutant. ChemBioChem 2018, 19, 272–279. [Google Scholar] [CrossRef]
- Galperin, M.Y.; Wolf, Y.I.; Makarova, K.S.; Vera Alvarez, R.; Landsman, D.; Koonin, E.V. COG database update: Focus on microbial diversity, model organisms, and widespread pathogens. Nucleic. Acids Res. 2021, 49, D274–D281. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic. Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA genes in genomic sequences. In Gene Prediction; Kollmar, M., Ed.; Springer: New York, NY, USA, 2019; pp. 1–14. [Google Scholar]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic. Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Skinnider, M.A.; Merwin, N.J.; Johnston, C.W.; Magarvey, N.A. PRISM 3: Expanded prediction of natural product chemical structures from microbial genomes. Nucleic. Acids Res. 2017, 45, W49–W54. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | Chromosome Characteristics |
|---|---|
| Genome topology | Linear |
| Chromosome size (bp) | 8,324,019 |
| Scaffolds | 3 |
| G+C content (%) | 71.32 |
| Protein-coding genes | 7115 |
| Gene density (per kb) | 0.85 |
| Average ORF size (bp) | 1018.05 |
| rRNA (16S-23S-5S) operons | 21 |
| sRNA | 1 |
| Number of tRNA | 78 |
| Secondary metabolites gene cluster | 40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Zhang, L.; Zhu, J.; Chen, H.; Chen, Z.; Si, T.; Liu, T. Genomic and Metabolomic Investigation of a Rhizosphere Isolate Streptomyces netropsis WLXQSS-4 Associated with a Traditional Chinese Medicine. Molecules 2021, 26, 2147. https://doi.org/10.3390/molecules26082147
Zhang S, Zhang L, Zhu J, Chen H, Chen Z, Si T, Liu T. Genomic and Metabolomic Investigation of a Rhizosphere Isolate Streptomyces netropsis WLXQSS-4 Associated with a Traditional Chinese Medicine. Molecules. 2021; 26(8):2147. https://doi.org/10.3390/molecules26082147
Chicago/Turabian StyleZhang, Songya, Lingxiao Zhang, Jing Zhu, Hong Chen, Zhicong Chen, Tong Si, and Tao Liu. 2021. "Genomic and Metabolomic Investigation of a Rhizosphere Isolate Streptomyces netropsis WLXQSS-4 Associated with a Traditional Chinese Medicine" Molecules 26, no. 8: 2147. https://doi.org/10.3390/molecules26082147
APA StyleZhang, S., Zhang, L., Zhu, J., Chen, H., Chen, Z., Si, T., & Liu, T. (2021). Genomic and Metabolomic Investigation of a Rhizosphere Isolate Streptomyces netropsis WLXQSS-4 Associated with a Traditional Chinese Medicine. Molecules, 26(8), 2147. https://doi.org/10.3390/molecules26082147